
Abstract
Ammonites belonging to the genus Latiwitchellia Imlay are described and figured for the first time outside the palaeogeographic domain of the Eastern Pacific. The Subbetic ammonites that are classified here as Latiwitchellia evoluta Imlay fit quite well with the North American forms (from Snowshoe Formation, Oregon) despite being slightly less evolute and having more regular ribbing. The range of stratigraphic distribution is similar, although the oldest Subbetic forms (lower part of the Discites Zone) are somewhat earlier than the North American ones (uppermost Discites Zone). The presence of the same species in the Betic Cordillera (westernmost Tethys) and in eastern Oregon (Eastern Pacific) confirms that during the early Bajocian the two palaeogeographic domains were connected via the Hispanic Corridor. Latiwitchellia possibly originated in the westernmost Tethys before expanding to the Eastern Pacific.
Resumen
Se describen y figuran por primera vez ammonites pertenecientes al género Latiwitchellia Imlay fuera del dominio paleogeográfico del Pacífico Oriental. Las ammonites subbéticos que se clasifican como Latiwitchellia evoluta Imlay encajan bastante bien con las formas norteamericanas (de la Formación Snowshoe, Oregón), aunque son un poco menos evolutos y tienen la ornamentación más regular. El rango de distribución estratigráfica es similar, aunque las formas subbéticas más antiguas (parte inferior de la Zona de Discites) son algo más antiguas que las norteamericanas (parte superior de la Zona de Discites). La presencia de la misma especie en la Cordillera Bética (extremo occidental del Tetis) y en el este de Oregón (Pacífico Oriental) confirma que durante el Bajociense temprano los dos dominios paleogeográficos estaban conectados a través del Corredor Hispánico. Latiwitchellia posiblemente se originó en el Tetis occidental antes de expandirse al Pacífico oriental.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
The genus Latiwitchellia was established and described by Imlay (1973, p. 70) to include about 52 ammonite specimens of the Lower Bajocian from the upper part of the Weberg Member, in the Snowshoe Formation, Eastern Oregon (USA). Subsequently, only a small number of citations are known for this genus (Donovan et al., 1981, p. 143; Fernández-López, 1985, p. 68; Taylor, 1988, p. 24, 2016, p. 459; Smith & Taylor, 1992, p. 72; Westermann, 1992, p. 257; Sandoval et al., 2012; Sandoval, 2022) that refer to the North American specimens originally described by Imlay (1973), but no citations of new species are known, except an Andean form ("L”. atacamensis Dietze and Hillebrandt, 2019, p. 44), which, only with certain doubts, should be included in this genus. Imlay (1980, p. 22) refers to Latiwitchellia gracilis, but without indicating the author of the species or any description or illustration of it.
The detailed sampling carried out since the 1970s in the Lower Bajocian of various stratigraphic sections of the Median Subbetic in the central sector of the Betic Cordillera have provided a large quantity of ammonoids, many of which have been taxonomically analysed, enabling fine biostratigraphic results for this stratigraphic interval (Sandoval, 1983, 1990, 2022 and references therein). Only a few of these ammonites (eight in total) have been attributed to the genus Latiwitchellia Imlay.
2 Geographical and geological setting
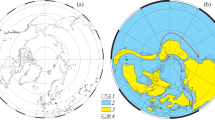
The geographical origin of the Latiwitchellia specimens analysed here (Fig. 1) formed part of the southern margin of the Iberian Plate, which, during the Bajocian (Middle Jurassic) was located near the eastern end of the Hispanic Corridor (Fig. 1b). This corridor was a relatively narrow sea channel that during certain time intervals (i.e. Early Bajocian–earliest Late Bajocian and Late Bathonian–Early Callovian) connected the Western Tethys with the Eastern Pacific (Sandoval, 2022 and references therein). This area was located at a palaeolatitude of about 26°N (Fig. 1c) within the southern Iberian continental margin by the Early Bajocian (~170 Ma). These sites are currently located in southern Spain (Fig. 1a), being part of the External Zones of the Betic Cordillera.

