
Abstract
Ataxioceratinae of the genus Olorizia are characteristic ammonites of the Kimmeridgian Eastern Iberian platform. Olorizia was established as a dimorphic monospecific genus that containing a single species (Olorizia olorizi) endowed with a remarkably wide phenotypic variability. Its morphological plasticity allowed recognizing three morphotypes. They are interpreted as endemic forms and the product of in situ evolution, through a cladogenetic split from the autochthonous lineage of Ardescia Atrops during the Early Kimmeridgian (Platynota Chron); they are also interpreted as one of the greatest exponents of derived morphology in the group due to the achievement of costulation with double connections (ataxioceratoid and subpolyplocoid ribs). In this work, a new dimorphic species is stablished, Olorizia calandensis sp. nov.; the scarce graphic illustration of specimens of the various morphotypes of Olorizia olorizi existing to date is also corrected and the change of institution for the custody of fossil remains is reported, as well as the new identification acronyms.
Resumen
Los Ataxioceratinae del género Olorizia Moliner [m, M] son ammonites característicos de la plataforma ibérica oriental en el Kimmeridgiense. Olorizia se estableció como un género monoespecífico dimórfico que incluía una única especie (O. olorizi) dotada de una variabilidad fenotípica muy amplia. Su plasticidad morfológica permitió reconocer tres morfotipos. Se interpretan como formas endémicas y producto de la evolución in situ, mediante una escisión cladogenética a partir del linaje autóctono de Ardescia Atrops [m, M] durante el Kimmeridgiense temprano (Cron Platynota); también se interpretan como uno de los máximos exponentes de morfología derivada en el grupo por la consecución de costulación con dobles conexiones (costillas ataxioceratoides y subpoliplocoides). En este trabajo se establece una nueva especie dimórfica, Olorizia calandensis sp. nov. [m, M]; también se subsana la escasa ilustración gráfica de ejemplares de los diversos morfotipos de O. olorizi existente hasta la fecha y se informa del cambio de institución para la custodia de los restos fósiles y de las nuevas siglas de identificación.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Ataxioceratins are late Jurassic ammonites that developed shells with complex ornamentation as a phenotypic expression of important evolutionary innovations (e.g., Callomon in Donovan et al., 1981) and thrived mainly on epicontinental platforms of the Tethys, developing extreme phenotypes with a recurrent pattern called diachronic homeomorphism, consisting in the appearance of complex ribs with double connections such as: Ataxioceras (Parataxioceras) praehomalinum Atrops, Schneidia Atrops and Olorizia Moliner during the Platynota Chron and Ataxioceras Fontannes, Parataxioceras Schindewolf and Geyericeras Moliner & Olóriz during the Hypselocyclum Chron, interpreted as forms derived from different species of the genera Ardescia Atrops [m, M] and Lithacosphinctes Oloriz [m, M].
Institutional abbreviations.—UGR, University of Granada, Spain (palaeontological collection of the Department of Palaeontology). MPZ, University of Zaragoza, Spain (palaeontological collection of the Museum of Natural Sciences of the University of Zaragoza).
Other abbreviations: maximum shell diameter measured (Dm), maximum fragmocone diameter (Ph); umbilicus size (U); whorl high (H); whorl width (W); number of umbilical ribs per complete whorl (UR), number of umbilical ribs per half − a − whorl (UR/2); ribbing index calculated as the number of peripheral ribs per ten umbilical ribs (RI), number of constrictions (C). Macroconchs (M), microconchs (m); both dimorphs, macroconch and microconch, identified in a given species (M, m). First Appearance Datum (FAD).
Material storage: The 2066 fossil remains of ataxioceratine ammonites counted in the province of Teruel (eastern Iberian Chain) and collected by Moliner (2009) were deposited in the paleontological collection of the Department of Paleontology of the University of Granada (UGR) from 2009 to 2019, when they were all transferred to the palaeontological collections of the Museum of Natural Sciences of the University of Zaragoza (MPZ), deposit act EMPZ 2019/31, where they are currently housed. The equivalence between the old and current acronyms of all Olorizia specimens is detailed in Table 1.
2 Geological and geographical setting
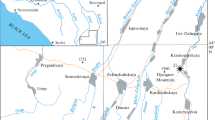
The examined ataxioceratins ammonites come from the detailed and systematic sampling of fifteen sections studied by Moliner (2009) for his doctoral thesis located in the localities of Alcorisa (Gallipuén reservoir and Las Umbrías), Calanda and Foz-Calanda (surroundings of the Calanda reservoir), Berge (San Cristóbal ravine), Molinos (Mezquitilla road), Montoro de Mezquita (Carrera ravine), Villarluengo (La Garrucha mountain range) and Alacón (Cerro Felío), all of them in the NE quadrant of the province of Teruel, Spain (Fig. 1).

Geographic location with reference to the sheets of the National Topographic Map at a scale of 1:50,000
They are located in outcrops of Lower Kimmeridgian materials of the Aragonese Branch of the eastern Iberian Chain and the north of the Maestrazgo, within the so-called Maestrazgo Basin (Figs. 1, 2).

Geological situation. Stars indicate location of the studied sections. Biostratigraphic correlation of materials deposited during the Platynota Chron (Lower Kimmeridgian) and, in yellow, those corresponding to the Olorizia olorizi biohorizon in the sections of Montoro de Mezquita (MBC), Villarluengo (MPR), Alacón (MBP), Molinos (MMM), Berge (MSC and MVP), Alcorisa (MTG1 and MCH), Foz-Calanda (MPC) and Calanda (MLG)
The Maestrazgo Basin of eastern Iberia was formed during the Late Jurassic-Early Cretaceous rifting event that affected Iberia coinciding with the northward spreading of the Central Atlantic rift and the opening of the Tethys Ocean (Aurell et al., 2019; Salas et al., 2001). The Maestrazgo Basin was divided into four sub-basins (depocentres): Aguilón, Oliete, Galve and Morella, controlled by the synsedimentary activity of local tectonic structures between the Late Jurassic and Early Cretaceous.
The outcrops are located in the Morella sub-basin (Aurell et al., 2019), within carbonate rocks (limestones and marls) deposited in a shallow marine carbonate ramp (e.g., Aurell, 1990; Aurell et al., 2019; Bádenas & Aurell, 2001; Moliner, 2009), in the first stage of synrift sequence 1 during the Lower Kimmeridgian.
The rhythmic succession of limestones and marls is included in the Calanda limestones and marls Member. (Meléndez et al., 1990) of the Loriguilla Calcareous Ritmite Formation (Gómez & Goy, 1979).
The Calanda Member contains a high number of ataxioceatine ammonites that allowed us to recognize, with minor changes, the three biochronozones widely identified in the Submediterranean Province and that support the secondary standard for the Submediterranean Lower Kimmeridgian (Fig. 3): Biochronozones of Sutneria platynota, of Ataxioceras hypselocyclum (reinterpreted at a regional level by Moliner in 2009 as Ataxioceras lothari) and of Crussoliceras divisum.

