
Abstract
Purpose of Review
This review explores sporotrichosis development as a disease in both cats and humans as well as options for diagnosis and treatment. This work also discusses the factors that might have culminated on the emergence of Sporothrix brasiliensis as the main etiological agent of this disease.
Recent Findings
Sporotrichosis is currently an epidemic in Brazil with cats acting as the primary vector of the disease. And, although molecular diagnostic techniques have been recently developed, the disease remains largely unchecked evidencing the need for novel therapeutic options as well as a more effective public health response.
Summary
It is becoming more evident that to manage and control sporotrichosis, a One Health approach needs to be globally adopted. In addition to that, global warming is creating increasingly favorable conditions to the emergence of fungal pathogens.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Sporotrichosis, caused by the thermodimorphic fungi of the Sporothrix genus, is the most common subcutaneous mycosis worldwide. Although ubiquitous globally in the environment, most fungi from the Sporothrix genus do not usually cause human or animal infections [1]. Until recently, sapronotic transmission was the most common source of human sporotrichosis, especially in North America and Europe, with infection usually beginning after cutaneous trauma related to recreational or occupational activities such as gardening, farming, and mining, giving the disease the epithet of “Rose Gardener’s Disease” [2].
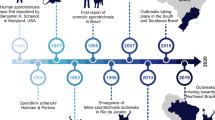
Until the 1990s, Sporothrix schenckii was presumed to be the sole agent of sporotrichosis until recent data defined the pathogenic species as Sporothrix globosa, Sporothrix brasiliensis, Sporothrix luriei, and Sporothrix schenckii sensu stricto [1, 3]. Some of these species have high endemicity in countries such as China, Japan, Australia, India, South Africa, and Brazil [4]. In Asia, particularly in China, S. globosa is the causative agent in 99.3% of cases of human sporotrichosis [5]. S. schenckii is responsible for 94% of cases in Australia and South Africa, and it accounts for 89% of cases in North and South America [6]. In Brazil, however, since the identification of a zoonotic sporotrichosis case transmitted from a cat to a human in 1998, sporotrichosis has emerged as a challenging epidemic in the region [7], with S. brasiliensis as the main etiological agent, accounting for 88% of cases reported in the south and southeast regions of the country, especially in Rio de Janeiro State, where the disease has been considered hyperendemic over the last two decades [3, 8•, 9, 10].
The Sporothrix Genus and the Emergence of S. brasiliensis
The recent emergence of some of these species, S. brasiliensis in particular, has shifted the historic paradigm of sapronotic sporotrichosis transmission, with zoonotic and enzootic infection becoming increasingly common in Brazil [1]. Previous contact with decaying plant material is commonly reported in S. globosa and S. schenckii infections, and sapronotic transmission of S. brasiliensis is possible [1]. S. brasiliensis is generally transmitted via bites, scratches, or direct contact with cutaneous lesions of infected cats [1, 4, 11, 12]. While dogs are generally not considered a significant source of human infections caused by S. brasiliensis, Sporothrix spp. has been isolated from the oral cavity and conjunctival mucosa of dogs, allowing for dog-to-human transmission [1]. Overall, sporotrichosis can be transmitted through various routes, including sapronotic, zoonotic, and animal horizontal transmission (Fig. 1).

Sporotrichosis transmission routes. Sporothrix spp. are commonly found in soil and decomposing plant matter in a mycelial form, which can lead to infections in humans and animals by sapronotic transmission (blue arrows). This occurs when filamentous propagules are traumatically inoculated into the skin. Deep scratches or contact with exudate from the cutaneous lesions of ill cats can transmit the infection horizontally to other animals (red arrows), cats being the most susceptible. Likewise, the infection can reach humans by zoonosis (purple arrows) where high load of yeast is inoculated through scratches, bites, and secretions from cats with sporotrichosis. Human–human transmission is extremely rare, but transmission is possible when daily interactions involve direct contact with injuries (dark gray arrow). The most involved species according to the routes of transmission are shown on the right side of the figure. Original figure, created with BioRender.com
Cat-transmitted sporotrichosis (CTS) has been reported in the American and Asian continents mostly as isolated cases and small outbreaks [11]. Isolated cases of feline sporotrichosis have also been reported in Spain, Japan, and Germany; however, there has been no evidence or report of cat-to-human transmission in these countries [13,14,15]. The situation is different in South America, especially in Brazil, where CTS emerged in the state of Rio de Janeiro in 1998 and currently remains hyperendemic and has begun to spread to other states and South American countries [16•, 17, 18]. Argentina, Paraguay, and Chile are among those countries where CTS has recently emerged [1, 19,20,21]. In 2022, in the UK, the first three cases of CTS due to S. brasiliensis were reported involving mother and daughter of a Brazilian family and the veterinarian of their cat with sporotrichosis, apparently only developed after three years living in the UK [22].
There are a few different factors that could explain the changes in the disease profile and recent emergence of S. brasiliensis and CTS epidemic such as environmental factors (changes in temperature and humidity), evolution, and urbanization coupled with changes in human culture and the instinct cat behavior, as well as an inadequate public health response [21].
Different approaches can help us analyze those factors and help us understand the problem. From a public health point of view, it is important to assess what has been done (or not) in the last two decades, to put sporotrichosis in check in its hyperendemic region. The geographic expansion of CTS cases is partially related to the fact that fungal infections are generally neglected with limited public health policies around the world [21, 23, 24]. Notably, Sporothrix was left off the 2022 World Health Organization Fungal Priority Pathogen List [25]. In Brazil, sporotrichosis is a known problem, but its rise and spread have been poorly addressed leading to an uncontrolled situation [21]. The expansion of the disease is a direct result of socioeconomic difficulties coupled with scarce or inadequate health services. A good example of this is that it took 16 years for the government of Rio de Janeiro, the major CTS endemic area, to implement an animal sporotrichosis control program, with free diagnosis and treatment for animals, and, even then, control measures have remained relatively inefficient with compulsory notification not universally performed and a general absence of educational campaigns to inform the population, leading to an ever-growing number of human and animal cases [21].
