
Abstract
Endophytic strains of Trichoderma species can be used as an alternative to chemicals to control vascular streak dieback (VSD) disease of cacao. Of 21 Trichoderma isolated from Theobroma cacao (cacao) in Indonesia, 19 were identified as Trichoderma asperellum. Four isolates of this species (ART-4, ART-5, ART-6 and ART-8) were reintroduced into young cocoa seedlings by root inoculation and after 4 weeks all were recovered from roots and stems, while ART-4 and ART-5 were recovered from leaves as well. Spraying seedlings pre-inoculated with T. asperellum ART-4, ART-5 and ART-6 with mycelium of the VSD pathogen Ceratobasidium theobromae resulted in no apparent VSD symptoms on the leaves. Those seedlings pre-inoculated with ART-8 showed 8.9 % incidence of VSD symptoms on the leaves when compared to a 20.4 % incidence of VSD on positive control leaves, 12 weeks after inoculation. The same isolates were also reintroduced into 3-month-old cocoa seedlings via the connecting site following shoot grafting, and after 12 weeks all isolates were recovered from stem and leaves. Seedlings grafted with buds infected by VSD and treated by ART-4 showed no VSD symptoms on their leaves 12 weeks after grafting and inoculation. However, those treated with ART-5, ART-6 and ART-8 showed 33.3 %, 50.0 % and 56.0 % incidence of VSD symptoms on their leaves, respectively, compared to an 88.9 % incidence of VSD on positive control leaves. Therefore, the study results demonstrates for the first time the potential of T. asperellum isolates to control VSD disease on cacao.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
Species of Trichoderma (teleomorph Hypocrea) are considered mostly to be common soil inhabitants, saprophytes and parasites of other fungi (Samuels et al. 2002). However, they are capable of more intimate associations with plant root systems in what has been characterized as an opportunistic avirulent symbiotic relationship (Harman et al. 2004). The critical phase of this association is the penetration of Trichoderma into the outer layers of cells of plant roots, where they persist. In the last decade, Trichoderma species have also shown to be among the most abundant avirulent endosymbionts in the living sapwood and leaves of tropical trees such as Cola spp., Herrania sp., Theobroma and Hevea spp. (Evans et al. 2003; Chaverri et al. 2011). On cacao (Theobroma cacao), diverse species of Trichoderma have been identified as endophytes, including T. stromaticum (Samuels et al. 2000), T. ovalisporum (Holmes et al. 2004), T. koningiopsis (Samuels et al. 2006a), T. theobromicola and T. paucisporum (Samuels et al. 2006b), T. martiale (Hanada et al. 2008), T. evansii (Samuels and Ismaiel 2009), and T. hamatum and T. harzianum (Bailey et al. 2009). Bailey et al. (2009) demonstrated that strains of T. hamatum and T. harzianum are capable of colonizing the above ground tissues of cacao and can be isolated from stem tissues, including the bark and xylem, glandular trichomes, the apical meristem, and leaves.
Strains of Trichoderma that were originally isolated as endophytes from sapwood of stems of Theobroma species have been shown to have a protective effect against fungal and oomycete pathogens. Some measure control of the cacao pathogen Moniliophthora roreri were achieved by topical application of an endophytic strain of T. ovalisporum (Krauss et al. 2010), and strains Trichoderma species that had been isolated as endophytes from cacao stems induced resistance to Phytophthora when they were introduced into roots of hot pepper (Capsicum annuum), although none of those strains were able to colonize the pepper stems (Bae et al. 2011). However, there is no information about the efficacy of endophytic Trichoderma strains against systemic diseases such as vascular streak dieback (VSD) disease of cacao. This disease is caused by the basidiomycete fungus Ceratobasidium theobromae (formerly Oncobasidium theobromae) (Guest and Keane 2007; Samuels et al. 2012), has devastated cocoa smallholdings in tropical Southeast Asian countries, including Indonesia. Indonesia is the world’s third largest producer of cacao (Shapiro et al. 2008; Rifin 2013), and most of that production is found in the Indonesian island of Sulawesi, the largest area of planted cacao in Indonesia. Basidiospores are dispersed from corticioid basidiomata by wind. They infect through leaves and grow through the xylem, killing branches or whole trees (Samuels et al. 2012). There is no effective control for this disease, which occurs only in Southeast Asia. In Sulawesi, the inability to control the disease has had disastrous consequences. A second species of Ceratobasidium, C. ramicola, is also associated with diseased cacao in Southeast Asia, but its role, if any, in VSD is unknown (Samuels et al. 2012).