Stratigraphic position of the Olorizia olorizi biohorizon recognised in the eastern Iberian Chain. Biostratigraphic correlation with the Secondary Standard Scheme for the Lower Kimmeridgian used in epicontinental sections of the Submediterranean Province in Europe. Taxonomic abreviations: A, Ataxioceras; Ard, Ardescia; C, Crussoliceras; G, Geyericeras; Garn, Garnierisphinctes; I, Idoceras; L, Lithacosphinctes; O, Olorizia; Orth, Orthaspidoceras; S, Sutneria and Sch, Schneidia
Olóriz et al. (1988) interpreted a turbid shallow-water environment, rich in nutrients and with a restricted connection to the open sea during the Platynota Chron.
3 Material and methods
The collection of ataxioceratins obtained by Moliner (2009) has been examined. Particular attention has been paid to the 80 specimens he included in the genus Olorizia [m, M]; recently, field sampling has also been carried out in the strata where the rarer morphotype of Olorizia and the new species were previously recorded, with negative results.
Bibliography on the Lower Kimmeridgian ataxioceratins which ataxioceratoid or subpolyplocoid ribbing, included in the dimorphic genera Olorizia, Schneidia, Ataxioceras, Parataxioceras and Geyericeras were revised to contextualize the new taxonomical proposals.
Micro and macroconchs of Olorizia olorizi have been photographed to fill in the lack of images since the definition of the taxon.
Priority is given to the use of FADs in biostratigraphic proposals for the definition of biozone and biohorizon or faunal horzonts, often used in biostratigraphy (e.g. Callomon, 1984) and, although although included, but not as a formal chronostratigraphic unit in the International Stratigraphic Guide, has a certain practical value in ataxioceratins studies (e.g. Atrops, 1982; Moliner, 2009Moliner & Olóriz, 2010). As proposed by Callomon (1984), among others, differential spelling has been applied to identify biostratigraphic units and for the index species (in lower case and italics) and their chronostratigraphic equivalents (initials in capital letters and normal font) for both the index species and the age of the chronostratigraphic unit.
4 Remarks on Early Kimmeridgian Ataxioceratinae with double-connected ribs
It has already been indicated that ataxioceratins ammonites thrived mainly on epicontinental platforms of the Tethys Ocean and showed a tendency to reach double-connected ribs (Fig. 4) from several lineages of the genera Ardescia and Lithacosphinctes during the Early Kimmeridgian.

Scheme of some common rib types in the genus Olorizia. a Free external or intercalatory rib. b Bifurcated intercalatory rib. c Simple rib. d Bifurcate rib. e Trifúrcate rib. f Polygirate rib. g, h Ataxioceratoid or polyplocoid ribs. i, j Subpolyplocoid ribs. k Incomplete subpolyplocoid rib. Arrow indicates position of the peristoma. The dashed line indicates half the height of the flank
Olorizia [m, M] Moliner (2009) is characterised by large adult size of microconchs and moderate size of macroconchs, with moderate to low coiling, costulation with widespread development of unrelated subpolyplocoid ribs with a high density of costulation and, to a lesser extent, ataxioceratoid elements. Its stratigraphic range is restricted to the upper part of the Sutneria platynota Biozone. They are considered to be derived forms of the Iberian lineage of Ardescia [m, M], specifically Ardescia casasi Moliner and represent the first known cladogenetic event in this lineage.
Ataxioceras (Parataxioceras) praehomalinum Atrops (1982) is exclusively differentiated from Ardescia desmoides Wegele [m] by the development of ataxioceratoid ribs at the end of the body chamber and, according to Atrops (1982, p.188), probably represents no more than a morphotype progressive from Wegele’s species during the late Desmoides Subchron in SE France. The determinations of this species in the Iberian Range (Fezer & Geyer, 1988; Finkel, 1992) are reinterpreted as phenotypes of Olorizia olorizi Moliner [m] because it comes from Ardescia casasi Moliner, a species considered endemic to the Iberian Peninsula.
Schneidia Atrops (1982) was defined as one of the microconchiate subgenera of Ataxioceras, its record is mainly confined to the youngest part of the Platynota Zone and the earliest part of Hypselocyclum Zone. They have ribs with double connections linked to their strong rib density. They are considered to be forms derived from phenotypes included in Ardescia of the desmoides Wegele group during the Early Platynota Chron (Atrops, 1982, p.156, 265). Schneidia Atrops [m, M], has been reinterpreted as a dimorphic genus (Moliner, 2009; Moliner & Olóriz, 2009; Villaseñor et al., 2015) that includes macroconchs previously referred to Ataxioceras striatellum Schneid (Atrops, 1982, p. 232, 233).
The genus Ataxioceras [m, M] Fontannes (1879) is considered monophyletic and includes the dimorphic pairs with the the most progressive costulation among the Kimmeridgian Ataxioceratinae, with double connected ribs and without parabolic structures. Ataxioceras [m] includes the lothari Oppel group forms, while Ataxioceras [M] includes the hypselocyclum Fontannes group forms (Moliner, 2009; Moliner & Olóriz, 2016). Ataxioceras [m, M] are considered to be derived forms of the more modern phenotypes of Ardescia gr. desmoides Wegele [m, M] during the Hypselocyclum Chron.
The genus Parataxioceras [m, M] Schindewolf (1925) is also considered monophyletic and gathers the dimorph pairs with the most conservative morphology among the Kimmeridgian ataxioceratins. These are phenotypes with slow or very slow coiling, with a somewhat compressed to equidimensional or even slightly depressed whorl, with vigorous ribbing, which, in addition, are provided with double connected ribs and with parabolic structures from the inner whorls. Parataxioceras [m] includes the forms of the oppeli Geyer group and related ones, while Parataxioceras [M] includes the forms of the cardoti Atrops—catenatum Schneid group and related ones (Moliner, 2009). These forms are considered to derive from various populations of Lithacosphinctes Olóriz. [m, M] developed during the Hypselocyclum Chron, as those of the inconditus Fontannes group, since in both cases (Lithacosphinctes and Parataxioceras) relatively vigorous costulation is recognised and provided with parabolic structures from the inner whorls on shells with slow coiling (Moliner & Olóriz, 2009).
Geyericeras [m, M] Moliner & Olóriz, 2010, is characterised by the relatively small adult size of both dimorphs, a relatively high degree of involution of the inner whorls, fine and delicate costulation in microconchs and in immature macroconchs, and the appearance of subpolyplocoid ribs. It is recorded in the upper part of the Ataxioceras lothari biozone identified in the eastern Iberian Range. Geyericeras [m] shows morphological convergence with the most evolute specimens of the stratigraphically older genus Schneidia [m] (Moliner & Olóriz, 2009). It is considered a derived form of the Iberian lineage of Ardescia and represents its last known cladogenetic event.
