

Abstract
Identifying genes linked to extreme phenotypes in humans has the potential to highlight biological processes not shared with all other mammals. Here, we report the identification of homozygous loss-of-function variants in the primate-specific gene ZNF808 as a cause of pancreatic agenesis. ZNF808 is a member of the KRAB zinc finger protein family, a large and rapidly evolving group of epigenetic silencers which target transposable elements. We show that loss of ZNF808 in vitro results in aberrant activation of regulatory potential contained in the primate-specific transposable elements it represses during early pancreas development. This leads to inappropriate specification of cell fate with induction of genes associated with liver identity. Our results highlight the essential role of ZNF808 in pancreatic development in humans and the contribution of primate-specific regions of the human genome to congenital developmental disease.
Similar content being viewed by others
Main
Studying the genetic basis of congenital diseases in humans can reveal unique mechanisms orchestrating human organ development that are distinct from those in other species. The development of the human pancreas has unique characteristics compared to its counterpart in rodents, underscoring the importance of investigating this process in humans to elucidate species-specific regulation mechanisms. Previous human genetic studies of individuals with pancreatic agenesis, a rare congenital condition resulting from inappropriate pancreas development, identified evolutionarily conserved genes (GATA6 (ref. 1), GATA4 (ref. 2) and CNOT1 (ref. 3)) involved in the early stages of pancreatic development which have different dosage-dependent effects in mouse and human. Here, we report the identification of loss-of-function variants in the primate-specific gene ZNF808 as a genetic cause of defective pancreatic development in humans.
To identify genetic causes of defective pancreas development, we initially studied two unrelated individuals with isolated pancreatic agenesis (defined by neonatal diabetes, which is diagnosed before 6 months of age and exocrine pancreatic insufficiency1) in whom all known genetic causes had been excluded. We performed exome sequencing in the two affected individuals and their consanguineous parents and found that both individuals were homozygous for ZNF808 loss-of-function variants (Supplementary Tables 1 and 2).
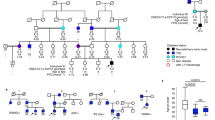
We next investigated the presence of rare ZNF808 biallelic variants in 233 more patients with neonatal diabetes without a pathogenic variant in the known etiological genes4. Homozygous loss-of-function ZNF808 variants were identified in a further 11 unrelated individuals and 2 affected siblings (Fig. 1a,b and Supplementary Table 3). All frameshift and stop-gain variants affect residues in the last exon of the gene and are predicted to result in a truncated protein lacking between 4 and all 23 of the zinc finger domains. Two patients (probands 2 and 3) were homozygous for deletions predicted to result in no ZNF808 messenger RNA. All families showed cosegregation of the ZNF808 homozygous variants with the disease consistent with recessive inheritance (Fig. 1b). No deleterious homozygous loss-of-function ZNF808 variants were identified in >680,000 individuals without neonatal diabetes or pancreatic agenesis (UK BioBank (n = 454,756), Gnomad v.2.1.1 (n = 141,071), 100,000 genomes project from Genomics England (n = 75,118) and Genes and Health (n = 8,921)). Overall, our findings show that ZNF808 biallelic loss-of-function variants cause pancreatic agenesis/neonatal diabetes.

a, Schematic representation of the ZNF808 gene and the pathogenic variants identified in 13 families. Deletions are highlighted in red above the gene cartoon while the loss-of-function variants are represented below. The KRAB and zinc finger domains are annotated at the gene level. b, Partial pedigrees of the 13 families with homozygous ZNF808 variants. NA, sample not available for testing; N/N, variant not detected; N/M, heterozygote for variant; M/M, homozygote for variant. Two black lines between parents indicate individuals who were known to be related. A black line and a dashed line between parents indicate individuals who were not known to be related at testing but were confirmed to be consanguineous by homozygosity mapping of next-generation sequencing data calculated with SavvyHomozygosity34.
The patients’ phenotype supports a role for ZNF808 in early pancreatic development affecting both endocrine and exocrine pancreatic functions (pancreatic agenesis). All individuals showed markedly reduced insulin secretion in utero, as they all had low birth weight (median −2.98 s.d., interquartile range (IQR) −3.56 to −2.47) reflecting reduced insulin-mediated fetal growth and also postdelivery, as they rapidly developed insulin-requiring diabetes (median age at diagnosis 17 weeks, IQR 3–23). There was evidence of early exocrine failure, with 5 of 5 patients tested having undetectable fecal elastase and 4 further patients having symptoms or investigations consistent with fat malabsorption (Supplementary Table 3). The patients’ phenotype is restricted to pancreas as no extra-pancreatic features were consistently observed in the cohort.
Analysis of transcriptomic data from GTEx reveals that ZNF808 is broadly expressed across human adult tissues (Extended Data Fig. 1a). Transcriptomic analysis of 15 human embryonic tissues5 showed that ZNF808 is maximally expressed in developing pancreas (Extended Data Fig. 1b).
ZNF808 is a member of the KRAB zinc finger protein (KZFPs) family, the largest group of DNA binding factors in the human genome (∼350 protein-coding genes). KZFPs primarily act as epigenetic silencers of transposable elements through establishment of heterochromatin-associated H3K9me3 (refs. 6,7). The KZFP family is rapidly evolving with new members found at most phylogenetic branches since its emergence at the dawn of tetrapods8,9.
ZNF808 is exclusively found in primates and its evolutionary origin can be functionally traced through its zinc finger signature to a common ancestor of Old World monkeys (Fig. 2a), with no similar array of zinc fingers found in New World monkeys or any other mammals8. Our genome-wide analysis of sequence identity between primate and non-primate mammalian orthologues for all human protein-coding genes did not highlight any other primate-specific gene confirmed to be causative for a human developmental disorder (Extended Data Fig. 1c and Supplementary Table 4). These findings confirm how extremely rare it is for a primate-specific gene to cause a human congenital developmental disorder.

a, Reconstructed phylogeny of ZNF808 using a zinc finger signature approach. The three amino acids of each zinc finger directly contacting DNA were used to build a specific functional signature to track evolution of ZNF808, as previously described8. Zinc finger domains are color-coded according to the number of variants in each triplet compared with the human version. Notable events of loss or gain of zinc fingers are also represented. No appreciable homology with any zinc finger array was detected in New World monkeys or in any other mammals. Silhouettes of representative species are all from PhyloPic.org. b, ZNF808 binds primarily MER11 transposable elements. Analysis of ZNF808 ChIP–seq data8 reveals that it primarily intersects with transposable elements, although a few binding sites are found on gene promoters and other genomic regions. Further analysis of transposons shows that ZNF808 binds primarily elements of the MER11 family—MER11A, MER11B and MER11C. c, MER11 transposable elements are primate-specific. The origin of each individual MER11 element in the human genome was traced using a comparative multiple alignment of 241 species35. The age of each element was determined as corresponding to the farthest phylogenetic branch where we could find a similar copy at a syntenic locus. Data points are plotted as the sum of elements found to have originated at each phylogenetic branch per subfamily—the x axis is scaled according to time in million years from estimates between the human genome and each phylogenetic branch common ancestor. A curve is interpolated between the data points to show the estimated rate of replication between phylogenetic branches. The scale is relative to each subfamily and is indicative of proportional changes between phylogenetic categories—the highest point for each subfamily is annotated with the number of new elements for scale.
Genome-wide binding profiling of ZNF808 shows that it primarily targets the long terminal repeat of endogenous retroviruses classified as MER11 elements which comprise subfamilies A, B and C (Fig. 2b). These elements also originated in Old World monkeys and spread during evolution in successive waves until recently (Fig. 2c). It was suggested that remnants of MER11 elements might be domesticated10,11,12,13, providing regulatory potential that can modulate gene expression in certain cellular contexts even though they have lost their ability to transpose. Using the chromatin immunoprecipitation ChIP-Atlas14 database, we found that MER11 elements show enriched occupancy for many DNA binding factors, including some involved in early embryonic development (Extended Data Fig. 2a,b). Three of these (GATA4, GATA6 and HNF4A) are known causes of pancreatic agenesis and/or diabetes in humans when mutated1,2,15,16 and are involved in endoderm, liver and pancreas specification. We also found that subsets of MER11 elements are targeted by other primate-specific KZFPs (Extended Data Fig. 2b). This suggests that MER11 elements have the potential to be regulatory elements and are silenced by a group of primate-specific KZFPs, with ZNF808 being the central effector.
