
Abstract
While siblings can be close allies, they can also be significant competitors. They are also family members that are typically with us for most of our lives. Research has raised questions about the factors shaping sibling relationships, and an adaptationist perspective would predict a role for a number of factors including sex, genetic relatedness, and childhood co-residence. Recent work has highlighted sex differences with regard to conflict and emotional closeness, greater conflict among full-siblings than half-siblings, and a role for co-residence in increasing sibling altruism. This study examines levels of both sibling conflict and sibling cooperation as a function of respondent sex, sex of sibling, birth interval (or absolute age difference), co-residence, and relatedness. Results indicate that sibling conflict and cooperation may not be shaped by the same set of factors. Sibling conflict was predicted by own sex, sex of sibling, birth interval, duration of co-residence, and the degree of relatedness. Greater levels of conflict were reported by sisters, those closer in age, those who have co-resided longer, and full-siblings compared to half-siblings. However, sibling prosocialness was only predicted by sex and relatedness with females and full siblings reporting greater levels of sibling prosocialness. More research investigating patterns of conflict and cooperation within families using more ecologically valid cues are necessary to determine whether the two are operating under the same mechanism, sensitive to the same cues, or are, indeed, operating under different mechanisms.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
There is no friend like a brother. There is also no enemy like a brother.
Bengali proverb
Introduction
Much has been written about the joy and the anguish of sibship. Siblings can be with us from our youngest days until our last days and can be incredible sources of support and cooperation as well as sources of conflict and aggravation. Many historical and fictional accounts focus on the most extreme sibling conflict, whether we are considering the siblicidal Biblical story of Cain and Abel or tales of ancient rulers who killed their siblings to ensure their own children would inherit resources and power. However, the majority of sibling conflict is not that extreme, though it may involve physical aggression and has been considered by some researchers a normal part of human development and something that declines over the lifespan (Campione-Barr & Smetana, 2010; Cicirelli, 1995; Jensen et al., 2018; McHale et al., 2012). When young, siblings spend a majority of time in close proximity which creates opportunities for cooperation and conflict. Along with conflict, cooperation and substantial affection between siblings is also highly common, perhaps especially among sisters (Buist et al., 2002; Cicirelli, 1995; Tibbetts & Scharfe, 2015). Siblings can be playmates, surrogate parents, and sources of substantial social support. From an evolutionary perspective, what factors should influence sibling conflict and cooperation? Do the same factors that influence conflict also influence cooperation, or are there different mechanisms shaping conflict versus cooperation for siblings? That is, are conflict and cooperation opposite ends of the same continuum or are they independent of each other?
Sibling Conflict and Cooperation Theory
Sibling relationships can be the most enduring relationships of our lives (Cicirelli, 1995). Inherent in such long-term ties is the potential for conflict as well as cooperation. Explaining cooperation and altruism from an evolutionary perspective, especially among kin, has been greatly influenced by Hamilton's (1964) proposal of kin selection theory. Hamilton introduced the concept of inclusive fitness (fitness not only due to only one’s own survival and reproduction, but also that of genetically related individuals) as an extension of classic Darwinian fitness. This new perspective modeled the likelihood that an individual will behave altruistically toward another depending on whether the degree of relatedness and the potential fitness benefit to the other person outweigh the potential fitness cost to oneself. Thus, altruistic behavior toward kin serves to indirectly increase one’s own inclusive fitness. This would suggest that genetic relatedness will influence the likelihood of conflict and cooperation between individuals with greater cooperation (and less conflict) occurring between genetically related individuals all else being equal. Studies investigating feelings of subjective closeness and received support across adult relationships as a function of degree of relatedness have produced results consistent with this theory. The highest levels of closeness and support were directed toward kin versus non-kin and, in addition, greater levels were directed to more closely related kin, like siblings, versus more distantly related kin such as cousins (Neyer & Lang, 2003). Twins have often been highlighted specifically in terms of developing intense intimate bonds (Segal, 1999). Since siblings are highly related, we would expect significant levels of solidarity and willingness to provide support.
However, shared genetic interests do not mean identical interests. One example of this is seen in parent-offspring conflict theory (Trivers, 1974). While offspring are the primary vehicles of parental fitness, their genetic interests are not identical. As a result, there is room for conflict, and one might expect offspring to have been selected to manipulate parents in the pursuit of greater amounts of parental investment while parents have been selected to manipulate offspring in the pursuit of their own best fitness interests. From any focal offspring’s point of view, the more parental investment, the better. From the parental point of view, the more invested in any one offspring, the less available to invest in other current or future offspring (Trivers, 1972). Parental investment theory (Trivers, 1972) suggests that the allocation of parental investment is a function of the amount of parental resources, offspring quality, and potential for future offspring. All else being equal, one would expect parents to value offspring equally and to encourage them to value their siblings perhaps more than they are naturally inclined to do. However, each offspring is more closely genetically related to themselves than to their siblings and, as such, generally values themself over their siblings. This raises the possibility of sibling conflict over parental resources with each sibling trying to maximize their “fair share” in comparison to their siblings.
