
Abstract
Impaired executive inhibition is a core deficit of attention deficit hyperactivity disorder (ADHD), which is a common childhood-onset psychiatric disorder with high heritability. In this study, we performed a two-stage genome-wide association study of executive inhibition in ADHD in Han Chinese. We used the Stroop color-word interference test to evaluate executive inhibition. After quality control, 780 samples with phenotype and covariate data were included in the discovery stage, whereas 922 samples were included in the replication stage. We identified one new significant locus at 7p22.3 for the Stroop word interference time (rs11514810, P=3.42E−09 for discovery, P=0.01176 for replication and combined P=5.249E−09). Regulatory feature analysis and expression quantitative trait loci (eQTL) data showed that this locus contributes to MICALL2 expression in the human brain. Most genes in the network interacting with MICALL2 were associated with psychiatric disorders. Furthermore, hyperactive-impulsive-like behavior was induced by reducing the expression of the zebrafish gene that is homologous to MICALL2, which could be rescued by tomoxetine (atomoxetine), a clinical medication for ADHD. Our results suggested that MICALL2 is a new susceptibility gene for executive inhibition deficiency related to hyperactive-impulsive behavior in ADHD, further emphasizing the possible role of neurodevelopmental genes in the pathogenic mechanism of ADHD.
Similar content being viewed by others


Introduction
Attention deficit hyperactivity disorder (ADHD) is a common psychiatric disorder with a worldwide prevalence of ~5%.1 Genetic epidemiological studies have revealed that gene variants constituted the primary etiology of ADHD, with a heritability estimated to be 0.76.2 Candidate gene association studies have identified several genes involved in the biosynthesis, release, transmission and metabolism of neurotransmitters contributing to the development of ADHD.3 However, these genes had only a 3.3% effect on the phenotypic variation of ADHD and interpreted only 4.3% of the average heritability.2, 4, 5, 6 In recent years, several genome-wide association studies (GWAS) of ADHD suggested that some genes related to neuronal development might be associated with ADHD,7, 8, 9, 10 but none of these genes had a significant genome-wide association with the behavioral phenotype. Thus, new and validated pathogenic genes remain to be discovered.
One reason for the lack of significant genetic results is that ADHD is a heterogeneous disorder. The same clinical presentation of inattention, hyperactivity and impulsivity may have different etiological contributors. Endophenotypes are intermediate phenotypes, which are involved at a level between genes and phenotypes, and are closer to the gene function. Many neuropsychological features have been suggested to be endophenotypes of ADHD, of which inhibition has the most supporting evidence.11, 12, 13, 14, 15 Inhibition is one important component of executive function,16, 17, 18, 19, 20 which regulates general cognitive processes. The psychological concept of inhibition refers to negative control by the higher centers over the lower centers of the nervous system. According to previous studies, impaired inhibition, which is closely related to the symptom impulsivity, was the core impairment in ADHD patients.21, 22, 23 Therefore, revealing the mechanism of impaired inhibition may help to develop a deep understanding of the etiology of ADHD.
As catechol-O-methyltransferase (COMT) largely regulates the synaptic availability of dopamine in the prefrontal cortex, which mediates executive function, several investigators have tested the association of COMT with aspects of executive function. The association of COMT with set shifting was first reported in a healthy sample.24 Using a stop-signal task, Mione et al.25 reported an interaction between COMT Val158Met and gender on response inhibition, showing that male subjects with the Val/Val genotype had poorer inhibition abilities. This association was further demonstrated by van Goozen et al.26 in 194 male adolescents with ADHD, in which the COMT Val allele predicted poorer response inhibition and set-shifting abilities. A significant association was also found between the 5-HTT LL genotype and inhibition in both adolescents and adults,27 in light of the regulatory effect of serotonin in impulsivity. Further neuroimaging analysis revealed that variation in 5-HTT affects neural activation in the inferior and medial frontal and temporal/parietal regions of the response inhibition network.28 In a gene–environment interaction study, short-allele carriers displayed increased activation of the right fusiform gyrus compared with long-allele carriers during failed inhibition.29 Additionally, several other genes were investigated to identify their behavioral-cognitive phenotype to explore ADHD-related endophenotypes. Cummins et al.30 used a larger sample size and identified the association of ADRA2A variants with increased intraindividual variability in response time as well as with ADHD-like behaviors. However, no GWAS of inhibition traits in ADHD had been conducted until now.
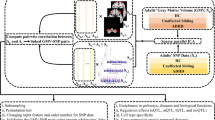
In this study, we conducted the Stroop color-word interference test in our ADHD samples to measure their inhibition function. GWAS for the two traits were then conducted in two independent ADHD samples to find and replicate inhibition-related genetic loci. Furthermore, a series of bioinformatics analyses and gene knockdown experiments in an animal model were performed to investigate the function of the associated loci and the potential effect of the associated genes on behavior. The findings may provide strong evidence for and new insight into the pathological mechanisms of executive inhibition and ADHD.