Other valid approach to understand why S. brasiliensis emergence is happening now is to explore the environmental and evolutionary point of view, and for that it is necessary to evaluate what differentiates S. brasiliensis to its counterparts and how that correlates to environmental changes in the last two decades. A collection of requirements exists for fungi to act as a primary pathogen [26, 27]. Among those are the ability to invade/bypass host barriers, evade/withstand the immune system, acquire nutrients in the human tissue, and, more importantly for our discussion, the ability to grow at or above host temperature [26, 27]. Not coincidentally, S. brasiliensis exhibits high thermotolerance and can effectively infect both humans and cats, which have a slightly higher body temperature ranging from 38 to 39 °C, whereas other members of the Sporothrix genus like S. globosa do not thrive well above 35 °C [28]. In addition, recent studies with clinical isolates suggest that S. brasiliensis has the capacity to undergo microevolutions within its host, increasing virulence in vivo over the course of infection [29,30,31,32]. These facts alone could help us explain why S. brasiliensis was so successful not only as a primary pathogen, but in its adaptation as a cat pathogen, which in turn collaborates to the spread of the disease, but we believe the correlation between its emergence and thermotolerance is not that simple and passes through environmental factors such as climate change.
S. brasiliensis grows in soil and decaying plant material, with warmer and damp conditions creating more opportunities for the fungus to proliferate and infect humans and animals. These environmental conditions justify why the majority of cases reported are concentrated in regions with tropical weather [33]. Nevertheless, in the last two decades, the number of cases reported in the hyperendemic regions seems disproportional, leading to a belief that global warming triggered the emergence of the thermotolerant species [19, 34,35,36,37].
The combination of endothermy and a complex immune system has been identified as the cause for the relatively high resistance of mammals against fungal infections [34]. Furthermore, the correlation between thermotolerance and virulence in fungal pathogens has been broadly discussed in the literature, as mammals are capable to maintain high body temperatures in comparison to environmental temperatures, which creates a thermally restrictive ambient for the majority of fungi, with all common fungal pathogens sharing the characteristic of being thermotolerant [26, 34,35,36,37].
In 2010, Garcia-Solache and Casadevall hypothesized that global warming is playing a key role in promoting the emergence of new fungal diseases in mammals by both increasing the geographic range of current pathogenic species and also selecting species for adaptive thermotolerance [34]. The emergence and spread of S. brasiliensis in Brazil and South America can be supported by both mechanisms described above. The rise in temperature favors evolution and adaptation of a thermotolerant Sporothrix species, while secondary effects of global warming like the increased rainfall totals help create the perfect environmental conditions to S. brasiliensis to thrive [38]. This, in combination with the large number of stray cats roaming the streets of Rio de Janeiro and other urban areas, created the “perfect storm” scenario to facilitate the emergence and spread of S. brasiliensis.
Pathogenesis
After traumatic inoculation from the environment or a cat, which enables Sporothrix propagules like conidia and mycelial fragments or yeast cells, respectively, to enter the host, the disease usually limits itself to the skin, subcutaneous tissue, and adjacent lymphatic vessels, accounting for the majority of cases reported [39•, 40]. Less frequently, but not rare in the cat transmission scenario, multiple skin lesions may arise from multiple inoculations (scratches and bites), or at the other extreme, skin or mucous lesions may arise after a non-traumatic contact with cats with sporotrichosis (touching exudates or being exposed to cat sneezing) [9, 39•, 41]. Atypical clinical presentations can also occur, especially in immunocompromised individuals or in the rare event of fungal conidia being inhaled from the environment, ranging from pulmonary sporotrichosis to disseminated and meningeal forms of the disease [40, 42,43,44]. Unfortunately, the Brazilian epidemic has had an increase in the number of disseminated infections [10].
The disease has a fairly broad incubation period, ranging from a few days to a few months [45]. Clinical manifestations begin with the development of a small papule or pustule at the inoculation site, which evolves into a nodule, ulcerating or not, involving the skin and usually with additional similar nodular-ulcerative lesions along the ascending lymphatic vessels [45]. This ascending distribution defines the classic lymphocutaneous form and, when there is a single lesion, it is classified as the fixed or localized cutaneous form with systemic symptoms typically absent [45]. The atypical clinical presentations, either extra-cutaneous or systemic, occur following contiguous or hematogenous spread from the primary affected site or from pulmonary infection, both cases more commonly observed in immunocompromised individuals, especially the hematogenous spread [9, 45]. It is worth mentioning that, besides the immune status of the patient, the clinical manifestations of sporotrichosis can vary based on several factors, including the virulence, inoculum site and size, and thermal tolerance of the strain [19].
The concept of virulence refers to the ability of a microbe to cause damage in a host, with injury resulting from the microbial processes, host immune response, or both, leading to a state of disease when homeostasis is disturbed [46]. The Sporothrix genus exhibits several virulence factors such as glycoproteins, secreted proteins, extracellular vesicles, thermotolerance, melanin production, ergosterol peroxide, dimorphism, and the ability to form biofilm in both filamentous and yeast forms [28, 47]. The genus produces a heat shock protein, namely, the chaperone HtpG or HSP90, which is crucial for preserving the yeast form of the fungus [48]. In 2009, Arrillaga-Moncrieff et al. performed a comparative study in the murine model that indicated that the potential virulence of the main human pathogenic species of the Sporothrix genus varies [49], with S. brasiliensis being considered the most virulent, S. globosa the least virulent, and S. schenckii exhibiting an intermediate virulence phenotype, findings that are also supported by epidemiological data [8•, 19, 49].
