

Abstract
The Altai region of Siberia was inhabited for parts of the Pleistocene by at least two groups of archaic hominins—Denisovans and Neanderthals. Denisova Cave, uniquely, contains stratified deposits that preserve skeletal and genetic evidence of both hominins, artefacts made from stone and other materials, and a range of animal and plant remains. The previous site chronology is based largely on radiocarbon ages for fragments of bone and charcoal that are up to 50,000 years old; older ages of equivocal reliability have been estimated from thermoluminescence and palaeomagnetic analyses of sediments, and genetic analyses of hominin DNA. Here we describe the stratigraphic sequences in Denisova Cave, establish a chronology for the Pleistocene deposits and associated remains from optical dating of the cave sediments, and reconstruct the environmental context of hominin occupation of the site from around 300,000 to 20,000 years ago.
Similar content being viewed by others


Main
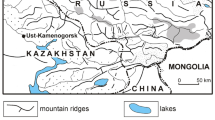
The period of existence and geographical distribution of Neanderthals and their sister group, the Denisovans1,2, are key unresolved issues in the study of human evolution and dispersal. Denisovan remains are known only from the type locality of Denisova Cave, which is located in the foothills of the Altai Mountains in southern Siberia (51° 23′ 51.3′′ N, 84° 40′ 34.3′′ E; Fig. 1a). Neanderthal remains have also been recovered from the deposits of this cave.

a, Map of southern Siberia and site location. b, Plan of cave interior showing Main Chamber, East Chamber and South Chamber; the entrance zone was not investigated in this study.
The cave consists chiefly of three large chambers—named Main Chamber, East Chamber and South Chamber (Fig. 1b)—each containing several metres of deposit. The stratified sequences are numbered by layer; layers with the same number in the different chambers are not necessarily equivalent in age2. Excavations have yielded Middle Palaeolithic stone artefacts and a variety of Upper Palaeolithic artefacts, as well as the remains of fauna and flora3,4,5,6,7. The fragmentary remains of four Denisovans, two Neanderthals and a daughter of Neanderthal and Denisovan parents have also been recovered8,9,10; their genomes have been sequenced1,2,11,12,13,14,15,16, as has DNA extracted from the Pleistocene sediments17.
The time of deposition of the youngest Pleistocene layers in the cave is currently constrained by radiocarbon (14C) ages for charcoal fragments and bone collagen. However, many of these ages approach or exceed the limit of 14C dating (about 50 thousand years ago (ka)) and the large spread in ages is consistent with field observations of post-depositional disturbance in some parts of the site (for example, layers 11.1 and 11.2 in East Chamber)2. Using a light-sensitive thermoluminescence signal18, seven dates have been reported for quartz grains extracted from the deeper deposits in Main Chamber3,19, ranging from 282 ± 56 ka (lower part of layer 22.2) to 69 ± 17 ka (layer 14.1). Although questions remain about some of the methodological aspects20, these dates broadly agree with biostratigraphic data for the cave deposits3. Geomagnetic excursions recorded in layers 22.1 and 22.2 (Main Chamber)21 were attributed to two events that are no longer supported22,23. DNA-based methods have been used to infer the ages of the hominin remains11,12,13,16,24. The oldest and youngest genetically dated Denisovan fossils—Denisova 2 (layer 22.1, Main Chamber) and Denisova 3 (layer 11.2, East Chamber), respectively—appear to differ in age by 54,200–99,400 years13, with Denisova 3 tentatively dated to 85 ± 8 ka (95.4% confidence interval)16,24. DNA-based ages have also been estimated for the so-called Altai Neanderthal (Denisova 5, 123 ± 7 ka)24 and the Neanderthal–Denisovan offspring (Denisova 11, dated to about 90 ka or later)16, the latter of which has been dated by 14C to more than 50 ka15. Further details are given in the Supplementary Information.
Collectively, these dates suggest that hominins have inhabited Denisova Cave intermittently since the middle of the Middle Pleistocene. However, accurate and more precise chronologies and a higher density of ages are required to address issues of site formation, hominin occupation and environmental conditions in the vicinity of the cave. Here we present details of the stratigraphy and report more than 100 new ages for the Pleistocene sequences in all three chambers, using optical dating of sediments25,26,27,28 to construct a common timeline for the cultural remains, hominin fossils and palaeoecological records.
Stratigraphy and site formation
Denisova Cave is formed in Silurian limestone and covers a total area of about 270 m2. Main Chamber is about 10 m high and contains approximately 6 m of deposits, which were first excavated 40 years ago. East Chamber and South Chamber are both narrow (less than 3 m wide) and extend away from Main Chamber (Fig. 1b and Extended Data Fig. 1a); their deposits have been excavated to depths of about 7 and 4.5 m, respectively. The upper 1.5–2 m of deposit in each chamber (layers 0–8) are Holocene in age. The underlying Pleistocene deposits consist of clay, silt and sand grains (that were blown or washed into the cave, or were reworked from pre-existing cave sediments) interstratified with coarser, angular limestone debris (gravel-size and larger) spalled from the floor, walls and roof of the cave owing to physical and chemical weathering of the bedrock.
The Pleistocene sequences (Fig. 2) are complex and include laterally discontinuous layers and unconformities between layers (due to erosional events or depositional gaps), as well as evidence of post-depositional modifications due to chemical diagenesis and freeze–thaw processes, subsidence and slumping, animal burrowing, and anthropogenic activities (Supplementary Tables 16–18). The layers cannot be traced stratigraphically between the chambers, but field observations of sedimentary characteristics and morphological features permit correlation of some Pleistocene layers (for example, 22).

a, Main Chamber. b, East Chamber. c, South Chamber. The southeast profile is shown for each chamber; these profiles were sampled for optical dating in 2012. Numbers denote stratigraphic layers, layer colours are notional and asterisks indicate areas of mixed deposit.