Materials and methods
Study samples and executive inhibition assessment
All the participants, who were recruited from the Child and Adolescent Psychiatric Outpatient Department of Peking University Sixth Hospital, met the DSM-IV ADHD diagnostic criteria. The clinical diagnosis was first made by a senior child and adolescent psychiatrist based on the parent- and teacher-completed ADHD Rating Scale-IV and was then confirmed by a semistructured interview with the parents and child, performed using the Chinese version of the Clinical Diagnostic Interview Scale.31 Those with major neurological disorders (e.g., epilepsy, schizophrenia, pervasive development disorder and mental retardation (IQ <70)) were excluded;7 IQ was assessed using the Chinese version of the Wechsler Intelligence Scale for Children, third edition. Most subjects were drug naive. For those who had been medicated, the drug was washed out for at least 1 month before the patient was recruited. The study was approved by the Institutional Review Board of Peking University Sixth Hospital. Written informed consent was obtained from parents of the ADHD probands. We used a two-stage study design and collected consecutive samples. The discovery sample was 1040 children and adolescents with ADHD, who were recruited before August 2011. Those recruited thereafter constituted the replication sample, with 1192 cases.
We used the Stroop color-word interference task to assess executive inhibition. The child and adolescent psychiatrist monitored all the tests conducted on patients and collected the data. The task included four sessions. At the beginning, thirty stimuli were presented in a 10 × 3 matrix for three cards each (21 × 29.7 cm2). In the first session, the subjects were asked to read the names of colors (red, green, yellow and blue) printed in black ink. In the second session, they were asked to name the colored squares (red, green, yellow and blue). In the third session, the subjects were asked to read the color words printed in different colors. In the fourth session, they were asked to name the colors of the ink. The time required to complete each session was recorded. The color interference time (CIT) equals to the time required to complete session 3 minus that for session 1, whereas the word interference time (WIT) equals to the time required to complete session 4 minus that for session 2. Both the CIT and WIT reflected interference inhibition.
Genotyping, quality control and association test
For the first stage of the study, genotypes were obtained using the Affymetrix 6.0 array from CapitalBio (Beijing, China). After mapping the single-nucleotide polymorphism (SNP) probes to SNPs with rs numbers, 653 428 SNPs remained. A total of 1026 cases remained after removing those individuals with per-individual autosomal heterozygosity >5 s.d. away from the mean, without age or IQ information, with a per-individual call rate <95% or with relatives with a genome identity PI_HAT ⩾0.185. Then, principal component analysis was conducted for the remaining samples using the SNPs with low linkage disequilibrium (LD) using the EIGENSOFT 4.2 software,32, 33 as described previously.34 Only the eigenvector 1 was significant in the Tracy–Widom test and thus was used as a covariate in the subsequent statistical analysis. Furthermore, 644 166 SNPs remained after removing those SNPs with a per-SNP call rate <98%, a Hardy–Weinberg equilibrium test P<0.001, or a minor allele frequency <1%. Association analysis was conducted for each quantitative trait using an additive linear regression model in PLINK,35 with age, IQ, sex and eigenvector 1 of principal component analysis as covariates. Genotyping at the replication stage was performed using the iPLEX MassARRAY Platform (Sequenom, San Diego, CA, USA). After excluding individuals with a call rate <95%, three SNPs with genotype data in 1174 samples were used for the replication stage and for the combined analysis with the discovery GWAS. Association analyses for the replication stage were implemented, with sex, IQ and age as covariates. In the combined analysis, eigenvector 1 was also used as a covariate (eigenvector 1 was set to zero for the samples from replication). P<5 × 10−8 was considered genome-wide significance. All reported P-values were two sided.
Imputation and association analysis after imputation
We used MACH-admix 1.036 to impute non-genotyped SNPs, using the ASN data (286 individuals) from the 1000 Genomes Project Integrated Phase 1 Release37 as the reference panel. Imputed SNPs with a squared correlation between imputed and true genotypes (rsq)<0.6 or SNPs with minor allele frequency <0.01 were removed. Association analysis after imputation was performed using mach2qtl.38 For significant loci, to combine the discovery stage data and the replication stage data, the genotype was extracted from the imputation data, and the association results were analyzed using PLINK.