In humans, sporotrichosis is categorized into cutaneous, mucosal, and extracutaneous forms based on the location of the lesions [39•, 50, 51]. Apart from the cutaneous presentations already described, S. brasiliensis has been associated with ocular involvement, disseminated disease, central nervous system (CNS) disease, and hypersensitivity reactions [52,53,54].
As for cats, which are the primary animal host and vector for human infection, multiple ulcerated skin lesions associated with enlarged lymph nodes and respiratory signs are the most common clinical manifestations of sporotrichosis [55]. The incubation period after infection ranges from three to 30 days [56]. While disseminated sporotrichosis cases in humans are more prevalent among individuals with immunosuppressive conditions, such an association was not found for feline sporotrichosis, as cats diagnosed with feline immunodeficiency virus (FIV) or feline leukemia virus (FeLV) did not exhibit an increased incidence of sporotrichosis [57]. For currently unknown reasons, S. brasiliensis is not easily controlled by the cat immune system, compared to response typical in humans and certain other mammals, such as dogs, leading to high fungal burdens in these susceptible animals.
Diagnosis and Treatment
In a context of hyperendemicity, especially considering areas with little access to mycology laboratories, the diagnosis of sporotrichosis is based on clinical-epidemiological probability. However, ideally, the isolation of the fungus from skin lesion scraping, exudate, or biopsy is the primary microbiologic diagnostic method for sporotrichosis. Other clinical specimens are used according to the affected sites. Biological materials from human skin lesions and other tissue samples present challenges in visualizing small yeast cells due to the low fungal load causing direct microscopic examination (DME) to be ineffective. Giemsa-stained purulent lesion imprints, biopsies, or aspirates enhance sensitivity [8•], and histopathological examinations with periodic acid-Schiff (PAS) or Gomori-methenamine silver (GMS) staining detect granulomas with epithelioid cells, yeast cells (rare in immunocompetent humans), and, in some cases, asteroid bodies [45]. In cases of extracutaneous or disseminated forms, the ideal clinical sample to isolate the fungus for definitive diagnosis will depend on the sites of Sporothrix infection and can include deep biopsies, sputum, cerebrospinal fluid, and blood cultures, for example. Stains like Gram, Giemsa, PAS, and GMS are helpful in these cases. Molecular techniques are considered ideal for epidemiological studies and correctly classify all species. The calmodulin (CAL), β-tubulin, and elongation factor (EF) genes are the main targets used in these techniques [16•, 58, 59]. It is also worth to mention that immunologic tools have been developed in the last two decades, and antibody detection through enzyme-linked immunosorbent assay (ELISA) is available for the presumptive diagnosis of both feline and human sporotrichosis [60,61,62]. Furthermore, nested PCR or quantitative PCR can be used to detect Sporothrix DNA in clinical samples with lower fungal burden such as the cerebrospinal fluid [63••].
Itraconazole is the drug of choice for treating sporotrichosis in humans and animals, but treatment is usually prolonged, especially in cats, with a treatment duration typically spanning from 3 to 6 months or even longer for severe or low-responsive cases [17, 64, 65]. A dosage of 100 to 400 mg per day is used for treating sporotrichosis in humans as it results in clinical improvement in nearly all patients and has an acceptable incidence of adverse effects, ranging from 10 to 40%, mostly mild [66,67,68]. Cats, on the other hand, are often reported as low or unresponsive to itraconazole [69] and little is known about predictors of the treatment response; however, the occurrence of respiratory signs and lesions on nasal mucosa and skin with high fungal loads was associated with treatment failure [70]. If patients are unresponsive to itraconazole, potassium iodide solution or terbinafine can be used as oral treatments for localized cutaneous forms [17]. Antifungal agents that act on ergosterol biosynthesis, such as polyenes, azoles, and allylamines, are also effective treatments. Amphotericin B, a polyene antifungal, is reserved for severe cases, but it can be cardiotoxic and nephrotoxic [71, 72].
Sporothrix species show the potential to develop in vitro resistance to conventional antifungals. S. schenckii [73] and S. globosa [74] have developed in vitro resistance to itraconazole in human infections, S. schenckii in felines [75, 76], and S. brasiliensis in dogs [77]. Recently, some cases of human S. brasiliensis infection caused by strains with reduced in vitro susceptibility to azoles, terbinafine, and/or amphotericin B have been described. These cases require extended treatment times and may develop sequelae [78]. The unequivocal correlation with clinical unresponsiveness and in vitro resistance remains to be proven, since host factors play an important role in the outcomes. Antifungal resistance in Sporothrix is related to the fungus’ ability to produce melanin, low genetic diversity, possibly due to the abnormal number of chromosomes, and single nucleotide polymorphisms in cytochrome P450 or other genes related to antifungal resistance [79, 80]. Sporotrichosis has a well-established therapeutic protocol, but the need for long courses of treatment and the occurrence of feline and human cases unresponsive to the treatment of choice or even with relapses, maybe related to resistant strains or low drug absorption and metabolism, highlight the importance of the development of novel antifungal drugs and therapeutic options, preferably more efficient and less toxic.
