
Abstract
Eric Davidson had a deep and abiding interest in the role developmental mechanisms played in generating evolutionary patterns documented in deep time, from the origin of the euechinoids to the processes responsible for the morphological architectures of major animal clades. Although not an evolutionary biologist, Davidson’s interests long preceded the current excitement over comparative evolutionary developmental biology. Here I discuss three aspects at the intersection between his research and evolutionary patterns in deep time: First, understanding the mechanisms of body plan formation, particularly those associated with the early diversification of major metazoan clades. Second, a critique of early claims about ancestral metazoans based on the discoveries of highly conserved genes across bilaterian animals. Third, Davidson’s own involvement in paleontology through a collaborative study of the fossil embryos from the Ediacaran Doushantuo Formation in south China.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Eric Davidson was a developmental biologist, not an evolutionary biologist or paleobiologist. He was driven to understand the mechanisms of gene regulatory control and how they controlled development, but this focus was deeply embedded within concerns about the relationship between development and evolution. Questions about the origin of major metazoan architectures or body plans were central to Eric’s concerns since at least the late 1960s. His 1971 paper with Roy Britten includes a section on “The Evolutionary Growth of the Genome” illustrated with a figure depicting variations in genome size in major animal groups and a metazoan phylogeny (Britten and Davidson 1971). A major focus of the latter part of the paper was “A proposal for the origin of evolutionary novelty”, which applied the Britten–Davidson model of gene regulatory control (Britten and Davidson 1969) to the formation of new structures such as organs within the developing embryo. These early models of gene regulatory networks focused on a putative regulatory role for repetitive DNA that disappeared in later work, but the importance of the connection between developmental mechanisms and evolution was well established.
The deep history of echinoids specifically, and animals more generally, is essential to understanding Davidson’s research program and his contributions to developmental biology. Comparative evolutionary developmental biology, or “evo-devo” has developed rapidly as a research program since the discovery of the extent of highly conserved developmental genes in the 1990s. Prior to these discoveries few developmental biologists predicted, or even considered, that eyes, hearts or appendages across bilaterian animals were generated by homologous and deeply conserved regulatory genes. Davidson’s interests in similarities in developmental mechanisms across bilaterians arose much earlier from his comparative studies of developmental mechanisms. He was among the pioneers in evo-devo, although he much preferred the term developmental evolution, or devo-evo.
In this contribution I examine three different facets of Davidson’s research program and their relationship to his goal of elucidating the processes of evolutionary change in developmental mechanism. I highlight three components of this program: First, his career-long interest in applying his growing understanding of the nature of gene regulation to the problem of the origin of metazoan body plans; second, what I see as a critique of many aspects of evo-devo as practitioners relied upon gene expression studies from the late 1990s onward (and in which I collaborated with Eric); finally, Davidson also engaged more directly with the fossil record through studies of remarkably preserved 600 million year old (Ma) fossil embryos and through efforts to test hypotheses about changes in regulatory patterning by re-engineering the split between cidaroids and euechinoids about 260 million years ago (Ma), applying what he termed synthetic experimental evolution.
2 Understanding the mechanisms of body plan formation
The relationship between evolutionary novelty and changes in gene regulation were raised in Britten and Davidson’s longer discussion (Britten and Davidson 1971) of their model of gene regulation (Britten and Davidson 1969). This model relied upon a regulatory function for repetitive DNA but reveals the importance that Davidson placed on evolutionary issues early in his career. In the 1971 paper they discussed the importance of genomic rearrangements for the formation and modification of regulatory networks, including saltatory replication, the diffusion of regulatory sequences via chromosomal rearrangements, and localized changes. From a largely conceptual standpoint, Britten and Davidson focused on new organ systems as a form of evolutionary novelty. Neither animal body plans nor deep time were considered. The primary focus of Davidson’s work at this time was mechanistic developmental biology as exemplified by his incredibly detailed syntheses in the three editions of his Gene Activity in Early Development. Although the 3rd Edition (Davidson 1986), for example, contains a wealth of potentially useful comparative developmental data there was little effort to apply it to evolutionary questions. The absence of an evolutionary context in these books is particularly remarkable when they are compared with the three books that Davidson produced from 2001 to 2015: Each has the word evolution in the subtitle (Davidson 2001, 2006; Peter and Davidson 2015).
