
Abstract
The purpose of the current study was to evaluate adaptability and phenotypic stability of 24 exotic advanced sugarcane genotypes across three major sugarcane producing agro-ecologies of Ethiopia and identify ideal location for genotype evaluation via GGE biplot analyses. The trials were conducted for 3 years involving three crop cycles or three cuttings corresponding to plant cane and first and second ratoon crops representing nine environments (location-cut combinations). Data for estimable recoverable sugar percent (ERS), cane yield (CY) (tons/ha), and sugar yield (SY) (tons/ha) were collected from the trials and subjected to ANOVA and GGE [genotype (G) plus genotype-environment (GE)] biplot analysis. ANOVA portrayed that genotype by environment interaction (GEI) accounted for 42.74%, 54.70% and 60.52% of the total variation for ERS, CY and SY, respectively, indicating GEI variation was a substantial proportion of the total variation in all the traits. High broad sense heritability (H2) was recorded for ERS (79.28) whereas that of SY was extremely low (2.90) showing difficulty of this trait to improve by simple phenotypic selection. GGE biplots for ERS, CY and SY showed that the first two principal components (PCs) together explained 59.01%, 80.54% and 71.73% of the total variation, respectively. GGE biplot analysis also revealed high positive correlations among crop cycles at all locations for all measured traits implying evaluation of genotypes across locations could give reliable information on the performance of the genotypes than testing for ratooning ability. The location Metehara, being both discriminating and representative, was identified as ideal for genotype evaluation. Genotypes G1 (PSR-9784) and G7 (VMC95-252) having high mean performance for cane yield (152 tons/ha and 142 tons/ha) and sugar yield (18 tons/ha and 18 tons/ha) and stability across environments, were recommended as ideal genotypes. These genotypes could be commercially grown at the studied locations or similar agro-ecologies.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Sugarcane (Saccharum spp.) is an important cash crop and is cultivated in the tropical and subtropical regions of the world. As is the case in many tropical countries sugarcane is the main source of sugar production in Ethiopia. It is an important crop widely cultivated for multiple purposes by smallholder farmers in sub-Saharan Africa and also by commercial farms. In Ethiopia sugarcane has been cultivated by smallholder farmers in the backyards and farmers’ fields since ancient times [8]. It was introduced to the country during the Axumite Kingdom in the first to fourth century period [20, 23].
In Ethiopia, sugarcane commercial cultivation commenced in 1951 and is currently expanded to every corner of the country in various agro-ecologies and geographic regions. It was started in the three old sugar estates starting in 1951 at Wonji, in 1969 at Metehara and in 1998 at Fincha. These three sugarcane growing agro-ecologies represent varied rainfall, soil, and other environmental conditions and has been used to test sugarcane genotypes since their inception. However, no systematic effort has been made so far to study genotype by environment interaction (GEI) in sugarcane and evaluate the test environments for genotype selection.
The sustainability of cane production in Ethiopia depends on productivity gains that could be achieved in each agro-climatic region of the industry with the release of new and high-yielding cultivars. This necessitates broadly and specifically adapted cultivars with desired traits. Although breeders consider many traits, such as disease resistance, harvesting characteristics, and ratooning ability, their top priority is to select for high sugar yield through its component traits of cane tonnage and sugar content.
Ethiopia has been relying on the importation of sugarcane varieties for commercial production. Many collections of clonal materials in different stages of breeding and selection programs have been introduced and evaluated for adaptation and yield performance. In this line, during 2012 advanced breeding clones have been imported from French Agricultural Research Centre for International Development (CIRAD), France. These new exotic clones needed to be evaluated across target agro-ecologies to select high-performing candidate varieties.
The development of sugarcane varieties that suit different agro-ecological and growing conditions involve genotypic evaluation across locations and crop cycles. In this process, the phenomena of GEI often complicate the identification of superior genotypes by confounding the effects of the environment and making it difficult to know the true genetic values of genotypes. GEI may cause changes in the relative ranking of genotypes across sites and/or crop years in trials and complicate identification of superior cultivars by confounding determination of true genetic values.