To characterize the molecular events triggered by ZNF808 loss, we functionally inactivated ZNF808 in H1 human embryonic stem cells (hESC) using CRISPR-Cpf1 (hereafter, ZNF808 KO) (Fig. 3a and Extended Data Fig. 3). Also, we generated induced pluripotent stem cells (iPSCs) derived from proband 2 harboring a homozygous deletion of ZNF808 exons 4 and 5 to validate our findings.

a, ZNF808 gene editing using CRISPR. Cpf1 and two guide RNAs targeting the zinc finger array were used in H1 stem cells to produce the ZNF808 KO. b, In vitro differentiation protocol to pancreatic progenitors. Overview of the multistep differentiation protocol used in this study for both epigenetic and transcriptomic analysis of differences induced by ZNF808 KO. Stage numbers from S0 to S4 are used through the text to refer to specific steps. c, ZNF808 is important for the maintenance of epigenetic repression on MER11 elements. Left, H3K9me3 ChIP–seq peaks intersecting with MER11 elements reveal that most sites are covered in heterochromatin-associated H3K9me3 and are bound by ZNF808. A proportion of these peaks lose H3K9me3 in ZNF808 KO clones at various stages of differentiation (top). Results show that many MER11 elements that had H3K9me3 signal in the WT gain H3K27ac in the ZNF808 KO, especially at the early stages of differentiation (bottom). d, A subset of MER11 elements is activated in the ZNF808 KO heatmap showing clustering of 220 MER11 elements displaying a loss of H3K9me3 followed by a gain of H3K27ac in at least one stage of differentiation. Normalized ChIP–seq signal is shown—color scale ranges from +3 (red) to −3 (blue) on a z-score scale.
We assayed epigenetic changes at key stages of in vitro differentiation from pluripotent stem cells to pancreatic progenitor cells (Fig. 3b)17. We first quantified the genome-wide presence of H3K9me3 (Fig. 3c, top), a hallmark of heterochromatin induced by KZFPs18 and focused our analysis on MER11 elements as they represented 91.3% of ZNF808-bound sites with H3K9me3 (Extended Data Fig. 4a–c). H3K9me3 was detected on 1,367 MER11 elements in wild-type (WT) stem cells, 1,041 of which are known to be targets of ZNF808. The ZNF808 KO cells show a loss of 693 H3K9me3-positive MER11 loci at S0 (embryonic stem cells), with 684 of these being known targets of ZNF808. The loss of H3K9me3 on MER11 elements known to be bound by ZNF808 is statistically significant when considering all stages (Fisher exact test P < 3.6 × 10−61). This implies that ZNF808 plays a key role in silencing MER11 elements in early development.
We then surveyed whether the loss of silencing revealed active regulatory potential at MER11 elements. In the ZNF808 KO, we observed emergence of 226 H3K27ac-positive MER11 elements at S0 (Fig. 3c, bottom), most (198) at known targets of ZNF808. The gain of H3K27ac on known MER11 elements targeted by ZNF808 is statistically significant when all stages are considered (Fisher exact test P < 1.8 × 10−16).
We identified a subset of 220 MER11 elements where both loss of H3K9me3 and gain of H3K27ac were observed at the same stage in the ZNF808 KO. These have distinct patterns of activity during differentiation which we could group in six clusters (Fig. 3d and Extended Data Fig. 5), reflective of when the elements gain H3K27ac. We found that MER11 elements gaining activity in the ZNF808 KO later during differentiation (cluster nos. 5 and 6) are more frequently bound by GATA4 or members of the HNF4 family (Extended Data Fig. 5c). These transcription factors are dynamically expressed during our differentiation protocol (Extended Data Fig. 6) and are known to be important in specification of endodermal derived tissues19,20 including pancreas21,22 and liver20,23,24. Analysis of chromatin accessibility from a single-cell ATAC-seq tissue atlas25 and the NIH Roadmap26 dataset shows that MER11 elements are not active in pancreas, however multiple MER11 loci are normally active in specific cell types, including fetal trophoblasts, enterocytes and hepatocytes (Extended Data Fig. 7a,d,e), with enrichment for those unmasked in the ZNF808 KO cells (Extended Data Fig. 7b,c). We found strong agreement between our ZNF808 KO and the patient-derived iPSCs in terms of loss of H3K9me3 and gain of H3K27ac (Extended Data Fig. 8a).
These results show that ZNF808 silences specific MER11 elements during differentiation toward pancreatic lineages and that loss of ZNF808 unmasks their regulatory potential which is normally found active in other cellular contexts.
We next quantified transcriptomic changes induced in the ZNF808 KO during differentiation. We found that the number of genes whose expression was dysregulated increased from 443 genes at S0 (stem cell state) to a peak of 2,124 genes at S3 (posterior foregut) (Fig. 4a). Comparable changes in gene expression were detected in the patient-derived iPSCs by both RNA-seq and quantitative PCR (qPCR) with reverse transcription (Extended Data Fig. 8b,c). We found that unmasked MER11 elements drive proximal gene activation in the ZNF808 KO predominantly at the first two stages of differentiation (S0, stem cell state; S1, definitive endoderm), with MER11 elements over-represented near activated genes at enhancer (10 kb–1 Mb) but not promoter distances (Fig. 4b and Extended Data Fig. 9a–c). We also found that the smaller group of MER11 elements that gain H3K27ac later in differentiation drive gene activation at S4 (pancreatic progenitor stage) (cluster no. 6, Fig. 3c and Extended Data Fig. 9c). These results show that the unmasking of MER11 elements in the ZNF808 KO impacts nearby gene expression in early differentiation with downstream effects as differentiation progresses.

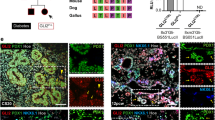
a, Loss of ZNF808 leads to perturbed gene expression throughput pancreatic differentiation. Bar chart showing total genes activated and repressed with FDR < 0.05 and |FC| > 1.25 for each stage of pancreatic differentiation. K, thousands. b, Dysregulated genes are found in proximity to unmasked MER11 elements in the ZNF808 KO early in differentiation. Proximity enrichment (−log10 Fisher exact right-tail P value) showing an excess of dysregulated genes compared to all genes in proximity to MER11 elements losing H3K9me3 and gaining H3K27ac in the ZNF808 KO as a function of distance between genes and binding sites for genes activated (orange) and repressed (blue) on log10 scale. Enrichment peaks between 10 kb and 100 kb suggest ZNF808 repressing distal gene enhancers. A total of 43.4% and 23.9% of activated genes at S0 and S1, respectively, are within 1 MB of a MER11 element. c, Hepatic cords genes are activated and dorsal pancreas bud genes are repressed in ZNF808 KO. Top, Fisher exact enrichment between ZNF808 KO activated and repressed genes and genes more highly expressed in CS12–14 hepatic cords or dorsal pancreatic buds, respectively28. Bottom, log2 fold-change ZNF808 KO over WT for dorsal pancreas bud (left, blue) and hepatic cords (right, orange) genes. Each dot represents a single gene. d, Genes exclusively expressed in liver and activated in hepatic cords are activated in ZNF808 KO. The log2 fold-changes of the 29 genes that are expressed in CS12–14 hepatic cords, exclusively expressed in GTEx liver and activated at S2 in ZNF808 KO. Error bars give DESeq2 standard error of the log2 fold-change, n = 3 independent biological replicates. e, Immunostaining of AFP and PDX1 at S3 (posterior foregut) stage. Confirmation of RNA-seq results by immunostaining, showing activation of AFP in ZNF808 KO in PDX1 positive cells (representative of three independent differentiation experiments; scale bar, 100 μm). Independent replicate stainings from the same cell lines are given in Extended Data Fig. 3d.