Several authors illustrated ataxioceratins phenotypes that reached the stage of double-connected ribs, during the Early Kimmeridgian in different epicontinental platforms in central-southern Europe, associated with both the Submediterranean and Subboreal domains (e.g., Oppel, 1862; Vilanova, 1863; Dumortier & Fontannes, 1876; Loriol, 1876; Fontannes, 1879; Quenstedt, 1888; Wegele, 1929; Schneid, 1944; Geyer, 1961; Karve-Corvinus, 1966; Schairer, 1967, 1974, 2000; Maubeuge, 1975; Ziegler, 1977; Atrops, 1982; Marques, 1983; Malinowska, 1986, Fezer & Geyer, 1988; Hantzpergue, 1989; Finkel, 1992; Rodriguez Tovar, 1993; Gradl & Schairer, 1997; Matyja & Wierzbowski, 1998; Gutowski, 1998; Caracuel et al., 1999; Gygi, 2003; Moor, 2009; Moliner, 2009; Grigore, 2011; Comment et al., 2015; Wierzbowski, 2017; Wierzbowski & Glowniak, 2018), in North West Africa (Bachnou & Atrops, 1996; Benzaggagh, 1988; Benzaggagh & Atrops, 1997), in palaeomargins of the northern Tethys, such as Iran (Parent et al., 2014; Zadesmaeil et al., 2021) and on other platforms outside of the Tethian domain, such as Cuba (Sánchez Roig, 1920, 1951) and Mexico (e.g., Cantú-Chapa, 1991, Villaseñor et al., 2015). Early Kimmeridgian ataxioceratins records are found in the Mediterranean epioceanic domain (e.g. Olóriz, 1978; Sapunov, 1977; Sarti, 1993).
Isolated records of Ataxioceras-like ammonites or putative Ataxioceras are also reported but without detailed stratigraphy from the Indo-Malgasy Province and Pacific región, such as India or Japan, (among others, Kobayashi & Fukada, 1947; Krishna et al., 1996, 2009; Sato, 1962, 1992).
However, the palaeobiological interpretation of the Early Kimmeridgian ataxioceratins as dimorphic genera (Comment et al., 2015; Fürsich et al., 2022; Hantzpergue, 1989; Moliner, 2009; Moliner & Olóriz, 2009; Villaseñor et al., 2015) continues to coexist with the partly morphological, partly phyletic interpretation established by Atrops (1982), who considered Ardescia as a microconchiate subgenus of the polyphyletic genus Orthosphinctes Schindewolf, and Schneidia and Parataxioceras as microconchiate subgenera of the genus polyphyletic Ataxioceras Fontannes (e.g., Kiebling & Zeiss, 1992; Gygi, 2003; Moor, 2009; Grigore, 2011; Wierzbowski, 2017; Wierzbowski & Glowniak, 2018; Zadesmaeil et al., 2021).
5 Systematic palaeontology
Class Cephalopoda Cuvier, 1795
Subclass Ammonoidea Zittel, 1885
Order Ammonitida Agassiz, 1847
Suborder Ammonitina Hyatt, 1889
Superfamily Perisphinctoidea Steinmann in Steinmann & Döderlein, 1890
Family Ataxioceratidae Buckman, 1921
Subfamily Ataxioceratinae Spath, 1930 emend. Zeiss, 1968
Génus Olorizia Moliner, 2009 [m, M]
urn:lsid:zoobank.org:act: 4A8D5EE7-8236-4F59-AEE6-9BCA12B119DC
Type species: Olorizia olorizi Moliner, 2009 [m, M]
Other species in the genus: Olorizia calandensis sp. nov. [m, M]
Etymology: Honour to Dr. Federico Olóriz, Professor at the University of Granada (Spain), tireless researcher and teacher; for his scientific contribution to the knowledge of ammonites, biostratigraphy and fossil associations of the Upper Jurassic.
Remarks: Olorizia brings together phenotypes with moderate to low coiling, that develop subpolyplocoid and, to a lesser extent, ataxioceratoid ribs unrelated to a high rib density during the Platynota Chron.
Other genera of contemporary ataxioceratins, such as Ardescia Atrops and Lithacosphinctes Olóriz do not develop ribs with double connections, while Oloriza does.
Schneidia Atrops is more involute and has subpolyplocoid ribs linked to the strong ribbing density.
Ataxioceras Fontannes, Parataxioceras Schindewolf and Geyericeras Moliner & Olóriz, which also develop double connections, are recorded in younger stratigraphic levels (traditional A. hypselocyclum Biozone).
Olorizia developed during the Platynota Chron in an area of the Morella sub-basin with restricted connection to open seas, in which post-mortem shell drift was not significant, nor were relevant alterations of the stratigraphic order such as reworking and associated effects (Moliner, 2009). In this context, the higher abundance of microconchs (4:1 ratio with respect to macroconchs) is interpreted as being due to the greater longevity of macroconchiate counterparts.
Biostratigraphy: The Olorizia record is restricted to the Lower Kimmeridgian, Sutneria platynota Biozone, from the top of the Ardescia desmoides Subzone up the bottom of the Schneidia guilherandense Subzone.
Geographic range: Olorizia is considered a endemic taxon to the eastern Iberian Range and northern Maestrazgo, in the province of Teruel.
5.1 Olorizia olorizi Moliner, 2009 [m, M]
Urn:lsid:zoobank.org:act: 8E3E041D-14DE-4BC8-AD9B-298BDD5920F1.
1988Ataxioceras (Parataxioceras) praehomalinum ATROPS in Fezer & Geyer, 1988, Taf. 4, fig. 1
? pars.1992Ataxioceras (Parataxioceras) hippolytense ATROPS in Finkel, p. 238, Abb. 22; non ? Abb. 26; non ? Abb. 48.
? pars.1992Ataxioceras (Parataxioceras) cf. evolutum ATROPS in Finkel, p. 239, Abb. 31, non Abb. 13.
? pars.1992Ataxioceras (Ataxioceras) homalinum SCHNEID in Finkel, p. 240, Abb. 33; non Abb. 21.
? pars.1992Ataxioceras (Ataxioceras) discobolum (FONTANNES) in Finkel, p. 240, Abb. 38; non? Abb. 25.
2009Olorizia olorizi MOLINER [m, M], p. 201, Lám. 4, figs. 1-5.
2009Olorizia olorizi MOLINER [m] in Moliner & Olóriz, p. 209, fig. 5.