So, from an evolutionary perspective, sibling conflict is rooted in conflict over limited resources. For human children, as well as other animal offspring, this often means competition over limited parental resources which historically, for humans, would have included food and basic resources such as shelter and time spent teaching useful skills (hunting and weaving). As a result, we would expect greater levels of sibling conflict in families with more limited parental resources and/or greater numbers of offspring. Some research does indicate that larger sibships are associated with more sibling violence (Straus et al., 2006). However, for many families in the modern western world (where our study was conducted), access to resources is adequate if not abundant. Studies focusing on proximate explanations of sibling conflict have suggested that the most common sources of conflict include issues of relative power, self-interest (sharing personal items), violation of rules, and activities outside the family (McGuire et al., 2000). Both younger and older siblings’ most common complaint is about sharing personal possessions with the least common complaint competition over parental attention from toddlers to adolescents (Campione-Barr & Smetana, 2010; Dunn & Munn, 1985; McGuire et al., 2000).
Possible Factors Shaping Levels of Cooperation and Conflict
Sex of Sibling
A number of studies have indicated that sex plays a role in sibling behavior. Research suggests that boys experience more conflict with their siblings than girls do (Brody et al., 1985) and that boys engage in more violent sibling conflict with the greatest levels between brothers (Salmon & Hehman, 2015; Straus et al., 2006). While older sisters often engage in caretaking of younger siblings, several studies indicate that sister-sister relationships are more emotionally close than those between brothers or between brothers and sisters (Pollet, 2007; Stocker et al., 2020; Tanskanen & Danielsbacka, 2019; White & Reidman, 1992a). However, from a theoretical standpoint, same-sex individuals may be in more direct conflict over the same resources than opposite-sex siblings. While all siblings may be equal competitors for food or attention, opposite-sex ones are less likely to be competing for the same mates, for example. A recent study of co-playing video games and sibling conflict and affection indicated that playing together increased sibling affection among boys and girls but also was associated with greater conflict in boys, although playing violent games together was associated with decreased relationship conflict in boys, perhaps the result of cooperating in their video game scenarios (Coyne et al., 2016).
Relatedness
Until relatively recently, the impact of relatedness on sibling conflict has been largely understudied in humans, despite the volume of work on this topic in non-human animals (Hodge et al., 2008; Holmes & Sherman, 1982; Wahaj et al., 2004). Emlen’s framework of family relations (1997) predicted that brothers and sisters will invest more in full siblings than in half-siblings or step-siblings as sibling ties likely weaken with decreasing degrees of relatedness. He also suggested that conflict would increase as relatedness decreases. A number of studies examining sibling solidarity have provided some support for Emlen's (1997) predictions. For example, Jankowiak and Diderich's (2000) study of polygynous Mormons demonstrated a preference for full siblings over half-siblings despite cultural pressure to treat them the same. Pollet (2007) found, in a sample of Dutch adults, that relatedness predicted sibling interaction patterns, and White and Reidmann (1992b) reported more contact between full siblings than between step-/half-siblings. Twin studies have also reported more cooperation and emotionally closer relationships between monozygotic twins than between dizygotic ones (Segal, 2005) as well as more intense grief following the death of a monozygotic twin than other more distant relatives (Reed et al., 2021). However, recent studies have also reported some results that do not match Emlen’s or Hamilton’s predictions. Salmon and Hehman's (2015) study of college students in the USA indicated that genetic relatedness influenced the frequency and intensity of conflict but that the most intense conflict was between non-biological siblings, followed by conflict between full siblings with the least conflict between half-siblings. Similar results were also reported in a sample from the British Millennium Cohort study in which conflict was assayed via children’s reports of how much siblings picked on or hurt each other. Children living with only full siblings experienced more sibling conflict than those living with only maternal half-siblings, perhaps the result of differential access to parental resources (Tanskanen et al., 2017). In addition, Steinbach and Hank's (2018) study of full versus half versus step-sibling contact and conflict based on German Family Panel data also found that full siblings had more contact and felt closer than half-siblings but that full siblings also had more conflicts. Similarly, a UK study also reported higher levels of conflict with full than with half-siblings, including physical aggression (Khan et al., 2020).
Birth Spacing and Length of Co-residence
Two additional environmental factors that are likely to influence levels of sibling conflict and cooperation are birth spacing (or birth intervals) and length of co-residence. A number of studies have reported greater levels of conflict between siblings with short birth intervals with the highest levels of conflict occurring between siblings born within 2 years of each other and who would presumably be competing for similar parental resources (Cicirelli, 1995). Larger birth intervals have been linked to less conflict between siblings during childhood but also less close relationships as adults (Pollet, 2007). Studies that have investigated the role of length of co-residence on levels of sibling conflict have reported that longer co-residence was associated with more intense conflict while greater birth spacing was associated with less intensity of conflict (Salmon & Hehman, 2015), which is not surprising considering sibling conflict theory and parental investment (Sulloway, 1996). An online study of sibling relationship quality (Gyuris et al., 2020) that also examined co-residence and relatedness indicated that a greater degree of relatedness was associated with better sibling relations but only when siblings were not co-resident during childhood. However, when conflict was the variable of interest, full siblings reported more conflict during childhood than half-siblings, similar to Salmon and Hehman (2015). Similar results looking at relationship quality have also been reported in a German sample in which those who co-resided with their half-siblings reported better relationship quality (measured as contact frequency and emotional closeness) as adults than those that did not reside with their half-siblings (Tanskanen & Danielsbacka, 2019). Studies focusing on sibling cooperation have also examined the role of co-residence, usually as part of assessing cues of relatedness, reporting that maternal perinatal association (a cue of relatedness) as well as co-residence duration predict sibling-directed altruism (Sznycer et al., 2016) across several cultures and that younger full- and half-siblings elicited more altruism than step-siblings.