We performed gene set enrichment analysis using Enrichr27 and observed fetal and adult liver genes activated in ZNF808 KO from S2 (primitive gut tube) onwards (fetal liver enriched at stage S2, false discovery rate (FDR) < 10−14, odds ratio (OR) 12.3) (Extended Data Fig. 9d). We tested whether this reflected the divergence between pancreatic buds and hepatic cords in vivo, with comparison to Carnegie stage 12–14 human embryos that correspond to our S2–S4 in vitro stages28 (Fig. 4c, top). We found hepatic cord genes activated in our ZNF808 KO (FDR < 10−45, OR 6.4, stage S2) and conversely dorsal pancreas genes repressed (FDR < 10−12, OR 3.1, stage S3). The divergence between hepatic cord and dorsal pancreas genes peaks at posterior foregut stage S3 (Fig. 4c, bottom), coinciding with the introduction of factors modulating propancreatic signaling pathways (for example, retinoic acid and BMP and SHH inhibition); we suggest that this extinguishes the hepatic fate in the ZNF808 KO.
Using GTEx we found that the induction of hepatic gene expression in the ZNF808 KO was enriched in genes whose expression is exclusive to liver (Extended Data Fig. 10; P < 10−16, OR 7.6, stage S2). We identified a set of 29 liver-exclusive, hepatic cord genes activated at S2, including genes such as AFP, ALB, APOA1 and LIPC (Fig. 4d). This included an example of a gene (TDO2) activated in proximity to a MER11 element, suggesting that MER11 unmasking is compatible with hepatic gene expression (Extended Data Fig. 10c–e). We confirmed the transcriptomic upregulation of the early liver marker alpha-fetoprotein (AFP) by immunostaining (Fig. 4e and Extended Data Fig. 3d). Taken together, our data show that ZNF808 is essential to prevent a liver gene expression program from being aberrantly activated during pancreas differentiation, suggesting a potential mechanism for pancreatic agenesis.
We report the identification of recessive loss-of-function variants in ZNF808, a primate-specific gene, as a cause of pancreatic agenesis, a congenital developmental disorder. This confirms the crucial importance of gene discovery efforts in patients with extreme phenotypes.
Our study shows that ZNF808 is critical for human pancreatic development. Previous human genetic studies identified pancreatic developmental genes, including GATA6 (refs. 1,29,30), GATA4 (refs. 2,15,29,30,31) and HNF4A (refs. 16,32,33), with different dosage-dependent effects between mouse and humans, suggesting key differences in pancreatic development between the two species. Our identification of the role of ZNF808 and MER11 element regulation during human pancreatic development offers important insights into how mechanisms regulating pancreas development have diverged between primates and other mammals and supports a key role of ZNF808 in regulating differentiation of endoderm progenitors between liver and pancreas lineages. We provide correlative evidence that the unmasking of MER11 elements in the ZNF808 KO ultimately leads to the downstream induction of a liver gene expression program during pancreas differentiation. Future work combining targeted genome editing and 3D conformation data will be necessary to confirm which of the regulatory regions repressed by ZNF808 are responsible for disrupting pancreas development in our patients.
Our characterization of the role of ZNF808 during early development offers insights into the role of KZFPs, showing that they provide a negative layer of regulation that masks transposable element regulatory potential in cellular settings where the right transcription factors are present, yet the domesticated regulatory activity is undesired. We believe that this is an evolutionary mechanism that allows for a broader range of transposons to be domesticated as they do not have to prove beneficial in all cellular contexts if KZFPs can selectively silence them where they would have a negative impact.
The crucial role of ZNF808 and MER11 elements in human pancreas development underscores that even primate-specific genes and transposable elements can be involved in important aspects of human biology. This discovery offers important insights into the evolution of gene regulation during human development and opens new avenues of research in the fields of human genetics and diabetes.
Methods
Subjects
The study was conducted in accordance with the Declaration of Helsinki and all subjects or their parents/guardian gave informed written consent for genetic testing. DNA testing and storage in the Beta Cell Research Bank was approved by the Wales Research Ethics Committee 5 Bangor (REC 17/WA/0327, IRAS project ID 231760). For Proband 2, parents gave written consent for collection of a skin biopsy to be dedifferentiated to iPSCs to study the patient’s cause of neonatal diabetes. No participant received compensation for entering this study.
Individuals with neonatal diabetes diagnosed before the age of 6 months were recruited by their clinicians for molecular genetic analysis in the Exeter Genomics Laboratory. For one patient (case 10 in Supplementary Table 3), a ZNF808 homozygous loss-of-function variant was identified by exome sequencing analysis by the Center for Genomic Medicine, King Faisal Specialist Hospital and Research Center, Riyadh.
Proband 1 and 2 were selected from a larger cohort of individuals with pancreatic agenesis3. Within this group, they were the only two individuals without a genetic diagnosis who were born to consanguineous parents.
Genetic analysis
Exonic sequences were enriched from genomic DNA using Agilent’s SureSelect Human All Exon kit (v.4) and then sequenced on an Illumina HiSeq 2000 sequencer using 100 base pair (bp) paired-end reads. The sequencing data were analyzed using an approach based on the GATK best-practice guidelines. GATK v.3.7 HaplotypeCaller was used to identify variants that were annotated using Alamut batch v.1.8 (human reference genome assembly hg19) and variants that failed the QD2 VCF filter or had less than five reads supporting the variant allele were excluded. Copy number variants were called by SavvyCNV36, which uses read depth to judge copy number states. SavvyVcfHomozygosity34 was used to identify large (>3 Mb) homozygous regions in the exome sequencing data (https://github.com/rdemolgen/SavvySuite).
A total of 232 patients diagnosed with diabetes before age 6 months in whom the known genetic causes of neonatal diabetes had been excluded were analyzed either by using a targeted next-generation sequencing assay, which includes baits for known neonatal diabetes genes and additional candidate genes followed up from gene discovery, such as ZNF808, or by independent genome sequencing analysis. Variant confirmation and cosegregation in family members were performed by Sanger sequencing (primers available on request).
Cell culture and in vitro differentiation of human stem cells
The hESC (WA01/H1 line, Wicell) and patient-derived iPSC were cultured on Matrigel-coated plates (BD Biosciences) in Essential 8 (E8) medium (Life technologies, A1517001) and passaged using EDTA. To carry out the differentiation experiments, hPSC (human pluripotent stem cells) were dissociated using EDTA, then seeded on new Matrigel-coated plates in E8 medium supplemented with 10 µM rho-associated kinase inhibitor (ROCKi, catalog no. Y-27632; Selleckchem catalog no. S1049) at a density of 0.21 million cells per cm2. After 24 h, the differentiation was started by washing the cells with PBS, then changing the medium to D0 medium. The differentiation was carried out using our optimized protocol as previously described17.
Genome editing
To create an in vitro model for studying the role of ZNF808 in pancreatic development, guide RNAs targeting the zinc fingers domain of the fifth exon of ZNF808 for deletion were designed using Benchling (https://benchling.com) (gRNAs sequence available in Supplementary Table 7). The gRNAs with the highest quality score and lowest off-targets score were selected and purchased, alongside the RNP components (Alt-R Cas12a (Cpf1) Ultra protein, crRNA), from Integrated DNA Technologies (IDT) and used according to the manufacturer’s recommended protocol. A total of 2 million cells were electroporated with the RNP complex using Neon Transfection system (Thermo Fisher, 1100 V, 20 ms, two pulses) and plated on Matrigel-coated plates in E8 medium containing 10 µM ROCK inhibitor overnight. Afterwards, cells were single-cell sorted, expanded and screened for the desired deletion using PCR. Positive clones were validated by Sanger sequencing at Eurofins Genomics and the sequences were aligned using Geneious Prime 2020.1.1. The KO clones were characterized for pluripotency, chromosomal integrity and the top three off-target hits predicted by the online tool CRISPOR37 were checked with no off-target indels found.
Formaldehyde crosslinking
To fix the cells for ChIP–seq samples preparation, cells were incubated with TrypLE for 5–10 min at 37 °C and gently homogenized with the pipette, then pooled in a 15 ml Falcon tube containing warm DMEM. Afterwards, cells were spun down at 250g at room temperature for 3 min, then resuspended in DMEM at a concentration of 5 million cells per ml and incubated with 333 mM fresh 16% methanol-free formaldehyde at room temperature for precisely 10 min. Formaldehyde was quenched using 250 mM Tris pH 8.0 for another 10 min at room temperature, then cells were spun down at 250g at 4 °C for 5 min. Cell pellet was resuspended gently in PBS, aliquoted into 1.5 ml Eppendorf tubes and spun down at 250g at 4 °C for 5 min. Supernatant was removed and samples were stored at −80 °C until further processing.