Britten and Davidson’s model of gene regulation had impact on evolutionary thinking. Valentine and Campbell identified three major modes of evolutionary change in regulatory control: (1) changes in protein-coding and downstream regulatory genes; (2) repatterning of regulatory control of existing genes; and (3) the “expansion of the regulatory apparatus”. While acknowledging that all three modes were likely to operate during the formation of “a wholly new animal type” they suggested: “growth of the regulatory genome may usually predominate during evolution of new grades of complexity; gene repatterning may predominate during the radiation of major new variations within grades; and structural gene frequency changes may be especially important during phyletic evolution within species lineages” (Valentine and Campbell 1975, p. 680). This prescient, hierarchical view of developmental evolutionary changes would largely be born out by later research.
During the 1980s and early 1990s Davidson and his laboratory group focused on developmental mechanisms (Rothenberg 2016) resulting in a series of reviews articulating his views of the important types of development (Davidson 1989, 1990, 1991). The lab also began intensive investigation of the regulation of several genes, particularly the structure of the endo16 gene and its associated regulatory elements (Yuh et al. 2001; Davidson et al. 2002).
Evolution re-emerged as an important thread in Davidson’s work in the early 1990s when Kevin Peterson moved to CalTech as a post-doc. Peterson was originally trained as a paleontologist at UCLA but had already begun focusing on the evolution of development. By this time Davidson had begun to focus on the origin of animal body plans as a compelling evolutionary problem where work on Gene Regulatory Networks (GRNs) could provide critical insights. Together they applied the insights from Davidson’s articulation of types of development to the issue of the origin of animal body plans (Davidson et al. 1995; Peterson et al. 1997; Peterson et al. 2000a, b; Peterson and Davidson 2000). In particular, they argued that what they termed “Type 1 embryogenesis” corresponded to the earliest stages of bilaterian animal evolution. It is worth quoting their argument at length because I think these early views remained central to Davidson’s view of early animal evolution for the remainder of his career:
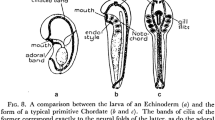
We think that the immediate developmental products of this basic but elegant process of embryogenesis may resemble in their cellular organization the earliest types of animal, those that were present long before animals evolved. These products are the marine larvae of modern indirectly developing species. Such larvae are small, free-living organisms usually less than 1mm across. They consist of only a few thousand cells, and they generate only a very modest repertoire of differentiated cell types. Usually they have a few neurons, a few muscle cells, a functional gut with mouth and anus, and a ciliated epidermis… A good example of a species that develops by this entirely indirect process is the familiar sea urchin. (Cameron et al. 1998; pp. 614–615)
In what they termed “maximally indirect development” the adult develops from a small rudiment of undifferentiated cells which are set aside from involvement in larval embyogenesis. The view of animal origins advanced by Cameron, Davidson and Peterson postulated that adult bilaterian body plans, whether of arthropods, chordates or echinoderms, were appended to ancestral larval-like forms. Although based primarily on his view of larval morphology, Nielsen’s scenario for the early evolution of animals invokes a very similar eumetazoan ancestor: a holopelagic, planktotrophic larvae (Nielsen 2013). The Davidson group’s hypothesis asserts that the broad distribution of indirect development across bilaterians reflects the ancestral metazoan condition, rather than Type I development arising independently in different clades. As discussed in the following two sections, Davidson’s views of early animal evolution were informed by this framework. Type I embyogenesis is key to one of the chapters in his 2006 volume (Davidson 2006) and remains an important intellectual scaffold (see, for example his concluding remarks to Chapter 4). In his final book the ideas return as Mode 1 and Mode 2 embryonic processes in Chapter 3 of Genomic Strategies for Embryonic Development (Peter and Davidson 2015).