Current Study
This study examined levels of conflict (indexed by overall frequency as well as intensity) and cooperation (indexed by sibling prosocialness) between siblings with the following predictions:
Prediction 1a
Sex of sibling influences the level of conflict with greater conflict expected between same-sex siblings relative to opposite sex siblings.
Prediction 1b
Sex of sibling influences the level of cooperation with greater cooperation expected between same-sex siblings relative to opposite sex siblings.
Prediction 2a
Duration of co-residence influences the level of conflict with increased levels of conflict expected between siblings who have co-resided longer.
Prediction 2b
Duration of co-residence influences the level of cooperation with increased levels of cooperation expected between siblings who have co-resided longer.
Prediction 3a
Degree of relatedness influences the degree of conflict with greater levels of conflict expected between full siblings than half-siblings (replicating Salmon & Hehman, 2015 findings).
Prediction 3b
Degree of relatedness influences the degree of cooperation with greater levels of cooperation expected between full siblings than half-siblings.
Method
Participants
Participants included 364 young adults (262 females and 102 males) between the ages of 17 and 30 (M = 18.79, SD = 1.52) who reported on 573 siblings. Out of the 573 siblings, 290 were same-sex sibling pairs, and 283 were opposite-sex sibling pairs. There were 481 full siblings and 92 half-siblings. Approximately 45% of participants self-reported their ethnicity as Caucasian, 33% Hispanic/Latino, 6% Asian, 5% African American, 3% Middle Eastern, 2% South Asian, 1% Pacific Islander, and 5% “Other.” Participants were recruited from introductory psychology courses at a southwestern US private university and received course credit for their participation in the study.
Measures
Demographics
Participants were asked to self-report their sex, age, ethnicity, number of siblings, and the birth order of themselves and their siblings.
Sibling Conflict
Sibling conflict was operationalized using the Sibling Issues Checklist, which is composed of a list of 20 potential sibling conflict issues (Campione-Barr & Smetana, 2010). For each potential sibling conflict issue, participants are asked to rate (i) how frequently the issue arose between themselves and a particular sibling on a 5-point Likert scale where 1 indicated never and 5 indicated very often, and (ii) how intense those issues were on a 5-point Likert scale where 1 indicated calm and 5 indicated very angry. The scale yields two overall scores—one for frequency of conflict and one for intensity of conflict—as well as two factor scores (i.e., conflict over equality and fairness and conflict over invasion of personal and psychological space). Conflict over equality and fairness includes items such as conflict over “parents’ attention” and “whose turn to do chores.” Conflict over invasion of personal and psychological space includes items such as “telling other what to do” and “borrowing things without permission.” The overall conflict scores range from 20 to 100, with higher scores indicating more frequent and more intense conflict. The factor scores range from 10 to 50, with higher scores indicating more frequent and more intense conflict of that particular type. Participants were asked to complete the Sibling Issues Checklist for each of their siblings, indicating that sibling’s sex, age, and degree of relatedness (full sibling, half-sibling, step-sibling, or adopted sibling). Participants were also asked to report how long they co-resided with that sibling (“I lived with this sibling from the time I was _____ (age) until I was _____ (age)”). Cronbach’s alphas indicated there was a high level of internal consistency for frequency (α = .87) and intensity (α = .88) of conflict subscales, as well as the overall conflict scale (α = .93) with respect to this specific sample.
Sibling Cooperation
Sibling cooperation was operationalized using the Prosocialness Scale for Adults (Caprara et al., 2005). The scale consists of 16 statements describing common situations involving sharing, helping, taking care of, and feeling empathy for others and their needs/requests. We modified the scale to measure behaviors and feelings specifically directed to siblings by changing the “friends/colleagues” and “others” in the original items to reflect “this sibling.” For example, “I do what I can to help this sibling.” Participants rated each statement on a 5-point Likert scale where 1 indicated never/almost never true and 5 indicated almost always/always true. Possible scores on this scale range from 16 to 80, with higher scores indicating more prosocial behavior/feelings directed toward that sibling. Participants were asked to complete the prosocialness scale for each of their siblings. Cronbach’s alpha indicated there was a high level of internal consistency for this scale with respect to this specific sample (α = .97).
Procedure
Participants were provided a link to complete the survey online via SurveyMonkey. After giving informed consent, participants were directed to the survey where they first responded to the demographic questions, followed by the sibling conflict questions and the prosocial questions for each sibling. After completion of the survey, participants were given course credit for their time. All measures and procedures were approved by the authors’ university’s Institutional Review Board (IRB).