a General sketch of the Betic Cordillera with approximate location of the areas with the sections containing Latiwitchellia: 1 Sierra de Alta Coloma (JAC3 and JAC11 sections); 2 Barranco de Agua Larga (JAQ1 and JAQ2 sections), Jaén Province; 3 Rio Fardes (JFB2 section), Granada Province. b Palaeogeographic map (Mollweide projection) showing the distribution of oceans and lands in the Early Bajocian (c. 170 Ma); stars indicate the approximate areas where Latiwitchellia has been recorded. c, Focus on the Western Tethys-Central Atlantic showing the approximate location of the Subbetic basin (rectangle) in relation to the trans-Pangaean seaway; the black triangles indicate the approximate position of the palaeodomains where Latiwitchellia has been recorded. Modified after Sandoval (2022).
3 Material and methods
As mentioned above, the specimens of the genus Latiwitchellia studied here come from selected stratigraphic sections of the Subbetic domain (Betic Cordillera, southern Spain). Whenever possible, these sections were numbered, measured, and sampled in detail, bed by bed. Localities and stratigraphic sections were abbreviated to a letter-number code following the customary rules of the Department of Stratigraphy and Palaeontology (University of Granada, Spain). Among the numerous stratigraphic sections sampled, the only sites where Latiwitchellia were collected are (Fig. 1a): Sierra de Alta Coloma area and Barranco de Agua Larga, Campillo de Arenas and Noalejo (Jaén Province); Rio Fardes, Gorafe (Granada Province).
After being prepared and cleaned in the laboratory, the ammonites were coated with magnesium oxide and then photographed.
The present work follows the standard ammonite zonation for the Mediterranean Province (Rioult et al., 1997), but raising the Ovale Subzone to zonal rank. The Propinquans Zone is equivalent to the Sauzei Zone used by German and some British researchers.
3.1 Abbreviations
3.1.1 Measurements
The most significant specimens were measured, for which the following abbreviations are used: D, diameter of the shell (mm); U, umbilical diameter of the shell (mm); H, whorl-section height of the shell (mm); W, whorl-section width of the shell (mm); h = H/D; u = U/D; w = W/D. Some of these parameters are approximate, because some of the specimens were laterally flattened (crushed) by compression.
3.1.2 Other abbreviations
PH, phragmocone; BC, body chamber; E external lobe; L, lateral lobe; U2-U5, umbilical lobes; HT, holotype; OD, original designation; [M], macroconchs; [m], microconchs.
3.1.3 Housing institution
The specimens studied are on deposit in the “Departamento de Estratigrafía y Paleontología”, University of Granada, Spain (UGR). Only representative specimens are figured.
4 Systematic palaeontology
Order AMMONITIDA Fischer, 1882
Suborder AMMONITINA Fischer, 1882
Superfamily HILDOCERATACEOIDEA Hyatt, 1867
Family HILDOCERATIDAE Hyatt, 1867
Subfamily GRAMMOCERATINAE Buckman, 1905
4.1 Genus LATIWITCHELLIA Imlay, 1973
Type species by OD. Witchellia (Latiwitchellia) evoluta Imlay, 1973, p. 70, pl. 32, Figs. 1, 2, 5, 6; HT.
