
Abstract
Scleractinian corals of late Cenomanian to early Turonian age from the western border of the Iberian Peninsula are described for the first time. They derive from middle to inner shelf limestones of the mid Albian-Turonian West Portuguese Carbonate Platform, near the localities of Figueira da Foz, Leiria, and Nazaré, where a succession of fossil-rich beds with cephalopods, rudists, and other invertebrates allows an accurate biostratigraphic control from the basal upper Cenomanian to the upper lower Turonian. During this interval, facies are deeper and carbonate-rich, that are more favourable to the occurrence of hermatypic corals and which are absent in previous sequences of the platform. The studied corals belong to the superfamilies Actinastreoidea, Cladocoroidea, Cyclolitoidea, Madreporoidea, Phyllosmilioidea, and Poritoidea. With a total of 11 species, the number of taxa is comparatively low when compared with other Late Cretaceous faunas. The corals show relationships to Albian coral faunas from the SE Iberian Platform and the Quillan Basin (SW France), both of which are located in the Tethyan Realm. The faunal turnover in scleractinian corals at the Cenomanian/Turonian (C/T) boundary is discussed. Scleractinian corals were rich in genera during the Cenomanian and reach about 110 co-existing genera at the base of the late Cenomanian. This number was reduced by 46 genera that became extinct at the Cenomanian/Turonian boundary, but were replaced rapidly after the boundary by 59 genera, of which six became extinct before the Coniacian. Sixty-six genera survived the C/T boundary, of which 11 became extinct before the middle Coniacian. This means that the Cenomanian richness in genera was almost restored in the Coniacian.
Resumen
Se describen por primera vez los corales escleractinidos del periodo Cenomaniense superior - inicio del Turoniense del margen occidental de la península Ibérica. Los ejemplares fueron obtenidos en secciones de la plataforma carbonática media e interna de edad Albiense-Turoniense del oeste de Portugal, próximas a las localidades de Figueira da Foz, Leiria, y Nazaré. Este registro también contiene una sucesión de capas ricas en otros fósiles como cefalópodos, rudistas y otros invertebrados que permiten un control preciso sobre la bioestratigrafía desde el Cenomaniense superior hasta el Turoniense superior. Durante este intervalo de tiempo, las facies son más profundas y ricas en carbonatos, siendo más favorable a la aparición de corales hermatípicos que estaban ausentes en el registro anterior de la plataforma. Los corales estudiados pertenecen a las superfamilias Actinastreoidea, Cladocoroidea, Cyclolitoidea, Madreporoidea, Phyllosmilioidea y Poritoidea. Aunque muestran un total de 11 especies, la cantidad de taxones es comparablemente baja en comparación con otras faunas del Cretácico Superior. Los corales muestran relaciones con otros corales del Albiense del margen sureste de la plataforma Ibérica, y con la cuenca de Quillan (SO de Francia), ambas zonas localizadas en el mar de la Tethys. Se discute el cambio de las faunas de escleractinidos en el límite Cenomaniense/Turoniense (C/T). durante el Cenomaniense orden Scleractinia era rico en géneros en la zona estudiada, llegando a alcanzar aproximadamente 100 géneros en la base del Cenomaniense superior. Esta cantidad se reduce hasta 46 géneros en el límite Cenomaniense/Turoniense, pero se recuperó nuevamente después de dicho límite hasta alcanzar los 59 géneros, de los cuales 6 se extinguieron durante el Turoniense. Sesenta y seis géneros sobrevivieron el límite C/T, de cuales 11 géneros se extinguieron antes del Coniaciense medio. Eso indica que la riqueza de génerosque había en el Cenomaniense fue casi restaurada en el Coniaciense.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Scleractinian corals are a widespread invertebrate group in Mesozoic and Cenozoic shallow marine environments. Their skeletal remains are common on the margins and inner areas of rimmed carbonate platforms with reef or constratal structures, where an outstanding fossil record reveals the key importance of hermatypic corals as the frame builders, alongside sponges and microbialites (e.g., James, 1983; Tucker & Wright, 1992). When symbiotic with zooxanthellae, their modern representatives are stenotypic and live in fully marine warm environments with rather narrow ecological requirements. These biological constraints can be used to infer aspects that are related to the habitats and the way of life of Mesozoic fossil corals, using their occurrence for integrated palaeoecological studies and palaeoenvironmental characterisation of carbonate series.
The study of coral assemblages becomes even more interesting and well documented regarding the Cretaceous Period. During this long interval of Earth’s history and until the late Maastrichtian major biotic changes occurred (Skelton, 2006). The same warm and shallow shelf environments of the Tethyan carbonate platforms where scleractinians had prospered, were extensively colonised by a large diversity of rudist bivalves, better adapted to a new range of ecological conditions that were allowed by high sealevel intervals, including reduced clastic input from the marginal marine areas of the continents, and changes in the Ca/Mg proportion (Gili & Götz, 2018). As a question of sub-environments, only rudists were major carbonate frame contributors, but they were not true reef builders. Coral richness at the genus level varied throughout the Cretaceous with peaks during the early Aptian and the Cenomanian, and considerable drops in the middle Albian and early to middle Turonian (Löser, 2016: 103). Generic richness varied not only through time but also from area to area such as, for example, in the Western Tethys where coral richness on the genus level was high from the Barremian to early Albian, and during the Cenomanian, Santonian and early Campanian, whereas in the Central Tethys it was high during the late Barremian and early Aptian, and during the early Cenomanian and Coniacian to Santonian. The Western Atlantic shows a completely different pattern where coral richness was high from the late Barremian into the early Albian and in the Maastrichtian (Löser, 2016: 125).
The present study focuses on the westernmost part of the Teythan platforms lying within the European branch of Tethys and its upper Cenomanian and lower Turonian coral assemblages, at present still poorly described regarding palaeontological aspects. The West Portuguese Carbonate Platform (WPCP) was set up at the palaeo-Atlantic margin of the Iberian microplate, acting as a post-rift shallow sea that was opened to repeated faunal changes between the Tethys Realm and the temperate Boreal domain. It was a wide shallow marine area colonised by scleractinian corals and rudists such as the Caprinulidae and Radiolitidae. Although these bivalves have long been studied, the Cenomanian–Turonian coral assemblages remained rather unknown concerning their taxonomy, age, palaeobiology and biogeographic range.
2 Location and stratigraphical setting
2.1 Geological setting
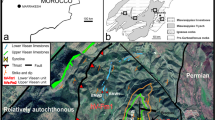
The studied area is located at the western border of the Iberian Peninsula, in the northern Estremadura and Beira Litoral ranges of west central Portugal, near the localities of Figueira da Foz, Leiria and Nazaré (Fig. 1). It is a low relief basin area with a geological setting of Mesozoic and Cenozoic sedimentary units from the Atlantic West Iberian Margin (WIM; Kullberg et al., 2013) bounded eastwards by a Variscan basement which acted as the main sediment source since the breakup of Pangaea (Soares et al., 2012).

Geographic location and geological context of the studied area. A Location on the western border of Iberia. B Detailed geographical and geological setting of the Beira Litoral and Estremadura ranges from the onshore region of the West Iberian Margin, with location of the sampled localities with upper Cenomanian and lower Turonian corals of the West Portuguese Carbonate Platform; a Caranguejeira, Leiria; b Nazaré; c Salmanha, Figueira da Foz. 1 Recent to Lower Cenozoic siliciclastics; 2 Turonian to Maastrichtian marginal marine and alluvial siliciclastics; 3 Middle Cenomanian to lower Turonian platform carbonates; 4 Aptian to middle Cenomanian alluvial siliciclastics; 5 Sinemurian to uppermost Jurassic syn-rift carbonates and Upper Jurassic interbedded sandstones of the Lusitanian Basin infill; 6 Triassic coarse siliciclastics and Hettangian evaporites; 7 Pre-Mesozoic Variscan basement
The regional stratigraphy shows a thick succession of Mid Triassic to Upper Cretaceous siliciclastic, mixed and carbonate formations. They record the tectono-sedimentary evolution of the Lusitanian Basin, through four rifting phases of the proto-Atlantic opening and several intervals of thermal detumescense and accommodation of marine carbonates (Azerêdo et al., 2003; Wilson et al., 1989). The post-rift transition to a passive margin took place from the late Aptian onwards, with the deposition of Albian to Maastrichtian siliciclastics and interbedded platform carbonates (Dinis et al., 2008; Segura et al., 2014).
From the mid Albian to the Cenomanian and early Turonian, during a long-term interval of eustatic rise and highstand with widespread marine sedimentation over many worldwide continental margins (Haq, 2014; Hardenbol et al., 1998), the Estremadura and Beira Litoral ranges of the WIM gradually become filled by inner shelf sediments of the West Portuguese Carbonate Platform (WPCP; Berthou, 1984; Soares, 1980; here Fig. 2). The first carbonated sequences were transgressive over post-rift coarse alluvial siliciclastics and confined to the region of Lisbon in south Estremadura (Rey, 2006). During the middle Cenomanian the platform carbonates gradually extended to the remaining onshore sectors of the continental margin. This part of the succession is dominated by oyster-rich inner-shelf facies with a rather high siliciclastic input, more related to a mixed carbonate platform environment less favourable for the occurrence of scleractinian corals.