Results
The means (and standard deviations) of duration of co-residence and the sibling conflict and prosocial scores as a function of degree of relatedness (half vs. full) and sex of sibling (same sex vs. opposite sex) appear in Table 1. Preliminary analyses conducted on the two-factor conflict scores indicated the pattern of the results was not significantly different than the analyses with the overall scores. Therefore, only the analyses with the overall conflict frequency and intensity scores are reported here. Inspection of Table 1 indicates that (i) respondents reported co-residing for longer duration with full siblings than with half siblings; (ii) respondents reported more frequent and more intense sibling conflict as well as more prosocialness with full siblings compared to half siblings; and (iii) there were not any significant differences in self-reported co-residence, frequency and intensity of conflict, or prosocialness between same-sex and opposite-sex sibling pairs.
Effect of Sex of Sibling, Duration of Co-Residence, and Genetic Relatedness on Sibling Conflict
Overall Frequency of Sibling Conflict
A hierarchical linear regression analysis was conducted to test whether (i) there would be more frequent conflict between same-sex siblings relative to opposite-sex siblings; (ii) there would be more frequent conflict as duration of co-residence increases; and (iii) there would be more frequent conflict between full siblings relative to half-siblings. In order to control for participant and other sibling variables before accounting for the effect of genetic relatedness on frequency of sibling conflict, respondents’ sex (coded as 0 = female, 1 = male) was added in step 1; sex of sibling (coded as 0 = same-sex, 1 = opposite-sex), absolute difference between respondent’s age and sibling’s age, and duration of co-residence were entered in step 2; degree of relatedness (coded as 0 = half-siblings, 1 = full siblings) was entered in step 3; and the two-way interactions between degree of relatedness with co-residence, respondents’ sex, sex of sibling, and age difference were added in step 4. Results from this analysis are summarized in Table 2.
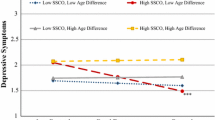
In step 1, the respondents’ sex explained approximately 1% of the variance in frequency of sibling conflict, F(1, 571) = 4.63, p = .03. Inspection of the standardized regression coefficient (β) indicates that females reported significantly more frequent sibling conflict than males. In step 2, sibling sex, age difference between siblings, and length of co-residence explained an additional 11% of the variance in frequency of sibling conflict, F(3, 568) = 23.54, p < .001. Inspection of the standardized regression coefficients (βs) indicates that whereas length of co-residence was not a significant predictor, sibling sex and the age difference between siblings were significant unique predictors of frequency of sibling conflict. The main effect of sibling sex indicates that there was significantly more frequent conflict between same sex siblings relative to opposite sex siblings. The main effect of age difference between siblings indicates that the frequency of conflict increases as the age difference decreases. In step 3, degree of relatedness explained an additional 1% of the variance in frequency of sibling conflict, F(1, 567) = 4.48, p = .04. Inspection of the standardized regression coefficient (β) indicates that there was significantly more frequent sibling conflict between full siblings than half-siblings. In step 4, the interactions between genetic relatedness and the other four predictor variables (respondent sex, sibling sex, age difference, and co-residence) explained an additional 2% of the variance in frequency of sibling conflict, F(4, 563) = 3.20, p = .01. Inspection of the standardized regression coefficients (βs) indicates that whereas the interactions between relatedness and respondent sex, sibling sex, and age difference were not significant predictors, the interaction between relatedness and co-residence was a significant unique predictor of frequency of sibling conflict. As shown in Fig. 1, frequency of sibling conflict significantly increased for half siblings the longer the duration of co-residence (p = .001) whereas frequency of conflict between full siblings was not greatly influenced by duration of co-residence (p = .53). Overall, approximately 14% of the variance in frequency of sibling conflict was explained by the final regression model, F(9, 563) = 10.53, p < .001, R2 = .144.

Interaction between genetic relatedness and duration of co-residence on frequency of sibling conflict
Overall Intensity of Sibling Conflict
A separate hierarchical linear regression analysis was conducted to test whether (i) there would be more intense conflict between same-sex siblings relative to opposite-sex siblings; (ii) there would be more intense conflict as duration of co-residence increases; and (iii) there would be more intense conflict between full siblings relative to half-siblings. The variables were entered into the model following the same procedure described above (i.e., respondents’ sex was entered in step 1; sex of sibling, absolute age difference, and duration of co-residence were entered in step 2; degree of relatedness was entered in step 3; and the two-way interactions between degree of relatedness with co-residence, respondents’ sex, sex of sibling, and age difference were added in step 4). Results from this analysis are summarized in Tables 3 and 4.