The Pleistocene deposits in Main Chamber comprise layers 9–22 and their subdivisions (Fig. 2a and Extended Data Figs. 2, 5a, b). Most of the layers are composed of a poorly sorted, sandy-silt matrix with abundant angular clasts of spalled bedrock. Some layers (for example, 9 and 21) have fewer coarse clasts. Many of the layer boundaries are indistinct and transitional. Layer 22.1 forms a peak-shaped feature in the southeast profile and incorporates many vertically tilted gravel clasts. The two overlying layers (21 and 20) follow the same topography. There is no evidence for substantial mixing of sediments between these three layers. Layer 12 marks the start of the sub-horizontal stratigraphy in this chamber. Parts of some layers show signs of animal burrowing (for example, 9 and 13), freeze–thaw processes (for example, 11 and 22.1), phosphatization (layer 9 in particular), manganese staining and other diagenetic alterations.
In East Chamber, the basal 2 m of deposit (layer 17.2) consists mainly of fine-grained sediments (silty clay) deposited in a former phreatic tube (Fig. 2b and Extended Data Figs. 3, 5c–e). The overlying layer (17.1) has a similar composition and is, in turn, overlain by a thin band of organic-rich sediment (layer 16); both layers follow the undulating topography of the narrow bedrock shelf that separates the phreatic tube from the chamber above. The remaining Pleistocene sequence (layers 15 to 9) is characterized by poorly sorted sediments composed of sandy silts with occasional-to-abundant angular clasts of limestone. Layers 17.1 to 11.1 dip inwards from the walls and sag towards the base of the chamber owing to post-depositional subsidence and possibly some slumping; carnivore coprolites (from cave hyenas) are common in these layers. Parts of layers 11 and 9 are heavily phosphatized29 and show signs of disturbance (lack of bedding), especially from animal burrowing activities. Sediment mixing may also have occurred close to the chamber walls, where fissures are evident and the stratigraphy is complex.
Excavations in South Chamber are currently limited to the entrance area, where layers 9–22 are tentatively recognized (Fig. 2c and Extended Data Fig. 4). The lowest excavated unit is provisionally identified as layer 22, on the basis of its similar colour and texture to layer 22 in Main Chamber and its unconformable contact with the overlying unit, tentatively identified as layer 20. The latter is overlain by layers 9–19, which consist of poorly sorted sediments (silty sand with scattered to abundant clasts of spalled bedrock), and are commonly separated by indistinct and transitional boundaries. Layer 9 contains few coarse clasts, but is extensively phosphatized and burrowed in places (Extended Data Fig. 4d).
Pleistocene chronologies
We constructed numerical chronologies for the Pleistocene sequences in all 3 chambers from optical dating25,26,27,28 of 103 sediment samples collected over 4 excavation seasons; 92 samples were dated from measurements of more than 280,000 individual grains of quartz and potassium-rich feldspar (K-feldspar), and 11 samples were dated using multi-grain aliquots of K-feldspar. Optical ages were determined using four methods of equivalent dose (De) estimation (see Methods) and were used to develop a Bayesian model for the depositional chronology in each chamber, to connect their stratigraphic sequences and to provide a composite age and environmental framework for hominin occupation of the site (Figs. 3, 4). We also compared these ages with previously published 14C ages2,15 for nine animal bones, a fragment of charcoal and a piece of Neanderthal bone from layers 11 and 12; no Denisovan remains have been dated directly. Details of sediment sample locations and collection, preparation, measurement, and data-analysis procedures are given in the Supplementary Information, together with the measured and modelled ages and related data.

Layers containing skeletal remains and sedimentary DNA of Denisovans and Neanderthals are denoted by circled specimen numbers and by ‘DNA’, respectively (red, Denisovans; blue, Neanderthals; both colours, mixed ancestry). Artefact assemblages are indicated to the right of each stratigraphic sequence. Climatic conditions are inferred from pollen and faunal records (orange, relatively warm; blue, relatively cold; white, no data). Optical dates (in ka) are the Bayesian model estimates with total uncertainties at 95.4% probability; layer 13 in Main Chamber (labelled ‘nm’) was not included in the age model. Isochrons (lines of equal age) are used to correlate stratigraphic layers between chambers, and time gaps are indicated by wavy lines.

a, Climatic conditions inferred from pollen and faunal records in Main Chamber, East Chamber and South Chamber (orange, relatively warm; blue, relatively cold; white, no data) plotted on a calendar-year timescale; the time span of each climatic interval is demarcated by the mean start and end ages for the relevant layers (Fig. 3). Marine isotope stage (MIS) boundaries follow a previous publication40. b, Fossils (circled specimen numbers) and sedimentary DNA (‘DNA’) of Denisovans (red) and Neanderthals (blue) and a fossil of mixed ancestry (both colours). c, Composite BDP-96 biogenic silica record of diatom productivity in Lake Baikal32, located about 1,600 km east of Denisova cave. d, LR04 stack of marine benthic foraminiferal isotopic (δ18O) data assembled from 57 globally distributed sites40 (VPDB, Vienna Pee Dee Belemnite).