Schematic diagram of the upper Aptian to lower Campanian post-rift series of the Atlantic West Iberian Margin with emphasis on the West Portuguese Carbonate Platform (WPCP) record of the Estremadura and Beira Littoral ranges of west central mainland Portugal. D1 Main intra-Aptian unconformity related to tectonic uplift and beginning of widespread coarse siliciclastic drainage; D2 Cenomanian–Turonian unconformity related to uplift and subaerial exposure, and karst development on the WPCP; D3 Main intra-Campanian unconformity followed by deep facies changes to alluvial reddish lutites (”Garum” series)
The basal upper Cenomanian is marked by widespread transgressive nodular limestone sequences (Callapez, 2008). They yield a typical Tethyan assemblage with the ammonite Neolobites vibrayeanus and a rich and diverse fauna of benthic invertebrates, including many molluscs and echinoids (Costa d’Arnes Formation; unit “C”; Berthou, 1984; Callapez, 2004; here Fig. 3A). These open shelf facies are overlain by a thick bed of coarse-grained bioclastic limestone with coral debris, nerineacean gastropods, and epibenthic bivalves (unit”D”), showing a significant lateral persistence throughout the studied area.

Stratigraphic setting of the sampled localities with Cenomanian and lower Turonian corals of the West Portuguese Carbonate Platform: Salmanha (Figueira da Foz), Nazaré and Caranguejeira (Leiria). A–C Sections and local stratigraphic units; a–d Location of the collected samples; a Middle upper Cenomanian outer shelf limestones of Salmanha, b Middle lower Turonian coarse bioclastic limestones of Salmanha, c Middle lower Turonian coarse bioclastic micaceous limestones of Nazaré (northern beach), d Middle upper Cenomanian rimmed shelf limestones of Caranguejeira. 1–12 Main fossil occurrences; 1 Scleractinian corals, 2 Pteriomorphia bivalves (except Ostreina), 3 Oysters and related bivalves, 4 Heterodonta bivalves (except rudists), 5 Caprinulid rudists, 6 Radiolitid rudists, 7 Nerineacean gastropods; 8 Tylostomid and other mesogastropods; 9 Acteonellid gastropods; 10 Cephalopods, 11 Echinoids, 12 Vertebrate remains
The next upper Cenomanian succession shows a noteworthy lateral diversification of facies over the WPCP. In the northern areas between Figueira da Foz and Coimbra (Baixo Mondego region), they consist of open shelf limestones and marly limestones with Vascoceras ammonites, Hemiaster echinoids and Tylostoma gastropods, among others, and more locally they consist of small branched corals (units “E” to “J” Berthou, 1984; Callapez, 2008). To the south, the correlative sequences of Leiria contain thick limestone beds of a rimmed carbonate platform with platy corals present in slope areas (unit “G”) and Caprinula, Durania and Radiolites buildups (unit “F” to “J”; here Fig. 3B). These rudist facies are also widespread across central and south Estremadura, from Nazaré towards the regions of Sintra and Lisbon (Berthou, 1984).
The uppermost Cenomanian record of the WPCP is limited by a large unconformity with subaerial exposure and palaeokarst development of the carbonate beds, which can be related to an uplift episode occurred at the Cenomanian–Turonian transition (Callapez, 2008). The overlying sequences are mid-early Turonian in age, showing a geographical range restricted to a few areas of Beira Litoral, Nazaré and Leiria. Near Figueira da Foz, they begin with massive micritic limestones (unit “K”) followed by beds of coarse-laminated marly limestone with a rich assemblage of ammonites with North-African affinities (unit “L”; Barroso-Barcenilla et al., 2011). These sequences are correlative to a narrow fringe of Radiolites biostromes that lies around Leiria, and to several layers of micaceous, bioclastic limestone in the Nazaré area (Callapez, 2008). The upper part of the lower Turonian WPCP succession is markedly regressive, including carbonate sandstones with small massive corals, rudists and Actaeonella gastropods in the Nazaré exposure (Callapez, 2004; Fig. 3C). Near Figueira da Foz, a correlative succession of cross-bedded calcarenite shoals extensively outcrops with remains of scleractinian corals, Radiolites rudists and nerineacean (Plesioptygmatis) gastropods (units “M”, “N”, and “O”). The occurrence of Actaeonella caucasica also suggests a late early Turonian age. These sequences interfinger with a progradational body of littoral plain micaceous sandstones that marks the final retreat of the WPCP to offshore domains.
Despite this rather diverse set of stratigraphic units, the coral record of the WPCP is comparatively scarce in the number of localities where representatives of this group are present as a major part of the invertebrate assemblages. For this study, the sampling of coral remains was centred on upper Cenomanian and lower Turonian carbonate beds of three main sections of the northern and central ranges of the carbonate platform (Figs. 1B and 3) where these invertebrates occur in significant numbers: Salmanha (Figueira da Foz), Souto de Cima and Carrasqueira (Caranguejeira, Leiria) and the Northern beach of Nazaré.
2.2 Localities
The samples of Salmanha, in the northern range of the WPCP, were collected from two distinct parts of the local composite section (Fig. 3A, sampling points “a” and “b”), all of them with accurate biostratigraphical data provided by ammonites, inoceramids and foraminifera (Barroso-Barcenilla et al., 2011; Berthou, 1984; Callapez, 1998, 2004, 2008; Segura et al., 2014). They include the Casal dos Touros quarry where the mid upper Cenomanian unit ”H” is well exposed, always with a facies of whitish massive limestone (Fig. 4A, B) rich in Pycnodonte vesiculare, Tylostoma ovatum and Hemiaster scutiger, besides many coral representatives. Additional material was collected near the top of the WPCP succession, where the upper lower Turonian unit “M” extensively outcrops in slopes of the highway and a nearby supermarket storage area (Fig. 4C). Here, the local facies appears as a whitish limestone with low-angle planar cross-bedded structure and calcarenitic to calciruditic texture, sometimes with rather large concentrations of bioclasts, including many coral fragments. The geographical coordinates of both these sampling points are, respectively: 40° 9′ 4.75″ N; 8° 49′ 28.46″ W and 40° 8′ 54.86″ N; 8° 49′ 53.64″ W. To the South, in the central range of the WPCP, corals were sampled in the localities of Nazaré, and Souto and Carrasqueira, near Caranguejeira (Leiria), where an extensive area of mid upper Cenomanian rudist rich bioclastic limestones occurs. The first of these localities has an impressive exposure of the Cenomanian and Turonian WPCP units, with a continuous section (Fig. 3C, sampling point “c”). The topmost layers of the succession are accessible through the Northern beach way, where a coral-rich bed of whitish-pink micaceous limestone outcrops (Fig. 4D), which is overlain by yellowish cross-bedded coarse sandstones with abraded debris of Radiolites and Actaeonella. This sampled coral bed is correlative to the upper lower Turonian unit “M” of Figueira da Foz in the above, with geographical coordinates of 39° 36′ 23.59″ N; 9° 5′ 2.30″ W. Located about 30 km Northeast of Nazaré, the sampling area of Souto de Cima and Carrasqueira near Caranguejeira (Leiria) is known by its accessible large exposures of calcarenite limestone with rudist buildups and coral-rich levels in slopes of the roads to Caranguejeira and Padrão (Fig. 4E). Two sampling points where chosen: near Souto de Cima, from a thick whitish limestone with planar cross-bedded structure, and Caprinula and Radiolites massive debris, and along the Carrasqueira karstic valley where a set of platy coral-rich beds outcrop near the main road (Fig. 3C, sampling points “d” and “e”). These parts of the local WPCP succession are both mid-late Cenomanian in age and are the first correlative of unit “H” of Figueira da Foz and the Carrasqueira unit which is laterally equivalent to unit “G”. Their geographical coordinates are: 39° 44′ 17.13″ N; 8° 43′ 15.44″ W, and 39° 45′ 28.06″ N; 8° 43′ 36.53″ W, respectively.

Photographic views of the sampled sections from the Cenomanian–Turonian carbonate platform succession of West Central Portugal. A Salmanha (Figueira da Foz); aspect of the upper Cenomanian local units”H”,”I” and”J”. Unit”H” is a massive whitish limestone with fragments of branched corals, infralittoral bivalves (Pycnodonte), gastropods (Haustator, Tylostoma), Vascoceratid ammonoids and infaunal echinoids (Mecaster); B Salmanha (Figueira da Foz); Detail of the sampled middle upper Cenomanian massive limestone with fragments of branched corals (Unit”H”; coin diameter = 1.6 cm); C Fontela (Figueira da Foz); aspect of the middle lower Turonian coarse limestone beds of unit”M” with scattered concentrations of corals and debris of rudist bivalves (Radiolites); D Nazaré (Northern beach of the Sítio); view of the local lower Turonian succession (units”L” and”M”) with pink micaceous limestones with corals, rudist bivalves (Radiolites) and gastropods (Actaeonella) followed by strongly regressive, alluvial coarse limestones with abraided debris of the same fossil-association; E Padrão (Leiria); panoramic view of upper Cenomanian massive and laminated limestone beds (unit”G”) with autochthonous concentrations of platy corals
3 Materials and methods
Specimens were sampled during fieldwork research of the Cenomanian and lower Turonian carbonate successions of Figueira da Foz, Leiria and Nazaré. All samples were collected from stratigraphic sections with a detailed biostratigraphical setting based on ammonites and foraminifera (Barroso-Barcenilla et al., 2011; Berthou, 1984; Callapez, 1998, 2004, 2008; Segura et al., 2014), and they had been previously studied for invertebrate palaeoecology purposes (Callapez, 1998, 2008).