In step 1, the respondents’ sex explained approximately 2% of the variance in intensity of sibling conflict, F(1, 571) = 9.92, p = .002. Inspection of the standardized regression coefficient (β) indicates that females reported significantly more intense sibling conflict than males. In step 2, sibling sex, age difference between siblings, and length of co-residence explained an additional 13% of the variance in intensity of sibling conflict, F(3, 568) = 29.23, p < .001. Inspection of the standardized regression coefficients (βs) indicates that whereas sibling sex was not a significant predictor, age difference and length of co-residence were significant unique predictors of intensity of sibling conflict. The main effect of age difference indicates that as the age difference decreased, intensity of sibling conflict increased. The main effect of co-residence indicates that as siblings’ duration of co-residence increased, so did the intensity of their conflict. In step 3, degree of relatedness did not explain any additional variance in intensity of sibling conflict, F(1, 567) = 2.43, p = .12. In step 4, the interactions between genetic relatedness and the other four predictor variables (respondent sex, sibling sex, age difference, and co-residence) explained an additional 2% of the variance in intensity of sibling conflict, F(4, 563) = 2.54, p = .04. Inspection of the standardized regression coefficients (βs) indicates that whereas the interactions between relatedness and respondent sex, sibling sex, and age difference were not significant predictors, the interaction between relatedness and co-residence was a significant unique predictor of intensity of sibling conflict. As shown in Fig. 2, intensity of sibling conflict significantly increased for half siblings the longer the duration of co-residence (p < .001) whereas frequency of conflict between full siblings was not influenced by duration of co-residence (p = .99). Overall, approximately 17% of the variance in intensity of sibling conflict was explained by the model, F(9,563) = 12.56, p < .001, R2 = .167.

Interaction between genetic relatedness and duration of co-residence on intensity of sibling conflict
Effect of Sex of Sibling, Duration of Co-Residence, and Genetic Relatedness on Sibling Prosocialness
A separate hierarchical linear regression analysis was conducted to test whether (i) there would be more cooperation between same-sex siblings relative to opposite-sex siblings; (ii) there would be more cooperation as duration of co-residence increases; and (iii) there would be more cooperation between full siblings relative to half-siblings. The variables were entered into the model following the same procedure described above (i.e., respondents’ sex was entered in step 1; sex of sibling, absolute age difference, and duration of co-residence were entered in step 2; degree of relatedness was entered in step 3; and the interactions between degree of relatedness with co-residence, respondents’ sex, sex of sibling, and age difference were added in step 4). Results from this analysis are summarized in Table 3.
In step 1, the respondents’ sex explained approximately 1% of the variance in sibling prosocialness, F(1, 571) = 4.57, p = .03. Inspection of the standardized regression coefficient (β) indicates that females reported significantly more sibling prosocialness than males. In step 2, sibling sex, age difference between siblings, and length of co-residence did not explain any additional variance in intensity of sibling prosocialness, F(3, 568) = 1.18, p = .32. In step 3, degree of relatedness explained an additional 1% of the variance in sibling prosocialness, F(1, 567) = 4.74, p = .03. Inspection of the standardized regression coefficient (β) indicates that there was significantly more sibling prosocialness between full siblings than half-siblings. In step 4, the interactions between genetic relatedness and the other four predictor variables (respondent sex, sibling sex, age difference, and co-residence) did not explain any additional variance in sibling prosocialness, F(4, 563) = 1.63, p = .17. Overall, approximately 3% of the variance in sibling cooperation was explained by the final regression model, F(9, 563) = 2.17, p = .02, R2 = .033.
Discussion
The purpose of the current study was to investigate the effect of sibling sex, duration of co-residence, and degree of genetic relatedness on sibling conflict and sibling prosocialness. The results provided support for prediction 1a in that although the intensity of conflict was not influenced by sibling sex, more frequent conflict was reported between same-sex siblings than opposite-sex siblings. However, inconsistent with prediction 1b, there was no evidence that sibling prosocialness varied as a function of sibling sex. The finding of greater conflict between same-sex siblings is consistent with previous studies (Salmon & Hehman, 2015). Same-sex siblings are in competition for more similar resources, perhaps outside the home even more than within, than opposite-sex siblings and so it would be surprising if this factor did not impact frequency of conflict. The lack of an effect on sibling prosocialness suggests that sibling sex does not influence conflict and prosocialness in the same way.
It should also be noted that sex of respondent influenced frequency and intensity of conflict as well as prosocialness with females reporting both more frequent and intense conflict as well as more sibling prosocialness than males. While a number of studies have reported greater conflict between brothers than sisters (Brody et al., 1985; Salmon & Hehman, 2015), in the current study, females (sisters) reported more frequent and intense sibling conflict. This discrepancy may have been partly influenced by the focus in some research on physical conflicts rather than other conflict behaviors (arguing) as males generally report greater physical aggression which is more often directed toward their same-sex siblings (Campione-Barr & Smetana, 2010; Straus et al., 2006). The inclusion of conflict behaviors besides physical aggression in the current study may account for the observed sex differences. It should be noted, though, that some studies have reported no sex differences in levels of sibling conflict (Khan et al., 2020). The observed sex difference in prosocialness is consistent with previous research suggesting more emotional closeness on the part of sisters (Stocker et al., 2020; Tanskanen & Danielsbacka, 2019) which would typically be associated with greater cooperative or prosocial behavior directed toward siblings. Together, the current findings suggest that both sibling conflict and prosocialness are influenced by one’s own sex with females reporting higher levels of both. Birth interval, however, was found to influence reported conflict (both frequency and intensity) but not prosocialness. Whereas the shorter the birth interval (closer in age), the more frequent and intense the reported conflict, the age difference between siblings did not affect prosocialness scores. Shorter birth intervals, like sex of sibling, will increase the likelihood of competition over similar resources (Pollet & Hoben, 2011; Salmon & Hehman, 2014) and hence result in greater levels of reported conflict the closer in age. The current study suggests that mechanisms regulating prosocialness may not be sensitive to age differences between siblings in the same way that mechanisms regulating conflict appear to be.