Fifty-five sediment samples have been dated from Main Chamber (Extended Data Figs. 2, 5a, b). The optical ages are mostly in stratigraphic order and show that the deposits have accumulated episodically since the middle of the Middle Pleistocene (Fig. 3 and Extended Data Fig. 6). Finite ages for the deeper deposits (layers 22–13) could be obtained only using the K-feldspar post-infrared infrared stimulated luminescence (pIRIR) signal, because the optically stimulated luminescence (OSL) signal of quartz was saturated. The wide spread in dates for layer 13 (175–102 ka at 95.4% probability; Supplementary Table 7) is consistent with disturbance by burrowing animals3, so these ages were omitted from the Bayesian model. Layer 22 accumulated before 287 ± 41 ka, followed by deposition of layers 21 and 20 between 250 ± 44 ka and 170 ± 19 ka, and layers 19–14 between 151 ± 17 ka and 97 ± 11 ka (here and below, we give modelled age estimates and total uncertainties at 95.4% probability). Layer 12 was deposited between 70 ± 8 ka and 58 ± 6 ka, before the accumulation of layer 11 between 44 ± 5 ka and 38 ± 3 ka. The final Pleistocene deposits are represented by layer 9, which unconformably overlies layer 11 and has start and end dates of 36 ± 4 ka and 21 ± 8 ka, respectively. There is some uncertainty about layer assignments where the stratigraphic boundaries are indistinct and transitional, and the small-scale mixing of parts of some layers is indicated by scatter in the single-grain De distributions and optical ages.
Thirty-seven sediment samples have been dated from East Chamber (Extended Data Figs. 3, 5c–e). The deposits in layers 17–12 have a stratigraphically coherent chronology (Fig. 3 and Extended Data Fig. 7), limited age variation within any particular layer and De distributions that show minimal evidence for mixing, which we attribute to the lack of substantial post-depositional disturbance. Layer 17 accumulated before 284 ± 32 ka (saturation of the pIRIR signal prevented estimation of a finite age for the deepest sample), followed by the deposition of layers 16–12.2 between 259 ± 28 ka and 129 ± 11 ka. Layers 12.1 and 11.4–11.1 were deposited between 120 ± 11 ka and 38 ± 9 ka. None of the samples collected from layers 11.4 or 11.3 show evidence of mixing. Five out of ten samples from layers 11.2 and 11.1 also show good integrity and give date ranges of 63 ± 6 ka to 55 ± 6 ka and 49 ± 6 ka to 38 ± 9 ka, respectively. The other five samples—and those from layers 9 and 8 in the southeast profile—have layer assignments that are uncertain or produced scattered De distributions from which reliable optical ages could not be determined (see Supplementary Information). We attribute these De distributions to disturbance of the deposit by hyenas and other burrowing animals, resulting in mixing of grains—and presumably also artefacts and fossils—of various ages, consistent with the wide spread in 14C ages (from over 50,000 to about 19,000 calibrated years before present (bp; present is ad 1950) obtained for the upper part of layer 11 in this chamber2.
Eleven sediment samples were dated from South Chamber (Extended Data Fig. 4). The lowest sampled unit (layer 22) accumulated before 269 ± 97 ka, followed by a lengthy time gap before deposition of layers 19 and 14 between 136 ± 26 ka and 101 ± 19 ka (Fig. 3 and Extended Data Fig. 8a). Layer 12 was deposited between 64 ± 9 ka and 47 ± 8 ka, consistent with 14C ages of about 52,000 and over 50,000 calibrated years bp for bones from layer 11.2 (ref. 2).
We also compared the OSL and pIRIR ages for 47 samples dated using both quartz and K-feldspar grains as an internal test of consistency (Extended Data Fig. 8b). The mean age-ratio is 1.00 ± 0.01 (standard error of the mean) and the mean ratio for each chamber is also consistent with unity. This implies that the cave sediments were bleached sufficiently at deposition to reset both dating signals (at least over the time span of this comparison), as OSL traps are emptied more rapidly and completely than are pIRIR traps30,31. Collectively, the three chambers contain a near-complete sequence of Pleistocene deposits (with some noticeable gaps in sedimentation) and hominin occupation extending from about 300 to about 20 ka (Figs. 3, 4).
History of hominin occupation
The oldest hominin fossil from the cave is a Denisovan molar (Denisova 2) recovered in 1984 from the top of layer 22.1 in Main Chamber (328–246 ka); however, the exact provenance of this tooth cannot now be established definitively. Denisovan DNA was retrieved from the sediments of layer 15 in East Chamber (217–185 ka) and from hominin remains in East Chamber (Denisova 8, 132–93 ka, and Denisova 3, 69–48 ka) and South Chamber (Denisova 4, less than 47 ± 8 ka). Neanderthal DNA was retrieved from sediments that were deposited between 168 and 86 ka in Main Chamber (layers 19, 17 and 14) and between 205 and 172 ka in East Chamber (layer 14), and from hominin remains in East Chamber (Denisova 9, 150–118 ka, and Denisova 5, 132–93 ka); the age range for Denisova 9 applies also to Denisova 11. Figures 3 and 4 show the association between the hominin fossils, sedimentary DNA remains and stratigraphic layers in each chamber. The DNA-based ages13,24 for Denisova 5 and Denisova 8 are consistent with the optical ages for the associated layers; however, the estimates for Denisova 3 and Denisova 4 are older than expected based on their stratigraphic positions, whereas the ages for Denisova 11 and Denisova 2 are younger than expected. The omission of uncertainties associated with population size, mutation rate and generation interval in the genetic estimates16,24 may account for some of these discrepancies, as might the re-deposition of these isolated and fragmentary fossils.
Four main artefact phases have been identified for hominin occupation of Denisova Cave during the Pleistocene: the early Middle Palaeolithic, middle Middle Palaeolithic, Initial Upper Palaeolithic and Upper Palaeolithic (examples of each of these phases are shown in Extended Data Fig. 9). Figure 3 and Extended Data Table 1 show the association between the artefact phases and layers in each chamber, and Supplementary Table 1 lists the number of artefacts recovered from each layer in Main Chamber and East Chamber. Early Middle Palaeolithic stone tools have been recovered from sediments deposited in Main Chamber and South Chamber before 287 ± 41 ka and 269 ± 97 ka, respectively, with the further development of early Middle Palaeolithic assemblages up to 170 ± 19 ka and 187 ± 14 ka in Main Chamber and East Chamber, respectively. In general, early Middle Palaeolithic stone tools are characterized by discoidal and Kombewa cores, the presence of Levallois cores for manufacturing flakes, and they include various types of scrapers, denticulates and notched tools (Extended Data Fig. 9d). The overlying middle Middle Palaeolithic layers were deposited between 156 ± 15 ka and 58 ± 6 ka in Main Chamber and East Chamber, and between 136 ± 26 ka and 47 ± 8 ka in South Chamber. In these assemblages, primary flaking is characterized by the use of flat, discoidal and Levallois cores; isolated sub-prismatic cores have also been identified. Scrapers dominate the tool assemblage, although notched-denticulate pieces are well represented, and end-scrapers, burins, chisel-like tools and truncated flakes are also present (Extended Data Fig. 9c).