When GEI exist, its statistical significance and precise characteristics must be investigated in detail to asses in detail the relationships between environments and the pattern of the response of genotypes across environments. By the detail study of GIE it is also necessary to assess implications for selection strategies and help optimize resource allocation across locations and years. Furthermore, we should investigate the similarity of environments and their ability to discriminate genotypes, identify redundant sites or mega environments and analyze the stability of the site response across genotypes, visualize the performance rank and stability of genotypes across environments for the purposes of decision-making regarding release of new cultivars.
Genotype plus genotype x environment interaction (GGE) biplot analysis is a very useful graphical tool to investigate GIE and visualize the performance rank and stability of genotypes across environments [35]. GGE biplot can facilitate a better understanding of complex GEI in multi-environment trials of breeding lines and agronomic experiments. GGE biplot has been used to identify the performance of crop cultivars under multiple environments, ideal cultivars, mega-environment, and core testing sites [1]. It also has been successfully used in crop trials to analyze multi environmental trials data to release new varieties, including wheat [6], cotton [31], sunflower [28], and sugarcane [13, 22, 27].
With these views in mind, the present study was conducted to study the adaptability and phenotypic stability of exotic sugarcane genotypes in major sugarcane-producing agro-ecologies of Ethiopia and identify ideal location for genotype evaluation via GGE biplot analyses.
Materials and Methods
Study Sites
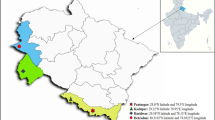
The experiments were conducted at three main Sugar Estates of Ethiopia during 2017/18 to 2019/20 (Fig. 1).

Location map of the experimental sites representing three major sugarcane producing agro-ecologies in Ethiopia
Wonji
Wonji Sugar Factory is located in Oromia Regional Government State, Eastern Shewa Zone, Adama Woreda, About 110 km from Addis Ababa and about 10 km south of Adama Town with latitude 8° 31′ N and longitude 39° 12′ E with elevation of 1550 masl. The average annual rainfall is 800 mm with maximum and minimum temperatures 26.9 °C and 15.3 °C respectively. The growth period for plant cane is 18–24 months while 13–15 months for ratoon crop. The plantation is covered by 95% of clay soils and vertisols and irrigation water is applied as furrow irrigation [8].
Metehara
Metehara sugar factory is located in Oromia Regional Government State, Eastern Shewa Zone about 200 km from Addis Ababa and about 8 km south of Metehara Town with latitude and longitude 8° 51′ N and 39° 52′ E respectively and with elevation of 950 masl. Annual rainfall is 554 mm with temperature maximum and minimum of 32.6 °C and 17.5 °C, respectively. The soils of the area are mainly vertisols. Furrow irrigation is practiced in the area. Harvesting age of sugarcane is 18–20 months for plant cane and 13–15 months for ratoon cane fields [8].
Fincha
Finchaa Sugar Estate is located in the western part of Oromia Reginal state at 9° 30′ to 10° 00′ N and 37° 15′ to 37° 30′ E. It is about 330 km North West of Addis Ababa with an elevation between 1350 and 1650 m.a.s.l. The average annual rainfall is 1280 mm with maximum and minimum temperatures 30.7 °C and 14.8 °C, respectively. The average length of the growing period of sugarcane in the area is 20–22 months for plant cane while 14–15 months for ratoon cane fields. The Estate practices dominantly sprinkler irrigation which does not critically require land forming operation. More than 95 percent of the cultivated and irrigated land soils in Finchaa are grouped in to Luvisols and Vertisols [8].
Plant Materials and Experimental Design
The experimental materials in this study involved 24 sugarcane genotypes. Twenty two of the materials were advanced breeding clones introduced from CIRAD, France and the remaining two were standard check varieties commercially cultivated in the areas (Supplemental Table 1). The experimental period lasts 3 years (2017/18, 2018/19 and 2019/20) involving three crop cycles or three cuttings corresponding to plant cane and first and second ratoon crops representing nine environments (location-cut combinations).
The experimental design used was Randomized Complete Block Design (RCBD) in three replications. The experimental plots consisted of six rows of 5 m in length and 1.45 m between rows with a plot area of 43.5 m2. The two rows on each side of the plot served as guard rows and the middle 4 rows were net experimental plots from which data was collected. Irrigation water was applied as furrow surface irrigation at Wonji and Metehara and using a sprinkler at Fincha. Other cultural practices were applied as recommended for the areas.