Olorizia [m, M]. Lower Kimmerdgian, Sutneria platynota Biozone, Schneidia guilherandense Subzone, Olorizia olorizi biohorizon. O. olorizi [m] olorizi morphotype. a Specimen MPZ2023/269, b specimen MPZ2023/263, founds in MCH section at Alcorisa. c O. olorizi [M] olorizi morphotype, specimen MPZ2023/294 found in MLG section at Calanda. d O. olorizi [m] gallipuense morphotype, specimen MPZ 2023/308 found in MSC section at Berge. The white dot shows the start of the body chamber

O. olorizi [m] olorizi morphotype. Lower Kimmerdgian, Sutneria platynota Biozone, Schneidia guilherandense Subzone, Olorizia olorizi biohorizon. a Holotype, specimen MPZ2023/253 found in MTG1 section at Alcorisa. b Specimen MPZ2023/271 found in MSC section at Berge. c Specimen MPZ2023/272 found in MPC section at Foz-Calanda. d Specimen MPZ2023/254 found in MTG1 section at Alcorisa. The white dot shows the start of the body chamber

Olorizia [m, M]. Lower Kimmerdgian, Sutneria platynota Biozone, Schneidia guilherandense Subzone, Olorizia olorizi biohorizon. a Olorizia calandensis [m]. Paratype, specimen MPZ2023/330 found in MVP section at Berge. b O. olorizi [M] olorizi morphotype, specimen MPZ2023/295 found in MPC section at Foz-Calanda. c, d O. calandensis [M]. Holotype, specimen MPZ2023/331 found in MPC section at Foz-Calanda. The white dot shows the start of the body chamber

O. olorizi [m] gallipuense morphotype. Lower Kimmerdgian, Sutneria platynota Biozone, Schneidia guilherandense Subzone, Olorizia olorizi biohorizon (specimens a, c, d); Ardescia desmoides Subzone, Ardescia desmoides biohorizon (specimen b). a Specimen MPZ2023/306 found in MSC section at Berge. b, c Specimens MPZ2023/304 and MPZ2023/305 found in MTG1 section at Alcorisa. d Specimen MPZ2023/311 found in MBP section at Alacón. The white dot shows the start of the body chamber

O. olorizi [M] gallipuense morphotype (a), O. olorizi [m] semihomalinum morphotype (b, c, d). Lower Kimmerdgian, Sutneria platynota Biozone, Schneidia guilherandense Subzone, Olorizia olorizi biohorizon. a Specimen MPZ2023/320 found in MPC section at Foz-Calanda. b Specimen MPZ2023/325 found in MMM section at Molinos. c Specimen MPZ2023/322 found in MSC section at Berge. d Specimen MPZ2023/323 found in MMM section at Molinos. The white dot shows the start of the body chamber
Etymology: Honour to Prof. Dr. Federico Olóriz (see etymology of the genus).
Holotype: Specimen MPZ 2023/253 (formerly UGR-MTG1.12.4), internal microconchiate mould, illustrated in Fig. 6a, collected from bed number 12 in the Gallipuén section MTG1 (Province of Teruel, Spain); included in the olorizi morphotype.
Paratypes: [m] 39 specimens. MPZ 2023/252, MPZ 2023/254–279 (27 specimens included in the olorizi morphotype), MPZ 2023/304–313 (10 specimens included in the gallipuense morphotype), MPZ 2023/322–323 (2 specimens included in the semihomalinum morphotype). [M] 8 specimens. MPZ 2023/290–295 (6 specimens included in the olorizi morphotype), MPZ 2023/319–320 (2 specimens included in the gallipuense morphotype).
Complementary material, interpreted as Olorizia cf. olorizi Moliner [m, M], consists of septate internal moulds and more or less complete body chambers showing coiling degree and sculpture equivalent to those described above: [m] 17 specimens. MPZ 2023/280–289 (10 specimens, olorizi morphotype), MPZ 2023/314–18 (5 specimens, gallipuense morphotype), MPZ 2023/324–325 (2 specimens, semihomalinum morphotype). [M] 9 specimens. MPZ 2023/296–303 (8 specimens, olorizi morphotype), MPZ 2023/321 (1 specimen, gallipuense morphotype).
Type locally: Las Umbrías – Gallipuén area, Alcorisa (Province of Teruel, Spain). 30 T 7178666 ED50 – 30 T 4528478.3 ED50.
Type horizon: Lower Kimmeridgian, Sutneria platynota Biozone, uppermost part of Ardescia desmoides Subzone, and base of the Schneidia guilherandense Subzone, Olorizia olorizi biohorizon (Figs. 2, 3).
Diagnosis: Microconchs with moderate to low coiling and tendency to enlarge, size up to 165 mm in diameter and compressed section; constrictions of variable number and amplitude, rare or absent parabolic structures and complex peristome provided with wide lateral lappets. Ribs are more or less dense and fine in the inner whorls, bifurcated and polygyrated with some free external ribs (residual elements); It usually includes subpolyplocoid ribs, which are sometimes incomplete and, less frequently, ataxioceratoid ribs, towards the end of the phragmocone and in the body-chamber. The ribbing index with high values, greater than or equal to 7. The rib curves decrease from small shell diameters, less than 60 mm.
Macroconchs of size less than 250 mm in diameter, section of the whorl ovate, constrictions present, rare parabolic structures in the phragmocone. Simple peristome. The ribbing of the macroconchs is similar to that of the microconchs, but with less frequent polyplocoid ribs and little development of bifurcate intercalatory ribs close ventral margin. In larger diameters, the ribbing is reduced to thick primary ribs close to the umbilical margin. The rib curves decrease from 70 mm or less.
Discussion: Olorizia olorizi is the index species of the O. olorizi biohorizon, which is the lower infrasubzonal unit of the Schneidia guilerandense Subzone in the local biostratigraphic scheme for the Iberian Chain. (Moliner, 2009; Moliner & Olóriz, 2009).
Moliner (2009) included almost all of the phenotypes interpreted as Olorizia in the dimorphic species Olorizia olorizi and, despite being large microconhs and, to a lesser extent, macroconhs, with slow coiling and a low to medium ribbing density at the end of ontogenetic development, Its intraspecific variability allowed it to recognize three morphotypes among the microconch forms and only two among the macroconchs: morphotypes gallipuense [m, M], olorizi [m, M] and semihomalinum [m]. The variability of Olorizia olorizi is manifested without stratigraphic or geographical segregation in the Aragonese Iberian Range. The two remaining specimens are gathered within the new species Olorizia calandensis sp. nov. [m, M].
The only specimens illustrated in the bibliography that could be included within the morphological variability recognized for O. olorizi were recorded in the same sector of eastern Iberian Chain, in noticeably close geographically sections, Calanda (Fezer & Geyer, 1988; Finkel, 1992) and La Ginebrosa by Finkel (1992); however, the section called Calanda by Fezer and Geyer (1988) and Finkel (1992) is not found in the municipality of Calanda, but in the neighbouring locality of Foz-Calanda.. The latter author classified them among several species of Ataxioceras FONTANNES [m, M]. The uncertainty to reinterpret them as Olorizia arises from the younger stratigraphic record indicated by Finkel (op. cit.) for all of them, Hypselocyclum Zone; however, the list of taxa recorded in the same strata, which includes several species of Schneidia, and the fact that the stratigraphic sections of Calanda (Foz Calanda in this work) and La Ginebrosa are not provided, and that the samples were not taken from numbered strata, raises some doubts about the reliability of the biostratigraphy indicated by him and allows, even with some doubt, considering them synonymous with O. olorizi. A. (P.) praehomalinum is not considered because its stratigraphic record is earlier and its most direct ancestors are in Ardescia desmoides, whereas the ancestors of O. olorizi are in Ardescia casasi, which was interpreted as an Iberian endemism.
The analysis of the intraspecific variability of O. olorizi allows establishing the differential characters between the various morphotypes.