The selection of levels to sample was chosen based on specific criteria, namely the carbonate facies, abundance and diversity of coral specimens, preservation, accessibility of beds, and degree of biostratigraphical control. The coral specimens were collected from the local units “F”, “G”, “H” and “M” of the WPCP in the outcrops of Salmanha (Figueira da Foz), Carrasqueira (Leiria) and northern beach of Nazaré. The studied collection consists of around 50 samples that were collected and processed, twenty-four of which were included in the taxonomic study.
Coral specimens were cut and polished from samples with a rather embedded carbonate matrix. Thin sections in both transversal and longitudinal orientation were prepared where possible. Thin sections were scanned by passing light through them using a flatbed scanner with an optical resolution of 6400 dpi. Scanned images were then transferred to greyscale bitmaps. Their quality was amended by histogram contrast manipulation (contrast stretching) where possible. Corallite dimensions were systematically measured. Values were calculated using the Palaeontological Database System PaleoTax, module PaleoTax/Measure (https://www.paleotax.de/measure). The procedure and statistical background are described in Löser (2012). For each type of measurement (corallite diameter and distance) in one thin section the following values were obtained and/or calculated: n, number of measurements; min–max, lowest and highest measured value; µ, arithmetic mean (average); s, standard deviation; cv, coefficient of variation; µ ± s, first interval. The obtained morphometric values of the fossils were compared against those on specimens in worldwide fossil coral collections using a PaleoTax database (Löser, 2004) on post-Paleozoic corals and associated image database which has 27,600 specimens, around 15,800 of which are illustrated, that are located in the Noroeste Regional Station (ERNO) in Sonora, Mexico which is a site of the National Autonomous University of Mexico (UNAM). The studied specimens and thin sections are housed in the Earth Sciences Department of the University of Coimbra (DCTUC) in Portugal under the numbers DCT/UC/CRET/2500, 2504, 2507, 2524, 5001–5019.
4 Description of the material
The determination of the taxonomic position (superfamilies and families) of the corals is principally based on the micromorphological characteristics of the septal blades. The genus assignment follows the organisational form of the coral and the species morphometric data. In its outline, the classification follows the traditional scheme that was introduced by Alloiteau (1952) and later improved by Löser (2016). The descriptions for superfamilies, families, and genera are principally based on the characteristics of the type material of the respective type species that were studied by the first author between 1990 and 2015 (and that resulted in the systematic revision published in Löser, 2016). All below given descriptions are not based on the literature because it often neither shows nor mentions important microstructural features. This is mainly because most literature published before 1960 and also many current publications do not show thin sections and do not allow to recognize important microstructural characteristics.
As for instance, the superfamily Actinastreoidea is based on the genus Actinastrea. To understand the characteristics of the superfamily the type material of Actinastrea goldfussi Orbigny, 1850, and additional topotypical material from the type locality was studied (the latter using thin sections). When several families are collected in one superfamily, the classification into families follows the name giving genera of the families. As for instance the family Agatheliidae is based on the genus Agathelia with the type species Agathelia asperella. In order to understand the constitution of the family, the type material of Agathelia asperella and topotypical material from the type area was studied (the latter by thin sections).
The following abbreviations are used in describing the dimensions of the corals: ccd, distance between corallite centres; cdw, distance between corallite centres within corallite series; clmax, large lumen; clmin, small lumen; cmax, larger outer corallite diameter; cmin, smaller outer corallite diameter; crd, distance of corallite series; septa, number of septa in the adult corallite.
Order Scleractinia Bourne, 1900.
Remarks
A general classification system that covers all Scleractinian corals (Triassic-today) and is valid for Mesozoic, Cenozoic, as well as extant coral is not available. For Mesozoic and Cenozoic/extant corals, different classification systems are still applied. The classification system of the order Scleractinia since Vaughan and Wells (1943) and Alloiteau (1952) is the application of suborders and families. The use of suborders is not practical for various reasons. Firstly, not all suborders are well-defined and/or limited to a relatively small group of genera. Secondly, when suborders are applied strictly, many families would remain without a suborder. These families would require the creation of new suborders. The creation of new taxa of higher levels always increases noise and may cause confusion. To avoid this scenario, a preliminary classification system was introduced by Löser (2016), which does not apply suborders, but rather superfamilies that group families together. Practically, suborders are for the moment replaced by superfamilies. Superfamilies have the advantage that they are covered by the rules of nomenclature, hence a superfamily has always a reference to a genus. This is not the case in suborders. A comparison of the former classification system using suborders and the new system is published in Löser (2016) and Löser et al. (2018). The concept of superfamilies was followed by Garberoglio et al., (2020, 2021) and Kolodziej (2020). Baron-Szabo (2021) only applies families but does not apply suborders. The relationship between former suborders and superfamilies is shown in Löser (2016) and Löser et al. (2018).
Ranges of genera, families, and superfamilies (as given in Löser, 2016) are principally based on material that was personally examined and that come from properly dated strata. Material without an exact stratigraphic control was not included. Of course, these ranges may change when new material is observed. Mesozoic corals have a low biostratigraphic value because the ranges of most species and genera are long. But at least they allow to apply probabilistic estimations based on associations (Löser, 2002, 2021).
Superfamily Actinastreoidea Alloiteau, 1952
Description Cerioid, phaceloid or plocoid colonies. Septa compact, mostly in a regular radial symmetry. Septa often connected to each other. Lateral faces with thorns, upper margin granulated. Microstructure of septa of medium-sized trabeculae. Lonsdaleoid septa present in one genus, main septa absent. Synapticulae absent. Pali in some genera present, columella in most genera. Endotheca varies, generally made of thin tabulae. Wall compact or subcompact, mainly septothecal. Marginarium in one genus. Coenosteum generally present, consists of isolated trabeculae or costae. Budding extracalicinal and intracalicinal.
Family Actinastreidae Alloiteau, 1952
Remarks
The family was formerly assigned to the suborder Archeocaeniina Alloiteau, 1952. The suborder has no reference to any family or genus, and should encompass corals with septa made of a few ‘simple’ trabeculae without a divergence system. The suborder cannot be identical with the suborder Astrocoeniina Vaughan & Wells, 1943 that is based on the Eocene genus Astrocoenia, which for a long period of time was poorly known. The material of Astrocoenia that is illustrated by Vaughan and Wells (plates 5 and 14) belongs to Actinastrea or Stelidioseris and corresponds to the definition of the suborder. The name-giving genus Astrocoenia has, in contrast, very small trabeculae (Löser, 2016: 186, fig. A55). The suborder Astrocoeniina is still applied to Cenozoic corals, but in a questionable sense.
Stelidioseris Tomes, 1893
Type species—Stelidioseris gibbosa Tomes, 1893.
Description Plocoid colony with a very narrow coenosteum that is made up of confluent costae. Corallites circular or polygonal. Septa in a regular radial symmetry. Septa of the first and second generation can be connected to the columella, younger septa can be connected to the older septa. Septa of the second generation may be centrally swollen. Columella styliform and large. No pali.
Stelidioseris retifera (Stoliczka, 1873).
Figure 5

Stelidioseris retifera (Stoliczka, 1873), DCTUC CRET 5003, coral surface, scale bar 1 mm
Material DCTUC CRET 5003.
Dimensions
(CRET 5003)
n | min–max | µ | s | cv | µ ± s | |
|---|---|---|---|---|---|---|
clmin | 25 | 1.64–2.36 | 1.97 | 0.19 | 10.0 | 1.77–2.17 |
clmax | 25 | 2.04–3.01 | 2.45 | 0.20 | 8.2 | 2.25–2.66 |
ccd | 27 | 1.79–2.61 | 2.21 | 0.19 | 8.7 | 2.02–2.41 |
septa | 24 |
Remarks
The species has a worldwide distribution between the Barremian and Coniacian.
Occurrence Upper lower Turonian (Nodosoides zone) of Portugal (Coimbra) Figueira da Foz, Casal dos Touros, Salmanha/M.
Superfamily Cladocoroidea Orbigny, 1851
Description Solitary and colonial (cerioid, phaceloid, plocoid) corals. Septa compact. Septal in regular thickness. Septal symmetry radial, regular or sub-regular. Septa often connected to each other. Septal lateral faces with thorns and/or granulae, septal upper margin with fine granulations. Lonsdaleoid septa and main septum absent. Microstructure of medium-sized trabeculae. Synapticulae absent. Pali present in some genera. Columella present in most genera, styliform, lamellar, parietal or by septal fusion. Endotheca generally present. Marginarium absent. Wall present, septothecal or parathecal. Coenosteum present in plocoid genera. Budding extracalicinal (cerioid, plocoid) or intracalicinal (phaceloid).
Family Cladocoridae Orbigny, 1851
Description Phaceloid colonies. Septal symmetry sub-regular radial. Pali present in some genera. Columella styliform, lamellar, or parietal.
Remarks
Most genera of the family are poorly documented due to the small size of the type specimens or the lack of thin sections. The family was formerly assigned to the suborder Faviina. As shown in Garberoglio et al. (2021), the septal microstructure of the name giving genus Favia differs from the Cretaceous families assigned to the suborder Faviina.
Procladocora Alloiteau, 1952
Type species—Calamophyllia gracilis Orbigny, 1850.
Description Phaceloid colony. Corallites with a small diameter (< 6 mm). Septa in a irregular radial symmetry; systems can be recognised. The number of septa increases with the corallite diameter. The septa of the first two cycles have pali. The columella is lamellar.