In Sulawesi, cacao has been cultivated since 1560, although intensive cultivation only began in the late 1970s (Ruf 2007; Anshary 2002), thus this ecosystem can be considered to be stable, and under this condition we expect the populations of beneficial microorganisms, including Trichoderma in the soil and as endophytes in trees, to be stable. The objective of the present research was to isolate and identify Trichoderma species from above ground tissues of cacao including pod, beans, and sapwood, and to evaluate their potential for suppressing VSD and for stimulating growth of cacao.
2 Materials and Methods
2.1 Sources of Isolates
Living cultures of Trichoderma species were obtained from pod surface, pod placenta, beans, sapwood of cacao, and larvae of cocoapod borer (CPB). For isolation from pods, pods were surface sterilized by sequential immersion in 2 % sodium hypochlorite, 70 % ethanol and sterilized water (Arnold et al. 2003). The husk was removed from the pod using a surface sterilized knife and immediately each piece of husk was cut into five small pieces (± 1 cm2). Three placentas with beans, and three beans were removed separately with a flamed scalpel along with any larvae of CPB, and placed in Petri dishes containing 20 mL potato dextrose agar (PDA, Difco, Detroit, MI). To expose the sapwood, bark of branches was removed, and pieces of sapwood (2.5–5 × 1–2 mm) were placed on PDA in 9-cm-diameter Petri dishes. Samples for this study were obtained from Bantaeng, Bone, Soppeng, Luwu and Polewali Mandar regencies. We obtained five apparently healthy pods from these sites. Additionally, we sampled ten branches from invested VSD trees from North Luwu and Mamuju regencies.
2.2 Isolate Identification
To obtain fresh mycelium for DNA extraction, 21 isolates were grown in cornmeal broth (Difco) in a 5-cm-diameter Petri dish for 4–5 days at 25 °C. The mycelia (±0.5 g) were harvested and dried using clean, absorbent, paper towels. The entire dried mycelial mat was then placed in a 1.5 mL Eppendorf tube for immediate DNA extraction. Extraction of the genomic DNA was performed as reported previously (Dodd et al. 2002). Polymerase chain reaction for amplification of translation elongation factor (tef1) was performed as reported previously (Samuels et al. 2006b). The primers used for tef1 amplification were EF1-728 F (Carbone and Kohn 1999) and TEF1 rev (Samuels et al. 2002). An amplicon of about 700 kbp was obtained and sequenced. Sequences of both DNA strands were obtained using the BigDye terminator cycle sequencing kit and an Applied Biosystems 3100 DNA sequencer (Applied Biosystems). DNA sequences were assembled using Sequencher 3.1 (Gene Codes). The tef1 sequences obtained were subjected to Basic Local Alignment Search Tool (BLAST) available at http://blast.ncbi.nlm.nih.gov/Blast.cgi. The DNA sequence of this section of tef1 served to identify Trichoderma at the species level (Samuels et al. 2006a).
2.3 Assessment of Endophytic Ability
Four isolates of Trichoderma, that were presumed to be endophytic (ART-4, ART-5, ART-6 and ART-8) were selected and tested for their ability to colonize cocoa seedlings. Selection of the isolates was based on presence of Trichoderma of pods origin and a preliminary study showed that these four isolates were most effective in controlling of VSD on seedling. Seedlings were inoculated either through roots or through connections at the site of shoot grafts. For root inoculation, for each isolate three beans were surface sterilized and then placed in layers of sterile moist cloth for 3 days to stimulate germination, following which they were transferred to sterile filter paper in dishes measuring 7 cm diameter × 12 cm high. After incubation for 3 days, the roots were inoculated by adding a suspension of Trichoderma containing 1 × 106 mL−1 conidia in sterile distilled water (SDW) into the dishes using a pipette. Three additional seedlings were inoculated with SDW as control. One week after inoculation, seedlings were transferred to poly-bags containing about 1 kg sterile soil and then were placed in a greenhouse. For inoculating shoot grafts, 3 × 4-month-old seedlings for each tested isolate were cut about 25 cm from the top and a suspension of Trichoderma containing 1 × 106 mL−1 conidia was dropped onto this cut before grafting with a 3-cm length of bud wood. Root-inoculated seedlings were incubated for 1 month, and the shoot-graft plants were incubated for 2 months before being assayed for the presence of endophytic Trichoderma. To determine the presence of endophytic Trichoderma in each case stems were cut into 1 cm sections and the leaves into 1 cm2 pieces. These were sterilized in 2 % sodium hypochlorite, 70 % ethanol and vigorously washed several times in SDW before being placed onto PDA in Petri dishes. These Petri dishes were incubated at room temperature and examined every day for the presence of Trichoderma.