This work coincided with the discovery of deep homologies among highly conserved transcription factors and signaling pathways, most famously the Hox and Pax genes. The extensive conservation of transcription factors and signaling molecules led to attempts to infer the ancestral morphology of early animals, with particular attention to earliest metazoans and the last common ancestor of all bilaterians (variously termed the “Urbilaterian” and the “protostome-deuterostome ancestor”, or PDA) (Shenk and Steel 1994; Scott 1994; De Robertis and Sasai 1996). By 2001 this had culminated in the identification of extensive morphological homologies across the Bilateria (Carroll et al. 2001). Carroll and colleagues wonderful introduction to evo-devo reconstructed a maximally complex PDA, with a head, heart, segmentation, proximo-distal and dorsal–ventral patterning, an image-forming eye and other features. Whether or not the PDA had appendages or a centralized nervous system was a continuing source of controversy (Arendt et al. 2008; Lichtneckert and Reichert 2005; Pueyo and Couso 2005; Lowe et al. 2006). Although Davidson was intensely interested in the relationship between development and evolution, it is important to differentiate his work from the main threads of evo-devo. Much of the work in evo-devo at this time involved identifying highly conserved genes and then comparing expression profiles in various clades. Davidson did not see this as sufficiently mechanistic, nor involved with understanding the role of GRNs. Thus despite the excitement about evo-devo Davidson did not see himself as part of this research agenda. (Davidson had a valid point about the difference in research agendas, but it also served his purposes to rail against any prevailing orthodoxy. Several times he complained to me about systems biology, although many saw him as one of the founders of the field.) In later conversations Davidson embraced the term devo-evo to distinguish what he saw as “good science” from some of the enthusiasms of the early years of evo-devo.
During the early 2000s new techniques enabled Davidson’s lab to progress rapidly in the elucidation of early development in Strongylocentrotus. This work began to identify common properties of developmental GRNs. Used in a comparative phylogenetic framework such information became an important tool to understand the evolution of GRNs over time. While in Davidson’s lab, Veronica Hinman compared the emerging sea urchin endomesoderm network to that for starfish (Hinman et al. 2003) to determine what components had been preserved over the c. 500 myr since the two clades diverged. The most surprising result was the conservation of a subnetwork of regulatory genes responsible for gut formation. The genes krox, otx, bra, foxa and gatae have the same topological relationship in both clades, and perturbation experiments showed that this recursively wired network of gene interactions is required for gut formation. This and related studies led to the insight that there was a structure to GRNs and that different components of a GRN could differ in their evolutionary lability (Davidson 2006). Later work showed how in echinoids Tbrain had been co-opted from control of otx, as in starfish to control the larval skeletogenic system (Hinman et al. 2007).
Davidson began to lay this out as a hierarchical relationship of the regulatory genome in his 2006 book The Regulatory Genome (Davidson 2006). He and I, in a paper published the same year, explored this in more detail in our explorations of the importance of GRN structure to the evolution of body plans. We suggested that GRNs comprised four different components: (1) recursively wired subcircuits of genes responsible for patterning parts of the developing embryo, which we described as kernels; (2) small subcircuits that are easily co-opted to form particular developmental roles (such as Notch), which we termed “plug-ins”; (3) switches which activated or deactivated particular subcircuits, which acted as input/output (I/O) switches in the GRN; and (4) the downstream differentiation gene batteries (Davidson and Erwin 2006). The highly conserved network subcircuit found in the endoderm of starfish and sea urchins served as the paradigmatic example of a kernel, and much of the paper discussed the evolutionary significance of this discovery. The recursively wired kernels defined the spatial domains of a particular region of the developing embryo, and the subcircuits are not reused elsewhere (although the individual genes often are deployed elsewhere during development). Modification of the regulatory relationships of genes within the kernel would lead to failure of the phenotype in that part of the developing embryo (Davidson and Erwin 2006; Erwin and Davidson 2009). A similar kernel had previously been described for heart specification across bilaterians. At the same time Wagner described the same recursively wired networks as character homology identity networks (CHiNs) (Wagner 2007), which played an important role in his views of homology and evolutionary novelty (Wagner 2014).
I had already become interested in how the process of evolution had itself changed over time. Since I was trained as a geologist this immediately brought to mind discussions of uniformitarianism, and I realized that most of evolutionary theory was essentially uniformitarian, assuming that the process of evolution had not changed over time (Erwin 2011). The recognition of kernels, however was an empirical demonstration of how developmental variability changed with the increasing complexity of GRNs. At the end of the 2006 paper we proposed that the four different components we had identified, kernels, plug-ins, I/O switches and differentiation gene batteries, differed greatly in their evolutionary lability (Davidson and Erwin 2006). We also suggested that the Linnaean hierarchy of taxonomic ranks was an imperfect reflection of differences in the evolutionary lability of different parts of developmental GRNs and the generation of evolutionary constraints.
The work of Davidson and his research group generated a more explicitly mechanistic view of how changes in developmental processes played out in evolution, particularly in deep time, than was true of much of evo-devo. Two issues in particular stand out: First, the articulation of a strong view of the nature of ancestral bilaterians, and second the hierarchical view of GRN structure, with varying evolutionary lability of different components.