Regulatory feature analysis and network construction
The regulatory features related to the significant SNP were obtained from rVarBase39 and ENCODE in UCSC Genome Browser.40 The LD-block region denoted by rs11514810 and its LD proxies were used as the input in rVarBase and UCSC ENCODE Browser. In rVarBase, we checked the regulatory features by mapping each SNP in this block. In UCSC ENCODE Browser, tracks included UCSC Genes, transcription, integrated regulation from ENCODE for the H3K27ac mark and DnaseI hypersensitivity clusters, transcription factor ChIP-Seq uniform peaks, chromatin state segmentation by HMM, chromatin interaction analysis paired-end tags and common SNPs (146). To explore the affected gene expression, the expression quantitative locus for rs11514810 was examined in the UK Brain Expression Cohort data set (GSE46706).41 Detailed processing and exclusion criteria have been described elsewhere,41 and eQTL analysis was described by Ramasamy et al.42 The expression plot was generated using BRAINEAC (http://www.braineac.org/) by searching for MICALL2, selecting its transcript, t3035223, and then stratifying its expression by SNP rs11514810. The MICALL2-interacting genes were from PINA v243 and InWeb,44 and the MICALL2-coexpressed genes were from GeneMANIA.45
Validation of MICALL2 function in zebrafish
The wild-type Tübingen strain zebrafishes used in this study were provided by the College of Life Science at the Peking University. The animal experiments were approved by the Institutional Animal Care and Use Committee of Peking University (LSC-LiuD-01). A Micall2b splice-blocking morpholino oligonucleotide (MO) was designed to bind exon 2/intron 2 to inhibit Micall2b splicing after transcription (Figure 3a). Another MO with 5 bp mismatches was used as a control (hereafter called MIS). Embryos were injected with 1 nl of MO/MIS at the one-cell stage and maintained at 28.5 °C in E2 zebrafish embryo medium. Thirty embryos were fixed in TRIzol reagent (Invitrogen, CarIsbad, CA, USA) at 3 days post fertilization (d.p.f.), and RNA was extracted using isopropyl alcohol and trichloromethane and then reverse transcribed into cDNA using a FastQuant RT Kit (with gDNase) (Tiangen, Beijing, China). Real-time PCR was performed to amplify cDNA and to detect changes in gene expression. The locomotion of larvae was analyzed at 6 d.p.f. Larvae injected with MO in E2 and four doses of tomoxetine (produced by Tocris Bioscience, Bristol, UK; alternative name is atomoxetine, 5, 10, 15, 20 μM) as well as larvae injected with the MIS control were placed in a 48-well plate, with one larva per well; 8 larvae for each group. The total sample size was estimated by experience in reference to previous studies. All larvae were habituated in the well for 10 min. The total swimming distance and average velocity of the larvae were recorded for 15 min using a Danio Vision Tracking System from Noldus Information Technology (Wageningen, The Netherlands). The data were analyzed using Ethovision 10.0 software (Noldus Information Technology).
Results
SNPs associated with the Stroop color-word interference test
After quality control for the genotype data as well as phenotype and covariate data cleaning were performed, a total of 780 individuals tested with the Stroop color-word interference test were included in the GWAS discovery stage; one individual had a missing CIT value. In the replication stage, 922 samples were included. The demographic description, IQ information and cognitive-behavioral phenotype data comparison for the discovery and replication samples are presented in Table 1. The distribution of the CIT and WIT for the discovery stage and the replication stage are shown in Supplementary Figure 1.
Three loci showing significance or trend significance with the Stroop WIT in the discovery stage were identified through association analysis (Table 2). One of these loci is the significant locus in 7p22.3 (rs11514810, P=3.42E−09, BETA=14.95 (10.05, 19.85)) (regional plot shown in Figure 1). Another locus located in 15q22.31 near gene MEGF11 (rs5016832, P=5.04E−07, BETA=18.26 (11.2, 25.33)) showed trend significance. Association analysis for the imputation data revealed another significant locus for the WIT near microRNA gene MIR8079 in 13q14.11 (rs73182927, P=6.92E−09, BETA=56.51 (37.6, 75.42), rsq=0.94). No significant results were found for the CIT. We attempted to replicate these three loci for the WIT in the replication stage with 922 samples. SNP rs11514810 reached the significance threshold in the combined analysis (P=0.01176, BETA=5.099 (1.14, 9.058) in replication stage, P=5.294E−09, BETA=9.31 (9.211, 9.41) in the combined samples). The other two SNPs were not replicated.

Regional plot for the significant locus rs11514810 for the Stroop word interference time.
Cross-trait validation for the inhibition-related loci
To further validate the loci identified as inhibition related, we performed cross-trait validation for SNPs rs11514810 and rs73182927 as well as their LD proxies (r2>0.75) on two Stroop traits (WIT and CIT). As shown in Supplementary Table 1, the most significant SNP, rs11514810, for the WIT as well as its several LD proxies also showed a nominal association with the CIT in both the discovery samples and the replication samples. Rs73182927 near MIR8079 and its LD proxies were also validated for the CIT in the discovery stage. These findings indicate that Stroop word interference and Stroop color interference may share some common genetic basis.