Significant research efforts have explored various therapeutic options for treating sporotrichosis. Over the past several years, these efforts have focused on diverse alternatives such as synthetic organic chemical compounds [81,82,83], essential oils [84], plant extracts [85], metal complexes [86, 87], nitric oxide (NO) releasing particles [88], and repositioned drugs [89, 90]. These compounds have exhibited promising results in vitro, as evidenced in Table 1. Moreover, some of these compounds show antifungal efficacy in animal models of sporotrichosis, further highlighting their potential as viable therapeutic options.
Conclusions
It is important to highlight that a One Health approach needs to be adopted globally to prevent and control the epidemic of S. brasiliensis and CTS. This means that surveillance, prevention, and treatment efforts should cover not only people and animals but also the environment, which is particularly crucial in densely populated urban areas where the risk of transmission is higher [95].
The possibility for gaining a better understanding of the transfer of both pathogenic and non-pathogenic microorganisms between humans, animals, and the environment is also of fundamental importance. This is achieved by analyzing evidence of the interactions that take place within this ecosystem [96]. Furthermore, global warming concerns are often focused elsewhere and the potential for its effects on fungal pathogens is broadly overlooked. A warmer climate has the potential to create favorable conditions to thermotolerant species [34], which can lead to a change in their geographic distribution. Adaptation to higher temperatures also has the potential to create conditions that could turn environmental species into mammal pathogens, as for every 1 °C gained in body temperature between 30 and 42 °C, approximately 6% of fungal species are currently excluded as potential pathogens [34, 37], meaning that global warming is particularly dangerous because it narrows down the thermal gradient between environmental conditions and mammalian temperatures [34].
Taking all the facts together, S. brasiliensis poses an especially challenging threat to control. We believe that only a One Health approach, coupled with increased research to enhance our understanding of the disease and to create alternative diagnostics and therapeutics, and implementation of educational campaigns for both health professionals and general population can stop the further spread of sporotrichosis.
Data Availability
Not applicable.
References
Papers of particular interest, published recently, have been highlighted as: • Of importance •• Of major importance
Rossow JA, Queiroz-Telles F, Caceres DH, Beer KD, Jackson BR, Pereira JG, Ferreira Gremião ID, Pereira SA. A One Health approach to combatting Sporothrix brasiliensis: narrative review of an emerging zoonotic fungal pathogen in South America. J Fungi. 2020;6:247.
Rodrigues AM, Hagen F, de Camargo ZP. A spotlight on Sporothrix and sporotrichosis. Mycopathologia. 2022;187:407–11.
Rodrigues AM, de Melo TM, de Hoog GS, Schubach TMP, Pereira SA, Fernandes GF, Bezerra LML, Felipe MS, de Camargo ZP. Phylogenetic analysis reveals a high prevalence of Sporothrix brasiliensis in feline sporotrichosis outbreaks. PLoS Negl Trop Dis. 2013;7: e2281.
Alvarez CM, Oliveira MME, Pires RH. Sporotrichosis: a review of a neglected disease in the last 50 years in Brazil. Microorganisms. 2022;10:2152.
Moussa TAA, Kadasa NMS, Al Zahrani HS, et al. Origin and distribution of Sporothrix globosa causing sapronoses in Asia. J Med Microbiol. 2017;66:560–9.
Zhang Y, Hagen F, Stielow B, et al. Phylogeography and evolutionary patterns in Sporothrix spanning more than 14 000 human and animal case reports. Persoonia. 2015;35:1–20.
Barros MBL, Schubach AO, Schubach TMP, Wanke B, Lambert-Passos SR. An epidemic of sporotrichosis in Rio de Janeiro, Brazil: epidemiological aspects of a series of cases. Epidemiol Infect. 2008;136:1192–6.
Rodrigues AM, Gonçalves SS, de Carvalho JA, Borba-Santos LP, Rozental S, Camargo ZP de. Current progress on epidemiology, diagnosis, and treatment of sporotrichosis and their future trends. J Fungi Basel Switz. 2022;8:776. In this work, Rodrigues and colleagues provide a in depth review of sporotrichosis epidemiology, diagnosis and treatment. They also discuss potential developments of the sporothrix-sporotrichosis system and some of the ways that the disease could be mitigated.
Izoton CFG, de Brito Sousa AX, Valete CM, et al. Sporotrichosis in the nasal mucosa: a single-center retrospective study of 37 cases from 1998 to 2020. PLoS Negl Trop Dis. 2023;17: e0011212.
Bittencourt AA, Oyafuso LKM, Cavalin RF, et al. A neglected disease. Human sporotrichosis in a densely populated urban area in São Paulo, Brazil: clinical–epidemiological and therapeutic aspects. Braz J Microbiol. 2022;53:739–48.
Gremião IDF, Miranda LHM, Reis EG, Rodrigues AM, Pereira SA. Zoonotic epidemic of sporotrichosis: cat to human transmission. PLoS Pathog. 2017;13: e1006077.
Almeida-Paes R, de Oliveira MME, Freitas DFS, do Valle ACF, Zancopé-Oliveira RM, Gutierrez-Galhardo MC. Sporotrichosis in Rio de Janeiro, Brazil: Sporothrixbrasiliensis is associated with atypical clinical presentations. PLoS Negl Trop Dis. 2014;8:e3094.
Gonzalez Cabo JF, de las Heras Guillamon M, Latre Cequiel MV, Garcia de Jalon Ciercoles JA. Feline sporotrichosis: a case report. Mycopathologia. 1989;108:149–154.
Hirano M, Watanabe K, Murakami M, Kano R, Yanai T, Yamazoe K, Fukata T, Kudo T. A case of feline sporotrichosis. J Vet Med Sci. 2006;68:283–4.