3 Critique of evo-devo
Davidson’s research program on the mechanisms of body plan formation and his view of the nature of the last common bilaterian ancestor (LCBA) led naturally to a critique of other research programs. The LCBA was naturally of considerable interest to paleontologists as well, and with some colleagues I had sought to bring a paleontological perspective to discussions (Valentine et al. 1999; Erwin et al. 1997). We argued that if the PDA were as morphologically complex as inferred by Carroll and many others (see preceding section), and was larger than about 1 cm, it would have necessarily left trace fossil evidence in the fossil record, and thus could not have been older than latest Ediacaran age, or about 550 Ma. When Davidson and I met in the late 1990s our early discussions focused on this issue. Together we extended the argument that he had made with Peterson and challenged the inference that highly conserved genes, and even some of the regulatory interactions, were necessarily evidence for conservation (homology) of the morphological structures.
In a 2002 paper Davidson and I reviewed the evidence from the fossil record, phylogenetics and from studies of highly conserved genes, and distinguished between the differentiation of cell types and the generation of morphological structures through morphogenesis (Erwin and Davidson 2002). Based largely on the views in Davidson’s 2001 book Genomic Regulatory Systems (Davidson 2001), we proposed that the generation of many cell types occurred early in metazoan history, associated with organisms with diverse, and possibly multipotent, cell types, with proximo-distal and anterior–posterior patterning, but with fewer morphogenetic patterning systems. The argument is encapsulated in the following quotation:
The evolution of given body parts probably began with the installation of cell differentiation programs to deploy specific cell types in a certain position in an organism, initially in a very simple morphological context. Later in evolution the transcriptional regulators of these differentiation gene batteries would have been coopted for use in increasingly complex, clade-specific programs of gene regulation that control pattern formation processes. These morphogenetic programs have often continued to be utilized at the same location in the embryo. (Erwin and Davidson 2002; p. 3025)
We examined the case of Pax6, by that time frequently used as an example of the conservation of morphogenetic patterning of the eye (Arendt and Wittbrodt 2001). We proposed that the ancestral bilaterian role of the gene was in controlling genes encoding visual pigments, and because of this role it was later co-opted independently in various bilaterian lineages to control eye development. From this alternative view, the PDA was a much less complex organism. The subsequent identification of many highly conserved genes in cnidarians (Miller et al. 2005) lent support to this argument.
There is now far more comparative data on the phylogenetic distribution of highly conserved developmental genes, including many found in cnidarians and sponges. Some of these genes also have comparative expression data, although relatively few have established GRN information. A recent review largely confirmed the hypothesis advanced in 2002 that the origin of characteristic cell types preceded the formation of morphogenetic systems (Tweedt and Erwin 2015).
4 The search for early organisms
In 1998 Shuhai Xiao, Yun Zhang and Andy Knoll published a paper in Nature reporting Scanning Electron Microscope (SEM) photographs of fossil algae and possible fossil embryos from the Ediacaran-age Doushantuo Formation in southern China (Xiao et al. 1998), which was perhaps the highest-profile of several papers that year establishing the preservation of cellular and subcellular structures. The Doushantuo Formation was already well known to paleontologists for exquisitely preserved fossil material in a phosphate-rich rock. Since the unit dated to the interval immediately preceding the explosion of animal diversity in the Cambrian, the announcement of fossil embryos received considerable attention. Until a year or so earlier, paleontologists had not realized that embryos could be preserved in the fossil record. The work of Xiao, Knoll and their colleagues promised new insights into the earliest phases of animal evolution and the paper linked their discoveries to the work on early bilaterians by Davidson and colleagues. They wrote:
Davidson et al. have … [hypothesized] that animals not only originated but underwent substantial early cladogenesis as minute, little-differentiated metazoans similar in form, function and ontogeny to the larvae of living invertebrates. Only later, after developmental toolkits and physiological tolerances had been well established, did macroscopic size and the adult body plans of extant phyla evolve within already discrete clades. (Xiao et al. 1998; p. 557)
Xiao et al. described cleaving embryos and inferred either direct or lecithotrophic larval development. The fossil embryos from the Doushantuo appeared to bear out the hypothesis laid out by Davidson, Peterson and colleagues. Later in 1998 a meeting was held in Kunming on the origin of animal body plans, at which the Doushantuo embryos were a featured part of the discussion. (Unbeknownst to the paleontologists, the meeting was partly supported by a creationist group and several American creationists attended. Eric never had much patience for intellectual dishonesty and consequently discussions at the meeting were rather spirited.) This meeting led to collaboration among Davidson, Chen Junyuan, one of the organizers of the meeting and a paleontologist from the Nanjing Institute of Geology and Palaeontology, and David Bottjer of USC. With funding from the NASA Astrobiology program Davidson, Bottjer, Chen and other colleagues and students developed a long-term research program on the Doushantuo embryos. (Bottjer has recounted the history of this collaboration; see Bottjer 2016). This group produced a continuing series of papers exploring the nature of these embryos and their possible implications for the evolution of development and for the early history of animals (Chen et al. 2000, 2002, 2004, 2010; Dornbos et al. 2005).