The deposition of the Initial Upper Palaeolithic layers started before 44 ± 5 ka and 63 ± 6 ka in Main Chamber and East Chamber, respectively, and after 47 ± 8 ka in South Chamber; we note that the earliest artefacts were recovered from layer 11.2 in East Chamber, which has been disturbed in places. The Initial Upper Palaeolithic assemblages include variants of parallel and radial flaking, and narrow-faced cores for blade production and Levallois flaking (Extended Data Fig. 9b). Scrapers dominate the stone artefacts and various Upper Palaeolithic-type lithic implements represent a large proportion of the assemblage, which also yielded a rich collection of bone tools and ornaments made from animal teeth and bones, ivory and gemstones. The accumulation of Upper Palaeolithic assemblages (Extended Data Fig. 9a) started in Main Chamber at 36 ± 4 ka, with a well-developed stone bladelet technology and a Upper Palaeolithic component that is distinct from that of the Initial Upper Palaeolithic assemblages, and from the prevailing background of scrapers.
The archaeological, fossil hominin and sedimentary DNA evidence therefore currently suggests that the cave was occupied by Denisovans from 287 ± 41 ka (or, conservatively, from 203 ± 14 ka) to 55 ± 6 ka or later, if Denisova 3 and Denisova 4 are not intrusive. Neanderthals were also present at the site between 193 ± 12 ka and 97 ± 11 ka.
Environmental context
We have reconstructed the Pleistocene environments in the vicinity of Denisova Cave over the last approximately 350,000 years from the remains of animals and plants in Main Chamber and East Chamber3,4,5,6,7 (Fig. 3 and Supplementary Information). Inferred conditions are relative to the current moderately continental climate (average January and July temperatures outside the cave of −16 °C and 18 °C, respectively). Reconstructions are based on analyses of the remains of 27 species of large mammals, more than 100 species of small vertebrates (40 species of small mammals, 66 taxa of birds and bones of fish, reptiles and amphibians) and pollen records for 30 species of tree, 36 shrub and grass taxa, and 6 species of spore-bearing plants.
The deposits that accumulated in late marine isotope stage (MIS) 9 did so under fairly warm and moderately humid conditions, characterized by forests dominated by pine and birch and a considerable admixture of broad-leaved species. The subsequent period of moderate cooling in early MIS 8 was marked by a reduction in forest cover, followed by an interval of relatively stable climate that was accompanied by the maximum spread of forest seen over the time span of our environmental reconstruction. Subsequent sedimentation in MIS 8 took place in a relatively cold and moderately humid climate, during which mountain tundra and forest–tundra landscapes developed in the vicinity of the cave. During MIS 7, and possibly early MIS 6, sediments were deposited under warm and relatively dry conditions, which were accompanied by the widespread emergence of pine–birch forests and broad-leaved species. Steppe and meadow–steppe communities flourished during the cold and dry periglacial conditions of terminal MIS 6.
Conditions were unstable during MIS 5, and incorporated environments that were variously drier, warmer, cooler and milder than the present-day environment. Climatic deterioration during MIS 4 was accompanied by multiple turnovers of plant associations under alternating dry and wet cold conditions; steppe and meadow–steppe communities expanded considerably, whereas birch, alder and broad-leaved species disappeared. During MIS 3, dark coniferous forests and meadow associations expanded when conditions were cool and humid, whereas mixed-grass communities and forest (including broad-leaved species) spread when conditions were warmer. In the millennia leading up to the last glacial maximum at about 20 ka, the climate fluctuated between moderately cold (dominated by spruce forests) and cold and dry (dominated by tundra–steppe associations).
There is broad agreement in the interpretation of climatic conditions between the three chambers—which can be linked using the chronological framework established in this study—and with global and other regional records32,33,34,35 (Fig. 4). The largest anomaly between inferred climate and timing is at approximately 150 ka, at which time the pollen assemblages in East Chamber and Main Chamber (layers 13 and 19.3, respectively) indicate the prevalence of hornbeam forest with a mixture of oak and Eurasian linden. This suggests a relatively warm and humid climate, whereas the optical ages place these layers in late MIS 6, during which period generally cold conditions are recorded in Lake Baikal32,33 (Fig. 4). We caution, however, that climatic conditions may fluctuate greatly within the time resolution of these records (350–1,000 years) and of the optical ages, which have uncertainties of several millennia.
Discussion
The combined Pleistocene sequences from all three chambers provide a near-continuous record of human and environmental history that spans the past three glacial–interglacial cycles (Figs. 3, 4 and Extended Data Table 1). Many layers show signs of post-depositional subsidence, but the deposits have by and large retained their stratigraphic integrity. Disturbance by animal burrowing and other activities is mostly restricted to parts of the uppermost Pleistocene layers, especially close to the cave walls. The timing of the four main artefact phases and the environmental reconstructions are broadly consistent between chambers when taking into account the sampling resolution and the uncertainties associated with palaeoenvironmental records and optical ages.