Scheufen S, Strommer S, Weisenborn J, Prenger-Berninghoff E, Thom N, Bauer N, Köhler K, Ewers C. Clinical manifestation of an amelanotic Sporothrix schenckii complex isolate in a cat in Germany. JMM Case Rep. 2015;2: e000039.
Falcão EMM, Pires MC de S, Andrade HB, Gonçalves MLC, Almeida-Paes R, do Valle ACF, Bastos FI, Gutierrez-Galhardo MC, Freitas DFS. Zoonotic sporotrichosis with greater severity in Rio de Janeiro, Brazil: 118 hospitalizations and 11 deaths in the last 2 decades in a reference institution. Med Mycol. 2020;58:141–143. In this work, Falcão and colleagues provide a portrait of the development of sporotrichosis in Rio de Janeiro, the area most affected by the disease, in the last two decades. This retrospective search in INI/Fiocruz database, an internationally recognized reference institution, provides valuable insight of the epidemiology of the disease.
Barros MB de L, de Almeida Paes R, Schubach AO. Sporothrix schenckii and sporotrichosis. Clin Microbiol Rev. 2011;24:633–654.
Hay RJ, Morris-Jones R. Outbreaks of sporotrichosis. Curr Opin Infect Dis. 2008;21:119–21.
Etchecopaz A, Toscanini MA, Gisbert A, Mas J, Scarpa M, Iovannitti CA, Bendezú K, Nusblat AD, Iachini R, Cuestas ML. Sporothrix brasiliensis: a review of an emerging South American fungal pathogen, its related disease, presentation and spread in Argentina. J Fungi Basel Switz. 2021;7:170.
Barros MB de L, Schubach TP, Coll JO, Gremião ID, Wanke B, Schubach A. Esporotricose: a evolução e osdesafios de uma epidemia. Rev Panam Salud Pública. 2010;27:455–460.
Gremião IDF, Oliveira MME, Monteiro de Miranda LH, Saraiva Freitas DF, Pereira SA. Geographic expansion of sporotrichosis, Brazil. Emerg Infect Dis. 2020;26:621–4.
Barnacle JR, Chow YJ, Borman AM, Wyllie S, Dominguez V, Russell K, Roberts H, Armstrong-James D, Whittington AM. The first three reported cases of Sporothrix brasiliensis cat-transmitted sporotrichosis outside South America. Med Mycol Case Rep. 2023;39:14–7.
Seyedmousavi S, Guillot J, Tolooe A, Verweij PE, de Hoog GS. Neglected fungal zoonoses: hidden threats to man and animals. Clin Microbiol Infect. 2015;21:416–25.
Rodrigues ML, Nosanchuk JD. Fungal diseases as neglected pathogens: a wake-up call to public health officials. PLoS Negl Trop Dis. 2020;14: e0007964.
WHO fungal priority pathogens list to guide research, development and public health action. https://www.who.int/publications-detail-redirect/9789240060241. Accessed 23 May 2023.
Köhler JR, Hube B, Puccia R, Casadevall A, Perfect JR. Fungi that infect humans. Microbiol Spectr. 2017. https://doi.org/10.1128/microbiolspec.FUNK-0014-2016.
Valdez AF, Miranda DZ, Guimarães AJ, Nimrichter L, Nosanchuk JD. Pathogenicity & virulence of Histoplasma capsulatum - a multifaceted organism adapted to intracellular environments. Virulence. 2022;13:1900–19.
Brilhante RSN, Fernandes MR, Pereira VS, et al. Biofilm formation on cat claws by Sporothrix species: an ex vivo model. Microb Pathog. 2021;150: 104670.
Freitas DFS, Santos SS, Almeida-Paes R, de Oliveira MME, do Valle ACF, Gutierrez-Galhardo MC, Zancopé-Oliveira RM, Nosanchuk JD. Increase in virulence of Sporothrix brasiliensis over five years in a patient with chronic disseminated sporotrichosis. Virulence. 2015;6:112–120.
Oliveira MME, Almeida-Paes R, Corrêa-Moreira D, et al. A case of sporotrichosis caused by different Sporothrix brasiliensis strains: mycological, molecular, and virulence analyses. Mem Inst Oswaldo Cruz. 2019;114: e190260.
Corrêa-Junior D, de Andrade IB, Alves V, et al. Unveiling the morphostructural plasticity of zoonotic sporotrichosis fungal strains: possible implications for Sporothrix brasiliensis virulence and pathogenicity. J Fungi. 2023;9:701.
Corrêa-Junior D, de Andrade IB, Alves V, et al. Metabolic plasticity and virulence-associated factors of Sporothrix brasiliensis strains related to familiar outbreaks of cat-to-human transmitted sporotrichosis. J Fungi Basel Switz. 2023;9:724.
Rabello VBS, Almeida-Silva F, Scramignon-Costa B de S, Motta B da S, de Macedo PM, Teixeira M de M, Almeida-Paes R, Irinyi L, Meyer W, Zancopé-Oliveira RM. Environmental isolation of Sporothrix brasiliensis in an area with recurrent feline sporotrichosis cases. Front Cell Infect Microbiol. 2022;12:894297.
Garcia-Solache MA, Casadevall A. Global warming will bring new fungal diseases for mammals. mBio. 2010;1:e00061–10.
Casadevall A. Fungal virulence, vertebrate endothermy, and dinosaur extinction: is there a connection? Fungal Genet Biol FG B. 2005;42:98–106.
Bergman A, Casadevall A. Mammalian endothermy optimally restricts fungi and metabolic costs. mBio. 2010;1:e00212–10.
Robert VA, Casadevall A. Vertebrate endothermy restricts most fungi as potential pathogens. J Infect Dis. 2009;200:1623–6.