2.4 Screening for VSD: Inoculation Through Roots
Cultures of C. theobromae were obtained from naturally infected cacao branches in the field. To isolate this fungus, fragments of branches were surface sterilized as was described above for the isolation of Trichoderma. First, the bark was removed from the surface sterilized segments, which were then cut into 0.5 cm sections and placed onto water agar (WA) in Petri dishes. C. theobromae was recognized by its cottony, non-sporulating white mycelium that grows slowly from the ends of the segments within a few days at 25 °C, as was described by Samuels et al. (2012). For inoculum, ±0.5 g fresh mycelium was crushed with a sterile spatula and shaken in 10 mL of sterile distilled water to break up the mycelium. This suspension was sprayed onto one seedling that had previously inoculated with one of four isolates of Trichoderma. Each treatment consisted of nine seedlings for each of the four Trichoderma isolates plus nine each for the positive and negative controls, for a total of 54 seedlings. These seedlings were placed in greenhouse, where they were observed weekly for the appearance of symptoms. Incidence of disease was calculated according to the formula: I = A/B, where A is the number of leaves showing VSD symptoms and B is the number of all leaves observed.
2.5 Screening for VSD: Inoculation Through the Connecting Site of Shoot Grafts
Bud wood uniformly infected with VSD was selected from the field and was grafted to 3 month-old seedling stocks and inoculated with Trichoderma at the time of grafting. The viability of the infected grafts was low; only ±20 % of the grafted plants survived to the end of the experiment. Because of the low viability of the grafted plants, each treatment consisted of only five shoot grafts. All shoot grafts were placed in the nursery at field temperature (27–31 °C) and the incidence of disease was observed for 12 weeks after grafting and inoculation with Trichoderma. The incidence of disease was measured using the formula as above.
2.6 Analysis
The data of VSD incidence were analyzed after transformation to Log x + 1. Duncan’s multiple range test (DMRT) was then used for evaluating significant differences between the treatment means.
3 Results
3.1 Identification of Trichoderma from Cacao Tissue and Cocoa Pod Borer Larvae
A total of 21 isolates from cacao in Sulawesi were obtained from pod surface, placenta, bean, sapwood of branches, and larvae of cocoa pod borer (CPB). These isolates were identified at the species level by analysis of their translation elongation factor (tef1) (Table 1). Of the 21 isolates, 19 were T. asperellum and two were T. virens. The only isolation from CPB larvae was one culture of T. asperellum.
3.2 Endophytic Inoculation
No growth was observed within 4 days of incubation on PDA in Petri dishes from material taken from control roots, stems or leaves, 1 month after Trichoderma strains were inoculated into test plants. On this basis, the Trichoderma that was reisolated from inoculated plants was assumed to be the strain that was used in the inoculation. Trichoderma asperellum strains ART-4, ART-5, ART-6 and ART-8 were reisolated from root and stem sections 1 month after inoculation through roots. Only isolates ART-4 and ART-5 were reisolated from leaf pieces. Strains ART-4, ART-5, ART-6 and ART-8 were reisolated from the connection point of the grafts at the end of the assay or after 3 months on stem and leaf pieces, but no Trichoderma were reisolated from the roots.
3.3 Screening for VSD: Inoculation Through Root
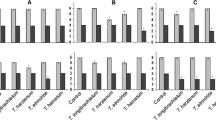
Screening experiments were conducted in the laboratory for 3 months to estimate the potential for controlling VSD with T. asperellum strains ART-4, ART-5, ART-6 and ART-8 when inoculated through roots. In plants inoculated with strains ART-4, ART-5 and ART-6 1 month before being inoculated with mycelium of C. theobromae no symptoms of VSD developed, and there was a lower level of symptom development in plants inoculated with ART-8 (Fig. 1). No symptoms appeared on negative controls or on seedlings treated with ART-4, ART-5 and ART-6 after the 3 months period. Symptoms of VSD appeared within 2 months on plants infected with C. theobromae, and the disease incidence in this positive control was 20.4 % (Fig. 1)

Vascular streak dieback (VSD) incidence on seedlings after treatment by four Trichoderma asperellum strains (ART-4, ART-5, ART-6 and ART-8) through roots. The first appearance of symptoms was 8 weeks after inoculation. Control (−): negative control; Control (+): positive control. Means of incidence in the same time followed by same letter are not significantly different according to DMRT (P ≤ 0.05)
3.4 Screening for VSD: Inoculation Through Shoot Grafts
Cacao bud wood lightly infected by VSD could be grafted and persisted as growing seedlings for the 12 weeks of the study. On seedlings not treated with T. asperellum (positive control), the VSD symptoms were conspicuous within 10 weeks of grafting, and a high disease incidence with 88.9 % was observed. On seedlings treated with strains ART-5, ART-6 and ART-8, a reduced disease incidence 33.0 %, 50.0 % and 56.0 %, respectively, was observed, while on seedlings treated with ART-4, there was no indication of disease, similar to the negative control (Fig. 2).