Besides the inhibition trait, we have collected three-dimensional symptoms, namely inattention (CDISatt), hyperactivity-impulsivity (CDIShi) and overall assessment (CDISall), for the patients according to the Clinical Diagnostic Interview Scale.21 First, we calculated the correlations between the Stroop test traits (CIT and WIT) and these three symptom traits (as shown in Supplementary Table 2). The result showed that increased Stroop test scores were associated with the increases in each of the ADHD symptom scores, in which WIT is significantly correlated with hyperactivity-impulsivity score and overall assessment. Furthermore, we examined the role of WIT as a mediator to mediate the association between rs11514810 with the ADHD symptoms. We used the model 4 in PROCESS46 to bootstrap the sampling distribution of the indirect effect (where the indirect effect is the reduction in the strength of the SNP/symptom association that is due to the WIT). The indirect effect of rs11514810 on the hyperactivity-impulsivity score (CDIShi) through WIT had a point estimate of 1.0238 and a 95% bias-corrected bootstrap confidence interval of 0.3311–2.5212, which means the mediation effect was different to zero even at the lower bound of the confidence interval. These data showed that SNP rs11514810 accounts for significant variation in ADHD hyperactivity-impulsivity symptoms, in part through the effects of the SNP on the intermediate phenotype of inhibitory control.
Genetic mechanism of the significant locus rs11514810
Given the strong association of rs11514810 with the WIT, we further examined the regulatory function and related genes of rs11514810. LD analysis identified 66 LD proxies for this SNP (r2>0.75). Related regulatory elements identified when searching for rs11514810 and its LD proxies included a chromatin interactive region in MCF-7 and K562 cell lines. The chromatin interactive region containing this LD block spans genes MICALL2, INTS1 and PSMG3, among which MICALL2 is the nearest gene to rs11514810. Regulatory features of SNP rs11514810 and its LD block from ENCODE are shown in Supplementary Figure 2. One active regulatory element (an H3K27ac mark) region exists in this LD block (blue box), along with several active chromatin state segmentations and transcription factor peaks, which may regulate the expression of nearby genes. To assess the potential function of this locus in brain tissue, we tested the association of SNP rs11514810 with MICALL2 gene expression. eQTL data from the UK Brain Expression Cohort41 showed that rs11514810 affected MICALL2 expression in the intralobular white matter, the hippocampus, the temporal cortex and the occipital cortex (specifically the primary visual cortex) (Figure 2). The minor allele T of rs11514810, associated with strong inhibition dysfunction, decreased MICALL2 expression in several brain regions. In normal samples, MICALL2 is expressed in brains at different development stages (Supplementary Figure 3); thus, decreased MICALL2 expression may affect brain functioning. Furthermore, MICALL2 interacting with CasL-like 2 has been reported to function in plexin-mediated axonal repulsion.47 To further explore the specific function of MICALL2, we mapped a network including MICALL2 and its interacting or coexpressed genes (as shown in Supplementary Figure 4). Among the 24 genes, all 6 interacting genes and 6 of 18 coexpressed genes had specific evidence indicating their association with psychiatric disorders, including ADHD,48 schizophrenia49 and major depressive disorder50 (Supplementary Table 3). Thus, MICALL2 may contribute to disorders related to impaired inhibitory control by interacting with these genes.

The effect of rs11514810 on MICALL2 expression. The study of expression quantitative trait loci in brain tissue demonstrates the effect of rs11514810 on MICALL2 gene expression in 10 different brain regions in 134 samples from the UK Brain Expression Cohort (UKBEC). Boxplot dashed bars mark the 25th and 75th percentiles. CRBL, cerebellar cortex; FCTX, frontal cortex; HIPP, hippocampus; MEDU, medulla (specifically inferior olivary nucleus); OCTX, occipital cortex (specifically primary visual cortex); PUTM, putamen; SNIG, substantia nigra; TCTX, temporal cortex; THAL, thalamus; WHMT, intralobular white matter.
Validation of MICALL2 function in zebrafish
Two homologous genes of MICALL2 are present in zebrafish; one of these genes, MICALL2b, is mainly expressed in the nervous system. After we injected micall2b-MOs into embryos at 3 d.p.f., MICALL2b gene expression was decreased compared with that in the MIS-control group (Figure 3b). Further behavioral testing showed that micall2b morphants displayed a marked increase in total swimming distance over 15 min (1343.00±87.73 mm in MO vs 875.60±76.44 mm in MIS, P=0.0001) (Figure 3c), and a higher mean swimming speed (2.716±0.1038 mm s−1 in MO vs 2.325±0.1093 mm s−1 in MIS, P=0.01) (Figure 3d). Furthermore, after treating the larvae with different concentrations of tomoxetine (TX) solution, which is a clinical medication for ADHD, fishes showed significantly decreased total swimming distance (Figure 3c) (MO E2 vs MO 5 μM TX, MO 10 μM TX, MO 15 μM TX, MO 20 μM TX: 1343±87.73 vs 717.9±97.89, 590.8±80.31, 479.7±70.01 and 506.9±73.41 mm, respectively; P<0.0001) and average velocity (Figure 3d) (MO E2 vs MO 5 μM TX, MO 10 μM TX, MO 15 μM TX, MO 20 μM TX: 2.716±0.1038 vs 1.802±0.1817, 1.529±0.1676, 1.182±0.2572, 1.106±0.1307 mm s−1, respectively; P<0.0001) compared with larvae only incubated in E2. A negative dose–response relationship was observed between the tomoxetine concentrations and behavioral performance.