Organization (WMO) WM, World Meteorological Organization (WMO). State of the climate in Latin America and the Caribbean 2021 (WMO-No. 1295). WMO, Geneva; 2022.
Orofino-Costa R, Freitas DFS, Bernardes-Engemann AR, et al. Human sporotrichosis: recommendations from the Brazilian Society of Dermatology for the clinical, diagnostic and therapeutic management. An Bras Dermatol. 2022;97:757–777. In this work, Orofino-Costa and colleagues provide standardized recommendations for the therapeutic management, diagnosis and clinical aspect of sporotrichosis. These instructions were put together by a panel of twelve experts in human sporotrichosis from different Brazilian regions, divided by three different work groups: clinical, diagnosis and treatment.
Alves M do M, Milan EP, Silva-Rocha WP da, et al. Fatal pulmonary sporotrichosis caused by Sporothrix brasiliensis in Northeast Brazil. PLoS Negl Trop Dis. 2020;14:e0008141.
de Andrade Galliano Daros Bastos F, Raimundo Cognialli RC, Rodrigues de Farias M, Dos Santos Monti F, Wu K, Queiroz-Telles F. Spread of Sporothrix spp. through respiratory droplets from infected cats: a potential route of transmission. Med Mycol. 2022;60:myac079.
Farooqui SM, Youness H. The infection returns: a case of pulmonary sporotrichosis relapse after chemotherapy. Case Rep Med. 2018;2018:1384029.
Rojas FD, Fernández MS, Lucchelli JM, Lombardi D, Malet J, Vetrisano ME, Cattana ME, Sosa M de LÁ, Giusiano G. Cavitary pulmonary sporotrichosis: case report and literature review. Mycopathologia. 2017;182:1119–1123.
Yagnik KJ, Skelton WP, Olson A, Trillo CA, Lascano J. A rare case of disseminated Sporothrix schenckii with bone marrow involvement in a patient with idiopathic CD4 lymphocytopenia. IDCases. 2017;9:70–2.
Mahajan VK. Sporotrichosis: an overview and therapeutic options. Dermatol Res Pract. 2014;2014: 272376.
Casadevall A, Pirofski LA. Host-pathogen interactions: redefining the basic concepts of virulence and pathogenicity. Infect Immun. 1999;67:3703–13.
García Carnero LC, Lozoya Pérez NE, González Hernández SE, Martínez Álvarez JA. Immunity and treatment of sporotrichosis. J Fungi Basel Switz. 2018;4:100.
Rodriguez-Caban J, Gonzalez-Velazquez W, Perez-Sanchez L, Gonzalez-Mendez R, Rodriguez-del Valle N. Calcium/calmodulin kinase1 and its relation to thermotolerance and HSP90 in Sporothrix schenckii: an RNAi and yeast two-hybrid study. BMC Microbiol. 2011;11:162.
Arrillaga-Moncrieff I, Capilla J, Mayayo E, Marimon R, Mariné M, Gené J, Cano J, Guarro J. Different virulence levels of the species of Sporothrix in a murine model. Clin Microbiol Infect Off Publ Eur Soc Clin Microbiol Infect Dis. 2009;15:651–5.
Tamez-Castrellón AK, Romeo O, García-Carnero LC, Lozoya-Pérez NE, Mora-Montes HM. Virulence factors in Sporothrix schenckii, one of the causative agents of sporotrichosis. Curr Protein Pept Sci. 2020;21:295–312.
Lopes-Bezerra LM, Schubach A, Costa RO. Sporothrix schenckii and sporotrichosis. An Acad Bras Cienc. 2006;78:293–308.
Freitas DFS, de Siqueira Hoagland B, do Valle ACF, et al. Sporotrichosis in HIV-infected patients: report of 21 cases of endemic sporotrichosis in Rio de Janeiro, Brazil. Med Mycol. 2012;50:170–178.
Cruz ILR, Freitas DFS, de Macedo PM, et al. Evolution of virulence-related phenotypes of Sporothrix brasiliensis isolates from patients with chronic sporotrichosis and acquired immunodeficiency syndrome. Braz J Microbiol Publ Braz Soc Microbiol. 2021;52:5–18.
Silva-Bailão MG, Lima P de S, Oliveira MME, Oliveira LC, Almeida-Paes R, Borges CL, Bailão AM, Coelho ASG, Soares CM de A, Zancopé-Oliveira RM. Comparative proteomics in the three major human pathogenic species of the genus Sporothrix. Microbes Infect. 2021;23:104762.
Boechat JS, Oliveira MME, Almeida-Paes R, et al. Feline sporotrichosis: associations between clinical-epidemiological profiles and phenotypic-genotypic characteristics of the etiological agents in the Rio de Janeiro epizootic area. Mem Inst Oswaldo Cruz. 2018;113:185–96.
Carlos IZ, Batista-Duharte A. Sporotrichosis: an emergent disease. In: Zeppone Carlos I, editor. Sporotrichosis New Dev. Future Prospects. Springer International Publishing, Cham, 2015. pp 1–23.
Schubach TMP, Schubach A de O, Okamoto T, et al. Sporothrix schenckii isolation from blood clot of naturally infected cats. Braz J Vet Res Anim Sci. 2004;41:404–408.
Zancope-Oliveira RM, de Almeida-Paes R, de Oliveira MME, Freitas DFS, Galhardo MCG. New diagnostic applications in sporotrichosis. Skin Biopsy - Perspect. 2011. https://doi.org/10.5772/23590.
Lopes-Bezerra LM, Mora-Montes HM, Zhang Y, Nino-Vega G, Rodrigues AM, de Camargo ZP, de Hoog S. Sporotrichosis between 1898 and 2017: the evolution of knowledge on a changeable disease and on emerging etiological agents. Med Mycol. 2018;56:126–43.