The analysis of the rib-curves of the olorizi [m] morphotype allowed us to determine the existence of two populations by ribbing density (Moliner, 2009). The highest cloud on the ordinate axis of the bivariate plots for the number of primary ribs per complete-whorl and per half-a-whorl, groups the most densely ribbed specimens and the lowest cloud groups the population with the lowest rib density. There are intermediate or transitional cases between the two. In addition, specimens from the less densely ribbed population show a tendency towards a slightly higher ribbing index for large shell diameters.
The microconhs of the gallipuense morphotype are more evolute than those of the olorizi morphotype, also by their slower coiling, lower rib density and by having more vigorous primary ribs with greater relief at the umbilical edge. When comparing the rib-curves, a certain overlap can be seen between the gallipuense morphotype and the less densely ribbed population of the olorizi morphotype; in this case the criteria of thickness of the whorl and involution of the shell are effective for their specific differentiation. The macroconchs of the gallipuense morphotype are more evolute than those of the olorizi morphotype, have lower rib density and a higher ribbing index at equivalent diameters; in addition, the primary ribs of the gallipuense morphotype are more reinforced and the constrictions are more pronounced than those of the olorizi morphotype.
The semihomalinum [m] morphotype is differentiated from the rest of the morphotypes of the species by the constant spacing of the primary ribs and by its rectilinear course on the flank; furthermore, this morphotype can be free of constrictions or have a narrow, shallow constriction in the body chamber. The olorizi morphotype has a greater density of ribbing and its primary ribs are less rigid and thick.
Biostratigraphy: Olorizia olorizi ranges in the Lower Kimmeridgian, from upper part of the S. platynota Biozone, uppermost part of the A. desmoides Subzone (A. desmoides biohorizon) up to the lower part of the S. guilherandense Subzone (O. olorizi biohorizon).
Geographic range: The species Olorizia olorizi [m, M] has only been recognized in the eastern Iberian Range and northern Maestrazgo, in the NE quadrant of the province of Teruel, Spain; to date it has been registered in the sections MTG1 and MCH (Alcorisa), MSC and MVP (Berge), MPC (Foz-Calanda), MLG (Calanda), MMM (Molinos), MBC (Montoro de Mezquita), MPR (Villarluengo) and MBP (Alacón). Some specimens included in the synonymy, but originally interpreted as Ataxioceras (Parataxioceras) spp., come from the same outcrops, such as Foz-Calanda (Fezer & Geyer, 1982 and Finkel, 1991), or from very close outcrops, such as La Ginebrosa (Finkel, 1991).
5.2 Olorizia calandensis sp. nov. [m, M]
urn:lsid:zoobank.org:act:17F5F680-9AC2-4967-914F-48442350E925
Figure 7a, c, d.
2009Olorizia n. sp. A [m, M], in Moliner, p. 227.
Etymology: From the village of Calanda (Teruel), around which many specimens of Olorizia have been found.
Type material: An internal macroconchiate mould is chosen as the holotype, specimen MPZ 2023/331 (formerly UGR-MPC.18.18), illustrated in Fig. 7c.d and as a paratype, an internal microconchiate mold, MPZ 2023/330 (formerly UGR-MVP.25.2), illustrated in Fig. 7a.
Type locally: Valdelapiedra ravine, next to the Calanda reservoir in Foz Calanda (province of Teruel) 30 T 733506.1 ED50 – 30 T 4531503.9 ED50.
Type horizon: Lower Kimmeridgian, Sutneria platynota biozone, Schneidia guilherandense subzone, Olorizia olorizi biohorizon (Figs. 2, 3).
Diagnosis: Ataxioceratin moderately large macroconch and moderately small microconch. Coiling degree ca. 40%. Ovate whorl section. Oblique, narrow, shallow constrictions, sparse on the microconch. Ribbing slightly prorsiradiate. Bifurcated and polygyrated ribs with some external intercalatory ribs. Both dimorphs develop subpolyplocoid ribs on the larger diameters. Primary ribs tend to reinforcement on the periumbilical edge at the larger diameters of the macroconch. The sculpture tends to weaken mid-flank in the body chamber and the connections between the primary and secondary ribs are not clear in both dimorphs.
Description: The holotype, MPZ 2023/331, is a macroconch 168 mm in diameter and shows a coiling degree ca. 40% in diameters greater than 125 mm. The end of the phragmocone is at ca. 95 mm and the body chamber is almost one whorl (Table 2). Ovate whorl section with a maximum thickness at mid-height on the flank, with a narrow and rounded venter and slightly convex flanks.
There are four narrow and relatively deep constrictions in the body chamber.
Crowded ribbing on the phragmocone, progressively less accentuated. Ribs are thin, progressively thicker, somewhat prorsiradiate and slightly concave towards the peristome.
Bifurcate and polygirate ribs with some intercalatory ribs (one or two) at the beginning of the body chamber, then sculpture consists of subpolyplocoid common ribs showing the primary/secondary connection more or less close to the mid flank, but always in its inner half.
Reinforcement of ribs on the periumbilical edge is incipient towards the end of the shell, where it also occurs the sculpture smoothing and weakening of connecting points among primary and secondary ribs. The ribbing index increases to 4.
MPZ 2023/330, is an incomplete microconch measuring 129.5 mm in diameter and 40% involution, which preserves only about two-thirds of the whorl corresponding to the body chamber (Table 2).
The section is ovate with the maximum thickness close to the umbilical edge, which is rounded; the flanks are slightly convex and converge in the ventral region, which is broad and convex; the umbilical wall is steep. A deep, narrow constriction is seen at the beginning of the preserved body chamber fragment.
Ribs are thin and maintains its thickness all the way along the flank, somewhat prorsiradiate and slightly concave towards the peristoma. Ribs are bifurcated, with sparse free external ribs intercalated between them; the final third of the preserved body chamber shows polygyrate ribs and at least four subpolyplocoid elements with clear connections. In larger diameters, there is a smoothing of the sculpture and weakening of the connection points between the primary and secondary ribs. The ribbing index is low (3.7–129 mm in diameter).
Discussion: Both dimorphs have similar coiling degree. At equivalent diameter, the ribbing of the macroconch is denser and the ribbing index higher. The microconch develops double-connected ribs at smaller diameter.
The semihomalinum morphotype [m] is the closest morphologically to Olorizia calandensis [m] of the three morphotypes of Olorizia olorizi, considering the uniformity ribbing and the scarcity of constrictions; however, at equivalent diameters, Olorizia calandensis [m] is less evolute, with higher ribbing density, finer primary ribs and the lowest ribbing índex, (3,7 at shell diameters 121–129,5 mm in O. calandensis vs 4,5–5,2 at shell diameters 120,5–134 mm).
Macroconchiates of Olorizia olorizi gallipuense morphotype have a lower ribbing density, the primary ribss are more vigorous, the constrictions are more developed and double-connected ribs are less well achieved.
Differences with other taxa, such as Ardescia, Lithacosphinctes, Schneidia, Ataxioceras, Parataxioceras and Geyericeras have been discussed in the introduction to the genus Olorizia.
Olorizia calandensis sp. nov. is the youngest record of the genus in the sections in which it appears.