Zebrafish with decreased micall2b expression showed hyperactive-impulsive behavior, and locomotion activity could be rescued by treatment with a attention deficit hyperactivity disorder (ADHD) drug. (a) Micall2b splice-blocking morpholino oligonucleotide (Micall2b-MO) was designed to block splicing at the exon 2/intron 2 boundary. (b) Micall2b expression in MO-injected zebrafish was lower than in mismatch MO (MIS)-injected control or wild-type (WT) zebrafish at 3 days post fertilization (d.p.f.). (c) The swimming distance of zebrafish injected with MO (n=64) was greater than that of MIS-injected animals (n=62) in 15 min at 6 d.p.f. Locomotion was reduced by treatment with tomoxetine at 5 μM (n=28), 10 μM (n=26), 15 μM (n=28) and 20 μM (n=26). (d) The average speed of larvae injected with MO (n=64) was faster than that of larvae injected with MIS (n=64). Changes in speed were rescued by treatment with tomoxetine at 5 μM (n=29), 10 μM (n=28), 15 μM (n=27) and 20 μM (n=26). The P-value was calculated using an unpaired t-test; error bars are±s.e.m. ***P<0.0005 and *P<0.05.
Discussion
In this study, a GWAS was conducted to explore genes associated with impaired executive inhibition in children with ADHD. One new significant locus, rs11514810, near the gene MICALL2 was identified to be associated with the word interference time in the Stroop test. Regulatory feature analysis for this locus showed that this region contained several transcription factor peaks and active regulatory elements, which may further influence the expression of nearby genes. eQTL data in the brain verified the effect of SNP rs11514810 on MICALL2 expression. Our animal experiment further validated the contribution of MICALL2 to the inhibitory phenotype in zebrafish. Decreased MICALL2 expression led to hyperactive-impulsive-like behavior, which could be rescued by treatment with an ADHD drug.
Mical represents a conserved family of cytosolic multidomain proteins. According to research on Mical family genes in flies, Mical genes are mainly expressed in the nervous system and the musculature and have roles in axon guidance, myofilament organization and synaptogenesis.47, 51 The connection of MICALL2 with several other genes associated with psychiatric disorders (Supplementary Figure 4) indicates that MICALL2 might exert important effects on the development of common phenotypic features in psychiatric disorders, including ADHD, through interactions with other genes. For example, MICALL2 could regulate neurite outgrowth by binding to Rab13,52 which is encoded by RAB13, and showed increased mRNA expression during neuronal regeneration. Additionally, MICALL2 could mediate the endocytic recycling of occludin in the tight junctions of MTD-1A cells53, 54, 55 and the migration of transfer Actinin-4 (encoded by ACTN4) from the cell body to the tips of neurites52 through binding to Rab13, which mediates the Dysbindin-1–necdin–p53 pathway in schizophrenia.56
Inhibition dysfunction is a key deficit of ADHD. By comparing the inhibitory function of a subset of patients recruited for this study with a control group, we revealed a consistent inhibitory deficit of ADHD patients in our previous study.57 Further evidence from literature suggested inhibitory deficit was heritable58 and aggregated in the family members of individuals with ADHD.59 Family segregation was also supported by Rommelse et al.60 in a large family-based study. In McAuley’s study,15 the difference of response inhibition between ADHD and controls remained significant in adolescence, which was independent of remittent or persistent of the disorder, suggesting state independence for inhibitory deficit. All the above evidence supported inhibition dysfunction as an endophenotype of ADHD. In this study, we identified the association of rs11514810 with inhibitory function in the ADHD sample. By the mediation analysis, we further found that, although rs11514810 was not associated with ADHD symptoms, it accounts for indirect effect on ADHD hyperactivity-impulsivity symptom through the effect of the SNP on inhibition. However, as we did not include a control group in the association analysis, we cannot conclude that the gene variant identified caused inhibitory dysfunction in ADHD. This variant may have also been associated with individual differences in inhibitory control in healthy individuals, which necessitated further elucidation in a comparable control group. However, given the important role of executive inhibition in research on psychiatric disorders, this study, as the first significant GWAS of behavioral-cognitive phenotypes, provided important information about the genetic mechanism of inhibitory function.
In summary, in this study, we used two independent sample groups to identify and verify the significant variant near MICALL2 associated with executive inhibition, as measured using the Stroop test. Within the same sample used for the GWAS of categorical ADHD,7 the dimensional neuropsychological endophenotype appeared to be more sensitive than the behavioral phenotype for discovering significant association signals. Further analysis of expression regulatory features and studies in an animal model helped to reveal the possible causal mechanism at the significant locus. The results provide more insight into the pathophysiology of ADHD through the impairment of executive inhibition, again highlighting neuronal development in the pathogenic process. Considering the complexity of cognition and behavior, it is likely that more genes remain to be discovered in larger samples and that more aspects of executive function should be discussed.