Baptista VS, Mothé GB, Santos GMP, et al. Promising application of the SsCBF ELISA test to monitor the therapeutic response of feline sporotrichosis caused by Sporothrix brasiliensis from Brazilian epidemics. Braz J Microbiol Publ Braz Soc Microbiol. 2021;52:145–53.
Almeida-Paes R, Pimenta MA, Pizzini CV, Monteiro PCF, Peralta JM, Nosanchuk JD, Zancopé-Oliveira RM. Use of mycelial-phase Sporothrix schenckii exoantigens in an enzyme-linked immunosorbent assay for diagnosis of sporotrichosis by antibody detection. Clin Vaccine Immunol. 2007;14:244–9.
BIDiagnostics - Sorodiagnóstico de doenças infecto-parasitárias. In: BIDiagnostics. https://bidiagnostics.com.br/. Accessed 24 May 2023.
Almeida-Silva F, Almeida M de A, Rabello VB de S, et al. Evaluation of five non-culture-based methods for the diagnosis of meningeal sporotrichosis. J Fungi Basel Switz. 2023;9:535. Because meningeal sporotrichosis diagnosis from cerebrospinal fluid is usually hard due to the low fungal burden presented, Almeida-Silva and colleagues tested and discussed five non-culture-based methods for diagnosis. The authors discovered that four out of the five methods tested had substantial levels of sensitivity and specificity and suggest that these methods should be implemented as soon as possible into clinical practice to improve prognosis in these cases.
Thomson P, González C, Blank O, Ramírez V, Río CD, Santibáñez S, Pena P. Sporotrichosis outbreak due to Sporothrix brasiliensis in domestic cats in Magallanes, Chile: a One-Health-approach study. J Fungi Basel Switz. 2023;9:226.
Gremião IDF, da Silva M, da Rocha E, Montenegro H, et al. Guideline for the management of feline sporotrichosis caused by Sporothrix brasiliensis and literature revision. Braz J Microbiol Publ Braz Soc Microbiol. 2021;52:107–24.
de Lima Barros MB, Schubach AO, de Vasconcellos Carvalhaes de Oliveira R, Martins EB, Teixeira JL, Wanke B. Treatment of cutaneous sporotrichosis with itraconazole--study of 645 patients. Clin Infect Dis Off Publ Infect Dis Soc Am. 2011;52:e200–206.
Sivagnanam S, Bannan AM, Chen SC-A, Ralph AP. Sporotrichosis (Sporothrix schenckii infection) in the New South Wales mid-north coast, 2000–2010. Med J Aust. 2012;196:588–90.
Tang MM, Tang JJ, Gill P, Chang CC, Baba R. Cutaneous sporotrichosis: a six-year review of 19 cases in a tertiary referral center in Malaysia. Int J Dermatol. 2012;51:702–8.
Schubach TMP, Schubach A, Okamoto T, et al. Canine sporotrichosis in Rio de Janeiro, Brazil: clinical presentation, laboratory diagnosis and therapeutic response in 44 cases (1998–2003). Med Mycol. 2006;44:87–92.
de Souza EW, Borba C de M, Pereira SA, et al. Clinical features, fungal load, coinfections, histological skin changes, and itraconazole treatment response of cats with sporotrichosis caused by Sporothrix brasiliensis. Sci Rep. 2018;8:9074.
Orofino-Costa R, de Macedo PM, Rodrigues AM, Bernardes-Engemann AR. Sporotrichosis: an update on epidemiology, etiopathogenesis, laboratory and clinical therapeutics. An Bras Dermatol. 2017;92:606–20.
Rodrigues AM, Della Terra PP, Gremião ID, Pereira SA, Orofino-Costa R, de Camargo ZP. The threat of emerging and re-emerging pathogenic Sporothrix species. Mycopathologia. 2020;185:813–42.
Vettorato R, Heidrich D, Fraga F, Ribeiro AC, Pagani DM, Timotheo C, Amaro TG, Vettorato G, Scroferneker ML. Sporotrichosis by Sporothrix schenckii senso stricto with itraconazole resistance and terbinafine sensitivity observed in vitro and in vivo: case report. Med Mycol Case Rep. 2018;19:18–20.
Fischman Gompertz O, Rodrigues AM, Fernandes GF, Bentubo HDL, de Camargo ZP, Petri V. Atypical clinical presentation of sporotrichosis caused by Sporothrix globosa resistant to itraconazole. Am J Trop Med Hyg. 2016;94:1218–22.
Gremião I, Schubach T, Pereira S, Rodrigues A, Honse C, Barros M. Treatment of refractory feline sporotrichosis with a combination of intralesional amphotericin B and oral itraconazole. Aust Vet J. 2011;89:346–51.
da Rocha RFDB, Schubach TMP, Pereira SA, Dos Reis ÉG, Carvalho BW, Gremião IDF. Refractory feline sporotrichosis treated with itraconazole combined with potassium iodide. J Small Anim Pract. 2018;59:720–1.
Guterres KA, de Matos CB, Osório LDG, Schuch ID, Cleff MB. The use of (1–3) β-glucan along with itraconazole against canine refractory sporotrichosis. Mycopathologia. 2014;177:217–21.
Bernardes-Engemann AR, Tomki GF, Rabello VB de S, Almeida-Silva F, Freitas DFS, Gutierrez-Galhardo MC, Almeida-Paes R, Zancopé-Oliveira RM. Sporotrichosis caused by non-wild type Sporothrix brasiliensis strains. Front Cell Infect Microbiol. 2022;12:893501.