Associated ammonites: The enumeration of ammonites identified in the strata where Olorizia calandensis, MPC.18 and MVP.25, was collected is: Ardescia celtiberica Moliner [m], Ardescia strangulata Moliner [m], Ardescia sp. [m], Lithacosphinctes schaireri (Atrops) [m], Schneidia guilherandense guilherandense Atrops [m], Schneidia guilherandense turolense Moliner [m], Schneidia collignoni Atrops [m, M], Schneidia sp. [m], Olorizia calandensis sp. nov. [m, M], Ataxioceratinae [m, M] unspecified, Aspidoceras s.l., Ochetoceras sp., Glochiceratidae unspecified and the zonal index, Sutneria platynota (Reinecke).
Stratigraphic and geographic distribution: The species come from levels of the Sutneria platynota Biozone, Schneidia guilherandense Subzone, Olorizia olorizi biohorizon. The macroconch was found in bed number 18 in the Foz Calanda section (MPC) and the microconch, in the bed number 25 in the Berge section (MVP).
6 Conclusions
The description of the species Olorizia calandensis sp. nov. was made possible by the revision of the Early Kimmeridgian ataxioceratins with with double connected ribs and, especially, by the revision of the genus Olorizia.
The new species, like all the specimens belonging to the genus Olorizia, achieves one of the maximum exponents of the morphology derived from the group, the ribs with double connections at large diameters. Olorizia calandensis sp. nov. develops only double connected ribs of subpolyplocoid type on the body chamber, with the lower connection below half flank height.
Macroconch and microconch dimorphs have been identified.
The palaeogeography, with the formation of the Maestrazgo Basin during the Late Jurassic-Early Cretaceous rifting event that affected Iberia, coupled with the overall geographical and stratigraphic record of Olorizia, allows us to consider the new species as another case of endemism among the Ataxioceratinae of the eastern Iberian platform during the Early Kimmeridgian and as a product of in situ evolution.
The stratigraphic record of Olorizia calandensis is restricted to the middle part of the Schneidia guilerandense Subzone, Olorizia olorizi biohorizon.
Data availability
The data that support the findings of this study are available on request.
References
Agassiz, L. (1847). An introduction to the study of Natural history, in a series of lectures delivered in the hall of the College of Physicians and Surgeons. Greeley & McElrath.
Atrops, F. (1982). La sous-Famillie des Ataxioceratinae (Ammonitina) dans le Kimméridgien inférieur de Sud-Est de la France: Systématique, évolution, chronostratigraphie des genres Orthosphinctes et Ataxioceras. Documents Des Laboratoires De Géologie Lyon, 83, 1–463.
Aurell, M. (1990). El Jurásico Superior en la Cordillera Ibérica Central (provincias de Zaragoza y Teruel). Tesis Doctoral, Dpto. Ciencias de la Tierra, Univ. Zaragoza.
Aurell, M., Bádenas, B., Canudo, J. I., Castanera, D., García-Penas, A., Gasca, J. M., Martín-Closas, C., Moliner, L., Moreno-Azanza, M., Rosales, I., Santas, L., Sequero, C., & Val, J. (2019). Kimmeridgian-Berriasian stratigraphy and sedimentary evolution of the central Iberian Rift System (NE Spain). Cretaceous Research, 103, 104–153. https://doi.org/10.1016/j.cretres.2019.05.011
Bachnou, A., & Atrops, F. (1996). Stratigraphie du Jurassique supérieur dans la région du Moyen Ouerrha (Prérif interne); Comparaison avec l’avant–pays rifain oriental. Mines, Géologie & Energie, 55, 21–30.
Bádenas, B., & Aurell, M. (2001). Kimmeridgian palaeogeography and basin evolution of northeastern Iberia. Palaeogeography, Palaeoclimatology, Palaeoecology, 168, 291–310. https://doi.org/10.1016/S0031-0182(01)00204-8
Benzaggagh, M. (1988). Études stratigraphiques du Jurassique supérieur dans le Prérif interne (Régions de Msila et de Moulay Bouchta, Maroc). Thèse 3 ème cycle, Univ. Lyon I.
Benzaggagh, M., & Atrops, F. (1997). Stratigraphie et association de faune d’ammonites des zones du Kimméridgien, Tithonien et Berriasien basal dans le Prérif interne (Rif, Maroc). Newsletters on Stratigraphy, 35(3), 127–163. https://doi.org/10.1127/nos/35/1997/127
Buckman, S. (1909–1930). Yorkshire type ammonites. Wheldon & Wesley (eds), London, 1–7.
Callomon, J. H. (1984). Biostratigraphy, chronostratigraphy and all that – again! In O. Michelsen, & A. Zeiss (Eds.), International Symposium on Jurassic stratigraphy. Geological Survey of Denmark, Copenhagen (Erlangen 1984) 3, 611–624.
Cantú-Chapa, A. (1991). Ataxioceras (Ataxioceras) lopeztichae sp. nov., amonoideo del Kimeridgiano inferior de Tlaxiaco, Oaxaca, sur de México. Revista De La Sociedad Mexicana De Paleontologia, 4, 17–21.
Caracuel, J., Olóriz, F., & Rodríguez-Tovar, F. J. (1999). Ammonite Biostratigraphy from the Planula and Platynota Zones in the Lugar Section (External Subbetic, southern Spain). Profil, 16, 107–120.
Comment, G., Lefort, A., Koppka, J., & Hantzpergue, P. (2015). Le Kimméridgien d’Ajoie (Jura, Suisse): Lithostratigraphie et biostratigraphie de la Formation de Reuchenette. Revue De Paléobiologie, 34(2), 161–194. https://doi.org/10.5281/zenodo.34341
Cuvier, G. (1797–98). Tableau élémentaire de l’histoire naturelle des animaux. Baudouin.
Donovan, D. T., Callomon, J. H. C., & Howart, M. K., (1981). Classification of the Jurassic Ammonitina. In: M. R. House, & J. R. Senior (Eds.), The Ammonoidea. The Evolution, Classification Mode of Life and Geological Usefulness of a Major Fossil Group. The Systematics Association, Special 18: 101–155.
Dumortier, E., & Fontannes, F. (1876). Description des Ammonites de la zone à Ammonites tenuilobatus de Crussol (Ardèche) et de quelques autres fossiles jurassiques nouveaux ou peu connus. Mémoires De L’académie De Lyon, Classe De Sciences, 21, 1–159.
Fezer, R., & Geyer, O. F. (1988). Der Oberjura von Calanda im nordöstlichen Keltibericum (Provinz Teruel, Spanien). I Stratigraphie. Arbeiten Aus Dem Institut Für Geologie und Paläontologie der Universität, Stuttgart, 84, 207–237.
Finkel, R. (1992). Eine Ammoniten-Fauna aus dem Kimmeridgium des nordöstlichen Keltiberikums (Spanien). Profil, 3, 227–297.
Fontannes, F. (1879). Description des Ammonites des calcaires du Château de Crussol, Ardèche (zones à Oppelia tenuilobata et Waagenia beckeri). Lyon et Savy F., París.