The oldest stone artefacts and Denisovan fossil indicate that hominins have occupied the cave since late MIS 9 (about 300 ka), with Denisovan DNA retrieved from sediments deposited during late MIS 7, at about 200 ka. The recovery of Denisovan skeletal and genetic remains in younger deposits implies their presence in the cave until early MIS 3 (about 55 ka) and possibly later. Neanderthals were also present from late MIS 7 or early MIS 6 until MIS 5 (about 100 ka), based on fossil and sedimentary DNA evidence. No remains of modern humans have yet been recovered that would confirm their presence at the site during the Late Pleistocene.
The data presented here and in recent genetic studies12,13,14,15,16,17,24 reveal that the cave was occupied during the last interglacial by Denisovans and two genetically distinct groups of Neanderthals. Denisovans may have survived until at least about 55 ka, by which time anatomically modern humans were present in other parts of Asia36,37,38,39. Ongoing investigations at this and other sites in the region may help to resolve the timing and possible causes of extinction of archaic hominins in southern Siberia, and the nature of any encounters—including interbreeding—between them and modern humans.
Methods
No statistical methods were used to predetermine sample size. The experiments were not randomized and investigators were not blinded to allocation during experiments and outcome assessment.
Sediment samples for optical dating were collected at Denisova Cave in 2012, 2014, 2016 and 2017. Samples were collected by hand at night using a red-light torch for illumination, and sealed in thick black plastic to prevent light exposure during transport to the University of Wollongong. Optical dating gives an estimate of the time since grains of minerals, such as quartz and K-feldspar, were last exposed to sunlight25,26,27,28. The time of sediment deposition is estimated by dividing the equivalent dose (De, the radiation energy absorbed by grains since deposition) by the environmental dose rate (the rate of supply of oinizing radiation to the grains since deposition). The De is estimated from laboratory measurements of the OSL from quartz or the pIRIR from K-feldspar. The OSL signal is bleached by light more rapidly and completely than the pIRIR signal30,31 (hence, consistent OSL and pIRIR ages suggests prolonged bleaching at deposition) but the latter signal saturates at much higher doses than the OSL signal27,28 (hence, finite ages can be obtained for older deposits). The dose rate is estimated from field and laboratory measurements of environmental radioactivity, plus the small contribution from cosmic rays.
We used both quartz and K-feldspar grains to develop a numerical chronology for the Pleistocene sequences in Denisova Cave. The ages of the older samples were determined solely from the K-feldspar pIRIR signal, owing to saturation of the quartz OSL signal. Standard procedures were used to isolate individual grains of quartz (180–212 μm in diameter) and K-feldspar (ranging in size between 90 and 212 μm in diameter, depending on availability), and grains were etched in hydrofluoric acid to remove the alpha-irradiated surface layer. De values for individual quartz grains were estimated using well-established OSL measurement and data-analysis procedures41,42,43, and De values for individual K-feldspar grains were determined using a two-step (200 °C and 275 °C) regenerative-dose pIRIR procedure44 and a recently developed standardized growth curve approach45,46. We analysed the single-grain pIRIR signals in two ways to estimate the final De values. A multiple-aliquot regenerative-dose procedure for K-feldspar was used to estimate the De values for the oldest deposits47. Details of sample preparation and De determination methods, experimental procedures, quality-assurance criteria and validation tests are given in the Supplementary Information.
The environmental dose rate includes contributions from beta particles (measured in the laboratory48 on a portion of each sample), gamma rays (measured by in situ gamma spectrometry at each sample location), cosmic rays49 and radioactive emitters internal to the K-feldspar and quartz grains. The latter dose rate is commonly trivial for quartz (we assumed a value of 0.032 ± 0.010 Gy per thousand years), but is substantial for K-feldspar owing to the beta dose from 40K; we measured the potassium contents of a subset of grains by wavelength-dispersive X-ray spectroscopy using an electron microprobe. The measured beta dose rates were corrected for grain-size attenuation, and the beta, gamma and cosmic-ray dose rates were adjusted for water content (based on the measured field values). The sample De values were divided by the corresponding total environmental dose rates to calculate the optical ages in calendar years before present; the associated uncertainties are at the 68.2% confidence level and include all known and estimated sources of random and systematic error. Details of dose-rate determination and the resulting data are presented in the Supplementary Information, together with the age determination and error estimation procedures.
A Bayesian age model was developed separately for Main Chamber, East Chamber and South Chamber on the OxCal platform50,51. Each age was input as a C_date in calendar years before ad 1950 with an associated uncertainty (the standard error of the mean); we used the weighted mean age for each of the samples with paired OSL and pIRIR ages. The sequence of stratigraphic layers in each chamber was used as prior information, and we also allowed for the probable existence of time gaps (modelled as intervals) in each stratigraphic sequence; the resulting ages, however, are insensitive to the choice of model. Each sequence was modelled as a series of phases, assuming that (a) the stratigraphically lowest phase is older than those above, and (b) the measured ages are unordered and uniformly distributed within each phase. Modelled age estimates were obtained for the start and end of each phase to facilitate chronological comparisons between the three chambers. Details of the Bayesian modelling and the resulting chronologies are provided in the Supplementary Information.
Code availability
All custom R code used to produce the results presented here are available from the corresponding authors on reasonable request.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this paper.
Data availability
All data generated and/or analysed during the current study are available from the corresponding authors on reasonable request.
References
Krause, J. et al. The complete mitochondrial DNA genome of an unknown hominin from southern Siberia. Nature 464, 894–897 (2010).
Reich, D. et al. Genetic history of an archaic hominin group from Denisova Cave in Siberia. Nature 468, 1053–1060 (2010).
Derevianko, A. P. et al. Paleoenvironment and Paleolithic Human Occupation of Gorny Altai: Subsistence and Adaptation in the Vicinity of Denisova Cave (Institute of Archaeology and Ethnography, Siberian Branch of the Russian Academy of Sciences, Novosibirsk, 2003).
Bolikhovskaya, N. S. & Shunkov, M. V. Pleistocene environments of northwestern Altai: vegetation and climate. Archaeol. Ethnol. Anthropol. Eurasia 42, 2–17 (2014).