4.1.1 Description (emended after Imlay, 1973)
Shells evolute with slow growth of the whorl, ovate or subquadrate-compressed to moderately compressed whorl-section, flat or slightly convex flanks, venter gently rounded to almost flat with a rather high keel limited by smooth flat bands or bordered by shallow furrows. Rectiradiate to rursiradiate ribs, mainly simple, some bifurcate on the middle third of the flanks, and rarely arise freely on the flanks; ribs bend sharply forward high on the flanks, vary considerably in strength from one specimen to another, and can vary during ontogeny; small nodes can develop on the upper part of flanks at the points where ribs bend forward, these occurring mostly on the intermediate and outer whorls of coarsely ribbed specimens. The aperture is marked by a deep constriction that is followed by a swelling. The septal suture is very simple with little ramification: the E-L saddle is extremely broad and irregularly rounded; the lateral lobe (L) is not deep, trifid, moderate in width, about the same depth as the E (external lobe), and considerably deeper than the umbilical (U2); the suspensive lobe rises slightly to the umbilical seam. Although forms of different sizes appear among the specimens figured by Imlay (1973, pls 31–33), the existence of dimorphism is not shown. All of these specimens are apparently macroconchs. However, some of the specimens included in Pelekodites by Imlay (1973, p. 74, pl. 34, Figs. 1–18) could represent the dimorphous partner [m] of Latiwitchellia [M].
4.1.2 Remarks
According to Imlay (1973, p. 70), who considered Latiwitchellia to be a subgenus of Witchellia, and consequently included it in the family Sonniniidae Buckman, 1892, this differs from Witchellia Buckman (1889) in having more evolute coiling, a relatively broader whorl section that does not become higher or compressed on the BC, less common ribs furcation near the base of the flanks, only slight weakening of its ribs adorally on the BC, and blunt nodes high on the flanks on some coarsely ribbed variants.
Donovan et al., (1981, p. 143) considered Latiwitchellia to be synonymous with Fontannesia and included it in the Family Sonniniidae.
Fernández-López (1985, p. 68) highlighted the significant differences between Witchellia and Latiwitchellia (coiling, whorl-section, ventral region, ribbing) and concluded that these morphological differences, in addition to their biostratigraphic distribution, bring Latiwitchellia closer to the forms of the genus Fontannesia than to Witchellia.
Fontannesia Buckman, 1902, p. 6 (type species, Dumortieria grammoceroides Haug, 1887, p. 137, pl. 5, Fig. 5; OD), is quite similar to Latiwitchellia, but it is more involute with greater whorl spiral expansion, has an ovate rather than subrectangular whorl-section and, the ribs, generally more uniform and persistent throughout ontogeny, are rarely bifurcate or twinned at the umbilical edge. Since the establishment of this taxon by Buckman, many authors, including Howarth (2013 in the new Treatise, p.119, who indicated that it is a Bajocian homeomorph of Grammoceras), have included Fontannesia in the family Sonniniidae. However, others, such as Westermann and Getty (1970, p., 240), Westermann & Wang (1988, p. 314), Linares and Sandoval (1988), and Sandoval et al. (2012) and Sandoval (2022), showed that Fontannesia should be included in the Grammoceratinae Buckman, 1905, with which it shares more common characters. The specimens described and figured by Imlay (1973, pp. 58–59; pl. 5, Figs. 1–19) as Fontannesia cf. F. luculenta (Buckman), F. cf. F. carinata (Buckman) and F. cf. F. evoluta (Buckman) show considerable resemblance to the various morphotypes of the type species of Latiwitchellia and should possibly be included in this genus.
Linaresites Sandoval, 2012 (in Sandoval et al., 2012, p. 115; type species, Fontannesia montillanensis Linares & Sandoval, 1988, p. 8, pl. 1, Figs. 12, 13 (OD)) is perhaps the genus most closely related to Latiwitchellia. The two genera share a markedly evolute coiling, the same type of whorl-section, keel, and similar shape as well as thickness of ribbing and very simple sutures, but Linaresites is much smaller and appears earlier in the stratigraphic record.