Morris-Jones R, Youngchim S, Gomez BL, Aisen P, Hay RJ, Nosanchuk JD, Casadevall A, Hamilton AJ. Synthesis of melanin-like pigments by Sporothrix schenckii in vitro and during mammalian infection. Infect Immun. 2003;71:4026–33.
Teixeira MM, Almeida-Paes R, Bernardes-Engemann AR, et al. Single nucleotide polymorphisms and chromosomal copy number variation may impact the Sporothrix brasiliensis antifungal susceptibility and sporotrichosis clinical outcomes. Fungal Genet Biol FG B. 2022;163: 103743.
Brilhante RS, Pereira VS, Oliveira JS, et al. Pentamidine inhibits the growth of Sporothrix schenckii complex and exhibits synergism with antifungal agents. Future Microbiol. 2018;13:1129–40.
Brilhante RS, Pereira VS, Oliveira JS, et al. Terpinen-4-ol inhibits the growth of Sporothrix schenckii complex and exhibits synergism with antifungal agents. Future Microbiol. 2019;14:1221–33.
Garcia Ferreira P, Pereira Borba-Santos L, Noronha LL, DeckmanNicoletti C, de Sá Haddad Queiroz M, de Carvalho da Silva F, Rozental S, Omena Futuro D, Francisco Ferreira V. Synthesis, stability studies, and antifungal evaluation of substituted α- and β-2,3-dihydrofuranaphthoquinones against Sporothrix brasiliensis and Sporothrix schenckii. Mol Basel Switz. 2019;24:930.
Waller SB, Cleff MB, de Mattos CB, et al. In vivo protection of the marjoram (Origanum majorana Linn.) essential oil in the cutaneous sporotrichosis by Sporothrix brasiliensis. Nat Prod Res. 2021;35:2977–81.
Almeida-Silva F, Bernardes-Engemann AR, Bérenger ALR, da Silva VP, Figueiredo MR, Freitas DFS. In vitro activity of Schinus terebinthifolius extract and fractions against Sporothrix brasiliensis. Mem Inst Oswaldo Cruz. 2022;117: e220063.
Artunduaga Bonilla JJ, Honorato L, Haranahalli K, et al. Antifungal activity of acylhydrazone derivatives against Sporothrix spp. Antimicrob Agents Chemother. 2021;65:e02593–20, AAC.02593–20.
Artunduaga Bonilla JJ, Honorato L, Guimarães AJ, Miranda K, Nimrichter L. Silver chitosan nanocomposites are effective to combat sporotrichosis. Front Nanotechnol. 2022;4.
Liu S, Zamith-Miranda D, Almeida-Paes R, da Silva LBR, Nacharaju P, Nosanchuk JD. Nitric oxide-loaded nano- and microparticle platforms serving as potential new antifungal therapeutics. Fungal Biol. 2023;127(7–8):1224–30. https://doi.org/10.1016/j.funbio.2023.01.007.
Borba-Santos LP, Barreto TL, Vila T, et al. In vitro and in vivo antifungal activity of buparvaquone against Sporothrix brasiliensis. Antimicrob Agents Chemother. 2021;65: e0069921.
Borba-Santos LP, Nucci M, Ferreira-Pereira A, Rozental S. Anti-Sporothrix activity of ibuprofen combined with antifungal. Braz J Microbiol Publ Braz Soc Microbiol. 2021;52:101–6.
Asquith CRM, Machado ACS, de Miranda LHM, Konstantinova LS, Almeida-Paes R, Rakitin OA, Pereira SA. Synthesis and identification of pentathiepin-based inhibitors of Sporothrix brasiliensis. Antibiotics. 2019;8:249.
Mathias LDS, Almeida JCDA, Passoni LC, Gossani CMD, Taveira GB, Gomes VM, Vieira-Da-Motta O. Antifungal activity of silver salts of Keggin-type heteropolyacids against Sporothrix spp. J Microbiol Biotechnol. 2020;30:540–51.
Borba-Santos LP, Vila T, Rozental S. Identification of two potential inhibitors of Sporothrix brasiliensis and Sporothrix schenckii in the Pathogen Box collection. PLoS ONE. 2020;15: e0240658.
Borba-Santos LP, Nicoletti CD, Vila T, et al. A novel naphthoquinone derivative shows selective antifungal activity against Sporothrix yeasts and biofilms. Braz J Microbiol Publ Braz Soc Microbiol. 2022;53:749–58.
Cabañes FJ. Sporotrichosis in Brazil: animals+humans=one health. Rev Iberoam Micol. 2020;37:73–4.
Wilfert L, Brown MJF, Doublet V. OneHealth implications of infectious diseases of wild and managed bees. J Invertebr Pathol. 2021;186: 107506.
Funding
AFV and DCJ were supported by grants from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES, Finance Code 001); AFV was also supported by NIH AI165204. LN, JJAB, and SF were supported by grants from Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ). LN and SF were also supported by grants from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq). JDN and DZM were supported in part by NIH R21 AI124797, NIH AI165204, and DOD OR200187.
Ethics declarations
Ethical Approval
Not applicable.
Conflict of Interest
The authors declare no competing interests.
Human and Animal Rights and Informed Consent
This article does not contain any studies with human or animal subjects performed by any of the authors.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is part of the Topical Collection on Tropical Mycoses
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Valdez, A.F., Corrêa-Junior, D., Bonilla, J.J.A. et al. A Review on Sporotrichosis and the Emergence of Sporothrix brasiliensis as a Pathogen. Curr Trop Med Rep 10, 252–261 (2023). https://doi.org/10.1007/s40475-023-00297-6
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40475-023-00297-6