Flow cytometry analysis
The hESC-derived cells were dissociated into single cells by incubation with TrypLE for 5–10 min at 37 °C and resuspended in cold FBS/PBS (5% v/v). For surface marker staining of CXCR4, 1 million cells were incubated with the directly conjugated antibody CD184/CXCR4 APC at a final dilution of 1:10 for 30 min at room temperature. For intracellular markers, 1 million cells were first fixed in 350 µl of Cytofix/Cytoprem Buffer (BD, no. 554722) for 20 min at 4 °C, then washed twice with BD Perm/Wash Buffer Solution (BD, no. 554723). Cell pellet was resuspended in 80 µl of FBS/BD Prem/Wash buffer (4% v/v) and incubated with the corresponding directly conjugated antibody at a final dilution of 1:80 overnight at 4 °C. After incubation with the antibody, cells were washed twice and analyzed using FACSCalibur cytometer (BD Bioscience), BD CellQuest Pro software v.4.0.2 and FlowJo software v.9 (Tree Star). Details of the antibodies are listed in Supplementary Table 6.
Immunocytochemistry
For adherent cultures, cells were fixed in 4% PFA for 15 min at room temperature, permeabilized with 0.5% Triton X100 in PBS, then blocked with UltraV block (Thermo Fisher) for 10 min and incubated with primary antibodies diluted in 0.1% Tween in PBS overnight at 4 °C. After incubation, cells were washed twice with PBS and incubated with corresponding secondary antibodies diluted in 0.1% Tween in PBS for 1 h at room temperature. Details of the antibodies are listed in Supplementary Table 6.
Chromatin immunoprecipitation
The following steps were performed with ice-cold samples and buffers containing a protease inhibitor cocktail (cOmplete ULTRA Tablets EDTA-free, Roche). In 1.5 ml DNA LoBind tubes, 4 million fixed cells were resuspended in 1 ml of lysis buffer 1 (50 mM HEPES-KOH pH 7.4, 140 mM NaCl, 1 mM EDTA, 0.5 mM EGTA, 10% glycerol, 0.5% NP40, 0.25% Triton X100, proteinase inhibitor 1×) incubated at 4 °C on a rotating wheel at 10 r.p.m. for 10 min. Cells were then centrifuged at 1,700g for 5 min at 4 °C. Supernatant was discarded and pellets were resuspended in 1 ml of lysis buffer 2 (10 mM Tris HCl pH 8.0, 200 mM NaCl, 1 mM EDTA, 0.5 mM EGTA, proteinase inhibitor 1×) and incubated at 4 °C on a rotating wheel at 10 r.p.m. for 10 min. After centrifugation at 1,700g for 5 min at 4 °C, supernatant was discarded and pellets were washed with 500 µl of SDS shearing buffer (1 mM Tris HCl pH 8.0, 1 mM EDTA, 0.15% SDS, proteinase inhibitor 1×), without disturbing the pellets, followed by centrifugation at 1,700g for 5 min. Washing was repeated twice and pellets were resuspended in 1 ml of SDS shearing buffer and transferred into Covaris milliTUBE 1 ml AFA Fiber. Chromatin was sheared on a Covaris E220 for 6 min at 5% duty cycle, 140 W, 200 cycles. The sheared chromatin was then transferred into 1.5 ml DNA LoBind tubes and centrifuged at 10,000g for 5 min at 4 °C. Supernatant was then used immediately for immunoprecipitation. Chromatin quality control was performed on Bioanalyzer 2100 (Agilent) to verify that most fragments ranged between 200 and 600 bp.
For H3K9me3 IP, chromatin corresponding to 1 million cells was put in a new 1.5 ml DNA LoBind tube and topped to 900 µl with SDS shearing buffer. For H3K27ac IP, chromatin corresponding to 3 million cells was put in a new 1.5 ml DNA LoBind tube and topped to 900 µl total with SDS dilution buffer (1 mM Tris HCl pH 8.0, 1 mM EDTA, 0.026% SDS, proteinase inhibitor 1×) so that the SDS in the final IP buffer would be 0.1%. IP conditions were further adjusted to 150 mM NaCl and 1% Triton final, 1 ml final volume. Either 1 µl of H3K9me3 antibody (catalog no. 39685, Active Motif) or 5 µg of H3K27ac antibody (catalog no. 39161, Active Motif) was added. The IP was then incubated on rotating wheel at 10 r.p.m. at 4 °C overnight. The next day, Dynabeads Protein G (5 µl for the H3K9me3 IP or 25 µl for the H3K27ac) were put on magnet and supernatant was removed. Beads were then resuspended in the full volume of the IP and incubated for 2 h on a rotating wheel at 10 r.p.m. at 4 °C.
For H3K9me3, low-salt washing buffer (10 mM Tris HCl pH 8.0, 150 mM NaCl, 1 mM EDTA, 1% Triton X100, 0.15% SDS, 1 mM PMSF) and high-salt washing buffer (10 mM Tris HCl pH 8.0, 500 mM NaCl, 1 mM EDTA, 1% Triton X100, 0.15% SDS, 1 mM PMSF) were used.
For H3K27ac, low-salt washing buffer (20 mM Tris HCl pH 8, 150 mM NaCl, 2 mM EDTA, 1% Triton X100, 0.1% SDS, 1 mM PMSF) and high-salt washing buffer (20 mM Tris HCl pH 8, 500 mM NaCl, 2 mM EDTA, 1% Triton X100, 0.1% SDS, 1 mM PMSF) were used.
All washes took place while IPs and buffers were ice cold. PMSF was always added in the buffers immediately before each wash. The IPs were placed on a magnetic rack and supernatant was discarded. Beads were resuspended in low-salt washing buffer and transferred into a clean DNA LoBind tube. Beads were then placed on a magnetic rack; supernatant was removed and beads were resuspended in low-salt washing buffer. The mixture was placed again on a magnetic rack, supernatant was discarded and beads were washed with high-salt washing buffer. Once more, samples were placed on the magnetic rack, supernatant was removed and beads were resuspended in LiCl buffer (10 mM Tris HCl pH 8.0, 1 mM EDTA, 0.5 mM EGTA, 250 mM LiCl, 1%NP40, 1%NaDOC, 1 mM PMSF). The mixture was placed on the magnetic rack, supernatant was removed and beads were washed with 10 mM Tris HCl pH 8.0 and transferred to a clean DNA LoBind tube. Finally, with the samples on the magnetic rack the supernatant was completely removed and beads were resuspended in elution buffer (10 mM Tris HCl pH 8.0, 1 mM EDTA, 1% SDS and 150 mM NaCl).
RNase A was added to the elution buffer at a final concentration of 0.5 µg µl−1 and samples were incubated at 37 °C for 1 h in a shaking incubator at 1,100 r.p.m. Subsequently proteinase K was added at a concentration of 400 ng µl−1 and chromatin was decrosslinked at 65 °C overnight. The supernatant was collected and purified using Serapure beads before library preparation.
To control for the efficiency of the IP, we used qPCR with primers targeting negative and positive regions in the genome for the histone marks H3K9me3 and H3K27ac, respectively (primer sequences available in Supplementary Table 7).
Library preparation
Libraries were prepared using NEBNext Ultra II DNA Library Prep Kit for Illumina following the manufacturer’s instructions. Adapters and indexed primers design, resuspension and annealing were as previously described38. The adapters used were iTrusR2-stubRCp andiTrusR1-stub.
The library was quantified by qPCR using KAPA SYBR FAST and the set of primers itru7_101_01 and itru5_01_A. After quantification the library was amplified and double indexed (primers details in Supplementary Table 7). Amplified libraries were double size selected using home-made Serapure beads to enrich for fragments between 200 and 600 bp.