Predictions 2a and 2b concerned the impact of co-residence on conflict and cooperativeness. Providing support for prediction 2a, we found that although the frequency of conflict was not influenced by duration of co-residence, intensity of conflict increased as the duration of co-residence increased. However, inconsistent with prediction 2b, sibling prosocialness was not significantly influenced by co-residence. A number of studies have reported a relationship between co-residence and conflict or relationship quality (Gyuris et al., 2020; Salmon & Hehman, 2015; Tanskanen & Danielsbacka, 2019) indicating more conflict with greater duration of co-residence and yet better relationship quality with greater duration, at least in the case of half-siblings. Studies looking at altruism toward siblings have viewed co-residence as an indirect cue of relatedness among siblings and reported that co-residence is a predictor of sibling altruism (Bressan et al., 2009), especially in the absence of maternal perinatal association (Sznycer et al., 2016). Co-residence has an impact via exposure, simple opportunities for conflict and cooperation as relationships are built but it may also play a role as a cue of relatedness, activating mechanisms of kin valuation. Findings from the current study suggest this valuation, however, may only influence intensity of conflict and not frequency of conflict or sibling prosocialness. This is discussed further below with regard to the interaction between co-residence and relatedness in terms of intensity of conflict.
Predictions 3a and 3b focused on the effect of degree of relatedness on levels of conflict and cooperation, and both predictions were largely supported. Consistent with prediction 3a, although degree of relatedness did not influence intensity of conflict, more frequent conflict was reported to occur between full siblings than half siblings. Similar results have now been reported in a number of studies (Khan et al., 2020; Salmon & Hehman, 2015; Tanskanen et al., 2016; Tanskanen et al., 2017). Furthermore, the current study found an interaction between co-residence and relatedness on both frequency and intensity of conflict. Longer co-residence increased the frequency and intensity of conflict for half siblings while it had no impact on the levels of conflict for full siblings. It appears that increased co-residence either serves as a cue of relatedness, making half-siblings more like full in terms of their levels of conflict or simply increases the opportunities for conflict to arise, and perhaps associated with decreased parental support from non-co-resident parents. In addition, consistent with prediction 3b, full siblings reported greater prosocialness than half siblings in line with expectations from kin selection theory in that altruistic behavior is more likely to be directed toward closely related individuals. Therefore, findings from the current study suggest that both conflict and prosocialness are influenced by genetic relatedness, with higher levels of each directed towards siblings that are more closely genetically related. Findings also suggest that with greater co-residence, half siblings become more like full in terms of higher levels of conflict but not in terms of higher levels of prosocialness.
Despite the similar impact of sex and relatedness on conflict and cooperation (being female and more related increase both), several other variables predicted conflict but not cooperation including sex of sibling, age difference, and co-residence. In addition, while sibling sex and relatedness predicted frequency of conflict, those factors did not predict intensity of conflict. This suggests that there may be utility in looking at frequency and intensity of conflict as separate constructs in future work. Though our final models were significantly predicting conflict and prosocialness, the models explained more variance in sibling conflict scores than they did in sibling prosocialness scores. This raises a number of interesting questions including what other factors influence both sibling conflict and cooperation that were not included in our study. Clear contenders would include overall quality of sibling relationship or emotional closeness, phenotypic similarity between siblings (Tifferet et al., 2016), and the reproductive value of siblings (which influences the fitness payoff of any investment). It also would be interesting to investigate potential interactions between genetic relatedness and those factors. Future studies might benefit from including a wider range of relevant factors as well as interactions between those factors and genetic relatedness in attempting to explain additional variance in sibling conflict and cooperation.
There were a few limitations in the current study, one of which is that this was a focal sibling self-report, not a within-family study where we would have self-reports from both members of a sibling pair. It would be informative to know whether siblings have similar perceptions of the levels of conflict and cooperation in their relationship as well as how those perceptions may vary as a function of other relevant factors. In addition, while we initially were hoping to be able to include full, half, and step siblings, we only had sufficient numbers of full and half siblings to include in the analyses. Therefore, these results do not speak to the impact of these variables when non-biologically related siblings are added to the mix. We also did not ask whether half siblings were related on the maternal or paternal side. Other studies have suggested that there may be differences between paternal and maternal half-siblings in terms of the probability of co-residence as well as relationship quality (Gyuris et al., 2020), and maternal half-siblings may be over-represented in our sample. Our use of a sibling prosocialness score as an index of sibling cooperation may also raise some questions. Was the different pattern of results for conflict vs cooperation the result of the prosocialness score not quite tapping into sibling cooperation sufficiently or a reflection of different mechanisms influencing these behaviors? What role might reproductive value or the ability to turn resources into fitness play in conflict or cooperation? In future research, we hope to utilize some more ecologically valid measures of cooperation, such as gift giving and child care, to assess this as well as to collect a wider range of relatedness cues to see what they may add in terms of explanatory power to our models and how they may relate to the interaction we found between co-residence and relatedness. We would also like to see more testing of such hypotheses about sibling relationships in non-WEIRD (Western Educated Industrialized Rich Democratic) populations in order to see what patterns of sibling conflict and cooperation generalize across different cultures and what behaviors might be due to local ecological conditions (including resource scarcity).