Fürsich, F. T., Schneider, S., Werner, W., Lopez-Mir, B., & Pierce, C. S. (2022). Life at the continental–marine interface: Palaeoenvironments and biota of the Alcobaça Formation (Late Jurassic, Central Portugal), with a formal definition of the unit appended. Palaeobiodiversity and Palaeoenvironments, 102, 265–329. https://doi.org/10.1007/s12549-021-00496-x
Geyer, O. F. (1961). Monographie der Perisphinctidae des unteren Unterkimmeridgium (Weisser Jura γ, Badenerschichten) im süddeutschen Jura. Palaeontographica Abt. A, 117, 1–157.
Gómez, J. J., & Goy, A. (1979). Las unidades litoestratigráficas del Jurásico medio y superior, en facies carbonatadas del sector levantino de la Cordillera Ibérica. Estudios Geológicos, 35, 569–598.
Gradl, H. V., & Schairer, G. (1997). Ammoniten von Kalberberg (Nördliche Frankenalb) (Oberoxford bis Unterkimmeridge). Mitteilungen der Bayererische Staatssamlung für Paläontologie und Historische. Geologie, 37, 9–26.
Grigore, D. (2011). Kimmeridgian-Lower Tithonian ammonite assemblages from Ghilcoş – Hăghimaş Massif (eastern Carpathians, Romania). Acta Paleontologica Romaniae, 7, 177–189.
Gutowski, J. (1998). Oxfordian and Kimmeridgian of the northeastem margin of the Holy Cross Mountains. Central Poland. Geological Quarterly, 42(1), 59–12.
Gygi, R. (2003). Perisphinctacean ammonites of the Late Jurassic in northern Switzerland A versatile tool to investigate the sedimentary geology of an epicontinental sea. Schweizerische Palaontologische Abhandlungen, 123, 1–232.
Hantzpergue, P. (1989). Les Ammonites kimméridgiennes du Haut-fond d’Europe occidentale. Biochronologie, Systématique, Évolution, Paléobiogéographie. Cahiers de Paléontologie. CNRS, Paris.
Hyatt, A. (1889). Genesis of the Arietidae. Smithsonian Contributions, 673. Memoirs of the Museum of Comparative Zoölogy. Cambridge, XVI (3).
Karvé-Corvinus, G. (1966). Biostratigraphie des Oxfordium und untersten Kimmeridgium am Mont Crussol, Ardèche, im Vergleich mit Suddeutschland. Neues Jahrbuch Für Geologie und Paläontologie Abhandlungen, 126(2), 101–141.
Kiebling, W., & Zeiss, A. (1991–1992). New palaeontological data from the Hochstegen Marble (Tauern Window, Eastern Alps). Geologisch-Paläontologische Mitteilungen Innsbruck, 18, 187–202.
Kobayashi, T., & Fukada, A. (1947). A new species of Ataxioceras in Nippon. Japanese Journal of Geology and Geography, Transactions and Abstracts, 20(2–4), 45–48.
Krishna, J., Pandey, B., Ojha, J. R., & Pathak, D. B. (2009). Reappraisal of the age framework, correlation, environment and nomenclature of Kachchh Mesozoic lithostratigraphic units in Wagad. Journal of Scientific Research, 53, 1–20.
Krishna, J., Pathak, D. B., & Pandey, B. (1996). Quantum refinement in the Kimmeridgian ammonoid chronology in Kachchh (India). GeoResearch Forum, 1(2), 195–204.
Loriol, P. de (1876–1878). Monographie paléontologique des couches de la zone á Ammonites tenuilobatus (Badener Schichten) de Baden (Argovie). Mémoires de la Société Paléontologique Suisse, 3, 1–32 (1876); 4, 33–76 (1877); 5, 77–200 (1878).
Malinowska, L. (1986). Paleozoogeography and Paleoecology of the Lower Kimmeridgian of Poland. Bulletin of the Polish Academy of Sciences. Earth Sciences, 34(2), 165–173.
Marques, B. (1983). O Oxfordiano-Kimeridgiano do Algarve Oriental: estratigrafia, paleobiologia (Ammonoidea) e paleobiogeografia. Ph.D. Thesis, Univ. Nova de Lisboa.
Matyja, B. A., & Wierzbowski, A. (1998). The stratigraphical and palaeogeographical importance of the Oxfordian and Lower Kimmeridgian succession in the Kcynia IG IV borehole. Biuletyn Państwowego Instytutu Geologicznego, 382, 35–69.
Maubeuge, P. L. (1971–1974). Catalogue des Ammonites du Jurassique supérieur (Callovien à Kimmeridgien) du Musée cantonal de Bâle-Campagne. Quatrième partie. Tätigkeitsberichten der Naturforschenden Gesellschaft Baselland, 29, 40–338.
Meléndez, G., Aurell, M., & Atrops, F. (1990). Las unidades del Jurásico superior en el sector nororiental de la Cordillera Ibérica: Nuevas subdivisiones litoestratigráficas. Cuadernos De Geología Ibérica, 14, 225–245.
Moliner, L. (2009). Ataxioceratinae (Ammonitina) del Kimmeridgiense inferior en el NE de la provincia de Teruel (Cordillera Ibérica Oriental y Maestrazgo). Granada.
Moliner, L., & Olóriz, F. (2009). Updated biostratigraphy of Jurassic (lower Kimmeridgian) deposits containing the ammonite Ataxioceras from the eastern Iberian Range, northeastern Spain. Geologiska Föreningens i Stockholm Förhandlingar, 131, 193–201. https://doi.org/10.1080/11035890902952894
Moliner, L., & Oloriz, F. (2010). New Lower Kimmeridgian ataxioceratin ammonite from the eastern Iberian Chain, Spain: Systematic, biogeographic and biostratigraphic relevance. Acta Palaeontologica Polonica, 55(1), 99–110. https://doi.org/10.4202/app.2008.0064
Moliner, L., & Oloriz, F. (2016). Revisión de los primeros Ataxioceratinae (Cephalopoda, Ammonitina) del Kimmeridgiense figurados en España: La contribución de Juan Vilanova y Piera, 1863 [-1870]. Boletín De La Real Sociedad Española De Historia Natural Sección Geológica, 110, 5–27.
Moor, E. (2009). Oxydiscites und Cymaceras vom Schaffhauser Randen. Mitteilungen der Naturforschenden Gesellschaft, Schaffhausen, 49, 1–38.
OIóriz, F., Marques, B., & Moliner, L. (1988): The Platform-effect: an example from Iberian Shelf areas in the lowermost Kimmeridgian. 2nd International Symposium on Jurasssic Stratigraphy, II, 534–562.
Olóriz, F. (1978). Kimmeridgiense-Tithónico inferior en el Sector Central de las Cordilleras Béticas (Zona Subbética). Paleontología. Bioestratigrafía. Tesis Doct. Univ. Granada, 184.
Oppel, A. (1862–1863). Über jurassische Cephalopoden. Mittheilungen, Palaeontologische aus dem Museum des Koenigl. Bayer. Staates, 3, 127–162 (1862); 163–266 (1863).