Agadjanian, A. K. & Shunkov, M. V. Evolution of the Quaternary environment in the northwestern Altai. Archaeol. Ethnol. Anthropol. Eurasia 37, 2–18 (2009).
Bolikhovskaya, N. S., Kozlikin, M. B., Shunkov, M. V., Uliyanov, V. A. & Faustov, S. S. New palynological data from the unique Paleolithic site of Denisova Cave in northwest Altai. Bull. Moscow Soc. Natural. Biol. Series 122, 46–60 (2017).
Vasiliev, S. K., Shunkov, M. V. & Kozlikin, M. B. in Problems of Archaeology, Ethnography and Anthropology of Siberia and Neighbouring Territories Vol. 23 (eds Derevianko, A. P. et al.) 60–64 (Institute of Archaeology and Ethnography, Siberian Branch of the Russian Academy of Sciences, Novosibirsk, 2017).
Turner, C. G. II. in Chronostratigraphy of the Paleolithic in North Central, East Asia and America (ed. Derevianko, A. P.) 239–243 (Institute of History, Philology and Philosophy, Siberian Branch of the USSR Academy of Sciences, Novosibirsk, 1990).
Shpakova, E. G. & Derevianko, A. P. The interpretation of odontological features of Pleistocene human remains from the Altai. Archaeol. Ethnol. Anthropol. Eurasia 1, 125–138 (2000).
Mednikova, M. B. A proximal pedal phalanx of a Paleolithic hominin from Denisova Cave, Altai. Archaeol. Ethnol. Anthropol. Eurasia 39, 129–138 (2011).
Meyer, M. et al. A high-coverage genome sequence from an archaic Denisovan individual. Science 338, 222–226 (2012).
Sawyer, S. et al. Nuclear and mitochondrial DNA sequences from two Denisovan individuals. Proc. Natl Acad. Sci. USA 112, 15696–15700 (2015).
Slon, V. et al. A fourth Denisovan individual. Sci. Adv. 3, e1700186 (2017).
Prüfer, K. et al. The complete genome sequence of a Neanderthal from the Altai Mountains. Nature 505, 43–49 (2014).
Brown, S. et al. Identification of a new hominin bone from Denisova Cave, Siberia using collagen fingerprinting and mitochondrial DNA analysis. Sci. Rep. 6, 23559 (2016).
Slon, V. et al. The genome of the offspring of a Neanderthal mother and a Denisovan father. Nature 561, 113–116 (2018).
Slon, V. et al. Neandertal and Denisovan DNA from Pleistocene sediments. Science 356, 605–608 (2017).
Vlasov, V. K. & Kulikov, O. A. Radiothermoluminescence dating and applications to Pleistocene sediments. Phys. Chem. Miner. 16, 551–558 (1989).
Derevianko, A. P., Laukhin, S. A., Kulikov, O. A., Gnibidenko, Z. N. & Shunkov, M. V. First Middle Pleistocene age determinations of the Paleolithic in the Altai Mountains. Dokl. Akad. Nauk 326, 497–501 (1992).
Huntley, D. J. Letters: Vlasov & Kulikov’s method. Anc. TL 10, 57–58 (1992).
Derevianko, A. P., Gnibidenko, Z. N. & Shunkov, M. V. Middle Pleistocene excursions of the geomagnetic field in the strata of Denisova Cave. Dokl. Akad. Nauk 360, 511–513 (1998).
Roberts, A. P. Geomagnetic excursions: knowns and unknowns. Geophys. Res. Lett. 35, L17307 (2008).
Laj, C. & Channell, J. E. T. in Treatise on Geophysics (Volume 5: Geomagnetism) 2nd edn (ed. Schubert, G.) 343–383 (Elsevier, Amsterdam, 2015).
Prüfer, K. et al. A high-coverage Neandertal genome from Vindija Cave in Croatia. Science 358, 655–658 (2017).
Huntley, D. J., Godfrey-Smith, D. I. & Thewalt, M. L. W. Optical dating of sediments. Nature 313, 105–107 (1985).
Hütt, G., Jaek, I. & Tchonka, J. Optical dating: K-feldspars optical response stimulation spectra. Quat. Sci. Rev. 7, 381–385 (1988).
Roberts, R. G. et al. Optical dating in archaeology: thirty years in retrospect and grand challenges for the future. J. Archaeol. Sci. 56, 41–60 (2015).
Athanassas, C. D. & Wagner, G. A. Geochronology beyond radiocarbon: optically stimulated luminescence dating of palaeoenvironments and archaeological sites. Elements 12, 27–32 (2016).
Shunkov, M. V. et al. The phosphates of Pleistocene–Holocene sediments of the Eastern Gallery of Denisova Cave. Dokl. Earth Sci. 478, 46–50 (2018).
Rhodes, E. J. Dating sediments using potassium feldspar single-grain IRSL: initial methodological considerations. Quat. Int. 362, 14–22 (2015).
Smedley, R. K., Duller, G. A. T. & Roberts, H. M. Bleaching of the post-IR IRSL signal from individual grains of K-feldspar: implications for single-grain dating. Radiat. Meas. 79, 33–42 (2015).
Prokopenko, A. A., Hinnov, L. A., Williams, D. F. & Kuzmin, M. I. Orbital forcing of continental climate during the Pleistocene: a complete astronomically tuned climatic record from Lake Baikal, SE Siberia. Quat. Sci. Rev. 25, 3431–3457 (2006).
Grygar, T. et al. Lake Baikal climatic record between 310 and 50 ky bp: interplay between diatoms, watershed weathering and orbital forcing. Palaeogeogr. Palaeoclimatol. Palaeoecol. 250, 50–67 (2007).