Conclusion
In summary, our results indicate that mechanisms regulating sibling conflict and sibling cooperation may not be sensitive to the same set of factors. Sibling conflict was predicted by one’s own sex, sex of sibling, birth interval, co-residence duration, and degree of relatedness with greater conflict reported by sisters, those closer in age, those who have co-resided longer, and full-siblings compared to half-siblings. Sibling prosocialness, however, was only predicted by sex and relatedness with females and full siblings reporting greater levels of sibling prosocialness. Together, these findings contribute to the existing literature by (i) adding to the weight of evidence against Hamilton’s rule in terms of more conflict existing between full-siblings rather than half-siblings (replicating findings of Salmon & Hehman, 2015, among others) and (ii) finding a different pattern for cooperation, suggesting that cooperation does follow Hamilton’s rule with greater cooperation between full-siblings versus half-siblings. However, other factors that should be cues to relatedness (such as duration of co-residence) and, as such, should also influence cooperation among kin were not found to influence cooperation in the current study, which is inconsistent with previous work on kin altruism. Therefore, in light of the potential methodological limitations in the current study, the question whether the same factors that influence sibling conflict also influence sibling cooperation or there are indeed different mechanisms shaping conflict versus cooperation still remains. The current study suggests different mechanisms may be involved. More research investigating patterns of conflict and cooperation within families using more ecologically valid cues is necessary to determine whether the two are operating under the same mechanism, sensitive to the same cues, or under different mechanisms.
Availability of Data and Material
Data and measures will be made available upon publication. Deidentified data is available from the authors or on Zenodo.
References
Bressan, P., Colarelli, S. M., & Cavalieri, M. B. (2009). Biologically costly altruism depends on emotional closeness among step but not half or full sibling. Evolutionary Psychology, 7(1), 147470490900700130.
Brody, G. H., Stoneman, Z., MacKinnon, C. E., & MacKinnon, R. (1985). Role relationships and behaviors between preschool-aged and school-age sibling pairs. Developmental Psychology, 21, 124–129.
Buist, K. L., Dekovic, M., Meeus, W., & Van Aken, M. A. G. (2002). Developmental patterns in adolescent attachment to mother, father and sibling. Journal of Youth and Adolescence, 31, 167–176.
Campione-Barr, N., & Smetana, J. G. (2010). “Who said you could wear my sweater?” Adolescent siblings’ conflicts and associations with relationship quality. Child Development, 81, 464–471.
Caprara, G. V., Steca, P., Zelli, A., & Capanna, C. (2005). A new scale for measuring adults’ prosocialness. European Journal of Psychological Assessment, 21(2), 77–89.
Cicirelli, V. G. (1995). Sibling relationships across the life span. Springer.
Coyne, S. M., Jensen, A. C., Smith, N. J., & Erickson, D. H. (2016). Super Mario brothers and sisters: Associations between coplaying video games and sibling conflict and affection. Journal of Adolescence, 47, 48–59.
Dunn, J., & Munn, P. (1985). Becoming a family member: family conflict and the development of socialunderstanding in the second year. Child Development, 56, 764–774.
Emlen, S. T. (1997). The evolutionary study of human family systems. Social Science Information, 34, 563–589.
Gyuris, P., Kozma, L., Kisander, Z., Láng, A., Ferencz, T., & Kocsor, F (2020). Sibling relations in patchwork families: co-residence is more influential than genetic relatedness. Frontiers in Psychology, 11. https://doi.org/10.3389/psyg.2020.00993
Hamilton, W. D. (1964). The genetic evolution of social behavior, I and II. Journal of Theoretical Biology, 7, 1–52.
Hodge, S. J., Manica, A., Flower, T. P., & Clutton-Brock, T. H. (2008). Determinants of reproductive success in dominant female meerkats. Journal of Animal Ecology, 77, 92–102.
Holmes, W. G., & Sherman, P. W. (1982). The ontogeny of kin recognition in two species of ground squirrels. American Zoologist, 22, 491–517.
Jankowiak, W., & Diderich, M. (2000). Sibling solidarity in a polygamous community in the USA: unpacking inclusive fitness. Evolution and Human Behavior, 21, 125–139.
Jensen, A. C., Whiteman, S. D., & Fingerman, K. L. (2018). “Can’t live with or without them:” transitions and young adults’ perceptions of sibling relationships. Journal of Family Psychology, 32(3), 385–395.
Khan, R., Brewer, G., & Archer, J. (2020). Genetic relatedness, emotional closeness and physical aggression: a comparison of full and half sibling experiences. Europe’s Journal of Psychology, 16(1), 167–185.
McGuire, S., Manke, B., Eftekhari, A., & Dunn, J. (2000). Children’s perceptions of sibling conflict during middle childhood: issues and sibling (dis)similarity. Social Development, 9(2), 173–190.
McHale, S. M., Updegraff, K. A., & Whiteman, S. D. (2012). Sibling relationships and influences in childhood and adolescence. Journal of Marriage and the Family, 74, 913–930.