Procladocora arnaudi (Alloiteau, 1957).
Figure 6a, b

a, b Procladocora arnaudi (Alloiteau, 1957), DCTUC CRET 5000, a transversal thin section, b transversal thin section, detail, scale bar 1 mm. c–e Astraeofungia cenomana (Orbigny, 1850), DCTUC CRET 5018, a transversal thin section, d transversal thin section, detail, e longitudinal thin section, scale bar 1 mm. f–h Dimorphastrea cf. duparci (Koby, 1898), DCTUC CRET 5005, f transversal thin section, g transversal thin section, detail, h longitudinal thin section, scale bar 1 mm. i–k Dimorphastrea fallax Becker, 1875, DCTUC CRET 5007, k transversal thin section, j transversal thin section, detail, k longitudinal thin section, scale bar 1 mm
Material DCTUC CRET 5000; 2 thin sections.
Dimensions
(CRET 5000)
n | min–max | µ | s | cv | µ ± s | |
|---|---|---|---|---|---|---|
cmin | 7 | 3.59–6.20 | 4.79 | 0.78 | 16.4 | 4.00–5.58 |
cmax | 6 | 3.89–8.32 | 5.80 | 1.44 | 24.8 | 4.36–7.24 |
clmin | 7 | 2.46–4.79 | 3.86 | 0.74 | 19.2 | 3.12–4.61 |
clmax | 6 | 2.67–7.11 | 4.81 | 1.41 | 29.4 | 3.40–6.23 |
septa | 4 | 24–34 | 26.50 | 5.0 | 18.8 | 22–32 |
Remarks P. arnaudi to date is only known from the Maastrichtian of France.
Occurrence Upper Cenomanian (Juddii zone) of Portugal (Coimbra) Figueira da Foz, Casal dos Touros, Salmanha/H1.
Superfamily Cyclolitoidea Milne Edwards & Haime, 1849
Description Solitary and colonial corals. Septa with regular perforations, with perforations only in certain parts of the septa, or almost compact. Septa generally thick, without notable symmetry (except Negoporitidae). Septa often connected to each other. Septal lateral faces with pennulae and thorns. Septal distal margin with large granulae. Lonsdaleoid septa and main septum absent. Microstructure of large trabeculae. Synapticulae present. Pali in some genera probably present but difficult to distinguish from the perforated inner margins of the septa. Columella poorly defined. Endotheca present or absent. When present, mostly thin tabulae. Marginarium absent. Wall poorly defined. Coenosteum varies depending on the organisation type. Budding varies.
Remarks
The superfamily is a very important element in all Cretaceous faunas. The families are numerous in genera and some of their genera are very common. The superfamily continues at least into the Eocene.
Family Latomeandridae Fromentel, 1861
Description The family encompasses numerous solitary and colonial coral genera that show septa with perforations that are concentrated in the inner margin of the septa. The septa are thinner than in the Synastraeidae and less perforated than the Microsolenidae. The thickness of septa and the space between them is equilibrated.
Remarks
For this group of genera, formerly the family name Leptophylliidae was applied (e.g., Löser, 2016). Since Leptophyllia was found to belong to the Synastraeidae (see for instance Löser et al., 2019), the family name can no longer be applied. For the moment, we decided to reinstall the Latomeandridae. The family is not well defined since the type material of the type species of Latomeandra is silicified. The family was formerly assigned to the suborder Microsolenina Morycowa and Roniewicz, 1995. The suborder encompasses families that were originally assigned to the Fungiina suborder. The main characteristic is the presense of pennulae, balkon-like formations related to a rhythmical growth that are visible on the septal lateral faces. The genus Microsolena itself is poorly defined; the type material of the type species is not available. The genus, family, and suborder are conceptual.
Astraeofungia Alloiteau, 1952
Type species—Astrea decipiens Michelin, 1841.
Description Thamnasterioid colony, with mostly only at the inner margin perforated septa, that distinguish hardly in length and thickness. The corallites are regularly distributed, and generally not in rows as in Dimorphastrea. Costae run between all corallites.
Astraeofungia cenomana (Orbigny, 1850).
Figure 6c–e
Material DCTUC CRET 5018; 2 thin sections.
Dimensions
(CRET 5018)
n | min–max | µ | s | cv | µ ± s | |
|---|---|---|---|---|---|---|
ccd | 7 | 3.76–4.94 | 4.23 | 0.45 | 10.8 | 3.77–4.69 |
septa | 4 | 25–28 | 26.50 | 1.73 | 6.5 | 25–28 |
Remarks A. cenomana is a very common species. It has a worldwide distribution between the Aptian and Cenomanian.
Occurrence Early Turonian (Nodosoides zone) of Portugal (Coimbra) Nazaré, beach section/2.
Dimorphastrea Orbigny, 1850
Type species—Dimorphastrea grandiflora Orbigny, 1850.
Description Thamnasterioid colony with corallites arranged in concentric rows. The colony surface is plane, the corallite centres may be slightly depressed. Septa connect more between corallites of neighboured rows than with corallites of the same row. Dimorphastrea is similar to Astraeofungia and juvenile colonies of Astraeofungia and Dimorphastrea are difficult to distinguish.
Dimorphastrea cf. duparci (Koby, 1898).
Figure 6f–h
Material DCTUC CRET 5005; 2 thin sections.
Dimensions
(CRET 5005)
n | min–max | µ | s | cv | µ ± s | |
|---|---|---|---|---|---|---|
crd | 3 | 14.2–17.6 | 15.5 | 1.80 | 11.6 | 13.7–17.3 |
cdw | 4 | 4.54–11.5 | 8.42 | 2.88 | 34.2 | 5.53–11.3 |
septa | 5 | 33–48 | 41.4 | 5.89 | 14.2 | 36–47 |
Remarks
The present material differs from D. duparci by slightly larger distances between the corallite rows and higher septal counts. The species is known from the Kimmeridgian to Cenomanian of the Tethys.
Occurrence Upper Cenomanian (Geslinianum zone) of Portugal (Coimbra) Leiria, Carrasqueira/F.
Dimorphastrea fallax Becker, 1875
Figure 6i–k
Material DCTUC 2517, CRET 5004, CRET 5007; 6 thin sections.
Dimensions
(CRET 5007)
n | min–max | µ | s | cv | µ ± s | |
|---|---|---|---|---|---|---|
crd | 8 | 9.23–11.9 | 10.82 | 0.99 | 9.1 | 9.83–11.8 |
cdw | 13 | 4.34–9.08 | 6.39 | 1.65 | 25.9 | 4.73–8.05 |
septa | 10 | 32–53 | 41.40 | 7.10 | 17.1 | 34–49 |
Remarks
The species occurs between the Kimmeridgian and Turonian of the Western and Central Tethys, in Boreal regions, and in the Western Atlantic.
Occurrence Upper Cenomanian (Geslinianum zone) of Portugal (Coimbra) Leiria, Carrasqueira/F.
Dimorphastrea cf. lorioli Koby, 1898
Figure 7a–c

a–c Dimorphastrea cf. lorioli Koby, 1898, DCTUC 2500, a transversal thin section, b transversal thin section, detail, c longitudinal thin section, scale bar 1 mm. d–f Eocomoseris sp., DCTUC CRET 5006, d transversal thin section, e transversal thin section, detail, f longitudinal thin section, scale bar 1 mm. g–i Negoporites quartus Eliášová, 1995, DCTUC CRET 5008, g transversal thin section, h transversal thin section, detail, i longitudinal thin section, scale bar 1 mm. j–l Actinacis ? remesi Felix, 1903a, DCTUC CRET 5016, j transversal thin section, k transversal thin section, detail, l longitudinal thin section, scale bar 1 mm
Material DCTUC 2500, 2504, CRET 5009, CRET 5010; 8 thin sections.
Dimensions
(CRET 2500)
n | min–max | µ | s | cv | µ ± s | |
|---|---|---|---|---|---|---|
crd | 15 | 7.04–11.0 | 9.15 | 1.22 | 13.3 | 7.93–10.4 |
cdw | 15 | 2.79–9.29 | 5.50 | 1.71 | 31.1 | 3.78–7.21 |
septa | 15 | 26–40 | 31.40 | 4.48 | 14.2 | 27–36 |
Remarks
The present material differs from D. lorioli by higher septal counts. The species is known from the Hauterivian and Cenomanian of France based on the study of unpublished material observed by the first author.
Occurrence Upper Cenomanian (Geslinianum zone) of Portugal (Coimbra) Leiria, Carrasqueira/F.
Family Microsolenidae Koby, 1889
Description Solitary and colonial (cerioid, hydnophoroid, meandroid, phaceloid, plocoid, thamnasterioid) colonies. Septa completely and regularly perforated. Interseptal space larger than or equal to septal thickness.
Remarks
The family was formerly assigned to the suborder Microsolenina (see above).
Eocomoseris Melnikova et al., 1993
Type species—Eocomoseris gurumdyensis Roniewicz, 2011 nom. nov. pro Eocomoseris ramosa Melnikova et al., 1993 non Heptastylopsis ramosus Frech, 1890.
Description Astreoid colony with small corallites. Symmetry of septa irregular, but two size orders can be distinguished. Number of septa up to 30. Septa occasionally connected to each other. Costae sub-confluent to non-confluent. Synapticulae fairly common. Columella styliform. Endotheca and wall absent. Coenosteum narrow.