Vascular streak dieback (VSD) incidence on seedling grafted after treatment by four Trichoderma asperellum strains (ART-4, ART-5, ART-6 and ART-8) through connecting site of shoot grafting. The first appearance of symptoms was 10 weeks after grafting of bud wood. Control (−): negative control; Control (+): positive control. Means of incidence in the same time followed by same letter are not significantly different according to DMRT (P ≤ 0.05)
The treatment with T. asperellum through the grafting site affected seedling growth. Treatment with ART-4 coincided with a higher number of shoots (average of 4.3) than other treatments. With ART-5, ART-6, ART-8, positive control and negative control, the average of shoots number was 1.5, 1.6, 2.0, 2.0 and 2.0, respectively
4 Discussion
The results reported here is the first assessment of the occurrence of endophytic Trichoderma in cacao plantations in Sulawesi, Indonesia. Two species were identified: 90.4 % of the 21 isolations were T. asperellum, and the rest were T. virens. In a study that included soil isolates from the Indonesian islands of Java and Bali, Kubicek et al. (2003) recorded T. harzianum, T. virens and T. spirale. Sriwati et al. (2011) isolated T. virens and T. longibrachiatum as endophytes from cacao in Aceh Province of Indonesia, but they did not encounter T. asperellum.
The four isolates of T. asperellum used in the present work were isolated from cacao pods. The reintroduction of these four isolates into cacao seedlings indicates that strains of Trichoderma can invade and endophytically colonize stems and leaves. The same endophytic capability has been also observed with other isolates from pod (Table 1) (data not shown here). Our results suggest a role for T. asperellum in combating VSD. When inoculated through roots, plants resisted artificial inoculation with C. theobromae. When previously infected plants were inoculated with the Trichoderma at the grafting site, symptoms of VSD were significantly reduced. In case of root inoculation we were not able to recover T. asperellum from leaves although the roots were colonized. This suggests that the Trichoderma stimulated resistance to infection of above-ground plant parts as it has been described for this and other Trichoderma spp. (Shoresh et al. 2010; Hermosa et al. 2012). When Bae et al. (2011) applied endophytic Trichoderma isolates to roots of hot pepper they obtained resistance to attack by Phytophthora capsici, but just as we have observed, these cultures were not efficient stem colonizers. On the other hand, when Evans et al. (2003) applied conidial suspensions of endophytic Trichoderma to the meristems of cacao seedlings there was colonization of above ground plant parts. The apparent ability of T. asperellum to induce resistance to C. theobromae is consistent with the current view that at least some strains of Trichoderma species are asymptomatic plant endosymbionts, colonizing the intercellular spaces at the surface of plant roots epidermis and cortex and interacting with the plant to balance the costs of growth and defense (Hermosa et al. 2013). Interestingly, when T. asperellum ART-4 was inoculated through the grafting site, symptom development in infected bud wood did not proceed when compared to grafts done in the absence of the Trichoderma. We are not aware of Trichoderma having been used in this way to actually cure infected bud wood. The mode of action here suggests a direct action against the pathogen through mycoparasitism or antibiosis. Mycoparasitism and antibiosis are well known modes of action for Trichoderma (Bailey et al. 2008; Bae et al. 2011; Atanasova et al. 2013). This result suggests that the routine application of T. asperellum at the time of grafting can be effective with new planting, especially when disease pressure is high.