Data Collected
During harvest 50 stalk samples were taken randomly from the middle 4 furrows and weighed and used to estimate single cane weight. The number of the millable stalk was counted from the whole net plot of 4 rows and used to calculate cane yield per plot by multiplying single cane weight by stalk number per plot and converted to tons of cane yield per hectare (CY) confirming to the area of the plot. From the 50 stalk samples, 15 stalks were used to determine estimable recoverable sugar percent (ERS) using the standard hydraulic press method [15]. Sugar yield ton per hectare (SY) was calculated by multiplying cane yield ton per hectare by ERS.
Data Analyses
Using the data for CY, ERS and SY, first analysis of variance was performed for each location or production environment and for each crop cycle that is plant cane, first ratoon and second ratoon.
Data were combined over locations after checking the homogeneity of error variance as in [14]. The combined analysis of variance resulted in a highly significant (p < 0.01) genotype-by-environment interaction (GEI) which paves the way to study the GEI and adaptability and phenotypic stability of the genotypes using GGE biplot analysis (Table 1).
The GGE biplot analysis was performed as in [34] using the model:
where \({\overline{Y} }_{ij}\) is the performance of genotype i in environment j, µ is the grand mean, βj is the main effect of environment j, k is the number of principal components (PC); \({\lambda }_{k}\) is the singular value of the kth PC; and \({\mathrm{\alpha }}_{ik}\) and \({\gamma }_{jk}\) are the scores of ith genotype and jth environment, respectively for PCk; \({\in }_{ij}\) is the residual associated with genotype i in environment j. Analysis was done using the software GenStat 18th edition [30].
To estimate the genotypic and phenotypic variance components, the data was subjected to the VARCOMP procedure of SAS statistical software V9 [28]. These components were used to estimate broad-sense heritability on genotype mean basis considering three replicates and three locations [10, 16].
where σ2g = genotype, σ2gl = genotype × location σ2e = error variances, r = number of replications and l = number of locations.
Results and Discussion
Analysis of Variance (ANOVA)
Combined analysis of variance for estimable recoverable sugar (ERS), cane yield (CY) and sugar yield (SY) showed highly significant (p < 0.01) effects of the environment, genotypes and GEI (Table 1). As can be observed from Table 1, the percentage of the total sum of squares for genotype in ERS and CY was higher than the environment. On the other hand, the total sum of squares accounted for the environment was greater than for genotypes in SY. This showed that ERS and CY were less affected by specific environments of locations i.e. cropping years or cropping cycles i.e. plant cane and ratoon crops than SY. [22, 26] found in multi-environment trials that SY was more affected by environment than genotype. Unlike the present study, many reports showed that the effect of environment was more pronounced than the genotype effect in ERS and CY [13, 22, 26]. This suggests that in the future, trials involving more environments may be necessary to see the effect of the environment on these traits. The ANOVA portrayed that GEI accounted for 42.74%, 54.70% and 60.52% of the total variation for ERS, CY and SY, respectively. Thus, GEI variation was a substantial proportion of the total variation in all the traits studied thereby indicating the inconsistent performance of genotypes across environments.
The significant effect for GEI showed that the study was carried out under variable agro-climatic conditions which paves the way to study the specific adaptability and phenotypic stability of genotypes. When GEI is significant, we need to accurately sample the target environmental conditions where the varieties will be grown after release, using the trials planted at several locations. Because sugarcane is a perennial crop, these environments are made up of locations and crop stages (plant, first, and subsequent ratoon crops).
Sugarcane is usually characterized as a crop with low fertility, high GEI and high heterozygosity. When GEI is significant and high it leads to differential responses of genotypes across growing locations and environments and may limit selection response [13]. Variation among genotypes in phenotypic sensitivity to the environment (GEI) may necessitate the development of locally adapted varieties [9].
Genotypes showed a larger contribution to GEI than the E component of the variance for estimable recoverable sugar (ERS) and cane yield (CY) determined through analysis of variance, indicating that their performance was differentially modified by environmental factors. The inconsistent performance of some genotypes across years (crop cycles) in the same location emphasized that G and GEI must be considered together to make meaningful selection decisions rather than G alone [17], especially when crossover GE interactions are abundant [18].