Melles, M. et al. Sedimentary geochemisty of core PG1351 from Lake El’gygytgyn—a sensitive record of climate variability in the East Siberian Arctic during the past three glacial–interglacial cycles. J. Paleolimnol. 37, 89–104 (2007).
Frank, U. et al. A 350 ka record of climate change from Lake El’gygytgyn, Far East Russian Arctic: refining the pattern of climate modes by means of cluster analysis. Clim. Past 9, 1559–1569 (2013).
Liu, W. et al. The earliest unequivocally modern humans in southern China. Nature 526, 696–699 (2015).
Westaway, K. E. et al. An early modern human presence in Sumatra 73,000–63,000 years ago. Nature 548, 322–325 (2017).
Hershkovitz, I. et al. The earliest modern humans outside Africa. Science 359, 456–459 (2018).
Groucutt, H. S. et al. Homo sapiens in Arabia by 85,000 years ago. Nat. Ecol. Evol. 2, 800–809 (2018).
Lisiecki, L. E. & Raymo, M. E. A Pliocene–Pleistocene stack of 57 globally distributed benthic δ18O records. Paleoceanography 20, PA1003 (2005).
Jacobs, Z. & Roberts, R. G. Advances in optically stimulated luminescence dating of individual grains of quartz from archeological deposits. Evol. Anthropol. 16, 210–223 (2007).
Wood, R. et al. Towards an accurate and precise chronology for the colonization of Australia: the example of Riwi, Kimberley, Western Australia. PLoS ONE 11, e0160123 (2016).
Clarkson, C. et al. Human occupation of northern Australia by 65,000 years ago. Nature 547, 306–310 (2017).
Blegen, N. et al. Distal tephras of the eastern Lake Victoria basin, equatorial East Africa: correlations, chronology and a context for early modern humans. Quat. Sci. Rev. 122, 89–111 (2015).
Li, B., Jacobs, Z., Roberts, R. G., Galbraith, R. & Peng, J. Variability in quartz OSL signals caused by measurement uncertainties: problems and solutions. Quat. Geochronol. 41, 11–25 (2017).
Li, B., Jacobs, Z., Roberts, R. G. & Li, S.-H. Single-grain dating of potassium-rich feldspar grains: towards a global standardised growth curve for the post-IR IRSL signal. Quat. Geochronol. 45, 23–36 (2018).
Li, B., Jacobs, Z. & Roberts, R. G. An improved multiple-aliquot regenerative-dose (MAR) procedure for post-IR IRSL dating of K-feldspar. Anc. TL 35, 1–10 (2017).
Bøtter-Jensen, L. & Mejdahl, V. Assessment of beta dose-rate using a GM multicounter system. Nucl. Tracks Radiat. Meas. 14, 187–191 (1988).
Prescott, J. R. & Hutton, J. T. Cosmic ray contributions to dose rates for luminescence and ESR dating: large depths and long-term time variations. Radiat. Meas. 23, 497–500 (1994).
Bronk Ramsey, C. Bayesian analysis of radiocarbon dates. Radiocarbon 51, 337–360 (2009).
Bronk Ramsey, C. & Lee, S. Recent and planned developments of the program OxCal. Radiocarbon 55, 720–730 (2013).
Acknowledgements
This study was funded by the Australian Research Council through fellowships to Z.J. (FT150100138), B.L. (FT140100384) and R.G.R. (FL130100116), the Russian Science Foundation (project 14-50-00036 to A.P.D. and A.K.A.), the Russian Foundation for Basic Research (project 17-29-04206 to M.V.S., M.B.K., V.A.U., N.S.B. and S.K.V.) and the state task of the Ministry of Education and Science of the Russian Federation (project 33.867.2017/4.6 to A.P.D.). We thank Y. Jafari, T. Lachlan, D. Müller, D. Tanner, V. Vaneev and the Australian Microscopy & Microanalysis Research Facility for assistance and P. Goldberg for comments on an earlier version of this Article.
Reviewer information
Nature thanks G. Duller, E. J. Rhodes and the other anonymous reviewer(s) for their contribution to the peer review of this work.
Author information
Authors and Affiliations
Contributions
Z.J., A.P.D. and R.G.R. conceived this study. M.V.S., M.B.K. and A.P.D. led the excavations and artefact analyses, and Z.J. and B.L. performed the optical dating, with contributions from K.O. and R.G.R. The other analyses were led by V.A.U. (stratigraphy and geoarchaeology), N.S.B. (palynology) and A.K.A. and S.K.V. (palaeontology). Z.J., M.V.S., M.B.K. and R.G.R. wrote the main text with contributions from all authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Plan maps of Main Chamber, East Chamber and South Chamber in Denisova Cave.
a, Detailed plan of cave interior, showing year of excavation in each of the chambers and at the cave entrance. b, Photograph of the present-day cave entrance, with people on the left for scale. c–e, Plan maps of Main Chamber (c), South Chamber (d) and East Chamber (e), with the sequence of excavations in each chamber denoted by colour shading: from orange (earliest) through blue, pink, green and yellow, to white (most recent), with unexcavated areas in grey. Crosses denote locations of optical dating samples in each profile.
Extended Data Fig. 2 Stratigraphic sequences and locations of optical dating samples in Main Chamber.
a, b, Southeast profile after excavations in 1984 (a) and locations of samples collected from this profile in 2012 (blue) and 2014 (red) (b). c, d, Southeast profile after excavations in 2016 (c) and locations of samples collected from this profile in 2016 (yellow) (d). The number next to each filled circle represents the sample code (for example, number 1 next to the blue-filled circle in b denotes sample DCM12-1).