Dietze & Hillebrandt (2019, p. 40) regarded (as Imlay, 1973) Latiwitchellia as being closely related to the genus Witchellia, including this evolute member in the subfamily Witchelliinae Callomon & Chandler, 2006 in Chandler et al., (2006, p. 370). They separated Latiwitchellia from the evolutionary line Vacekia- Asthenoceras–Linaresites, as Sandoval et al., (2012, p. 117, Fig. 3) had proposed, because of the much smaller size and the thin and notably high keel of these Grammoceratinae. However, as can be seen in some of the specimens described and figured by Imlay (1973, pls 32 and 33), these also have a quite thin and high keel, while the various specimens of Linaresites (see Sandoval et al., 2012, Figs. 2L–O), a genus with clear phylogenetic relationships to Asthenoceras and Latiwitchellia, do not have a tall, thin keel.
Linaresites and Latiwitchellia (both being overlooked by Howarth, 2013 in the new version of the Treatise, and Linaresites also being overlooked by Dietze and Hillebrandt, 2019) show clear phylogenetic relationships with Asthenoceras Buckman, 1899, of which the former two genera are possibly close descendants (Sandoval et al., 2012, p. 117, Fig. 3; Sandoval, 2022, p. 809). According to Sandoval et al., (2012, p. 117), Latiwitchellia, which has the oldest records in the lowermost Bajocian (Discites Zone, Walkeri Subzone), appears to be the latest representative of the subfamily Grammoceratinae, rather than a primitive Witchelliinae. L. evoluta is possibly a neotenic form of L. montillanensis, and it may represent the last Grammoceratinae. The taxonomic status and phylogenetic relationships of Fontannesia with other taxa are less clear, possibly descending from some of the more strongly ribbed Asthenoceras (Fig. 2). New analyses on more abundant and well-preserved material will be necessary to clarify such relationships.

Proposed phylogenetic relationships among the latest Grammoceratinae (calibrated against the Mediterranean standard zonation). The figured specimens (not the septal sutures) are the HTs of the type species of each genus. Scale bars 1 cm (modified after Sandoval et al., 2012)
4.1.3 Geographic and stratigraphic distribution
All the North American specimens (from the Weberg Member of the Snowshoe Formation) are from the Lower Bajocian (Tuberculatum Zone, Ochocoense Subzone, and lower part of the Burkei Zone), more or less equivalent to the upper part of the Discites Zone, the Ovale Zone, and the basal part of the Laeviuscula Zone of the Mediterranean standard zones (see Taylor, 2016, pp. 459–460, Figs. 7 and 8). “L.” atacamensis Dietze & Hillebrandt comes from Lower Bajocian (Giebeli Zone, Multiformis Subzone, equivalent to the lower part of the Sauzei Zone; see Dietze & Hillebrandt, 2019, p. 26, text-Fig. 2) of Portezuelo, El Padre Section, Manflas area, Copiapó Province, Atacama, Northern Chile. In the Subbetic, Latiwitchellia has been recorded in the Lower Bajocian (Discites and Ovale zones) from the Barranco de Agua Larga and Sierra de Alta Colomaa, Campillo de Arenas, and Noalejo (Jaén Province) and the Rio Fardes area, Gorafe (Granada Province).
4.1.4 Latiwitchellia evoluta Imlay, 1973
Fig. 3a–e

Latiwitchellia evoluta Imlay, 1973 [M]: a, b JAQ1.-32. 2, lateral and ventral views, Discites Zone, Subsectum Subzone, Barranco de Agua Larga (JAQ1 section); c, d JFB2.4.1, lateral and ventral views, Ovale Zone?, Río Fardes (JFB2 section), specimen figured in Hernández-Molina et al., 1991, Fig. 15.2; e JAC3.(14–20)0.1, lateral view, Ovale Zone, Sierra de Alta Coloma (JAC3 section). f Fontannesia sp., JRi2.1.1, Discites Zone, Walkeri Subzone, Sierra de Ricote (JRi2 section), Murcia province. Scale bars 1 cm
1973? Fontannesia cf. F. evoluta (Buckman). - Imlay, p. 59, pl. 4, Figs 1–3.