The mean ERS of each test genotype and the checks in each environment is presented in Supplemental Table 2. The genotypes G4 (CPOO-1527) and G24 (FGO4-798) showed superior performance in terms of ERS. G2 (CPOO-1301) and G15 (FGO4-896) also had arguably the good performance of this trait. These genotypes could be used as potential parents in future crossing programs to improve sugar content. The genotype G13 (VNC-9655) scored the highest cane yield (Supplemental Table 3) whereas the genotypes with the highest sugar yield were G1 (PSR-9784), G7 (VMC95-252) and one of the check variety G9 (NCO-334) (Supplemental Table 4). These genotypes scored the highest sugar yield than G13 because of relatively higher performance in their ERS as sugar yield is the product of cane yield and ERS.
The Which-Won-Where Pattern
The GGE biplot shows the which-won-where pattern in a genotype-by-environment dataset and its graphical presentation addresses important concepts [35]. The symmetric scaling of genotype and environment scores in GGE biplots allows direct visualization of the magnitude of genotypic and environmental variations in the same units for both PC1 and PC2 [33]. Figure 2a presents GGE biplots for ERS based on symmetric scaling where the first two principal components (PCs) together explained 59.01% of the total variation with PC1 and PC2 explaining 43.98% and 15.03% of the variation, respectively. The two components PC1 and PC2 approximated less efficiently the GGE data of ERS. For ERS, data variability can be represented sufficiently when taking into account the additional contribution of its PC3 (12.96%). In contrast to the present finding, Guilly et al. [13] reported a GGE biplot represented by both PC1 and PC2 adequately approximating (71.55%) of the GGE data for ERS. The variation with the current finding could be they have used more genotypes and environments in their study.

By connecting vertex genotypes that are further away from the biplot origin, a polygon was formed. A line drawn originating from the biplot origin and intersecting the sides of the polygon at right angle divided the biplot into different sectors (Fig. 2a). The vector genotypes had the largest vector in their respective direction which showed the extent of the response of the genotypes to the tested environments. Genotypes contained within the polygon in a respective sector show less response or interaction to the environments contained in that sector. Accordingly, three mega environments were identified contained in three sectors. One of the mega environments contained only one environment E3 (Wonji during 2019/20). Genotypes G2 and G24 were the highest performers in this environment. The second mega environment contained environments E1 (Wonji in 2017/18), E4 (Metehara in 2017/18), E7 (Fincha in 2017/18) and E8 (Fincha in 2018/19). G24 being closest to this mega environment was the best performer in this group of environments (Fig. 2a and Supplemental Table 3). Genotype G4 had the longest vector in the sector containing the third mega environment with environments E2 (Wonji in 2018/19), E5 (Metehara in 2018/19), E6 (Metehara in 2019/20), and E9 (Fincha in 2019/20) and was favorable for these group of environments (Fig. 2a and Supplemental Table 3).
The which-won-where feature of the GGE biplot revealed the polygon view of cane yield where the first two principal components (PCs) represented 80.54% of the total variation with the first and second PCs explaining 53.66% and 26.88%, respectively (Fig. 2b). This was congruent with previous reports [25, 29]. This GGE biplot revealed four mega environments where the check variety NCO-334 (G9) and G22 were the winners in environments E4, E5 and E6 (Metehara); G13 in E1, E2, E7 and E8 (Wonji and Fincha); G1 in E2, E3, E7 and E8 (Wonji and Fincha) and G14 in E8 and E9 (Fincha).
For the sugar yield GGE biplot (Fig. 2c) the first two PCs together captured 71.73% of the total observed variation quite sufficient to explain the variation and allow us to draw concrete conclusions. Nine environments fell into four sectors. G22, G17 and one of the checks G9 were the winner genotypes in environments E4 and E5 (Metehara); G14 in E8 and E9 (Fincha); G1 in E1, E2 and E3 (Wonji) and E7, E8 and E9 (Fincha) revealing their specific adaptation to these locations.
The mean performance of genotypes for the three traits across environments (Supplemental Tables 2, 3 and 4) was similar to the results for winning genotypes based on the ‘which-won-where pattern’ of respective biplots (Fig. 2).