Long before 1998, the Doushantuo fossil material had been well known from studies of algae and microfossils. In their 1998 paper Xiao and Knoll etched fossils out of the rock matrix using dilute acid and produced the exquisite SEM images that attracted so much attention. By 2006 paleontologists began using synchrotron X-ray tomographic microscopy to produce stunning three-dimensional reconstructions which revealed the internal structure and the arrangement of the dividing cells (Donoghue et al. 2006).
The Caltech group was one of several working on the Doushantuo embryos in the early 2000s. Xiao continued his research at Virginia Tech and a strong group developed at Bristol. Participants in the different research groups had different strengths. Davidson’s co-leadership of the Caltech-Nanjing group with Chen, and the participation of his post-doc Paola Oliveri and his colleague Andy Cameron, brought considerable insight into larval biology and developmental processes. The other groups had greater experience in evaluating the preservational issues associated with the embryos and these differing perspectives often led to very different interpretations. For example, in 2004 Davidson and his colleagues described what they viewed as the oldest bilaterian organism—a minute form they named Vernanimalcula (Chen et al. 2004). This fossil, if the report were confirmed, would predate other fossil evidence for bilaterians by about 50 myr, and would support suggestions that the earliest bilaterians were small, relatively simple forms (Erwin and Davidson 2002). However, the announcement of Vernanimalcula was hugely controversial, with important questions raised about the extent of diagenetic changes to the original structures (Bengtson and Budd 2004; Bengtson et al. 2012). At this point there is relatively little support for Vernanimalcula as an early bilaterian.
The work continues, however. By 2015 Davidson and Bottjer were collaborating with Maoyan Zhu who had taken over leadership of the late Proterozoic–Cambrian research group at Nanjing after Chen retired. They published a fascinating description of what might be the oldest well-preserved early sponge (Yin et al. 2015) and further work is in progress.
Davidson’s interests in body plan formation were particularly focused on echinoderms, not surprisingly. With the release of the Strongylocentrotus genome in 2006 Davidson began thinking about combining data from the fossil record, the growing understanding of developmental genes and regulatory patterning, and phylogenetic information to address the origin of key developmental genes. Borrowing from an earlier paper (Birnbaum et al. 2000), Davidson and his colleagues addressed the origin of the echinoderm stereom, the distinctive skeletal structure of the clade. With the Strongylocentrotus genome available they identified the key biomineralization genes and proposed that comparative genomic data couple be used to date the origin of such genes (Bottjer et al. 2006). Two years later Feng Gao and Davidson described the co-option of the adult skeletonization GRN into the micromeres of the developing sea urchin embryo to form the larval skeleton (Gao and Davidson 2008), a feature which distinguishes the euechioids from their sister clade, the cidaroids. Jeff Thompson, a student of Bottjer’s and a member of the research group also identified some older cidaroid fossils in museum collections, pushing back the split between the cidaroids and the euechinoids (Thompson et al. 2015). A comparative analysis of the skeletonegenic genes across the five living classes of echinoderms allowed a reconstruction of the ancestral structure of the GRN kernel (Erkenbrack and Davidson 2015; Erkenbrack et al. 2016). The summer before Davidson died, he, Bottjer and I wrote an initial research proposal to use this information to experimentally ‘rewire’ a cidaroid genome to test whether this data actually was sufficient to explain the morphological differences between the two clades.