1973 Witchellia (Latiwitchellia) evoluta Imlay n. sp. - Imlay, p. 70, pls 31-33 (HT in pl. 32, Figs. 1–3, 5, 6).
1991 Fontannesia cf. grammoceroides (Haug). - Hernández-Molina et al., p. 95, Fig. 15, 2.
4.1.4.1 Material
JAC3.5.1, JAC3.14.2, JAC3.(14–20)0.1, JAC11.(3–7)0.1, JAQ1.(-32)0.2, JAQ1.(-16)0.2, JAQ2.20.23 and JFB2.4.1
4.1.4.2 Measurements
Specimen | D | U | u | H | h | W | w | R | Remarks | Biostratigraphy |
|---|---|---|---|---|---|---|---|---|---|---|
JAQ1.-32. 2 | 47.0 | 22.0 | 0.47 | 14.0 | 0.30 | 9.00 | 0.20 | 25 | [M], adult | Discites Zone |
43.0 | 20.0 | 0.47 | 12.7 | 0.30 | 8.7 | 0.20 | ||||
JAQ2.20.23 | 51.0 | 22.0 | 0.43 | 14.5 | 0.28 | [M], adult. Complete? | Discites Zone | |||
40.0 | 15.3 | 0.38 | 12.5 | 0.31 | ||||||
JAC3.(14–20)0.1 | 60.0 | 25.0 | 0.42 | 20.0 | 0.33 | [M], adult, complete? | Ovale Zone | |||
43.0 | 17.5 | 0.41 | 15.2 | 0.35 | ||||||
JAC11.3.7 | 46.0 | 19.5 | 0.42 | 15.5 | 0.34 | 10.0 | 0.22 | 18 | [M] | Ovale Zone |
36.5 | 14.0 | 0.38 | 13.3 | 0.36 | ||||||
JFB2.4.1 | 49.0 | 22.5 | 0.46 | 15.0 | 0.31 | 13.5 | 0.26 | 17 | [M] | Ovale? Zone |
37.0 | 17.0 | 0.46 | 12.0 | 0.32 |
4.1.4.3 Description
Medium-sized shells, evolute (O/D varying from 0.49 to 0.42), although somewhat less than for the specimens described by Imlay (1973, pp. 70–71), with slow whorl-spiral expansion. In specimens not laterally crushed, the whorl-section varies from subquadratic in internal whorls to suboval compressed in the BH; the oblique to rounded umbilical wall merges with the rounded umbilical edge; the flanks vary from almost flat to gently convex and converge to the venter, which is gently rounded to almost flat and bears a more or less high keel bordered by furrows sometimes replaced by flat bands in the BH of most evolutes and strongly ornamented specimens or even in earlier stages in less evolute and less strongly ribbed specimens. None of the Subbetic specimens seems to preserve the adult aperture, which in the North American forms inclines gently forward on the flanks and projects strongly forward on the venter. The ribbing, quite irregular, can vary considerably throughout ontogeny and from one specimen to another; ribs are mainly simple, some bifurcate on the lower third of the flanks and rarely a few arise freely near the middle flanks. Inner and intermediate whorls bear ribs that are dense, well marked, and nearly rectiradiate on the lower parts of the flanks; these become higher and generally rursiradiate on the middle parts, becoming still higher and projected forward high on the flanks and near the venter. On outer whorls, on the PH end, and on the BH the ribs become progressively more spaced, weaken or even vanish in the lower part of the flanks, while remaining very wide and strong in the upper part of the flank and in the venter region, where they project forward. Some specimens develop nodes or thickenings high on the flanks at the points where the flank ribs bend forward, this occurring mostly on the intermediate and outer whorls of coarsely ribbed forms (see JFB2.4.1 and JAQ1.-32.2). The less evolute specimens, stratigraphically higher up, may not have bifurcate or free intercalate ribs. The septal suture, partially observable in specimens JAQ1-32.2 and JAC11(3–7)0.1, is very simple with the E-L saddle and L-U2 broad and barely ramified; L is somewhat shallow, trifid, narrow, barely ramified, and somewhat deeper than the U2.