Extended Data Fig. 3 Stratigraphic sequences and locations of optical dating samples in East Chamber.
a, b, Southeast profile after excavations in 2012 (a) and locations of samples collected from this profile in 2012 (blue) and 2014 (red) (b). c, d, Southeast profile after excavations in 2015 (c) and locations of samples collected from this profile in 2016 (yellow) (d). e, f, Lower section of northwest profile (that is, below the choke-stone blocking the gap between the middle and lower sections of the hourglass-shaped cave profile) after excavations in 2014 (e) and locations of samples collected from this profile in 2014 (f). The number next to each filled circle represents the sample code (for example, number 1 next to the blue-filled circle in b denotes sample DCE12-1).
Extended Data Fig. 4 Stratigraphic sequences and locations of optical dating samples in South Chamber.
a–c, Southeast profile after excavations in 2003 (a, b) and after excavations in 2016 (c), showing locations of samples collected in 2012 (blue) and 2016 (yellow). The number next to each filled circle represents the sample code (for example, number 1 next to the blue-filled circle denotes sample DCS12-1). d, Area enclosed by the white square in c. Sample DCS16-2 is located about 13 cm below the base of the Holocene deposits and to the left of an infilled animal burrow, and immediately beneath a zone of phosphatization.
Extended Data Fig. 5 Additional stratigraphic sequences and sampling locations in Main Chamber and East Chamber.
a, Upper section of east profile in Main Chamber, showing locations of optical dating samples collected in 2014. b, Lower section of east profile in Main Chamber, showing locations of samples collected in 2014. c, Upper section of northwest profile in East Chamber, showing locations of samples collected in 2014 (red) and 2016 (yellow). d, Middle section of northwest profile in East Chamber, showing locations of samples collected in 2016. The lower section of this profile is shown in Extended Data Fig. 3f. e, West profile in East Chamber, showing location of sample DCE14-1.
Extended Data Fig. 6 Bayesian model of optical ages for deposits in Main Chamber.
Ages (n = 43) have been modelled in OxCal version 4.2.4. Only random errors are included in the age model. Pale probability distributions represent the unmodelled ages (likelihoods) and dark grey distributions represent the modelled ages (posterior probabilities). The narrow and wide brackets beneath the distributions represent the 68.2% and 95.4% probability ranges, respectively. Start and end ages have been modelled for each phase, with age ranges (95.4% confidence interval, random-only errors) given in years and rounded off to the closest decade.
Extended Data Fig. 7 Bayesian model of optical ages for deposits in East Chamber.
Ages (n = 28) have been modelled in OxCal version 4.2.4. Only random errors are included in the age model. Pale probability distributions represent the unmodelled ages (likelihoods) and dark grey distributions represent the modelled ages (posterior probabilities). The narrow and wide brackets beneath the distributions represent the 68.2% and 95.4% probability ranges, respectively. Start and end ages have been modelled for each phase, with age ranges (95.4% confidence interval, random-only errors) given in years and rounded off to the closest decade.
Extended Data Fig. 8 Bayesian model of optical ages for deposits in South Chamber and comparison of OSL and pIRIR ages for all three chambers.
a, Ages (n = 11) have been modelled in OxCal version 4.2.4. Only random errors are included in the age model. Pale probability distributions represent the unmodelled ages (likelihoods) and dark grey distributions represent the modelled ages (posterior probabilities). The narrow and wide brackets beneath the distributions represent the 68.2% and 95.4% probability ranges, respectively. Start and end ages have been modelled for each phase, with age ranges (95.4% confidence interval, random-only errors) given in years and rounded off to the closest decade. b, Comparison of single-grain OSL ages for quartz and single-grain pIRIR ages for K-feldspar for 47 samples (28 from Main Chamber, 15 from East Chamber and 4 from South Chamber), with age uncertainties shown at 2σ. The dashed line indicates the 1:1 ratio.
Extended Data Fig. 9 Palaeolithic artefacts from Main Chamber, East Chamber and South Chamber.
a, Upper Palaeolithic artefacts. 1–3, bladelets; 4, retouched blade; and 5, end-scraper. b, Initial Upper Palaeolithic artefacts. 1, marble ring; 2, tubular beads; 3, ivory pendant; 4, pendant made of red deer tooth; 5, ivory ring; 6, pendant made of elk tooth; 7, chloritolite bracelet; 8, bone needle; 9, end-scraper; 10, retouched point; 11, biface; and 12, pointed blade. c, Artefacts of middle Middle Palaeolithic assemblage. 1, blade; 2 and 5, Mousterian points; 3, Levallois point; and 4, scraper. d, Early Middle Palaeolithic assemblage. 1, core; 2 and 4, scrapers; 3, denticulate tool.
Supplementary information
Supplementary Information
This file contains: Section 1 Methods of excavation and analysis of the Pleistocene deposits, fauna and flora; Section 2 Previous chronologies and optical dating; Section 3 Stratigraphy and sedimentology; Section 4 Pleistocene environments, palynology and palaeontology; and Supplementary References
Rights and permissions
About this article

Cite this article
Jacobs, Z., Li, B., Shunkov, M.V. et al. Timing of archaic hominin occupation of Denisova Cave in southern Siberia. Nature 565, 594–599 (2019). https://doi.org/10.1038/s41586-018-0843-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-018-0843-2
- Springer Nature Limited
This article is cited by
-
More than a decade of genetic research on the Denisovans
Nature Reviews Genetics (2024)
-
Late Pleistocene emergence of an anthropogenic fire regime in Australia’s tropical savannahs
Nature Geoscience (2024)
-
New dating indicates intermittent human occupation of the Nwya Devu Paleolithic site on the high-altitude central Tibetan Plateau during the past 45,000 years
Science China Earth Sciences (2024)
-
Lithic technologies at Guanyindong cave, Southwest China: diversity and innovation during the Chinese Middle Palaeolithic
Archaeological and Anthropological Sciences (2024)
-
Qaleh Kurd Cave (Qazvin, Iran): Oldest Evidence of Middle Pleistocene Hominin Occupations and a Human Deciduous Tooth in the Iranian Central Plateau
Journal of Paleolithic Archaeology (2024)