There was variation among the four strains of T. asperellum that we studied. While three of them, ART-4, ART-5 and ART-6 protected young seedlings, ART-4 was more effective in ameliorating the effects of VSD in grafted budwood. This strain had the added ability of enhancing growth of cacao shoot graft seedlings. We do not know the mechanism that accounts for growth stimulation in cacao seedlings, but individual strains of some Trichoderma species are known to induce increases in plant biomass, leaf area and number of roots, and photosynthetic ability in addition to disease suppression (Chang et al. 1986; Shoresh et al. 2010). Because of its ability to suppress VSD while enhancing growth of cacao seedlings in experimental trials, strain T. asperellum ART-4 is now being applied under field conditions in Sulawesi.
The incidence of VSD on seedlings was lower than on shoot grafts. Artificial inoculation of leaves is a slow and somewhat ineffective process. Hyphae of the pathogen need to sporulate and the spores then must germinate on young leaves and penetrate this surface of leaf. Therefore, in this study, the concentration of inoculums developed in cacao seedling tissue was possibly lower than that developed in cacao shoot grafts tissue because of direct inoculation through bud wood infected.
References
Anshary A (2002) Karakteristik tanaman kakao (Theobroma cacao L) yang resisten terhadap penggerek buah kakao Conopomorpha cramerella (Snellen) (Lepidoptera: Gracillariidae). Desertasi Program Pascasarjana. Universitas Hasanuddin, Makassar
Arnold AE, Mejia LC, Kyllo D, Rojas EI, Maynard Z, Robbins N, Herre EA (2003) Fungal endophytes limit pathogen damage in a tropical tree. Proc Natl Acad Sci U S A 100:15649–15654
Atanasova L, Le Crom S, Gruber S, Coulpier F, Seidl-Seiboth V, Kubicek CP, Druzhinina IS (2013) Comparative transcriptomics reveals different strategies of Trichoderma mycoparasitism. BMC Genomics 14:121
Bae H, Roberts DP, Lim H-S, Strem MD, Park S-C, Ryu C-M, Melnick RL, Bailey BA (2011) Endophytic Trichoderma isolates from tropical environments delay disease onset and induce resistance against Phytophthora capsici in hot pepper using multiple mechanisms. Mol Plant-Microbe Interact 24:336–351
Bailey BA, Bae H, Strem MD, Crozier J, Thomas SE, Samuels GJ, Vinyard BT, Holmes KA (2008) Antibiosis, mycoparasitism, and colonization success for endophytic Trichoderma isolates with biological control potential in Theobroma cacao. Biol Control 46:24–35
Bailey BA, Strem MD, Wood D (2009) Trichoderma species form endophytic associations within Theobroma cacao trichomes. Mycol Res 113:1365–1376
Carbone I, Kohn LM (1999) A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 91:553–556
Chang YC, Chang YC, Baker R, Kleifeld O, Chet I (1986) Increased growth of plants in the presence of the biological control agent Trichoderma harzianum. Plant Dis 70:145–148
Chaverri P, Gazis RO, Samuels GJ (2011) Trichoderma amazonicum, a new endophytic species on Hevea brasiliensis and H. guianensis from the Amazon basin. Mycologia 103:139–151
Dodd SL, Lieckfeldt E, Chaverri P, Overtom BE, Samuels GJ (2002) Taxonomy and phylogenetic relationship of two species of Hypocrea with Trichoderma anamorphs. Mycol Prog 1:409–428
Evans HC, Holmes KA, Thomas SE (2003) Endophytes and mycoparasites associated with an indigenous forest tree, Theobroma gileri, in Ecuador and a preliminary assessment of their potential as biocontrol agents of cocoa diseases. Mycol Prog 2:149–160
Guest D, Keane PJ (2007) Vascular streak dieback: a new encounter disease of cacao in Papua New Guinea and Southeast Asia caused by the obligate basidiomycete Oncobasidium theobromae. Phytopathology 97:1654–1657
Hanada RE, Jorge Souza TD, Pomella AW, Hebbar KP, Pereira JO, Ismaiel A, Samuels GJ (2008) Trichoderma martiale sp. nov., a new endophyte from sapwood of Theobroma cacao with a potential for biological control. Mycological Research 112:1335–1343
Harman GE, Howell CR, Viterbo A, Chet I, Lorito M (2004) Trichoderma species -opportunistic, avirulent plant symbionts. Nat Rev Microbiol 2:43–56