References
Polanczyk G, de Lima MS, Horta BL, Biederman J, Rohde LA . The worldwide prevalence of ADHD: a systematic review and metaregression analysis. Am J Psychiatry 2007; 164: 942–948.
Faraone SV, Perlis RH, Doyle AE, Smoller JW, Goralnick JJ, Holmgren MA et al. Molecular genetics of attention-deficit/hyperactivity disorder. Biol Psychiatry 2005; 57: 1313–1323.
Gizer IR, Ficks C, Waldman ID . Candidate gene studies of ADHD: a meta-analytic review. Hum Genet 2009; 126: 51–90.
Li D, Sham PC, Owen MJ, He L . Meta-analysis shows significant association between dopamine system genes and attention deficit hyperactivity disorder (ADHD). Hum Mol Genet 2006; 15: 2276–2284.
Smoller JW, Biederman J, Arbeitman L, Doyle AE, Fagerness J, Perlis RH et al. Association between the 5HT1B receptor gene (HTR1B) and the inattentive subtype of ADHD. Biol Psychiatry 2006; 59: 460–467.
Kuntsi J, Neale BM, Chen W, Faraone SV, Asherson P . The IMAGE project: methodological issues for the molecular genetic analysis of ADHD. Behav Brain Funct 2006; 2: 27.
Yang L, Neale BM, Liu L, Lee SH, Wray NR, Ji N et al. Polygenic transmission and complex neuro developmental network for attention deficit hyperactivity disorder: genome-wide association study of both common and rare variants. Am J Med Genet B 2013; 162B: 419–430.
Neale BM, Medland SE, Ripke S, Asherson P, Franke B, Lesch K-P et al. Meta-analysis of genome-wide association studies of attention-deficit/hyperactivity disorder. J Am Acad Child Adolesc Psychiatry 2010; 49: 884–897.
Hinney A, Scherag A, Jarick I, Albayrak O, Putter C, Pechlivanis S et al. Genome-wide association study in German patients with attention deficit/hyperactivity disorder. Am J Med Genet B 2011; 156B: 888–897.
Stergiakouli E, Hamshere M, Holmans P, Langley K, Zaharieva I, de CG et al. Investigating the contribution of common genetic variants to the risk and pathogenesis of ADHD. Am J Psychiatry 2012; 169: 186–194.
Bidwell LC, Willcutt EG, Defries JC, Pennington BF . Testing for neuropsychological endophenotypes in siblings discordant for attention-deficit/hyperactivity disorder. Biol Psychiatry 2007; 62: 991–998.
Irizarry RA, Hobbs B, Collin F, Beazer-Barclay YD, Antonellis KJ, Scherf U et al. Exploration, normalization, and summaries of high density oligonucleotide array probe level data. Biostatistics 2003; 4: 249–264.
Gao L, Li Z, Chang S, Wang J . Association of interleukin-10 polymorphisms with schizophrenia: a meta-analysis. PLoS ONE 2014; 9: e90407.
Nikolas MA, Nigg JT . Moderators of neuropsychological mechanism in attention-deficit hyperactivity disorder. J Abnorm Child Psychol 2015; 43: 271–281.
McAuley T, Crosbie J, Charach A, Schachar R . The persistence of cognitive deficits in remitted and unremitted ADHD: a case for the state-independence of response inhibition. J Child Psychol Psychiatry 2014; 55: 292–300.
Pennington BF, Ozonoff S . Executive functions and developmental psychopathology. J Child Psychol Psychiatry 1996; 37: 51–87.
Barkley RA . Attention-deficit/hyperactivity disorder, self-regulation, and time: toward a more comprehensive theory. J Dev Behav Pediatr 1997; 18: 271–279.
Smith EE, Jonides J . Storage and executive processes in the frontal lobes. Science 1999; 283: 1657–1661.
Miyake A, Friedman NP, Emerson MJ, Witzki AH, Howerter A, Wager TD . The unity and diversity of executive functions and their contributions to complex 'Frontal Lobe' tasks: a latent variable analysis. Cogn Psychol 2000; 41: 49–100.
Pineda D, Ardila A, Rosselli M, Cadavid C, Mancheno S, Mejia S . Executive dysfunctions in children with attention deficit hyperactivity disorder. Int J Neurosci 1998; 96: 177–196.
Barkley R . Attention-Deficit Hyperactivity Disorder (Third Edition): A Handbook for Diagnosis and Treatment. The Guilford Press: New York, NY, USA, 2006.
Barkley RA . Behavioral inhibition, sustained attention, and executive functions: constructing a unifying theory of ADHD. Psychol Bull 1997; 121: 65–94.
Nigg JT, Blaskey LG, Stawicki JA, Sachek J . Evaluating the endophenotype model of ADHD neuropsychological deficit: results for parents and siblings of children with ADHD combined and inattentive subtypes. J Abnorm Psychol 2004; 113: 614–625.