Eocomoseris sp.
Figure 7d–f
Material DCTUC CRET 5006, CRET 5011; 4 thin sections.
Dimensions
(CRET 5006)
n | min–max | µ | s | cv | µ ± s | |
|---|---|---|---|---|---|---|
ccd | 20 | 1.89–2.68 | 2.25 | 0.23 | 10.5 | 2.01–2.48 |
cmin | 10 | 1.73–2.11 | 1.92 | 0.11 | 5.8 | 1.81–2.03 |
septa | 12 | 17–23 | 19.16 | 1.89 | 9.9 | 17–21 |
Remarks
The genus Eocomoseris was originally only described with one Cretaceous species. Later revisions of type material revealed that another two formerly described species are in existence. The distinction of species is difficult because the principal characteristic—the distance of the corallite centres—varies within one colony. In other genera of the family (Microsolena, Dimorpharaea) the distance of corallites presents a lower variability (see as for instance Pandey et al., 1999) and hence the corallite distance is more suitable for the distinction of species. The material presented here is not well enough preserved to establish a new taxon. Material that coincides in the corallite diameter and septal counts with the present specimens occurs worldwide from the Valanginian to Cenomanian.
Occurrence Upper Cenomanian (Geslinianum zone) of Portugal (Coimbra) Leiria, Carrasqueira/F.
Family Negoporitidae Eliášová, 1995
Description Colonial (plocoid) corals. Septa with perforations. Septal symmetry bilateral. Septa connected to each other. Pali and columella present. Endotheca with tabulae. Wall incomplete, synapticulothecate. Coenosteum extended, with isolated trabeculae. Budding extracalicinal.
Remarks
The family was formerly assigned to the suborder Microsolenina (see above).
Negoporites Eliášová, 1989
Type species—Porites michelini Reuss, 1846.
Description Plocoid colony. Corallite outline circular, corallites small (generally below 3 mm in diameter). Septa irregularly perforated. Symmetry of septa bilateral. Septal cycles differ in length, but hardly at all in thickness. Septa of younger cycles often and regularly connected to the septa of preceding cycles. Not well-separated pali irregularly present. Costae non-confluent. Synapticulae fairly common, mainly in the wall. Columella small, substyliform. Endotheca consists of numerous thin tabulae. Wall subcompact, synapticulothecal. Coenosteum medium broad, consists of trabeculae and tabulae.
Negoporites quartus Eliášová, 1995
Figure 7g–i
Material DCTUC CRET 5008; 2 thin sections.
Dimensions
(CRET 5008)
n | min–max | µ | s | cv | µ ± s | |
|---|---|---|---|---|---|---|
cmin | 12 | 2.19–2.80 | 2.45 | 0.18 | 7.4 | 2.26–2.63 |
cmax | 12 | 2.50–3.30 | 2.93 | 0.21 | 7.3 | 2.71–3.14 |
ccd | 13 | 3.42–6.31 | 4.67 | 0.89 | 19.2 | 3.77–5.56 |
septa | 10 | 28–36 | 32.90 | 2.51 | 7.6 | 30–35 |
Remarks
The genus Negoporites is restricted to the Cenomanian. The species N. quartus to date is only known from the Cenomanian of the European Boreal.
Occurrence Upper Cenomanian (Geslinianum zone) of Portugal (Coimbra) Leiria, Carrasqueira/F.
Superfamily Madreporoidea Dana, 1846
Description Colonial (phaceloid, plocoid) corals. Septa compact, very thin, in an irregular radial symmetry. Septa often connected to each other. Lateral faces with fine thorns, upper margin unknown. Lonsdaleoid and main septa absent. Microstructure of septa of small trabeculae. Synapticulae absent. Pali absent, columella present in most genera, lamellar or by septal fusion. Endotheca present. Marginarium absent. Wall thick, lamellar. Coenosteum made of lamellae. Budding extracalicinal and intracalicinal.
Family Madreporidae Dana, 1846
Palaeohelia Alloiteau, 1958
Type species—Palaeohelia collignoni Alloiteau, 1958.
Description Phaceloid colony with thin branches. Septa without symmetry. A weak columella is formed by the fusion of septa in the corallite centre.
Remarks
The family was formerly assigned to the suborder Meandrinina Alloiteau, 1952. The definition of this suborder was rather conceptual when it was established. It should gather genera with compact septa and smooth lateral septal faces and smooth septal upper margins. No data were provided on the septal microstructure. Subsequently, a large amount of material with very small trabeculae was assigned to this suborder. Indeed, the name-giving genus Meandrina Lamarck, 1801 shows septal ornamentation as described above, but it possesses rather large trabeculae (Löser, 2016: Fig. 3.3.1.3A). So, the family Madreporidae cannot remain here.
Palaeohelia sp.
Figure 8a–c

a–c Palaeohelia sp., DCTUC 2524, a transversal thin section, b transversal thin section, detail, c longitudinal thin section, scale bar 1 mm. d–f ?Barysmilia sp., DCTUC CRET 5002, d transversal thin section, e transversal thin section, detail, f longitudinal thin section, scale bar 1 mm
Material DCTUC 2524; 4 thin sections.
Dimensions
(CRET 2524)
n | min–max | µ | s | cv | µ ± s | |
|---|---|---|---|---|---|---|
cmin | 20 | 1.56–2.71 | 2.14 | 0.37 | 17.2 | 1.77–2.51 |
cmax | 20 | 2.34–3.48 | 2.88 | 0.29 | 10.3 | 2.58–3.17 |
clmin | 25 | 1.03–1.47 | 1.20 | 0.13 | 11.2 | 1.07–1.34 |
clmax | 25 | 1.06–2.02 | 1.56 | 0.26 | 16.8 | 1.30–1.83 |
septa | 24 |
Remarks
Palaeohelia received little attention since its creation. Four species are known: P. albiensis Beauvais, 1982, P. collignoni Alloiteau, 1958, P. dartoni (Wells, 1933), and P. graeca (Baron-Szabo & Steuber, 1996). There exists more undescribed material. The present specimen belongs with certainty to a new species, that is only known from the Cenomanian of Portugal. In its morphology it does not differ from other species of the genus, except for its corallite dimensions. These are very small compared to the other species. The material described by Löser et al. (2019) as Palaeohelia sp. from the Upper Cretaceous of Austria has even smaller dimensions.
Occurrence Upper Cenomanian (Juddii zone) of Portugal (Coimbra) Leiria, Souto de Cima/H.
Superfamily Phyllosmilioidea Felix, 1903b
Description Solitary and colonial (astreoid, flabelloid, meandroid, phaceloid, plocoid) corals. Septa compact. First septal generation thicker than all others. Symmetry irregular or subregular radial. Septa in some genera connected to each other. Septal distal margins smooth, lateral faces with fine thorns, inner margins often swollen or T-shaped. Lonsdaleoid septa very rare. Main septum absent. Microstructure of very small trabeculae, only marked by a dark line. The costae are made by medium-sized trabeculae that result in granulated costal surfaces. Synapticulae and pali absent. Columella present or absent. Endotheca mostly present. Marginarium absent. Wall generally present, septothecal. Coenosteum present in some genera. Budding varies.
Family Phyllosmiliidae Felix, 1903b
Remarks
The family was formerly assigned to the suborder Meandrinina Alloiteau, 1952 (see above for discussion).
Barysmilia Milne Edwards & Haime, 1848
Type species—Dendrophyllia brevicaulis Michelin, 1841.
Description Plocoid colony with elliptical corallites, often as very short corallite rows. The septal symmetry is irregularly radial. The coenosteum is present, often very narrow.
?Barysmilia sp.
Figure 8d–f
Material DCTUC CRET 5001, CRET 5002, CRET 5013, CRET 5015; 6 thin sections.
Dimensions
(CRET 5002)
clmin | 7.4–9.5 |
|---|---|
clmax | 8.3–11.2 |
septa | 48 |
Description Plocoid colony. Corallite outline circular. Septa compact, in cross section tapering towards the centre. Septal maximum thickness 300 µm. Septal microstructure of small trabeculae. Symmetry of septa radial and irregularly hexameral with subregular cycles of septa. Septal cycles differ in length and thickness. First two septal cycles extend to the corallite centre, later cycles are shorter. Younger septa are regularly connected to older septa. Septa of the first two cycles irregularly connected in the corallite centre. Septal distal margin unknown, lateral face with few thorns, inner margin swollen in places. Costae very short, non-confluent or sub-confluent. Synapticulae absent. Columella probably lamellar but difficult to distinguish from the inner margins of the septa. Endotheca consists of numerous tabulae. Wall compact, parathecal. Coenosteum narrow. Budding extracalicinal.
Remarks
This material very probably represents a new genus, but the material is too poorly preserved to be formally established as a new taxon. It differs from Barysmilia by the formation of the wall (parathecal in place of septothecal), the columella (parietal in place of lamellar) and the formation of the first septal cycles (normal thickness in place of thickened). Even the position within the family Phyllosmiliidae is questionable. The material is also similar to members of the Placocoeniidae family but differs clearly in its septal microstructure.
Occurrence Upper lower Turonian (Nodosoides zone) of Portugal (Coimbra) Figueira da Foz, Casal dos Touros, Salmanha/M; Nazaré, beach section/1; Nazaré, beach section/2.