After amplification, confirmation that the IP remained efficient was carried out using qPCR with primers targeting genomic regions negative and positive regions for the histone marks H3K9me3 and H3K27ac, respectively, as mentioned above. Libraries were sent for 150 bp paired-end sequencing at Novogene.
ChIP–seq analysis
Reads from each library were mapped on hg19 using Bowtie2 v.2.4.4 (ref. 39) with the ‘very-sensitive-local’ setting. Mapped reads were compressed in BAM files and indexed using SAMtools v.1.18 (ref. 40). We used MACS2 v.2.2.7.1 (ref. 41) to call peaks using the corresponding input datasets as background controls. Peaks were excluded if the average read had MAPQ < 20. Reads intersecting intervals were counted with HTSeq 0.13.5, with a filter of MAPQ > 20 for properly paired reads applied. To identify MER11 elements losing H3K9me3 and gaining H3K27ac, we took peak regions defined by an overlap with H3K9me3 (pybedtools 0.8.1 and bedtools v.2.30.0, f = 0.5, F = 0.5; e, True; u, True; wa, True) in WT cells with no overlap in the KO and the opposite for H3K27ac to derive a final list of 220 MER11 elements activated in ZNF808 KO. To identify epigenetic responses over differentiation, we quantified depth normalized reads in each region and standardized H3K9me3 and H3K27ac signals separately by applying a z-score normalization (zero-meaned and standard deviation scaled) to each region over the differentiation stages. To assign regions to clusters of similar behavior, we performed k-means clustering with parameters n_clusters = 6, init = ‘random’, max_iter = 3,000, n_init = 100 using Scipy. To visualize these, we also performed hierarchical clustering using fastcluster v.1.2.4 with optimal leaf ordering which we mapped onto our k-means clustering to provide a heatmap with consistent within cluster and between cluster ordering. ChIP–seq tables and data analyzed in Python used pandas 1.3.5 and NumPy 1.12.5. Enrichment of transcription factors was by downloading hg19 ChIP peak data from the repository ChIP-Atlas (https://chip-atlas.org14). We used bedtools fisher v.2.30.0 with options –f 0.5 -F 0.5 -e to calculate intersection contingency table between MER11 elements of distinct subfamilies (MER11A, MER11B and MER11B) and each ChIP-Atlas dataset. We then calculated right-tail Fisher exact P values for over-representation and ORs using the Julia Distributions.jl package and applied Benjamini–Hochberg FDR control using MultipleTesting.jl.
Analysis of chromatin accessibility at MER11 elements was performed using the chromatin atlas25. Peak calls of chromatin accessibility for each cell type deconvoluted from single-cell signal clustering were downloaded and overlapped with bedtools with f = 0.5, F = 0.5; e, True (at least 50% overlap from either peak).
Analysis of the epigenetic status of MER11 in various cell types was done using the expanded NIH Roadmap analysis by ref. 26. Only datasets with measured H3K9me3 and at least one of H3K27ac or H3K4me1 were retained. Chromatin states were assigned by overlapping with precalculated HMM states from ref. 26 and were collapsed by cell type using the provided metadata—overlap in at least one cell type with a particular chromatin state was counted as present and Enhancer/TSS status were given precedence over heterochromatin if both were found in a cell type at a particular locus in two different biosamples.
Gene expression analysis by RNA-seq
Stranded, poly-A selected RNA-seq libraries were prepared and sequenced from three independent replicates of our differentiation time course (paired-end 150 bp reads) by Novogene.
Stranded paired-end 150 bp RNA-seq reads were aligned to the hg19 genome using STAR v.2.7.3a (ref. 42) and quantified against Gencode v.36 release liftover to hg19 by RSEM v.1.3.2 (ref. 43) using the RSEM-STAR pipeline, with further options --seed 1618 --calc-pme --calc-ci --estimate-rspd --paired-end. RSEM estimated read counts per sample were rounded for use with DESeq2 v.1.30.1 (ref. 44). We perform differential expression analysis for each stage WT versus ZNF808 KO for all genes with at least ten raw counts in all replicates of one condition. Gene expression varies substantially over the differentiation time course with subsets of genes only expressed at early or late stages, therefore when testing each stage, we supply DESeq2 with samples from adjacent stages for information sharing in estimating dispersions. We have three independent replicates of our differentiation time course and we perform a paired analysis between WT and KO pairs within DESeq2 with the model ∼ExpNum + Genotype, where ExpNum is a factor indicating the replicate and GenoType is a factor indicating WT or ZNF808 KO, we then perform a contrast between the two levels of the GenoType factor and calculate differential expression with independentFiltering=FALSE. We consider genes with FC (fold change) > 1.25 and FDR < 0.05 differentially expressed.
To determine enrichments of differentially expressed genes in proximity to clusters of epigenetic response in ZNF808 loss, we used the package ProximityEnrichment.jl (https://github.com/owensnick/ProximityEnrichment.jl). Specifically, we calculate hypergeometric right-tail P values for the association of differentially expressed genes within x bp of ZNF808-bound region cluster against a background of genes for x in [1, 1 × 106]. For proximity-enrichment heatmaps, we take the maximal enrichment from this interval.
To determine gene set enrichments for sets of differentially expressed genes, we use the Enrichr API27, with the package (https://github.com/owensnick/Enrichr.jl) to recover enrichments for BioPlanet 2019, GO_Biological_Process 2018, GO_Cellular_Component 2018, GO_Molecular_Function 2018, Human_Gene_Atlas and KEGG_2019_Human gene sets. As Enrichr calculates enrichments using a generic background, we downloaded all definitions of Terms and Gene sets and recalculated Fisher exact right-tail P values and ORs (using HypothesisTesting.jl) for terms over-represented in the dysregulated genes against a background of all genes tested for differential expression, we then corrected these for multiple testing using the Benjamini–Hochberg method (using MultipleTesting.jl). We report all enrichments in Supplementary Table 5, we selected the most prominent and relevant. To assess the intersection between our data and the laser capture of human embryo hepatic cords and dorsal pancreas28, we took the set of genes detected in each stage of our data and hepatic cords versus dorsal pancreas and calculated the association between direction of dysregulation in our data with the direction of differentially expressed genes in the hepatic cords versus dorsal pancreas comparison with FDR < 0.05.
To identity genes exclusively expressed in the adult liver, we downloaded v8 gene level transcripts per million over all GTEx samples from https://gtexportal.org (file GTEx_Analysis_2017-06-05_v8_RNASeQCv1.1.9_gene_tpm.gct.gz). We calculated the IQR for all genes in all tissues and took a pragmatic definition of liver-exclusive expression: we took those genes for which the lower quartile of liver expression exceeded the upper quartile of expression in all other tissues, yielding 357 genes (Extended Data Fig. 10a). The genotype-tissue expression (GTEx) project was supported by the of the Office of the Director of the National Institutes of Health (NIH) and by NCI, NHGRI, NHLBI, NIDA, NIMH and NINDS. The data used for the analyses described in this manuscript were obtained from the GTEx Portal on 6 January 2023.
Data and code to perform transcriptomic analysis and to generate figure panels are available from https://github.com/owensnick/ZNF808Genomics.jl.
Statistics and reproducibility
No statistical method was used to predetermine sample size for the patient cohort as the aim of the study was to identify the genetic cause of pancreatic agenesis, which is a rare monogenic condition.
Three independent replicates of H1 embryonic stem cell-derived WT and ZNF808 KO differentiation time courses were collected. For transcriptomic analysis we used n = 3 replicates for WT and ZNF808 KO and n = 1 for the patient-derived iPSC. From each of WT, ZNF808 KO and patient-derived iPSC, one replicate per stage was assayed for H3K9me3 and H3K27ac epigenomics.
No data were excluded from any of the experiments described.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Clinical and genotype data are available only through collaboration as this can be used to identify individuals and so cannot be made openly available. Requests for collaboration will be considered following an application to the Genetic Beta Cell Research Bank (https://www.diabetesgenes.org/current-research/genetic-beta-cell-research-bank/). Contact by email should be directed to A. Hattersley (A.T.Hattersley@exeter.ac.uk). All requests for access to data will be responded to within 28 days. Transcriptomic and epigenomic data for WT and ZNF808 KO are available from the NCBI Gene Expression Omnibus under accession GSE205164. Source data are provided with this paper.