Barnett JH, Jones PB, Robbins TW, Muller U . Effects of the catechol-O-methyltransferase Val158Met polymorphism on executive function: a meta-analysis of the Wisconsin Card Sort Test in schizophrenia and healthy controls. Mol Psychiatry 2007; 12: 502–509.
Mione V, Canterini S, Brunamonti E, Pani P, Donno F, Fiorenza MT et al. Both the COMT Val158Met single-nucleotide polymorphism and sex-dependent differences influence response inhibition. Front Behav Neurosci 2015; 9: 127.
van Goozen SH, Langley K, Northover C, Hubble K, Rubia K, Schepman K et al. Identifying mechanisms that underlie links between COMT genotype and aggression in male adolescents with ADHD. J Child Psychol Psychiatry 2016; 57: 472–480.
Thissen AJ, Bralten J, Rommelse NN, Arias-Vasquez A, Greven CU, Heslenfeld D et al. The role of age in association analyses of ADHD and related neurocognitive functioning: a proof of concept for dopaminergic and serotonergic genes. Am J Med Genet B 2015; 168: 471–479.
van Rooij D, Hartman CA, van Donkelaar MM, Bralten J, von Rhein D, Hakobjan M et al. Variation in serotonin neurotransmission genes affects neural activation during response inhibition in adolescents and young adults with ADHD and healthy controls. World J Biol Psychiatry 2015; 16: 625–634.
Richards JS, Arias Vasquez A, van Rooij D, van der Meer D, Franke B, Hoekstra PJ et al. Testing differential susceptibility: plasticity genes, the social environment, and their interplay in adolescent response inhibition. World J Biol Psychiatry advance online publication, 12 May 2016, 1–14; doi: 10.3109/15622975.2016.1173724.
Cummins TD, Jacoby O, Hawi Z, Nandam LS, Byrne MA, Kim BN et al. Alpha-2A adrenergic receptor gene variants are associated with increased intra-individual variability in response time. Mol Psychiatry 2014; 19: 1031–1036.
Yang L, Wang YF, Qian QJ, Biederman J, Faraone SV . DSM-IV subtypes of ADHD in a Chinese outpatient sample. J Am Acad Child Adolesc Psychiatry 2004; 43: 248–250.
Patterson N, Price AL, Reich D . Population structure and eigenanalysis. PLoS Genet 2006; 2: e190.
Price AL, Patterson NJ, Plenge RM, Weinblatt ME, Shadick NA, Reich D . Principal components analysis corrects for stratification in genome-wide association studies. Nat Genet 2006; 38: 904–909.
Li Y, Zhang K, Chen H, Sun F, Xu J, Wu Z et al. A genome-wide association study in Han Chinese identifies a susceptibility locus for primary Sjogren's syndrome at 7q11.23. Nat Genet 2013; 45: 1361–1365.
Purcell S, Neale B, Todd-Brown K, Thomas L, Ferreira MA, Bender D et al. PLINK: a tool set for whole-genome association and population-based linkage analyses. Am J Hum Genet 2007; 81: 559–575.
Liu EY, Li M, Wang W, Li Y . MaCH-admix: genotype imputation for admixed populations. Genet Epidemiol 2013; 37: 25–37.
Genomes Project C Genomes Project C Abecasis GR, Genomes Project C Auton A, Genomes Project C Brooks LD, Genomes Project C DePristo MA, Genomes Project C Durbin RM et al. An integrated map of genetic variation from 1,092 human genomes. Nature 2012; 491: 56–65.
Li Y, Willer C, Sanna S, Abecasis G . Genotype imputation. Annu Rev Genomics Hum Genet 2009; 10: 387–406.
Guo L, Du Y, Qu S, Wang J . rVarBase: an updated database for regulatory features of human variants. Nucleic Acids Res 2016; 44: D888–893.
Gerstein MB, Kundaje A, Hariharan M, Landt SG, Yan KK, Cheng C et al. Architecture of the human regulatory network derived from ENCODE data. Nature 2012; 489: 91–100.
Trabzuni D, Ryten M, Walker R, Smith C, Imran S, Ramasamy A et al. Quality control parameters on a large dataset of regionally dissected human control brains for whole genome expression studies. J Neurochem 2011; 119: 275–282.
Ramasamy A, Trabzuni D, Guelfi S, Varghese V, Smith C, Walker R et al. Genetic variability in the regulation of gene expression in ten regions of the human brain. Nat Neurosci 2014; 17: 1418–1428.
Cowley MJ, Pinese M, Kassahn KS, Waddell N, Pearson JV, Grimmond SM et al. PINA v2.0: mining interactome modules. Nucleic Acids Res 2012; 40: D862–D865.
Lage K, Karlberg EO, Storling ZM, Olason PI, Pedersen AG, Rigina O et al. A human phenome-interactome network of protein complexes implicated in genetic disorders. Nat Biotechnol 2007; 25: 309–316.