5 Davidson’s impact
No evolutionary biologist would confuse Davidson with a member of the clan—he was a mechanistic, molecularly focused developmental biologist and he occasionally exhibited lacunae in his understanding of evolutionary biology. But he had a deep appreciation for the importance of evolution and for the role of deep time in understanding evolution. The tens and hundreds of millions of years of evolution of echinoderms and animals in general provided him with a much broader canvas upon which to think about developmental mechanisms and their influence on the history of life. Davidson was fond of the following syllogism: (1) The morphology of an organism is the product of development; (2) development is the result of the activity of gene regulatory networks; (3) therefore, evolution is a consequence of changes in the structure of gene regulatory networks. One was tempted to say: “Well, yes, but…” To him GRNs were more than sufficient to explain all the interesting aspects of evolution (he would occasionally acknowledge that there were other aspects to evolution, such as microevolutionary adaptation, behavior,… but dismiss them as of little interest). Consequently, while he acknowledged, at least in principle, the importance of the ecological dimensions of animals, Davidson fundamentally held that most of the interesting aspects of evolutionary change involved developmental mechanisms. He recognized that the developmental mechanisms which generated the phenotype had a far greater influence on evolutionary processes than was encompassed by the mathematical models of standard population genetics.
Although I do not think I recognized this when our collaboration began, I came to realize that we were approaching macroevolutionary problems in a very different manner from many other paleobiologists. Although quite a few paleobiologists were fascinated by the evo-devo revolution, macroevolutionary theory largely focused on the differential success of species and clades once they had formed, rather than on sources of macroevolutionary variation. The intriguing, and as yet unresolved challenge is whether evolutionary changes in GRN structure reveal changes in the supply of morphological variation that challenge standard views of microevolution and macroevolution.
References
Arendt, D., Denes, A. S., Jekely, G., & Tessmar-Raible, K. (2008). The evolution of nervous system centralization. Philosophical Transactions of the Royal Society, B, 363, 1523–1528.
Arendt, D., & Wittbrodt, J. (2001). Reconstructing the eyes of Urbilateria. Proceedings of the Royal Society London, Series B, 356, 1545–1563.
Bengtson, S., & Budd, G. (2004). Comment on “Small bilaterian fossils from 40 to 55 million years before the Cambrian”. Science, 306, 1291a.
Bengtson, S., Cunningham, J. A., Yin, C. Y., & Donoghue, P. C. J. (2012). Merciful death for the “earliest bilaterian,” Vernanimalcula. Evolution & Development, 14, 421–427.
Birnbaum, D., Coulier, F., Pebusque, M.-J., & Pontarotti, P. (2000). “Paleogenomics”: Looking in the past into the future. Journal of Experimental Zoology (Molecular and Developmental Evolution), 288, 21–22.
Bottjer, D. J. (2016). Eric Davidson’s career as a paleontologist. Developmental Biology, 412, S38–S40. doi:10.1016/j.ydbio.2016.01.026.
Bottjer, D. J., Davidson, E. H., Peterson, K. J., & Cameron, R. A. (2006). Paleogenomics of echinoderms. Science, 314, 956–960.
Britten, R. J., & Davidson, E. H. (1969). Gene regulation for higher cells: A theory. Science, 165, 349–357.
Britten, R. J., & Davidson, E. H. (1971). Repetitive and non-repetitive DNA sequences and speculation on the origins of evolutionary novelty. Quarterly Review of Biology, 46, 111–138.
Cameron, A. R., Peterson, K. J., & Davidson, E. H. (1998). Developmental gene regulation and the evolution of large animal body plans. American Zoologist, 38, 609–620.
Carroll, S., Grenier, J., & Weatherbee, S. (2001). From DNA to diversity. Malden: Blackwell Scientific.
Chen, J. Y., Bottjer, D. J., Davidson, E. H., Li, G., Gao, F., Cameron, R. A., et al. (2010). Phase contrast synchrotron X-ray microtomography of Ediacaran (Doushantuo) metazoan microfossils: Phylogenetic diversity and evolutionary implications. Precambrian Research, 179, 221. doi:10.1016/j.precamres.2010.03.001.
Chen, J. Y., Bottjer, D. J., Oliveri, P., Dornbos, S. Q., Gao, F., Ruffins, S., et al. (2004). Small bilaterian fossils from 40 to 55 million years before the cambrian. Science, 305, 218–222.
Chen, J. Y., Oliveri, P., Gao, F., Dornbos, S. Q., Li, C. W., Bottjer, D. J., et al. (2002). Precambrian animal life: Probable developmental and adult cnidarian forms from Southwest China. Developmental Biology, 248, 182–196.
Chen, J. Y., Oliveri, P., Li, C.-W., Zhou, G. Q., Gao, F., Hagadorn, J. W., et al. (2000). Precambrian animal diversity: Putative phosphatized embryos from the Doushantuo Formation of China. Proceedings of the National Academy of Sciences, USA, 97, 4457–4462.