4.1.4.4 Remarks
To date, apart from Latiwitchellia evoluta (Imlay), only one other ammonite species (“L.” atacamensis Dietze & Hillebrandt, 2019, p. 44, pl. 11, Figs. 2, 4, HT by monotype) has been originally assigned to the genus Latiwitchellia. This Andean species is more involute and has the faintest ribbing on inner whorls and very regular ribbing in the BC; except for the ventral grooves, these features bring it closer to Fontannesia or even to Dorsetensia, than to Latiwitchellia.
The Subbetic specimens here analysed and classified as L. evoluta match quite well with several of the specimens figured by Imlay (1973, pls 31–33), mainly with the variants that are less coarsely ribbed or intermediate, although North American forms have a slightly wider umbilicus and a slightly stronger and irregular ribbing. The specimen classified by Imlay (1973, p. 59, pl. 4, Figs. 1–3) as Fontannesia cf. F. evoluta (Buckman) is similar to some of the morphotypes of L. evoluta and could fall within the variability range of this species. Some Fontannesia species bear quite resemblance to L. evoluta (see Fig. 3f), but, are more involute, have an ovate whorl-section and a more sigmoidal, more persistent and regular ribbing throughout ontogeny. The more involute Subbetic specimens (JAQ2.20.23, JAC3.(14–20)0.1, JAC11.3.7) show similarities with some specimens of Asthenoceras intermedium (Imlay, 1973, p. 57, pl. 4, Figs. 7–15), but this species, which precedes L. evoluta in the stratigraphic record, is more involute and has finer ribs, many of which are divided in the lower third of the flank. Likewise, the same specimens share some similarities with the specimens from Tibet described and figured as Fontannesia kiliani (Kruzinga) by Westermann & Wang (1988, p. 316, pl. 20, Figs. 1–4, 6–7), which are also more involute and have more irregular ribs, many of which are furcate. Linaresites montillanensis (Linares & Sandoval, 1988) shares morphological similarities with L. evoluta, mainly in the evolute coiling, whorl-section, type of ribs, and very simple suture; however, these forms, later in time, are substantially larger. L. evoluta, possibly a neotenic form of L. montillanensis, may represent the last Grammoceratinae.
4.1.4.5 Distribution
The Subbetic specimens are from Lower Bajocian, Discites Zone of the Barranco Agua Larga (sections JAQ1 and JAQ2), Noalejo (Jaén Province) and from Ovale Zone of sections of Barranco de la Jarropa (JAC3), La Torquilla (JAC11) in Sierra de Alta Coloma area, Campillo de Arenas, and Noalejo (Jaén Province), and Rio Fardes (JFB2), Gorafe (Granada Province).
5 Conclusions
-
(1)
The analysis of the abundant Lower Bajocian ammonites collected in several stratigraphic sections from Subbetic domain (Betic Cordillera, southern Spain) has revealed the presence of a few specimens belonging to Latiwitchellia, an ammonite genus that to date has been found only in the Eastern Pacific Domain (North American Western Cordillera, Oregon, USA) and Chile.
-
(2)
The Subbetic Latiwitchellia specimens show clear similarities with the North American forms and have been considered as variants (morphotypes) of a same species, L. evoluta Imlay.
-
(3)
The analysis of the taxonomic relationships among Latiwitchellia and other Middle Jurassic ammonite genera such as Asthenoceras, Linaresites, and Fontannesia reveals a close phylogenetic relationship between these four genera (Fig. 2), which have typical characters of the Grammoceratinae, especially ornamentation and suture. This definitively excludes Latiwitchellia from Witchelliinae (Sonniniidae), leading it to be included in Grammoceratinae (Hildoceratidae). The relationship among Asthenoceras, Linaresites, and Latiwitchellia seems clear, whereas the possible relationship with Fontannesia remains more doubtful, although it seems feasible (based on morphology as well as biostratigraphy) that both Linaresites and Fontannesia descended directly from Asthenoceras in the late Aalenian.