Superfamily Poritoidea Gray, 1842
Description Colonial (astreoid, plocoid) corals. Septa irregularly perforated. Symmetry of septa irregular radial or bilateral. Septa connected to each other. Septal distal margin coarsely dentated, lateral face with rare granulae, inner margin slightly swollen in places. Lonsdaleoid septa and main septum absent. Microstructure of medium-sized trabeculae. Synapticulae abundant. Pali present in some genera. Columella and endotheca present. Marginarium absent. Wall varies. Coenosteum generally present. Budding extracalicinal.
Family Actinacididae Vaughan & Wells, 1943
Description Astreoid colonies. Septa in a regular radial or bilateral septal symmetry.
Remarks
The family was formerly assigned to the suborder Fungiina Verrill, 1870. The suborder was applied by Alloiteau (1952) and subsequent authors in a very broad sense for any coral with perforate septa and/or synapticulae (except the suborder Dendrophylliina). The suborder collects families that vary considerably in their septal microstructure and has a very informal character. Fungiina sensu stricto are corals with fulturae (see Gill, 1981) (e.g., only the Fungiidae, and perhaps the Asteroseriidae families). All other families cannot remain in this suborder. Some families have already been separated into the suborder Microsolenina.
Actinacis Orbigny, 1849
Type species—Actinacis martiniana Orbigny, 1849.
Description Astreoid colony. The corallites are small, elliptical, well marked and with large distances to each other. Septal symmetry is between radial and bilateral. Septa often and regularly connected to each other. Pali present but because of the septal perforations not well visible.
Actinacis ? remesi Felix, 1903a
Figure 7j–l
Material DCTUC CRET 5012, CRET 5014, CRET 5016, CRET 5017; 7 thin sections.
Dimensions
(CRET 5016)
n | min–max | µ | s | cv | µ ± s | |
|---|---|---|---|---|---|---|
clmin | 15 | 1.62–2.26 | 1.90 | 0.19 | 10.1 | 1.71–2.09 |
clmax | 15 | 2.07–2.83 | 2.38 | 0.26 | 11.2 | 2.11–2.65 |
ccd | 20 | 2.50–4.24 | 3.17 | 0.49 | 15.5 | 2.67–3.66 |
septa | 12 | 24–30 | 27.33 | 2.01 | 7.3 | 25–29 |
Remarks
The material can clearly be assigned to the genus Actinacis, but could not be assigned to an existing species with certainty. Closely related is material labelled as Actinacis vignyensis Alloiteau, 1957 (MNHN; but never formally described) and material labelled as Actinacis remesi Felix, 1903a (NHMW, coll. Trauth; PU, coll. Zuffardi-Comerci) from the Eocene of Klokocov (Moravia, Czech Republic) but the type of Actinacis remesi itself is unknown. The unnamed species is distributed worldwide between the Upper Albian and Eocene.
Occurrence Upper lower Turonian (Nodosoides zone) of Portugal (Coimbra) Nazaré, beach section/1; Nazaré, beach section/2.
5 Discussion
The Cenomanian/Turonian (C/T) boundary was a prominent event in the Cretaceous Period. Oceanic Anoxic Event II and a global sea level highstand had an impact on coral diversity and evolution. At the Cenomanian/Turonian boundary, 46 coral genera disappeared (Fig. 9). This number is constituted by genera that occurred at least in the late Cenomanian but did not pass the C/T boundary. Genera that had their last occurrence during the early or middle Cenomanian are not counted. Sixty-six genera survived the C/T boundary, but 11 of them did not reach the middle Coniacian. Another 59 genera evolved after the C/T boundary, but six of them occurred only during the Turonian. At the base of the late Cenomanian 110 genera occurred. The richness of genera almost recovered in the Coniacian and reached 115 genera in the late Santonian. The reduction of taxa at the C/T boundary is the same predicament that occurred at the Cretaceous/Paleogene boundary (Löser, 2016: 123). The loss of taxa during the Turonian is not only related to the sea level fall during the same period that reduced the available shallow marine area for colonisation; the low number of lower to middle Turonian coral localities to any extent (Löser, 2016: 119) is also caused by the regression during the Turonian that eroded the faunas deposited from the C/T boundary on. It is noteworthy that among all of these genera are numerous examples that are endemic, monospecific and/or poorly known.

Coral genera with their stratigraphic range between the upper Albian to Coniacian. Genera that became extinct before the upper Cenomanian are not shown. Genera endemic in one faunule in red. E, genera that became extinct during the upper Cenomanian; se, genera that survived the CT boundary but hardly reached the Coniacian; S, genera that survived the C/T boundary; T, exclusive Turonian genera; U, newly evolved, post-Cenomanian genera. Range data from Löser (2016). Only genera for which a range could be obtained are included
5.1 Species richness
When outcrops with shallow marine corals are sampled thoroughly, they always yield large faunas. Particularly for Cenomanian faunas this has been shown in multiples occasions for Boreal (Bohemian Basis, Eliášová, 1997; Westphalian Basin, Löser, 1994; Saxonian Basin, Löser, 2014, 2015) and Tethyan faunas (Penninic Ocean, Löser et al., 2013; Quillan Basin, Löser & Bilotte, 2017; West Pelagonian zone, Löser et al., 2018). For each of the three Boreal faunas nearly sixty species were found, and for the three Tethyan faunas values between 16 and 40 species are reported.
The 11 coral species described here are distributed among six localities or levels. With the exception of the Carrasqueira locality with five species, the localities have between one and three species. Since the report is not based on collection material but rather on systematic field studies, complete sampling was achieved. Compared to other shallow marine coral faunas, the number of species is very low and conditions seemed to be unfavourable for corals. The level that was richest in species is the Carrasqueira locality (Leiria) with an association of platy corals that are exclusively made up by pennular corals thus supposing low-level light conditions. A comparable situation was described from the upper Cenomanian of southern France (Löser & Bilotte, 2017) with 16 coral species. The Carrasqueira locality is the only place to have yielded rich coral material per number of specimens; in all other outcrops, corals have been searched for.
The level H1 in the Salmanha section yielded only one species, at the same level as the Souto de Cima outcrop (Leiria). Both species are phaceloid, inferring low water energy and probably also a slightly deeper environment. The lower Turonian localities at the Nazaré beach yielded a total of three species. The low number is due to the low concentration of corals and their very poor state of conservation. The lower Turonian level M in the Salmanha section yielded only two species. Compared to other coral localities of a comparable age, this number is very low.
5.2 Palaeoecology
The study of several Portuguese coral faunas has shown that even profound sampling cannot yield numerous species when the environment was not favourable for corals. Unfavourable conditions are mostly related to temperature or salinity. So low temperature has inhibited coral growth in the Western Pacific from the Albian on (Iba & Sano, 2007), and low salinity has inhibited coral growth in the Western Atlantic between the late Cenomanian and the Santonian because of the opening of the Western Interior Seaway (Hay, 2008; Löser, 2013a). Nevertheless, for the case of the Cenomanian–Turonian WPCT the surface water temperatures should have been sufficiently high to support benthic communities with corals, along with many examples of Tethyan invertebrates. Salinity variations and high turbidity related to the influx of fine-grained siliciclastics could have been much more effective in explaining the rather low diversity values, and coral growth inhibition. This can be partly explained by the palaeogeographic setting of the carbonate platform during the late Cenomanian and early Turonian, when its inner shelf ranges of Figueira da Foz, Leiria, and Nazaré were bordered by a depositional system of siliciclastic alluvial plains, which provided fine-grained micaceous sediments to the shallow platform. Besides progradation during regressive intervals, the fluvial drainage to the platform and significant volumes of sediment transported in suspension were certainly responsible for frequent episodes of salinity decrease and higher turbidity, which would have affected coral diversity and growth.
The presence of corals does not automatically imply the existence of reefs. Corals can construct reefs when the conditions are favourable. The Cretaceous was characterised by high temperatures, on the one hand (Forster et al., 2007), and a seawater chemistry that was unfavourable for the scleractinian corals which build their skeletons from aragonite, on the other hand (Stanley & Hardie, 1998). Throughout the Cretaceous, coral colonies remained small, rarely reaching dimensions beyond 20 cm. There are very few exceptions (Bover Arnal et al., 2012). Coral reefs—here understood as rigid biogenic constructions that are clearly elevated above the sea floor—are rarely reported from the Cretaceous (Bonilla González et al., 2018; Fernández Mendiola et al., 2015). Most of these rare reefs are built up by phaceloid corals of the genus Actinastreopsis (Löser, 2013a). Therefore, we cannot discuss coral reef formations in the Cretaceous of Portugal.
5.3 Distribution
The low number of species makes a comparison to other localities very difficult. The species from the study area all have long ranges (Fig. 10) with slightly more occurrences in the Albian and Cenomanian, and less in the Turonian to Maastrichtian. The palaeobiogeographic comparison with other areas does not show many results because of the paucity of data (Fig. 11). Species of the study area were found in boreal regions, such as the Bohemian-Saxonian Basin (Eliášová, 1997; Löser, 2014), and in two other areas of the Iberian Peninsula and the Quillan Basin (Löser & Bilotte, 2017; Löser et al., 2013).