Code availability
Code and software versions used to analyze the data presented are indicated in the Methods and provided in https://github.com/owensnick/ZNF808Genomics.jl with persistent Zenodo https://doi.org/10.5281/zenodo.8375708.
References
Allen, H. L. et al. GATA6 haploinsufficiency causes pancreatic agenesis in humans. Nat. Genet. 44, 20–22 (2011).
Shaw-Smith, C. et al. GATA4 mutations are a cause of neonatal and childhood-onset diabetes. Diabetes 63, 2888–2894 (2014).
De Franco, E. et al. A specific CNOT1 mutation results in a novel syndrome of pancreatic agenesis and holoprosencephaly through impaired pancreatic and neurological development. Am. J. Hum. Genet. 104, 985–989 (2019).
Zhang, H., Colclough, K., Gloyn, A. L. & Pollin, T. I. Monogenic diabetes: a gateway to precision medicine in diabetes. J. Clin. Invest. 131, e142244 (2021).
Gerrard, D. T. et al. An integrative transcriptomic atlas of organogenesis in human embryos. eLife 5, e15657 (2016).
Schultz, D. C., Friedman, J. R. & Rauscher, F. J. 3rd Targeting histone deacetylase complexes via KRAB-zinc finger proteins: the PHD and bromodomains of KAP-1 form a cooperative unit that recruits a novel isoform of the Mi-2alpha subunit of NuRD. Genes Dev. 15, 428–443 (2001).
Schultz, D. C., Ayyanathan, K., Negorev, D., Maul, G. G. & Rauscher, F. J. 3rd SETDB1: a novel KAP-1-associated histone H3, lysine 9-specific methyltransferase that contributes to HP1-mediated silencing of euchromatic genes by KRAB zinc-finger proteins. Genes Dev. 16, 919–932 (2002).
Imbeault, M., Helleboid, P. Y. & Trono, D. KRAB zinc-finger proteins contribute to the evolution of gene regulatory networks. Nature 543, 550–554 (2017).
Liu, H., Chang, L. H., Sun, Y., Lu, X. & Stubbs, L. Deep vertebrate roots for mammalian zinc finger transcription factor subfamilies. Genome Biol. Evol. 6, 510–525 (2014).
Bi, S., Gavrilova, O., Gong, D. W., Mason, M. M. & Reitman, M. Identification of a placental enhancer for the human leptin gene. J. Biol. Chem. 272, 30583–30588 (1997).
Pavlicev, M., Hiratsuka, K., Swaggart, K. A., Dunn, C. & Muglia, L. Detecting endogenous retrovirus-driven tissue-specific gene transcription. Genome Biol. Evol. 7, 1082–1097 (2015).
Pontis, J. et al. Primate-specific transposable elements shape transcriptional networks during human development. Nat. Commun. 13, 7178 (2022).
Prudhomme, S., Oriol, G. & Mallet, F. A retroviral promoter and a cellular enhancer define a bipartite element which controls env ERVWE1 placental expression. J. Virol. 78, 12157–12168 (2004).
Oki, S. et al. ChIP-Atlas: a data-mining suite powered by full integration of public ChIP–seq data. EMBO Rep. 19, e46255 (2018).
D’Amato, E. et al. Genetic investigation in an Italian child with an unusual association of atrial septal defect, attributable to a new familial GATA4 gene mutation and neonatal diabetes due to pancreatic agenesis. Diabet. Med. 27, 1195–200 (2010).
Yamagata, K. et al. Mutations in the hepatocyte nuclear factor-4 alpha gene in maturity-onset diabetes of the young (MODY1). Nature 384, 458–460 (1996).
Balboa, D. et al. Functional, metabolic and transcriptional maturation of human pancreatic islets derived from stem cells. Nat. Biotechnol. 40, 1042–1055 (2022).
Matsui, T. et al. Proviral silencing in embryonic stem cells requires the histone methyltransferase ESET. Nature 464, 927–931 (2010).
Sherwood, R. I., Chen, T. Y. & Melton, D. A. Transcriptional dynamics of endodermal organ formation. Dev. Dynam. 238, 29–42 (2009).
Zaret, K. S. From endoderm to liver bud: paradigms of cell type specification and tissue morphogenesis. Curr. Top. Dev. Biol. 117, 647–669 (2016).
Jennings, R. E. et al. Development of the human pancreas from foregut to endocrine commitment. Diabetes 62, 3514–3522 (2013).
Jennings, R. E., Berry, A. A., Strutt, J. P., Gerrard, D. T. & Hanley, N. A. Human pancreas development. Development 142, 3126–3137 (2015).
DeLaForest, A. et al. HNF4A is essential for specification of hepatic progenitors from human pluripotent stem cells. Development 138, 4143–4153 (2011).
Si-Tayeb, K., Lemaigre, F. P. & Duncan, S. A. Organogenesis and development of the liver. Dev. Cell 18, 175–189 (2010).
Zhang, K. et al. A single-cell atlas of chromatin accessibility in the human genome. Cell 184, 5985–6001(2021).
Boix, C. A., James, B. T., Park, Y. P., Meuleman, W. & Kellis, M. Regulatory genomic circuitry of human disease loci by integrative epigenomics. Nature 590, 300–307 (2021).
Kuleshov, M. V. et al. Enrichr: a comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 44, W90–7 (2016).
Jennings, R. E. et al. Laser capture and deep sequencing reveals the transcriptomic programmes regulating the onset of pancreas and liver differentiation in human embryos. Stem Cell Rep. 9, 1387–1394 (2017).
Carrasco, M., Delgado, I., Soria, B., Martin, F. & Rojas, A. GATA4 and GATA6 control mouse pancreas organogenesis. J. Clin. Invest 122, 3504–3515 (2012).
Morrisey, E. E. et al. GATA6 regulates HNF4 and is required for differentiation of visceral endoderm in the mouse embryo. Genes Dev. 12, 3579–3590 (1998).
Watt, A. J., Zhao, R., Li, J. & Duncan, S. A. Development of the mammalian liver and ventral pancreas is dependent on GATA4. BMC Dev. Biol. 7, 37 (2007).
Chen, W. S. et al. Disruption of the HNF-4 gene, expressed in visceral endoderm, leads to cell death in embryonic ectoderm and impaired gastrulation of mouse embryos. Genes Dev. 8, 2466–2477 (1994).
Pearson, E. R. et al. Macrosomia and hyperinsulinaemic hypoglycaemia in patients with heterozygous mutations in the HNF4A gene. PLoS Med. 4, e118 (2007).
Wakeling, M. N. et al. Homozygosity mapping provides supporting evidence of pathogenicity in recessive Mendelian disease. Genet. Med. 21, 982–986 (2019).
ZoonomiaConsortium. A comparative genomics multitool for scientific discovery and conservation. Nature 587, 240–245 (2020).
Laver, T. W. et al. SavvyCNV: Genome-wide CNV calling from off-target reads. PLoS Comput. Biol. 18, e1009940 (2022).
Haeussler, M. et al. Evaluation of off-target and on-target scoring algorithms and integration into the guide RNA selection tool CRISPOR. Genome Biol. 17, 148 (2016).
Glenn, T. C. et al. Adapterama I: universal stubs and primers for 384 unique dual-indexed or 147,456 combinatorially-indexed Illumina libraries (iTru & iNext). PeerJ 7, e7755 (2019).
Langmead, B. & Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359 (2012).
Li, H. et al. The sequence alignment/map format and SAMtools. Bioinformatics 25, 2078–2079 (2009).
Feng, J., Liu, T. & Zhang, Y. Using MACS to identify peaks from ChIP-Seq data. Curr. Protoc. Bioinforma. 2, 2.14.1–2.14.14 (2011).
Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21 (2013).