For cane and sugar yield biplots (Fig. 2b, c), the interesting feature of these view of the GGE biplots is that the genotypes on the vertex for each sector had the highest cane yield and sugar yield values in all environments that fall in the sector, providing that the percentage of GGE data explained by the biplot is high enough to accurately reflect the original data. The reliability of the which-won-where pattern revealed in these biplots by vertex genotypes depends on (i) the cumulated weights of PC1 and PC2 axes, (80.54% for cane yield) and (71.73% for sugar yield), and (ii) the narrowness of the angles that encompass all the vectors of environments falling in each sector of interest. The angles between environments in respective sectors for cane and sugar yield were acute which can also be evidenced from Fig. 4b, c.
Mean Performance and Stability of Genotypes
Figure 3 showed the rank performance of the genotypes and stability for the three traits. The average environment coordination (AEC) abscissa, has one direction, with the arrow pointing to the greater genotype main effect. In the AEC ordinate moving either direction away from the biplot origin indicates a greater GEI effect and reduced stability. The genotypes were ranked according to their projections on the AEC ordinate. Yan and Hunt [32] studied the performance and stability of genotypes for yield via the AEC method. The AEC ordinate separates genotypes with below-average means from those with above-average means. Hence, for ERS (Fig. 3a), G4 being close to the tip of the arrowhead on the AEC abscissa was the best genotype followed by G24, G15 and G2. Both the check genotypes G9 and G10 had below-average performance, though, nearest to the average value (Fig. 3a and Supplemental Table 2). G13 had the lowest mean ERS. Moreover, in terms of stability, the genotype G15 with relatively short projection on the AEC ordinate and with consistent performance across environments was the most stable. Genotype G2 was the most unstable because it had the largest projection. It had above-average performance in most of the environments and one of the lowest performance in environment E6 and below-average performance in E9 (Fig. 3a and Supplemental Table 2).

Similarly, the AEC showed that G13 followed by G9 (one of the checks), G1 and G7 were the genotypes with the highest cane yield performance (Fig. 3b). Likewise, the highest sugar yield performance was observed for genotypes G9 and G1 followed by G7 and G4 (Fig. 3c). Genotypes G24 and G15 had the lowest cane yield and sugar yield, respectively. A similar pattern of rank performance of genotypes for cane yield and sugar yield in this regard was expected as sugar yield is the product of cane yield and estimable recoverable sugar percent (ERS). Sandhu et al. [27] also observed similar results where averaged over environments and relative to average sugar recovery, genotypes with higher cane yield also gave high sugar yield. G3 and G5 were the most stable for cane yield performance whereas G7 and G13 had stable sugar yield performance. Among the two checks G9 although scored the highest cane and sugar yield was unstable.
Similarly, in the other check G10 was also unstable with an above-average mean of cane and sugar yield.
In sugarcane, the final aim is to get high sugar yield which is the function of both cane yield and quality trait estimable recoverable sugar percent and with best environmental stability. Therefore, the ideal genotype should have high sugar yield performance and high genotypic stability. In the current study, the genotypes (G7) and (G1) were the ideal genotypes for the studied environments (Fig. 3c). These genotypes had also sugar yields comparable with the highest performing check variety G9 (Supplemental Table 4). Such genotypes possessing wide adaptability and high mean performance of cane yield and sugar yield can be commercially utilized for all studied locations and similar agro-ecologies.
Generally, based on the GGE ranking biplots (Fig. 3 and Supplemental Tables 2, 3 and 4) it was noted that most of (90%) the French-Guyana (FG) genotypes (G15–G24) except G19 had above-average performance in ERS while most had below average performance in CY and SY. These genotypes could be utilized in breeding programs to improve ERS. The other genotypes (G1–14) which also included the two check varieties NCO-334 and B52-298 performed better in CY and SY in the present studied environments. In contrast to the present study, an investigation conducted in similar environments involving FG genotypes reported that these genotypes had more cane yield performance whereas their estimable recoverable sugar percent was relatively low [11].
These materials had been selected for cane yield in the source country where imported. This might also give a hint on the difficulty to improve both cane yield and sugar quality traits simultaneously. The report by [4, 19] showed the negative correlation of pol percent and recoverable sugar percent with cane yield is one of the major constraints in the improvement of sugarcane. In a given population, the adaptability of genotypes follows the environmental variation and the particular selection pressure to which they are exposed [3]. Therefore the observations of this study gives a clue as to how our import strategy of sugarcane genotypes in the future should be geared on, importing breeding lines selected specifically for the traits of interest.