Stratigraphic distribution of species found in the study area. The thickness of bars corresponds to the number of localities. The material of the present study is not shown but the age of the study area is marked in grey

Correlation of faunules with the studied faunas. Only the time period Albian to Coniacian was analysed, and only faunules with more than one species. The Correlation Ratio coefficient was applied. The Prebetic zone corresponds geographically to SE Spain, the Quillan Basin principally to SW France
6 Conclusion
Shallow marine strata of the West Portuguese Carbonate Platform of the upper Cenomanian to lower Turonian have yielded coral associations from six sample points with a surprisingly low number of exclusively hermatypic corals. The number of species varies between one and five per sample point, even though profound searching was carried out. The rarity of corals in the strata and the resulting low number of species is explained by a certain variation in salinity and a high water turbidity. This situation compares well to the Western Interior Seaway where coral growth was inhibited by lower temperatures and a lower salinity. Its occurrence stopped during the middle Cenomanian and returned in the Campanian.
References
Alloiteau, J. (1952). Embranchement des coelentérés. In J. Piveteau (Ed.), Traité de Paléontologie (pp. 376–684). Masson.
Alloiteau, J. (1957). Contribution à la systématique des Madréporaires fossiles. Centre National de la Recherche Scientifique.
Alloiteau, J. (1958). Monographie des Madréporaires fossiles de Madagascar. Annales Géologiques de Madagascar, 25, 1–218.
Azerêdo, A. C., Duarte, L. V., Henriques, M. H., & Manuppella, G. (2003). Da dinâmica continental no Triásico aos mares do Jurássico Inferior e Médio. Lisboa: Instituto Geológico e Mineiro.
Baron-Szabo, R. C. (2021). Upper Barremian–lower Aptian scleractinian corals of central Europe (Schrattenkalk Fm., Helvetic Zone, Austria, Germany, Switzerland). Zootaxa, 4960(1), 1–199. https://doi.org/10.11646/zootaxa.4960.1.1
Baron-Szabo, R. C., & Steuber, T. (1996). Korallen und Rudisten aus dem Apt im tertiären Flysch des Parnass-Gebirges bei Delphi-Arachowa. Berliner Geowissenschaftliche Abhandlungen, 18, 3–75 (E).
Barroso-Barcenilla, F., Callapez, P. M., Ferreira Soares, A., & Segura, M. (2011). Cephalopod assemblages and depositional sequences from the upper Cenomanian and lower Turonian of the Iberian Peninsula (Spain and Portugal). Journal of Iberian Geology, 37, 9–28. https://doi.org/10.5209/rev_JIGE.2011.v37.n1.1
Beauvais, L. (1982). Révision du genre Palaeohelia Alloiteau (Scleractiniaire méso-crétacé). Eclogae Geologicae Helvetiae, 75(3), 669–687.
Becker, E. (1875). Die Korallen der Nattheimer Schichten (1). Palaeontographica, 21, 1–60.
Berthou, P. Y. (1984). Albian-Turonian stage boundaries and subdivisions in the Western Portuguese Basin, with special emphasis on the Cenomanian-Turonian boundary in the Ammonite Facies and Rudist Facies. Bulletin of the Geological Society of Denmark, 33, 41–45.
Bonilla González, O. A., López-Horgue, M. A., Löser, H., & Fernández Mendiola, P. A. (2018). Un ejemplo clave en la evolución de los corales: los arrecifes del Aptiense-Albiense (Cretácico Inferior) de la Cuenca Vasco-Cantábrica. In A. Badiola, A. Gómez-Olivencia, & X. Pereda Suberiola (Eds.), Registro fósil y patrimonio de los Pirineos occidentales: una visión desde la Geodiversidad. (pp. 81–83). Vitoria Gasteiz: Servicio Central de Publicaciones del Gobierno Vasco.
Bover Arnal, T., Löser, H., Moreno Bedmar, J. A., Salas, R., & Strasser, A. (2012). Corals on the slope (Aptian, Maestrat Basin, Spain). Cretaceous Research, 37, 43–64.
Callapez, P. M. (1998). Estratigrafia e Paleobiologia do Cenomaniano-Turoniano. O significado do eixo da Nazaré-Leiria-Pombal. PhD Thesis. University of Coimbra.
Callapez, P. M. (2004). The Cenomanian-Turonian central West Portuguese carbonate platform. In J. L. Dinis & P. P. Cunha (Eds.), Cretaceous and Cenozoic events in West Iberia margins. 23rd IAS Meeting of Sedimentology, field trip 2 guidebook (pp. 39–51). International Association of Sedimentologists.
Callapez, P. M. (2008). Palaeobiogeographic evolution and marine faunas of the Mid-Cretaceous Western Portuguese Carbonate Platform. Thalassas, 24, 29–52.
Dana, J. D. (1846). Structure and classification of Zoophytes. United States Exploring Expedition during the years 1838, 1839, 1840, 1841, 1842 under the command of Charles Wilkes, U.S.N. Philadelphia: Lea & Blanchard.
Dinis, J. L., Rey, J., Cunha, P. P., Callapez, P. M., & Reis, R. P. (2008). Stratigraphy and allogenic controls on the western Portugal Cretaceous, an updated synthesis. Cretaceous Research, 29, 772–780. https://doi.org/10.1016/j.cretres.2008.05.027
Eliášová, H. (1989). Genres nouveaux des Scléractiniaires du Crétacé de la Bohême (Tchécoslovaquie). Casopis pro Mineralogii a Geologii, 34(2), 113–121.
Eliášová, H. (1995). Famille nouvelle des Scléractiniaires du Crétacé Supérieur de Bohême (Cénomanien supérieur - Turonien inférieur, République Tchèque). Vestník Ceského Geologického Ústavu, 70(3), 27–34.
Eliášová, H. (1997). Coraux crétacés de Bohême (Cénomanien supérieur; Turonien inférieur - Coniacien inférieur). République Tchèque. Vestník Ceského Geologického Ústavu, 72(3), 245–266.
Felix, J. (1903a). Verkieselte Korallen als Geschiebe im Diluvium von Schlesien und Maehren. Zentralblatt für Mineralogie, Geologie und Paläontologie, for 1903, 561–577.
Felix, J. (1903b). Studien über die korallenführenden Schichten der oberen Kreideformation in den Alpen und den Mediterrangebieten (1) Die Anthozoën der Gosauschichten in den Ostalpen. Palaeontographica, 49, 163–360.
Fernández Mendiola, P. A., Pérez-Malo, J., & García-Mondéjar, J. (2015). Stratigraphy and facies of the Early Aptian Robayera section (Cantabria, Northern Spain). Geogaceta, 55, 1–5.
Forster, A., Schouten, S., Baas, M., & Sinninghe Damsté, J. S. (2007). Mid-Cretaceous (Albian–Santonian) sea surface temperature record of the tropical Atlantic Ocean. Geology, 350, 919–922.
Frech, F. (1890). Die Korallenfauna der Trias (1:) Die Korallen der Juvavischen Triasprovinz. Palaeontographica, 37, 1–116.
Fromentel, E. (1861). Introduction à l’étude des polypiers fossiles. Mémoires de la Société d’émulation du Doubs, 5, 1–357.
Garberoglio, R. M., Löser, H., & Lazo, D. G. (2020). Early Cretaceous corals from the Agrio Formation, Neuquén Basin, west-central Argentina: Family Actinastraeidae. Cretaceous Research, 114(104503), 1–18. https://doi.org/10.1016/j.cretres.2020.104503
Garberoglio, R. M., Löser, H., & Lazo, D. G. (2021). Lower Cretaceous corals from the Agrio Formation, Neuquén Basin, west-central Argentina: Family Columastraeidae. Cretaceous Research, 124(104817), 1–19. https://doi.org/10.1016/j.cretres.2021.104817
Gili, E., & Götz, S. (2018). Part N, Revised, Volume 1, Chapter 26B: Paleoecology of Rudists. Treatise Online, 103: 1–29.
Gill, G. A. (1981). The fulturae (“compound synapticulae”), their structure and reconsideration of their systematic value. Acta Palaeontologica Polonica, 25(3/4), 301–310.
Gray, J. E. (1842). Pocilloporidae. Synopsis of the Contents of the British Museum (44th ed.). British Museum Natural History.
Haq, B. U. (2014). Cretaceous eustasy revisited. Global and Planetary Changes, 113, 44–58. https://doi.org/10.1016/j.gloplacha.2013.12.007
Hardenbol, J., Thierry, J., Farley, M. B., Jacquin, T., de Graciansky, P. C., & Vail, P. R. (1998). Mesozoic and Cenozoic sequence chronostratigraphic framework of European basins, Chart 1, Mesozoic and Cenozoic sequence chronostratigraphic chart. In P. C. Graciansky, J. Hardenbol, T., Jacquin, & P. Vail (Eds.), Mesozoic and Cenozoic Sequence Stratigraphy of European Basins. (Appendix). SEPM Special Publication 60. Tulsa: Society for Sedimentary Geology.
Hay, W. W. (2008). Evolving ideas about the Cretaceous climate and ocean circulation. Cretaceous Research, 29, 725–753. https://doi.org/10.1016/j.cretres.2008.05.025
Iba, Y., & Sano, S. I. (2007). Mid-Cretaceous step-wise demise of the carbonate platform biota in the Northwest Pacific and establishment of the North Pacific biotic province. Palaeogeography, Palaeoclimatology, Palaeoecology, 245(3/4), 462–482. https://doi.org/10.1016/j.palaeo.2006.09.008
James, N. P. (1983). Reef environment. In P. A. Scholle, D. G. Bebout, & C. H. Moore (Eds.), Carbonate depositional environments. AAPG Memoir 33. Tulsa: The American Association of Petroleum Geologists.
Koby, F. (1889). Monographie des polypiers jurassiques de la Suisse (9). Abhandlungen der Schweizerischen Paläontologischen Gesellschaft, 16, 457–586.