Davidson, E. H. (1986). Gene activity in early development (3rd ed.). Orlando, FL: Academic Press.
Davidson, E. H. (1989). Lineage-specific gene-expression and the regulative capacities of the sea-urchin embryo—a proposed mechanism. Development, 105, 421–445.
Davidson, E. H. (1990). How embryos work—a comparative view of diverse modes of cell fate specification. Development, 108, 365–389.
Davidson, E. H. (1991). Spatial mechanisms of gene-regulation in metazoan embryos. Development, 113, 1–26.
Davidson, E. H. (2001). Genomic regulatory systems. San Diego: Academic Press.
Davidson, E. H. (2006). The regulatory genome. San Diego: Academic Press.
Davidson, E. H., & Erwin, D. H. (2006). Gene regulatory networks and the evolution of animal body plans. Science, 311, 796–800.
Davidson, E. H., Peterson, K. J., & Cameron, R. A. (1995). Origin of bilaterian body plans: Evolution of developmental mechanisms. Science, 270, 1319–1325.
Davidson, E. H., Rast, J. P., Oliveri, P., Ransick, A., Calestani, C., Yuh, C. H., et al. (2002). A provisional regulatory gene network for specification of endomesoderm in the sea urchin embryo. Developmental Biology, 246, 162–190.
De Robertis, E. M., & Sasai, Y. (1996). A common plan for dorsoventral patterning in bilateria. Nature, 380, 37–40.
Donoghue, P. C. J., Bengtson, S., Dong, X. P., Gostling, N. J., Huldtgren, T., Cunningham, J. A., et al. (2006). Synchrotron X-ray tomographic microscopy of fossil embryos. Nature, 442, 680–683.
Dornbos, S. Q., Bottjer, D. J., Chen, I. A., Oliveri, P., Gao, F., & Li, C.-W. (2005). Precambrian animal life: taphonomy of phosphatized metazoan embryos from southwest China. Lethaia, 38, 101–109.
Erkenbrack, E. M., Ako-Asare, K., Miller, E., Tekelenburg, S., Thompson, J. R., & Romano, L. (2016). Ancestral state reconstruction by comparative analysis of a GRN kernel operating in echinoderms. Developmental Genes and Evolution, 226, 37–45. doi:10.1007/s00427-015-0527-y.
Erkenbrack, E. M., & Davidson, E. H. (2015). Evolutionary rewiring of gene regulatory network linkages at divergence of the echinoid subclasses. Proceedings of the National Academy of Sciences, USA, 112, E4075–E4084. doi:10.1073/pnas.1509845112.
Erwin, D. H. (2011). Evolutionary uniformitarianism. Developmental Biology, 357, 27–34.
Erwin, D. H., & Davidson, E. H. (2002). The last common bilaterian ancestor. Development, 129, 3021–3032.
Erwin, D. H., & Davidson, E. H. (2009). The evolution of hierarchical gene regulatory networks. Nature Reviews Genetics, 10, 141–148. doi:10.1038/nrg2499.
Erwin, D. H., Valentine, J. W., & Jablonski, D. (1997). The origin of animal bodyplans. American Scientist, 85, 126–137.
Gao, F., & Davidson, E. H. (2008). Transfer of a large gene regulatory apparatus to a new developmental address in echinoid evolution. Proceedings of the National Academy of Sciences, USA, 105, 6091–6096.
Hinman, V. F., Nguyen, A. T., Cameron, R. A., & Davidson, E. H. (2003). Developmental gene regulatory network architecture across 500 million years of echinoderm evolution. Proceedings of the National Academy of Sciences, USA, 100, 13356–13361.
Hinman, V. F., Nguyen, A., & Davidson, E. H. (2007). Caught in the evolutionary act: Precise cis-regulatory basis of difference in the organization of gene networks of sea stars and sea urchins. Developmental Biology, 312, 584–595.
Lichtneckert, R., & Reichert, H. (2005). Insights into the urbilaterian brain: Conserved genetic patterning mechanisms in insect and vertebrate brain development. Heredity, 94, 465–477.
Lowe, C. J., Terasaki, M., Wu, M. M., Freeman, R. M., Jr., Runft, L., Kwan, K., et al. (2006). Dorsoventral patterning in hemichordates: Insights into early chordate evolution. PLoS Biology, 4, e291.