Li, B. & Dewey, C. N. RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinforma. 12, 323 (2011).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Acknowledgements
We are grateful to the patients and their families for taking part in our gene discovery study. We also thank S. Eurola, J. Ustinov and R. Ward for expert technical assistance. This work was supported by a Wellcome Trust Collaborative Award in Science to E.D.F., N.D.L.O., T.O., A.T.H. and M.I. (grant no. 224600/Z/21/Z). E.D.F. is a Diabetes UK RD Lawrence Fellow (19/005971) and has been the recipient of an EFSD Rising Star fellowship during this study. A.T.H. and S.E. were the recipients of a Wellcome Trust Senior Investigator award (grant no. WT098395/Z/12/Z) during this study and A.T.H. is employed as a core member of staff in the NIH Research-funded Exeter Clinical Research Facility and is an NIHR Emeritus Senior Investigator. M.I. has a Sir Henry Dale Fellowship jointly funded by the Wellcome Trust and the Royal Society (grant no. 206688/Z/17/Z). S.E.F. has a Wellcome Trust Senior Research Fellowship (grant no. 105636/Z/14/Z). N.D.L.O. has a lectureship funded by a Research England’s Expanding Excellence in England (E3) award. M.B.J. and M.N.W. are recipients of an Exeter Diabetes Centre of Excellence Independent Fellowship funded by E3. The work was supported by the National Institute for Health Research (NIHR) Exeter Biomedical Research Centre, Exeter, UK. R.E.J. is a Diabetes UK Harry Keen Clinician Scientist fellow (20/0006263) and received an Academy of Sciences Starter grant during this project. N.A.H. is funded by the Medical Research Council (grant nos. MR/000638/1 and MR/S036121/1). Most of the experimental studies were funded by the Academy of Finland Center of Excellence MetaStem (grant no. 312437), the Novo Nordisk Foundation (grant no. 0057286) and the Sigrid Juselius Foundation. H.M. is a member of the Doctoral Program in Integrative Life Science at University of Helsinki. A.T.’s doctoral studies were funded by the Foundation for Education and European Culture (IPEP) in Greece and by the Cambridge Trust. D.B. received funding from a European Molecular Biology Organization long-term fellowship (ALTF 295-2019). King Salman Center for Disability Research funded exome sequencing analysis for one case through Research Group no. RG-2022-010. For the purpose of open access, the author has applied a CC BY public copyright licence to any author accepted manuscript version arising from this submission.
Author information
Authors and Affiliations
Consortia
Contributions
E.D.F., N.D.L.O., T.O., A.T.H. and M.I. designed the study. E.D.F., M.N.W., S.E.F., R.C., S.E., S.M. and M.J.B. performed the genetic analysis. M.N.W., C.F.W., N.D.L.O. and M.I. performed the conservation analysis. E.D.F., A.T.H., F.S.A. and the Pancreatic Agenesis Gene Discovery Consortium analyzed the clinical data. R.E.J. and N.A.H. provided human embryo RNA-seq data. N.D.L.O. and M.I. analyzed the transcriptomic and epigenetic data, with J.A.K. contributing to transcriptome analysis. H.M., J.S.-V., H.I., D.B. and S.M. performed the genome editing and differentiation studies in hESCs. A.T. performed the ChIP–seq assay. E.D.F., N.D.L.O., H.M., T.O., A.T.H. and M.I. wrote the first draft of the manuscript. All authors edited the paper.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Genetics thanks Miguel Branco, Didier Trono and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 The ubiquitously expressed ZNF808 is the first primate-specific gene confirmed to cause a congenital developmental disease.
a. ZNF808 is expressed across GTEx adult tissues. ZNF808 is expressed at variable levels across adult tissues with no tissue absent in expression. Boxplots describe ZNF808 expression in the GTEx (Genotype-Tissue Expression) project. Data shows sum of GTEx v8 isoform level transcripts per million data calculated with RSEM. Boxplot central line denotes median, box limit the interquartile range and whiskers extend to furthest point within 1.5 interquartile range, data points are outliers exceeding whiskers, median sample size n = 291. b. ZNF808 is maximally expressed in the embryonic pancreas and minimally expressed in the embryonic liver. Human embryo RNA-seq spanning CS14-225 (data normalized as in original publication) for all MER11-binding KZFPs, tissues ordered by expression and mean of replicates shown. Dots give expression for all replicates, central line of boxplots denotes median, box marks interquartile range and whiskers 1.5x interquartile range. c. ZNF808 is the only primate-specific gene confirmed to cause a congenital developmental disease. Homology scores (including 1-to-1 and 1-to-many orthologues) for every protein-coding gene (Ensembl Biomart,10th February 2020, release 98) for 26 primates and 70 non-primate mammals. The difference between the maximum % identity difference across all primates versus the maximum across all non-primates to human was calculated for each gene. Frequency densities (density estimations scaled to group size at 0.5% bin size) shown. Genes without a non-primate ortholog are plotted as percent identity between humans and primates (red), equivalent to a gene with 0% identity between primates and non-primates. Genes are grouped into non-disease causing (not present in OMIM-morbid; gray), disease causing (present in OMIM-morbid; green) and genetic causes of developmental disorders (present on DDG2P; blue). ZNF808 is highlighted (dotted line, ZNF808 is erroneously annotated as having a non-primate ortholog and its homology between primates and non-primates given). All OMIM-morbid and DDG2P genes with sequence identity difference >40 were manually checked with no evidence of a primate-specific disease gene causing a congenital developmental disorder found (Supplementary Table 4).
Extended Data Fig. 2 MER11 elements subsets are enriched for various transcription factors.
a. Full list of hits from the ChIP-Atlas database with Fisher exact test right-tail p-value < 1e-100 enriched in various MER11 subfamilies, without multiple comparison adjustment. The color of each bubble is scaled with p-value and the radius with enrichment. If multiple experiments were found to be enriched for any given factor only the most significant value is shown. b. Signal from selected factors overlay on a multiple alignment of MER11 sequences to show various subsets of sequences being targeted by specific transcription factors and KZFPs. Top left - Overlay of signal from published KZFPs ChIP–seq8 on a multiple alignment of MER11A, MER11B and MER11C elements reveal that ZNF808 binds strongly in the centre of these elements. Five other KZFPs can be found on smaller subsets of elements - a clear pattern of semi-exclusive binding between ZNF808 and ZNF525 / ZNF578 is visible. All KZFPs binding MER11 elements represented here were found to be primate-specific at various levels. Multiple alignments generated with MAFFT v7.475 as performed in8.
Extended Data Fig. 3 ZNF808 KO and protocol of differentiation toward beta cells.
a. Differentiation protocol to generate pancreatic endoderm, progenitors and islets from human embryonic stem cells. b. Flow cytometry analysis for the definitive endoderm marker CXCR4. n = 3 independent differentiation experiments, data are presented as mean values ± SEM. Axis labels state the marker and fluorochrome used. (a.u.=arbitrary unit, S1=definitive endoderm stage). c. Flow cytometry analysis for the pancreatic progenitors markers PDX1 and NKX6-1 at S4 (pancreatic progenitors stage). n = 3 independent differentiation experiments, data are presented as mean values ± SEM. Axis labels state the marker and fluorochrome used. (a.u.=arbitrary unit). d. Immunohistochemistry analysis of S3 (posterior foregut stage) monolayer cells for PDX1 and alpha-fetoprotein (AFP); and S4 (pancreatic progenitors stage) monolayer cells for PDX1, NKX6-1, SOX9 and NGN3 (representative of 3 independent differentiation experiments, scale bar = 200 um).
Extended Data Fig. 4 Epigenetic profile of ZNF808 peaks.
a. Analysis of H3K9me3 ChIP–seq peaks intersecting with ZNF808 peaks reveals that the majority of sites are covered in heterochromatin-associated H3K9me3 at early differentiation stages in the wild type. There is a partial loss of H3K9me3-positive loci in the ZNF808 KO in stem cells and during differentiation. Analysis of H3K27ac reveals that a few sites are positive in wild-type cells but many more gain activity in the ZNF808 KO at all stages, particularly at S0 and S1. b. Analysis of ZNF808 peaks shows that around half have H3K9me3 status in at least one stage of differentiation and that the vast majority of those are transposable elements of the MER11 family. Amongst those, the majority loses H3K9me3 in at least one differentiation stage in the ZNF808 KO – 21.7% of those also gain H3K27ac. These are similarly highly enriched in MER11 elements, although with a different distribution of subfamilies. Interval intersections were performed using pybedtools 0.81 using repeats annotation from repeatmasker.org for hg19, version RepeatMasker open-4.0.5 - Repeat Library 20140131. Promoter coordinates were downloaded from Ensembl Biomart and extended 2.5 kb from the start site of all protein-coding genes. c. Heatmap showing H3K9me3 and H3K27ac signals over all MER11 elements in wild type and ZNF808 KO during differentiation. Different patterns of loss of H3K9me3 and gain of H3K27ac are visible. Color scale is based on z-score per element of reads with MAPQ > 20, ranging from +2 (red) to –2 (blue).