-
(4)
The biostratigraphic range of Latiwitchellia is quite similar in the Subbetic and North American Western Cordillera, although the oldest Subbetic forms (from lowermost Discites Zone) are somewhat earlier in time than the North American forms (uppermost Discites Zone). This implies that although Latiwitchellia is more abundant in the eastern Pacific domain, it could have originated in the western end of the Tethys.
-
(5)
The Hispanic Corridor connecting Westernmost Tethys and Eastern Pacific was the oceanic channel that, at least temporarily, allowed the migration of Latiwitchellia and other ammonite taxa between the two palaeogeographic domains.
References
Buckman, S. S. (1902). Emendations of ammonite nomenclature (p. 7). Privately published.
Buckman, S. S. (1905). A monograph of the Inferior Oolite ammonites of the British Islands, part 13. Monograph of the Palaeontographical Society, London. supplement p. clxix–ccviii, supplement pl. 20–24.
Buckman, S. S. (1887–1907). A Monograph of the Ammonites from the Inferior Oolite Series. Monograph of the Palaeontographical Society, 1 (1–6), 1–456 + cclxii p., 103 + 24 supplementary pls. https://doi.org/10.1017/CBO9781316257203.
Chandler, R. B., Callomon, J. H., King, A., Jeffreys, K., Varah, M., & Bentley, A. (2006). The stratigraphy of the Inferior Oolite at South Main Road Quarry, Dundry, Avon. Proceedings of the Geologists’ Association, 117, 345–375. https://doi.org/10.1016/S0016-7878(06)80043-X
Dietze, V., & Hillebrandt, A. V. (2019). Lower Bajocian (Middle Jurassic) Ammonites of the Manflas area in Atacama Province, Northern Chile, Part 2: Giebeli Zone. Zitteliana, 93, 25–46.
Donovan, D. T. Callomon, J. H. & Howarth, M. K. (1981). Classification of the Jurassic Ammonitina. Systematics Association, London, vol. 18, pp. 101–155. Academic Press, London.
Fernández-López, S. (1985). El Bajociense en la Cordillera Ibérica. PhD. Thesis, Departamento de Paleontología, Facultad de Ciencias Geológicas, Univ. Complutense, Madrid, 850 p.
Fischer, P. H. (1882). Manuel de conchyliologie et de paléontologie conchyliologique. Librairie F. Savy. Paris. 1369 p., 23 pl.
Haug, E. (1887). Über die “Polymorphitidae”, eine neue Ammonitenfamilie aus dem Lias. Neues Jahrbuch Für Mineralogie, Geologie und Paläontologie, 1887(2), 89–163.
Hernández-Molina, F. J., Sandoval, J., Aguado, R., O’dogherty, L., Comas, M. C., & Linares, A. (1991). Olistoliths from the Middle Jurassic in Cretaceous materials of the Fardes Formation: Biostratigraphy (Subbetic Zone, Betic Cordillera). Revista De La Sociedad Geológica De España, 4, 79–104.
Howarth, M. K. (2013). Treatise Online No. 57, Part L, Revised, Volume 3B, Chapter 4: Psiloceratoidea, Eodoceratoidea, Hildoceratoidea. The University of Kansas, Lawrence, Kansas, 139 p. https://doi.org/10.17161/to.v0i0.4441
Hyatt, A. (1867). The fossil Cephalopoda of the Museum of Comparative Zoology. Bulletin of the Museum of Comparative Zoology, 5, 71–102.