Miller, D. J., Ball, E. E., & Technau, U. (2005). Cnidarians and ancestral gene complexity in the animal kingdom. Trends in Genetics, 21, 536–539.
Nielsen, C. (2013). Life cycle evolution: Was the eumetazoan ancestor a holopelagic, planktotrophic gastraea? BMC Evolutionary Biology. doi:10.1186/1471-2148-13-171.
Peter, I. S., & Davidson, E. H. (2015). Genomic control processes. Development and evolution. London: Academic Press.
Peterson, K. J., Arenas-Mena, C., & Davidson, E. H. (2000a). The A/P axis in echinoderm ontogeny and evolution: Evidence from fossils and molecules. Evolution & Development, 2, 93–101.
Peterson, K. J., Cameron, A. R., & Davidson, E. H. (1997). Set-aside cells in maximal indirect development: Evolutionary and developmental significance. BioEssays, 19, 623–631.
Peterson, K. J., Cameron, R. A., & Davidson, E. H. (2000b). Bilaterian origins: Significance of new experimental observations. Developmental Biology, 219, 1–17.
Peterson, K. J., & Davidson, E. H. (2000). Regulatory evolution and the origin of the bilaterians. Proceedings of the National Academy of Sciences, USA, 97, 4430–4433.
Pueyo, J. I., & Couso, J. P. (2005). Parallels between the proximal-distal development of vertebrate and arthropod appendages: Homology without an ancestor? Current Opinion in Genetics & Development, 15(4), 439.
Rothenberg, E. V. (2016). Eric Davidson: Steps to a gene regulatory network for development. Developmental Biology, 412, S7–S19.
Scott, M. P. (1994). Intimations of a creature. Cell, 79, 1121–1124.
Shenk, M. A., & Steel, M. A. (1994). A molecular shapshot of the metazoan ‘Eve’. Trends in Biochemical Science, 18, 459–463.
Thompson, J. R., Petsios, E., Davidson, E. H., Erkenbrack, E. M., Gao, F., & Bottjer, D. J. (2015). Reorganization of sea urchin gene regulatory networks at least 268 million years ago as revealed by oldest fossil cidaroid echinoid. Scientific Reports. doi:10.1038/srep15541.
Tweedt, S. M., & Erwin, D. H. (2015). Origin of metazoan developmental toolkits and their expression in the fossil record. In I. Ruiz-Trillo & A. M. Nedelcu (Eds.), Evolution of multicellularity (pp. 47–77). London: Academic Press.
Valentine, J. W., & Campbell, C. A. (1975). Genetic regulation and the fossil record. American Scientist, 63, 673–680.
Valentine, J. W., Jablonski, D., & Erwin, D. H. (1999). Fossils, molecules and embryos: New perspectives on the Cambrian explosion. Development, 126, 851–859.
Wagner, G. P. (2007). The developmental genetics of homology. Nature Reviews Genetics, 8, 473–479.
Wagner, G. P. (2014). Homology, genes, and evolutionary innovation. Princeton, NJ: Princeton University Press.
Xiao, S. H., Yun, Y., Knoll, A. H., & Bartley, J. K. (1998). Three-dimensional preservation of algae and animal embryos in a Neoproterozoic phosphorite. Nature, 391, 553–558.
Yin, Z. J., Zhu, M. Y., Davidson, E. H., Bottjer, D. J., Zhao, F. C., & Tafforeau, P. (2015). Sponge grade body fossil with cellular resolution dating 60 Myr before the Cambrian. Proceedings of the National Academy of Sciences, USA, 112, E1453–E1460. doi:10.1073/pnas.1414577112.
Yuh, C. H., Bolouri, H., & Davidson, E. H. (2001). Cis-regulatory logic in the endo16 gene: Switching from a specification to a differentiation mode of control. Development, 128, 617–629.
Acknowledgements
Earlier versions of this paper were presented at a workshop on “From Genome to Gene” at the Jacques Loeb Centre for the History and Philosophy of the Life Sciences at Ben Gurion University of the Negev in November 2015, and at a memorial symposium for Eric Davidson at Caltech in April 2016. I appreciate the invitation to contribute this paper from Ute Deichmann and Michel Morange. This paper incorporates research funded by the NASA National Astrobiology Institute.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Erwin, D.H. Eric Davidson and deep time. HPLS 39, 29 (2017). https://doi.org/10.1007/s40656-017-0156-z
Published:
DOI: https://doi.org/10.1007/s40656-017-0156-z