Neyer, F. J., & Lang, F. R. (2003). Blood is thicker than water: kinship orientation across adulthood. Journal of Personality and Social Psychology, 84, 310–321.
Pollet, T. V. (2007). Genetic relatedness and sibling relationship characteristics in a modern society. Evolution and Human Behavior, 28, 176–185.
Pollet, T. V., & Hoben, A. D. (2011). An evolutionary perspective on siblings: Rivals and resources. In C. Salmon & T. K. Shackelford (Eds.), The Oxford Handbook of Evolutionary Family Psychology (pp. 128–148). Oxford University Press.
Reed, B. W., Segal, N. L., Miller, S. A. (2021). Evolutionary perspective on decreases in grief intensity for deceased twin and non-twin relatives: An update. Personality and Individual Differences, 178. https://doi.org/10.1016/j.paid.2021.110822
Salmon, C., & Hehman, J. (2015). Evolutionary perspectives on the nature of sibling conflict: the impact of sex, relatedness, and co-residence. Evolutionary Psychological Science, 1(2), 123–129.
Salmon, C., & Hehman, J. A. (2014). The evolutionary psychology of sibling conflict and siblicide. In T. K. Shackelford & R. D. Hansen (Eds.), The evolution of violence (pp. 137–157). Springer.
Segal, N. L. (1999). Entwined lives: Twins and what they tell us about human behaviour. Penguin Books.
Segal, N. L. (2005). Evolutionary studies of cooperation, competition, and altruism: a twin-based approach. In R. L. Burgess & K. B. MacDonald (Eds.), Evolutionary perspectives on human development (2nd ed., pp. 265–304). Sage.
Steinbach, A., & Hank, K. (2018). Full-, half-, and step-sibling relations in young and middle adulthood. Journal of Family Issues, 39(9), 2639–2658.
Stocker, C. M., Gilligan, M., Klopack, E. T., Conger, K. J., Lanthier, R. P., Neppl, T. K., & Wickrama, K. A. S. (2020). Sibling relationships in older adulthood: Links with loneliness and well-being. Journal of Family Psychology, 34(2), 175–185.
Straus, M., Gelles, R. J., & Steinmetz, S. K. (2006). Behind closed doors: Violence in the American family. Transaction Publishers.
Sulloway, F. J. (1996). Born to rebel: Birth order, family dynamics, and creative lives. Pantheon.
Sznycer, D., De Smet, D., Billingsley, J., & Lieberman, D. (2016). Coresidence duration and cues of maternal investment regulate sibling altruism across cultures. Journal of Personality and Social Psychology, 111(2), 159–177.
Tanskanen, A. O., & Danielsbacka, M. (2019). Relationship quality among half siblings: the role of childhood co-residence. Evolutionary Psychological Science, 5(1), 13–21.
Tanskanen, A. O., Danielsbacka, M., Jokela, M., David-Barrett, T., & Rotkirch, A. (2016). Diluted competition? Conflicts between full-and half-siblings in two adult generations. Frontiers in Sociology, 1, 6. https://doi.org/10.3389/fsoc.2016.00006
Tanskanen, A. O., Danielsbacka, M., Jokela, M., & Rotkirch, A. (2017). Sibling conflicts in full-and half-sibling households in the UK. Journal of Biosocial Science, 49, 31–47.
Tibbetts, G., & Scharfe, E. (2015). Oh, brother (or sister)!: an examination of sibling attachment, conflict, and cooperation in emerging adulthood. Journal of Relationships Research, 6(8), 1–11.
Tifferet, S., Pollet, T., Bar, A., & Efrati, H. (2016). Predicting sibling investment by perceived sibling resemblance. Evolutionary Behavioral Sciences, 10(1), 64–70.
Trivers, R. L. (1972). Parental investment and sexual selection. In B. Campbell (Ed.), Sexual section and the descent of man (pp. 136–179). Aldine Publishing Company.
Trivers, R. L. (1974). Parent-offspring conflict. American Zoologist, 14, 249–264.
Wahaj, S. A., Van Horn, R. C., Van Horn, T. L., Dreyer, R., Hilgris, R., Schwarz, J., & Holekamp, K. E. (2004). Kin discrimination in the spotted hyena (Crocuta crocuta): Nepotism among siblings. Behavioral Ecology and Sociobiology, 56(3), 237–247.
White, L. K., & Riedmann, A. (1992). Ties among adult siblings. Social Forces, 71, 85–102.
White, L. K., & Riedmann, A. (1992). When the Brady bunch grows up: Step/half- and full sibling relationships in adulthood. Journal of Marriage and the Family, 54, 197–208.
Author information
Authors and Affiliations
Contributions
Both authors contributed equally to this research project.
Corresponding author
Ethics declarations
Ethics Approval
Survey was approved by the University of Redlands Institutional Review Board.
Consent to Participate
All participants provided informed consent.
Consent for Publication
All participants provided informed consent.
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Salmon, C.A., Hehman, J.A. Good Friends, Better Enemies? The Effects of Sibling Sex, Co-Residence, and Relatedness on Sibling Conflict and Cooperation. Evolutionary Psychological Science 7, 327–337 (2021). https://doi.org/10.1007/s40806-021-00292-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40806-021-00292-y