Hermosa R, Viterbo A, Chet I, Monte E (2012) Plant-beneficial effects of Trichoderma and of its genes. Microbiology 158:17–25
Hermosa R, Rubio MB, Cardoza RE, Nicolas C, Monte E, Gutierrez S (2013) The contribution of Trichoderma to balancing the costs of plant growth and defense. Int Microbiol 16:69–80
Holmes KA, Schroers HJ, Thomas SE, Evans HC, Samuels GJ (2004) Taxonomy and biocontrol potential of a new species of Trichoderma from the Amazon basin in South America. Mycol Prog 3:199–210
Krauss U, Hidalgo E, Bateman R, Adonijah V, Arroyo C, García J, Crozier J, Brown NA, Hoopen GM, Holmes KA (2010) Improving the formulation and timing of application of endophytic biocontrol and chemical agents against frosty pod rot (Moniliophthora roreri) in cocoa (Theobroma cacao). Biol Control 54:230–240
Kubicek CP, Bisset J, Druzhinina I, Kullnig-Gradinger C, Szakacs G (2003) Genetic and metabolic diversity of Trichoderma: a case study on South-East Asian isolates. Fungal Genet Biol 38:310–319
Rifin A (2013) Competitiveness of Indonesia’s cocoa beans export in the world market. International Journal of Trade, Economics and Finance 4:279–281
Ruf F (2007) The cocoa sector: Expansion, or green and double green revolutions. Background note presented on ODI/International Food Policy Research Institute Workshop. Accra, Ghana. IFPRI
Samuels GJ, Ismaiel A (2009) Trichoderma evansii and T. lieckfeldtiae two new T. hamatum-like species. Mycologia 101:142–156
Samuels GJ, Pardo-Schultheiss RA, Hebbar KP, Lumsden RD, Bastos CN, Costa JC, Bezerra JL (2000) Trichoderma stromaticum sp. nov., a parasite of the cacao witches broom pathogen. Mycol Res 104:760–764
Samuels GJ, Dodd SL, Gams W, Castlebury LA, Petrini O (2002) Trichoderma species associated with the green mold epidemic of commercially grown Agaricus bisporus. Mycologia 94:146–170
Samuels GJ, Dodd SL, Lu B-S, Petrini O, Schroers H-J, Druzhinina IS (2006a) The Trichoderma koningii aggregate species. Stud Mycol 56:67–133
Samuels GJ, Suarez C, Solis K, Holmes KA, Thomas SE, Ismaiel A, Evans HC (2006b) Trichoderma theobromicola and T. paucisporum: two new species from South America. Mycol Res 110:381–392
Samuels GJ, Ismaiel A, Rosmana A, Junaid M, Guest D, McMahon P, Keane P, Purwantara A, Lambert S, Rodriguez-Carres M, Cubeta MA (2012) Vascular Streak Dieback of cacao in Southeast Asia and Melanesia: in planta detection of the pathogen and a new taxonomy. Fungal Biol 116:11–23
Shapiro LH, Scheffer SJ, Maisin N, Lambert S, Purung HB,Sulistyowati E, Vega FE, Gende P, Laup S, Rosmana A, Djam S, Hebbar PK (2008) Conopomorpha cramerella (Lepidoptera: Gracillariidae) in the Malay Archipelago: genetic signature of a bottlenecked population? Annals of the Entomological Society of America 10: 930–938
Shoresh M, Harman GE, Mastouri F (2010) Induced systemic resistance and plant responses to fungal biocontrol agents. Ann Rev Phytopathol 48:21–43
Sriwati R, Chamzurni T, Sukarman (2011) Deteksi dan identifikasi cendawan endofit Trichoderma yang berasosiasi pada tanaman kakao. Jurnal Agrista 15:15–20
Acknowledgments
The authors are grateful to the Indonesian Agency for Agricultural Research and Development, Ministry of Agriculture and Directorate General of Higher Education, Ministry of Education and Culture who have supported this research through KKP3T and PAR-C research grants, respectively. Also, we would like to thank the University of Maryland and the United States Department of Agriculture, Beltsville, MD, USA, for giving permission to complete the research in their laboratories.
Author information
Authors and Affiliations
Corresponding author
Additional information
Section Editor: Wagner Bettiol
Rights and permissions
About this article

Cite this article
Rosmana, A., Samuels, G.J., Ismaiel, A. et al. Trichoderma asperellum: A Dominant Endophyte Species in Cacao Grown in Sulawesi with Potential for Controlling Vascular Streak Dieback Disease. Trop. plant pathol. 40, 19–25 (2015). https://doi.org/10.1007/s40858-015-0004-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40858-015-0004-1