Parent, H., Raoufian, A., Seyed-Emami, K., Ashouri, A. R., & Majidifard, M. R. (2014). The Bajocian-Kimmeridgian fauna of the Dalichai Formation in the SE Binalud Mountains, Iran. Reports of the Instituto De Fisiografía y Geología, 1, 1–60.
Quenstedt, F. A. (1887–1888). Die Ammoniten des Schwäbischen Jura. III. Der Weisse Jura. E. Schweizerbart’sche Verlags, III, 817–1140.
Rodríguez-Tovar, F. J. (1993). Evolución sedimentaria y ecoestratigráfica en plataformas epicontinentales del margen Sudibérico durante el Kimmeridgiense inferior. Tesis Doctoral, Univ. Granada.
Salas, R., Guimera, J., Mas, R., Martín-Closas, C., Meléndez, A., & Alonso, A. (2001). Evolution of the Mesozoic central Iberian rift system and its Cenozoic inversion (Iberian chain). Mémoires Du Muséum National D’histoire Naturelle, 186, 145–185.
Sánchez Roig, M. (1920). La fauna jurásica de Viñales. Secretaria de Agricultura, Comercio y Trabajo, Boletin especial, 1–61.
Sánchez Roig, M. (1951). La fauna jurásica de Viñales. Anales De La Academia De Ciencias Médicas, Físicas y Naturales De La Habana, 89(2), 46–94.
Sapunov, I. G. (1977). Ammonite Stratigraphy of the Upper Jurassic in Bulgaria. III. Kimmeridgian: substages, zones and subzones. Geologica Balcanica, 7(1), 63–80.
Sarti, C. (1993). Il Kimmeridgiano delle Prealpi Veneto-Trentine: Fauna e biostratigrafía. Memorie Del Museo Civico Di Storia Naturale Di Verona (IIa Serie). Sezione Scienze Della Terra, 5, 1–145.
Sato, T. (1992). Japan. In G. E. G. Westermann (Ed.), The Jurassic of the Circum-Pacific. Part III: Regional geology and stratigraphy, 9 Southeast Asia and Japan (pp. 194–206). Cambridge.
Sato, T. (1962). Etudes biostratigraphiques des Ammonites du Jurassique du Japon. Mémoires De La Société Géologique De France, Nouvelle Série, 41, 1–122.
Schairer, G. (1967). Biometrische Untersuchungen an Perisphinctes, Ataxioceras, Lithacoceras der Zone der Sutneria platynota (REINECKE) (unterster Unterkimmeridgium) der Fränkischen Alb. Dissertation, Univ. München.
Schairer, G. (1974). Quantitative Untersuchungen an Perisphinctidae (Ammonoidea) des untersten Unterkimmeridgium der Fränkischen Alb (Bayern). Zitteliana, 3, 37–124.
Schairer, G. (2000). Die Ammoniten des Schurfs von Mantlach (Oberoxfordium-Unterkimmeridgium, Südliche Frankenalb, Bayern). Allgemeines und die Ammonitenfauna aus Bank 60. Mitteilungen der Bayerischen Staatssammlung Für Paläontologie und Historische Geologie, 40, 81–107.
Schindewolf, O. H. (1925). Entwurf einer Systematik der Perisphincten. Neues Jahrbuch Für Mineralogie, Geologie und Paläontologie, 52(B), 309–343.
Schneid, T. (1944). Uber Ataxioceratiden der nördlichen Frankenjura. Palaeontographica Abteilung, A, 96, 1–43.
Spath, L. F. (1927–1933). Revision of the Jurassic Cephalopod Fauna of Kachh (Cutch). Memoirs of the Geological Survey of India, Palaeontologia Indica, 9(2), 551–945.
Steinmann, G., & Doderlein, L. (1890). Elemente der Paläontologie.
Vilanova, J. (1863 [1870]). Ensayo de descripción geognóstica de la provincia de Teruel, en sus relaciones con la agricultura de la misma. Junta General de Estadística. Madrid.
Villaseñor, A. B., Moliner, L., & Olóriz, F. (2015). Schneidia zacatense sp. nov. First population level study of Ataxioceratine from the Lower Kimmeridgian in northern Mexico. Biostratigrphic and palaeobiogeographic significance. Journal of Sputh American Earth Sciences, 63, 217–243. https://doi.org/10.1016/j.jsames.2015.07.018
Wegele, L. (1929). Stratigraphische und faunistische Untersuchungen im Oberoxford und Unterkimmeridge Mittelfrankens. Palaeontograhica, LXXI, 117–210; LXXII, 1–94.
Wierzbowski, A. (2017). The Lower Kimmeridgian of the Wieluń Upland and adjoining regions in central Poland: lithostratigraphy, ammonite stratigraphy (upper Planula/Platynota to Divisum zones), palaeogeography and climate-controlled cycles. Volumina Jurassica, XV, 41–120. https://doi.org/10.5604/01.3001.0010.5659
Wierzbowski, A., & Glowniak, E. (2018). The Early Kimmeridgian succession at Kodrąb (Radomsko elevation, central Poland) and its palaeogeographical and palaeotectonic implications. Geological Quarterly, 62(3), 509–521. https://doi.org/10.7306/gq.1421
Zadesmaeil, M., Majidifard, M. R., Vaziri, S., & Jahani, D. (2021). Lithostratigraphy and Biostratigraphy of the Dalichai Formation in Absharaf Section, Northwest of Damghan on the base of Ammonites (East Alborz). Journal of Geosciences, 31(120), 91–100. https://doi.org/10.22071/GSJ.2017.97439.1250
Zeiss, A. (1968). Untersuchungen zur Paläontologie der Cephalopoden des UnterTithon der Südlischen Frankenalb. Abhandlungen Bayerische Akademie der Wissenschaften Mathematisch-Naturwissenschaftliche Klasse, 132, 1–190.
Ziegler, B. (1977). The “White” (Upper) Jurassic in Southern Germany. Stuttgarter Beiträge Zur Naturkunde. Serie B, 26, 1–79.
Zittel, K. A. von (1884). Cephalopoda. In: Handbuch der Paläontologie (Part 1, Vol. 2, pp. 329–522). Munich.
Acknowledgements
I appreciate the effort and patience of my Mentor Teacher, Dr. Federico Olóriz Sáez, Professor at the University of Granada, to whom I will always be indebted for all he taught me. The ammonites were photographed by Zarela Herrera and Isabel Pérez Urresti. I would like to thank the editors of this volume and the reviewers of this paper, Eva Glowniak and an anonymous person, wich reviewed the paper in detail. Their comments and suggestions were very helpful.
Funding
This work has not received specific support from public sector agencies, the commercial sector or non-profit organisations.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
I know of no conflicts of interest associated with this publication.
Ethical statement
I declare that this manuscript is original, has not been published before and is not currently being considered for publication elsewhere.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Moliner, L. Update of the dimorphic genus Olorizia (Ataxioceratidae, Ammonitina) and establishment of the species Olorizia calandensis sp. nov. J Iber Geol (2024). https://doi.org/10.1007/s41513-024-00245-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s41513-024-00245-x