Imlay, R. W. (1973). Middle Jurassic (Bajocian) Ammonites from Eastern Oregon. Professional Paper of the U.S. Geological Survey, 756, 1–100. https://doi.org/10.3133/pp756
Imlay, R. W. (1980). Jurassic paleobiogeography of the conterminous United States in its continental setting. Professional Paper of the U.S. Geological Survey, 1062, 1–134. https://doi.org/10.3133/pp1062
Linares, A. & Sandoval, J. (1988). Asthenoceras y Fontannesia (Grammoceratinae?, Ammonitina) del tránsito Aaleniense-Bajociense de la Zona Subbética (Sur de España). Revista Española de Paleontología, 3, 3–11.
Rioult, M., Contini, D., Elmi, S. & Gabilly, J. (1997). Bajocien, in Cariou E. & Hantzpergue P. (Eds.), Biostratigraphie du Jurassique ouest-européen et méditerranéen. Bulletin des Centres de Recherches Exploration-Production Elf-Aquitaine, 17, 41–53.
Sandoval, J. (1990). A revision of the Bajocian divisions in the Subbetic Domain (southern Spain). Memorie Descrittive Della Carta Geologica D’italia, 40, 141–162.
Sandoval, J. (2022). Sonniniidae (Ammonitina, Middle Jurassic) from Southern Spain: taxonomic biostratigraphical and palaeobiogeogral analysis. Geodiversitas, 44(27), 801–851. https://doi.org/10.5252/geodiversitas2022v44a27
Sandoval, J., Henriques, M. H., Chandler, R. B., & Ureta, M. (2012). Latest Toarcian-earliest Bajocian (Jurassic) Grammoceratinae (Hildoceratidae, Ammonitina) of the western Tethys: their palaeobiogeographic and phylogenetic significance. Geobios, 45, 109–219. https://doi.org/10.1016/j.geobios.2011.11.001
Sandoval, J. (1983). Bioestratigrafía y Paleontología (Stephanocerataceae y Perisphinctaceae) del Bajocense y Bathonense de las Cordilleras Béticas. PhD Thesis. Secretariado de Publicaciones, Universidad de Granada, Granada, 613 pp.
Smith, P., & Taylor, D. G. (1992). Eastern Oregon and adjacent areas. In G. E. G. Westermann (Ed.), The Jurassic of the Circum-Pacific (pp. 67–72). Cambridge University Press.
Taylor, D. G. (1988). Middle Jurassic (Late Aalenian and Early Bajocian) ammonite biochronology of the Snowshoe Formation, Oregon. Oregon Geology, 50(11–12), 123–137.
Taylor, D. G. (2016). The Weberg and Warm Spring members of the Snowshoe Formation in Suplee area, Oregon: Lithofacies and Aalenian-early Bajocian ammonoid zonation. Carnets De Géologie, 16, 449–490. https://doi.org/10.4267/2042/61389
Westermann, G. E. G. (1992). Ammonite zones of the Circum-Pacific region: Middle Jurassic. In G. E. G. Westermann (Ed.), The Jurassic of the Circum-Pacific (pp. 253–261). Cambridge University Press.
Westermann, G. E. G., & Getty, T. A. (1970). New Middle Jurassic Ammonitina from New Guinea. Bulletin of American Paleontology, 57, 229–321.
Westermann, G. E. G., & Wang, Y.-G. (1988). Middle Jurassic ammonites of Tibet and the age of the lower Spiti Shales. Paleontology, 312, 295–339.
Acknowledgements
This study forms part of the results of the EMMI Research Group (RNM 0178 Junta de Andalucía, Spain). I am also grateful to my colleague G. Jiménez for photographic assistance and to Mr. D. Nesbitt for reviewing the English text. I am very grateful to two anonymous reviews whose constructive comments led to improvements in the final version of this paper.
Funding
This research was supported by EMMI Research Group (RNM 0178 Junta de Andalucía, Spain).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The author declares that there is no conflict of interest.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Sandoval, J. Latiwitchellia (Grammoceratinae?, Ammonitina, Middle Jurassic); an eastern Pacific ammonite in the Betic Cordillera (Southern Spain). J Iber Geol (2024). https://doi.org/10.1007/s41513-024-00253-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s41513-024-00253-x