Koby, F. (1898). Monographie des polypiers crétacés de la Suisse (3). Abhandlungen der Schweizerischen Paläontologischen Gesellschaft, 24, 63–100.
Kolodziej, B. (2020). A new coral genus with prominent, ramified main septum (Aptian, Tanzania). Ameghiniana. Revista de la Associación Paleontológica Argentina, 57(6), 555–565. https://doi.org/10.5710/AMGH.26.06.2020.3341
Kullberg, J. C., Rocha, R. B., Soares, A. F., Rey, J., Terrinha, P., Azerêdo, A. C., Callapez, P., Duarte, L. V., Kullberg, M. C., Martins, L., Miranda, J. R., Alves, C., Mata, J., Madeira, J., Mateus, O., Moreira, M., & Nogueira, C. R. (2013). A Bacia Lusitaniana, estratigrafia, paleogeografia e tectónica. In R. Dias, A., Araújo, P., Terrinha, & J. C. Kullberg (Eds.), Geologia de Portugal, II, Geologia Meso-cenozóica de Portugal (pp. 195–347). Lisbon: Livraria Escolar Editora.
Löser, H. (1994). La faune corallienne du mont Kassenberg à Mülheim-sur-la-Ruhr (Bassin crétacé de Westphalie, Nord Ouest de l’Allemagne). Coral Research Bulletin, 3, 1–93.
Löser, H. (2002). Biostratigraphical dating of Cretaceous coral communities using large data sets. Paläontotologische Zeitschrift, 76, 75–81.
Löser, H. (2004). PaleoTax—a database program for palaeontological data. Computer & Geosciences, 30(5), 513–521. https://doi.org/10.1016/j.cageo.2004.03.009
Löser, H. (2012). Intraspecific variation in the genus Stelidioseris (family Actinastraeidae, suborder Archeocaeniina, order Scleractinia; Jurassic-Cretaceous). Geologica Belgica, 15(4), 382–387.
Löser, H. (2013a). Taxonomy and distribution of the Early Cretaceous coral genus Actinastraeopsis. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 269(2), 189–202.
Löser, H. (2013b). Revision of the family Felixaraeidae (Scleractinia; Cretaceous). Geodiversitas, 35(4), 775793. https://doi.org/10.5252/g2013n4a1
Löser, H. (2014). 3. Korallen/3. Corals. Geologica Saxonica, 60(1), 17–56.
Löser, H. (2015). Les coraux. In N. Morel (Ed.), Stratotype Cénomanien (pp. 280–282). Muséum national d’Histoire naturelle.
Löser, H. (2016). Systematic part. Catalogue of Cretaceous Corals, 4, 1–710.
Löser, H. (2021). Corals from the Early Cretaceous (?Late Valanginian - Aptian) of Puebla (Mexico): Family Solenocoeniidae. Paleontologia Mexicana, 10, 37–51.
Löser, H., & Bilotte, M. (2017). Taxonomy of a platy coral association from the Late Cenomanian of the southern Corbières (Aude, France). Annales de Paléontologie, 103, 3–17. https://doi.org/10.1016/j.annpal.2016.10.005
Löser, H., Heinrich, M., & Schuster, U. (2019). Korallen von Rußbach und Gosau (Coniac-Santon; Österreich). CPress.
Löser, H., Steuber, T., & Löser, C. (2018). Early Cenomanian coral faunas from Nea Nikopoli (Kozani, Greece; Cretaceous). Carnets de Géologie/Notebooks on Geology, 18(3), 23–121. https://doi.org/10.4267/2042/66094
Löser, H., Werner, W., & Darga, R. (2013). A middle Cenomanian coral fauna from the Northern Calcareous Alps (Bavaria, Southern Germany)—new insights into the evolution of Mid-Cretaceous corals. Zitteliana, A53, 37–76.
Melnikova, G. K., Roniewicz, E., & Löser, H. (1993). New microsolenid coral genus Eocomoseris (Scleractinia, Early Lias-Cenomanian). Annales Societatis Geologorum Poloniae, 63, 3–12.
Michelin, H. (1841). Iconographie zoophytologique. Description par localités et terrains des polypiers fossiles de France (1). Bertrand.
Milne Edwards, H., & Haime, J. (1848). Observations sur les polypiers de la famille des astréides. Comptes Rendus Hebdomadaires des Séances de l’académie des Sciences, 27(19), 465–469.
Milne Edwards, H., & Haime, J. (1849). Mémoire sur les polypiers appartenant à la famille des oculinides, au groupe intermédiaire des Pseudoastréides et à la famille des Fongides. Comptes Rendus Hebdomadaires des Séances de l’académie des Sciences, 29(4), 67–73.
Orbigny, A. (1849). Note sur les polypiers fossiles. Masson.
Orbigny, A. (1850). Prodrôme de Paléontologie stratigraphique universelle des animaux mollusques et rayonnés (1–2). Masson.
Orbigny, A. (1851). Cours élémentaire de Paléontologie (3:) Polypiers ou Zoophytes. Masson.
Pandey, D. K., McRoberts, C. A., & Pandit, M. K. (1999). Dimorpharaea de Fromentel, 1861 (Scleractinia, Anthozoa) from the Middle Jurassic of Kachchh, India. Journal of Paleontology, 73, 1015–1028.
Reuss, A. E. (1846). Die Versteinerungen der böhmischen Kreideformation (2). Schweitzerbart.
Rey, J. (2006). Stratigraphie séquentielle et séquences de dépôt dans le Crétacé Inférieur du Bassin Lusitanien. Ciências da Terra, Special, 6, 1–120.
Roniewicz, E. (2011). Early Norian (Triassic) corals from the Northern Calcareous Alps, Austria, and the intra-Norian faunal turnover. Acta Palaeontologica Polonica, 56(2), 401–428. https://doi.org/10.4202/app.2009.0092
Segura, M., Barroso-Barcenilla, F., Callapez, P. M., García-Hidalgo, J. F., & Gil, J. (2014). Depositional sequences and cephalopod assemblages in the upper Cenomanian-lower Santonian of the Iberian Peninsula (Spain and Portugal). Geologica Acta, 12, 19–27. https://doi.org/10.1344/105.000002056
Skelton, P. W. (2006). Introduction to the Cretaceous. In P. W. Skelton (Ed.), The Cretaceous world (pp. 9–41). Cambridge University Press.
Soares, A. F. (1980). A «Formação Carbonatada» na região do Baixo Mondego. Comunicações dos Serviços Geológicos de Portugal, 66, 99–109.
Soares, A. F., Kullberg, J. C., Marques, J. F., Rocha, R. B., & Callapez, P. M. (2012). Tectono-sedimentary model for the evolution of the Silves Group (Triassic, Lusitanian Basin, Portugal). Bulletin de la Société Géologique de France, 183(3), 203–216. https://doi.org/10.2113/gssgfbull.183.3.203
Stanley, S. M., & Hardie, L. A. (1998). Secular oscillations in the carbonate mineralogy of reef-building and sediment-producing organisms driven by tectonically forced shifts in seawater chemistry. Palaeogeography, Palaeoclimatology, Palaeoecology, 144, 3–19.
Stoliczka, F. (1873). The corals or Anthozoa from the Cretaceous rocks of South India. Memoirs of the Geological Survey of India, Palaeontologia Indica, (4), 8(4/5), 130–202.
Tomes, R. F. (1893). Observations on the affinities of the genus Astrocoenia. Quarterly. Journal of the Geological Society of London, 49, 569–573.
Tucker, M. E., & Wright, V. P. (1992). Carbonate depositional environments. Blackwell Scientific Publications.
Vaughan, T. W., & Wells, J. W. (1943). Revision of the suborders, families and genera of Scleractinia. Special Papers. Geological Society of America, 44, 1–363.
Wells, J. W. (1933). Corals of the Cretaceous of the Atlantic and Gulf Coastal plains and Western Interior of the United States. Bulletins of American Paleontology, 18(67), 83–292.
Wilson, R. C. L., Hiscott, R., Willis, M., & Gradstein, F. (1989). The Lusitanian Basin of west central Portugal, Mesozoic and Tertiary tectonic, stratigraphic and subsidence history. In A. Tankard, & H. Balkwill (Eds.), Extensional tectonics and stratigraphy of the North Atlantic margins. AAPG Memoir, 46, 341–361.
Acknowledgements
Fieldwork and material preparation was supported by the UNAM/DGAPA PAPIIT project IN101111 (Mexico). Authors are grateful to the Earth Sciences Department at the University of Coimbra for access to the laboratory facilities. Pedro Callapez would like to thank the institutional support of CITEUC – Centro de Investigação da Terra e do Espaço da Universidade de Coimbra (Earth and Space Research Centre from the University of Coimbra) and FCT – Fundação para a Ciência e Tecnologia. Preparation of thin sections in the ERNO laboratory by Aimée Orcí (Hermosillo, Sonora, Mexico) is gratefully acknowledged. The English text correction was realised by Matthew Copley (Barcelona). Two anonymous reviews and advice of the editors helped to improve the quality of the publication.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Rights and permissions
About this article

Cite this article
Löser, H., Callapez, P.M. Upper Cenomanian and lower Turonian (Cretaceous) corals from the Tethyan West Portuguese Carbonate Platform. J Iber Geol 48, 141–162 (2022). https://doi.org/10.1007/s41513-022-00186-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s41513-022-00186-3