Extended Data Fig. 5 Dynamic epigenetic clusters of MER11 elements are enriched in transcription factors matching the stage where they are active.
a. Heatmap showing the epigenetic status of MER11 elements – this is a replicate from Fig. 3d to allow visual reference of clusters that gain H3K27ac at different points of the differentiation. b. Per cluster breakdown of subfamily of MER11 elements. Some trends can be observed, such as MER11A elements being found in increased proportion in cluster (#1, 5 and #6) which show H3K27ac signal gain at S4. c. Heatmap showing the percentage of intersection with ChIP–seq of selected transcription factors and their relationship with either all MER11 elements (top) or epigenetic active clusters identified in Fig. 3c. d. Similarly, here is shown the intersection of activated clusters with KZFPs found to be enriched in MER11 elements.
Extended Data Fig. 6 KZFPs and transcription factors binding MER11 elements have dynamic expression profiles during differentiation.
a, b. Expression of MER11-binding KZFPs. a- Heatmap showing maximum normalized mean expression in wild-type cells. b. Expression of all replicates in wild type and ZNF808 KO. c–d. Expression of MER11-binding transcription factors. As (a-b) for transcription factors GATA3, GATA4, GATA6, HNF4A, HNF4G.
Extended Data Fig. 7 MER11 elements are active in various cell types.
a. Overlap between accessible regions in 222 cell types from scATAC-seq human and fetal atlas and either all MER11 elements (blue) or the 220 elements that lose repression and gain activity in the ZNF808 KO (red). On the right, a zoom of the 15 cell types with the highest overlap in the ZNF808 KO activated MER11 elements. b. Heatmap of all MER11 elements activated in the ZNF808 KO intersecting with a peak of chromatin accessibility in the scATAC-seq dataset. c. Percentage of overlap with scATAC-seq peaks in select cell types and clusters of H3K27ac activity found in the ZNF808 KO during differentiation. Fetal syncytio/cytotrophoblasts are found in all 6 clusters at a higher percentage compared to MER11 background while other cell types are more enriched in cluster #6, which gains H3K27ac at S4. d. Examples of MER11 elements active in selected cellular contexts. All examples are taken from the list of MER11 losing H3K9me3 and gaining H3K27ac in the ZNF808 KO. e. Epigenetic status of MER11 elements in the NIH Roadmap dataset – biosamples were collapsed per cell type and chromatin state predictions at the single base pair level were used. Here we focus on two chromatin states classes we have built from the aggregate of multiple smaller ones, H3K9me3-positive heterochromatin (containing H3K9me3-associated categories “ZNF_Rpts” and “Het”) or Enhancer/TSS (aggregate of TSS-associated categories such as “TssA”, “TssFlnk”, “TssFlnkU”, “TssFlnkD” and enhancer-associated categories such as “EnhG1”, “EnhG2”, “EnhA1”, “EnhA2”, “EnhWk”). There are 18 active MER11 elements for ‘Pancreas’ versus 100 for ‘Liver’. Results show the same trends observed by ATAC-seq, notably that multiple MER11 elements are active in placenta or liver, but not in pancreas.
Extended Data Fig. 8 Validation of epigenetic and transcriptomic dysregulation observed in the ZNF808 KO cells using patient-derived iPSCs.
a. Heatmap of the 220 MER11 elements that lose H3K9me3 and gain H3K27ac in the ZNF808 KO as presented in Fig. 3, with the addition of signal obtained when differentiating iPSCs up to S3. b. Left, boxplots showing fold-change of activated and repressed dysregulated genes identified in ZNF808 KO and patient-derived iPSCs, showing agreement in direction and magnitude of the gene expression perturbation. Boxes mark interquartile range, with central line describing the median and whiskers 1.5x interquartile range. N = total activated and repressed dysregulated genes, see Fig. 4a. Right, linear regressions between ZNF808 KO and iPSC log2 fold-change KO over WT at same set of genes. Regression coefficients and p-value of slope term given. c. qRT-PCR for five hepatic marker genes assayed at the posterior foregut stage (S3) in cells derived from H1 control, H1-ZNF808-KO and the patient iPSC (line HEL340.7) carrying the ZNF808 deletion (n = 3-4 independent differentiation experiments). Data are presented as mean values ± SEM. Unpaired two-tailed t test.
Extended Data Fig. 9 Unmasked MER11 elements drive proximal gene activation in ZNF808 KO and dysregulated genes are associated with a loss of pancreatic identity and a gain of hepatic identity.
a. Example locus showing loss of repression and activation of a MER11 element in the ZNF808 KO with activation of the adjacent gene. Locus shows H3K9me3 and H3K27ac data in reads per million upstream of the TMTC1 promoter. H3K9me3-marked MER11 element in wild type is lost at S0 in ZNF808 KO concomitant with gain of H3K27ac. b. Gene expression for TMTC1 shown in transcripts per million (TPM), showing robust upregulation at S0, S1 and S2 stages. c. Proximity enrichments between pairs of MER11 clusters identified in Fig. 3 and genes activated at each stage. Fisher exact test right-tail –log10 p-value is denoted by color and odds ratio by size of dot. Minimal p-value for gene-element pairs in the range [1 bp, 100 Mb] and the odds ratio at that minimum given. d. Enrichr gene set enrichments for activated (orange) or repressed (blue) genes in ZNF808 KO selected gene sets and terms (see Supplementary Table 8) shown. Enrichments for activated genes are given for fetal liver, liver and regulation of TGF-beta, repressed otherwise. Fisher exact test right-tail –log10 p-value is denoted by color and odds ratio by size of dot.
Extended Data Fig. 10 Genes exclusively expressed in liver are activated in ZNF808 KO.
a. Heatmap of GTEx liver-exclusive genes. 357 genes for which their lower quartile of expression in liver exceeds the upper quartile of expression in all other tissues. Shown as z-score of median expression. Liver and pancreas highlighted. b. Over-representation of liver-exclusive genes in the list of genes activated in ZNF808 KO. Bubble plot with Fisher Exact test -log10 p-value denoted by color and odds ratio denoted by dot size. c-e. TDO2 is a liver-exclusive gene (c), activated at S2, S3 and S4 in ZNF808 KO (d). TDO2 is adjacent to two MER11 elements for which the leftmost loses H3K9me3 throughout the time course and gains H3K27ac from S1 onwards. Boxplots in (c) describe gene level TDO2 expression in the GTEx (Genotype-Tissue Expression) project. Data shows sum of GTEx v8 isoform level transcripts per million data calculated with RSEM. Boxplot central line denotes median, box limit the interquartile range and whiskers extend to furthest point within 1.5 interquartile range, data points are outliers exceeding whiskers, median sample size n = 291. The GTEx Project was supported by the Common Fund of the Office of the Director of the National Institutes of Health and by NCI, NHGRI, NHLBI, NIDA, NIMH and NINDS. The data used for the analyses described in this manuscript were obtained from the GTEx Portal on 08/26/21.
Supplementary information
Supplementary Information
Supplementary Methods and References.
Supplementary Tables
Supplementary Tables 1–8.
Source data
Source Data Fig. 2
Alignment of ZNF808 in higher primates.
Source Data Fig. 3
Uncropped gel image.
Source Data Extended Data Fig. 7
Source data of bar charts in Fig. 7a,e.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

{kind=link}
Cite this article
De Franco, E., Owens, N.D.L., Montaser, H. et al. Primate-specific ZNF808 is essential for pancreatic development in humans. Nat Genet 55, 2075–2081 (2023). https://doi.org/10.1038/s41588-023-01565-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41588-023-01565-x
- Springer Nature America, Inc.
This article is cited by
-
New ingredients for old recipes
Nature Genetics (2023)