Warde-Farley D, Donaldson SL, Comes O, Zuberi K, Badrawi R, Chao P et al. The GeneMANIA prediction server: biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res 2010; 38: W214–W220.
Hayes AF . Introduction to Mediation, Moderation, and Conditional Process Analysis: A Regression-Based Approach. Guilford Press: New York, USA, 2013.
Terman JR, Mao T, Pasterkamp RJ, Yu HH, Kolodkin AL . MICALs a family of conserved flavoprotein oxidoreductases, function in plexin-mediated axonal repulsion. Cell 2002; 109: 887–900.
Zhang L, Chang S, Li Z, Zhang K, Du Y, Ott J et al. ADHD gene: a genetic database for attention deficit hyperactivity disorder. Nucleic Acids Res 2012; 40: D1003–D1009.
Jia P, Sun J, Guo AY, Zhao Z . SZGR: a comprehensive schizophrenia gene resource. Mol Psychiatry 2010; 15: 453–462.
Guo L, Zhang W, Chang S, Zhang L, Ott J, Wang J . MK4MDD: a multi-level knowledge base and analysis platform for major depressive disorder. PLoS ONE 2012; 7: e46335.
Beuchle D, Schwarz H, Langegger M, Koch I, Aberle H . Drosophila MICAL regulates myofilament organization and synaptic structure. Mech Dev 2007; 124: 390–406.
Sakane A, Honda K, Sasaki T . Rab13 regulates neurite outgrowth in PC12 cells through its effector protein, JRAB/MICAL-L2. Mol Cell Biol 2010; 30: 1077–1087.
Terai T, Nishimura N, Kanda I, Yasui N, Sasaki T . JRAB/MICAL-L2 is a junctional Rab13-binding protein mediating the endocytic recycling of occludin. Mol Biol Cell 2006; 17: 2465–2475.
Nishimura N, Sasaki T . Cell-surface biotinylation to study endocytosis and recycling of occludin. Methods Mol Biol 2008; 440: 89–96.
Nishimura N, Sasaki T . Identification and characterization of JRAB/MICAL-L2, a junctional Rab13-binding protein. Methods Enzymol 2008; 438: 141–153.
Sjoholt G, Gulbrandsen AK, Lovlie R, Berle JO, Molven A, Steen VM . A human myo-inositol monophosphatase gene (IMPA2) localized in a putative susceptibility region for bipolar disorder on chromosome 18p11.2: genomic structure and polymorphism screening in manic-depressive patients. Mol Psychiatry 2000; 5: 172–180.
Shuai L, Chan RC, Wang Y . Executive function profile of Chinese boys with attention-deficit hyperactivity disorder: different subtypes and comorbidity. Archiv Clin Neuropsychol 2011; 26: 120–132.
Schachar RJ, Forget-Dubois N, Dionne G, Boivin M, Robaey P . Heritability of response inhibition in children. J Int Neuropsychol Soc 2011; 17: 238–247.
Schachar RJ, Crosbie J, Barr CL, Ornstein TJ, Kennedy J, Malone M et al. Inhibition of motor responses in siblings concordant and discordant for attention deficit hyperactivity disorder. Am J Psychiatry 2005; 162: 1076–1082.
Rommelse NN, Altink ME, Oosterlaan J, Buschgens CJ, Buitelaar J, Sergeant JA . Support for an independent familial segregation of executive and intelligence endophenotypes in ADHD families. Psychol Med 2008; 38: 1595–1606.
Acknowledgements
We thank all the patients for participating in this study. This work was supported by grants from the Major State Basic Research Development Program of China (973 Program, 2014CB846100 to YW), the National Natural Science Foundation of China (31401139 to SC, 81471381 to LY, and 31571496 to DL), Beijing Municipal Science and Technology Commission (Z151100003915122), and National Institutes of Health (R01HL129132 and R01HG006292 to YL), the Medical Research Council Grant ‘c-VEDA’ (Consortium on Vulnerability to Externalizing Disorders and Addictions) (MR/N000390/1), the Horizon 2020 funded ERC Advanced Grant ‘STRATIFY’ (695313).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies the paper on the Molecular Psychiatry website
Supplementary information
Rights and permissions
About this article

Cite this article
Yang, L., Chang, S., Lu, Q. et al. A new locus regulating MICALL2 expression was identified for association with executive inhibition in children with attention deficit hyperactivity disorder. Mol Psychiatry 23, 1014–1020 (2018). https://doi.org/10.1038/mp.2017.74
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/mp.2017.74
- Springer Nature Limited
This article is cited by
-
Shared polygenic risk for ADHD, executive dysfunction and other psychiatric disorders
Translational Psychiatry (2020)
-
CK1δ over-expressing mice display ADHD-like behaviors, frontostriatal neuronal abnormalities and altered expressions of ADHD-candidate genes
Molecular Psychiatry (2020)
-
Genetic variant for behavioral regulation factor of executive function and its possible brain mechanism in attention deficit hyperactivity disorder
Scientific Reports (2018)