Evaluation of Environments
Relationship among Test Environments
The relationship among test environments based on the mean performance of the genotypes for the three traits is depicted in the GGE biplots in Fig. 4. As described by [33] in biplot analysis an acute angle between two environmental vectors indicates a positive correlation, an obtuse angle a negative correlation, and a right angle deduces no correlation. Moreover, a small acute angle between the vectors of two environments strongly suggests that the two environments, being similar, would discriminate among genotypes similarly [21].

GGE Biplot for environment testers used to test the mean performance of sugarcane genotypes for the three traits: estimable recoverable sugar percent (a), cane yield (b) and sugar yield (c)
The vector view of the GGE biplot for ERS, CY and SY in Fig. 4a–c showed that the vectors representing the three crop seasons plant cane, first ratoon and second ratoon (environments E1, E2 and E3 at Wonji; E4, E5 and E6 at Metehara and E7, E8 and E9 at Fincha) at all three locations showed similar positive correlations amongst them, as the angle among the environmental vectors were acute angles.
The insinuations of the results of this study regarding the relationship of test environments for the three economic traits showed that evaluation of genotypes in either one of the cropping cycles or seasons should suffice to imply that testing of multi-location trials involving only plant cane crops (one season or one crop cycle) could give us a clear picture of the performance of the genotypes regarding the traits considered. A similar study in sugarcane [13, 25] showed the same trend for ERS, cane and sugar yield and recommended testing only for one crop season in multi-location trials giving sound results for the traits evaluation. Therefore, evaluation of genotypes across locations could give reliable information on the performance of the genotypes than testing for ratooning ability.
Discriminating Ability and Representativeness of Environments
For ERS, among the nine environments, E6 (Metehara in 2019/20, second ratoon crop) was the most discriminating (informative) as depicted by the longest vector, whereas E2 (Wonji in 2018/19, first ratoon crop) was the least discriminating, having the shortest vector (Fig. 4a). Environment E6 was also the most representative whereas E4, E7 and E8 were the least representatives. For cane yield and sugar yield, Metehara in all three cropping seasons or crop cycles (E4, E5 and E6) was the location with the most discriminating ability of the genotypes, whereas Fincha at plant cane crop (E7) was the least discriminating environment (Fig. 4b, c).
Test environments, which were consistently non-discriminating (non-informative), provided little information on the genotypes and, therefore, can be omitted as test locations. In the current study, for cane and sugar yield the three crop seasons/crop cycles (plant cane and first and second ratoon crop) at Metehara (E4, E5 and E6) were found to be most discriminating and correlated where the correlation between plant cane and first ratoon crop was positive and strong. Therefore, testing of the sugarcane genotypes at plant cane only at Metehara should be informative, and testing at Wonji and Fincha can be dropped as environments as these locations were found to be the least discriminating. Similarly, Metehara (E4, E5 and E6) is the most representative location for both cane and sugar yield as the environmental vectors of E4, E5 and E6 had smaller angle with average environment axis (AEC) (Figs. 3b, c, 4b, c). Blanche and Myers [5] used the GGE biplot method to identify test locations that optimized genotype selection based on their discriminating ability and representativeness. Rea et al. [25] also used this methodology to identify environments which are discriminating and representative of cane yield and sugar quality traits. According to [13] cane yield and ERS traits revealed significant GEI, always higher than the genotype x crop year (GC) interaction, indicating that testing genotypes across locations is more important than testing for ratooning ability.
As described in [33], the ideal test environment should be the most discriminating (informative) and also most representative of all test environments. Therefore, Metehara being the most discriminating and representative location for both cane and sugar yield, it is the ideal environment for genotype evaluation. Thus, testing of sugarcane genotypes at Metehara would be appropriate for selecting high-yielding genotypes having wide adaptability. This would reduce cost and saves time.
Genotypic and Phenotypic Variance Components and Broad Sense Heritability
The genotypic and phenotypic variance components and broad sense heritability of the traits are summarized in Table 2. The genetic variance was somehow lower than phenotypic variance indicating the role played by the environment in the variability of the genotypes for these traits. Similar to the findings of this study, Alam et al. [2], Chaudhary [7] found genotypic variance lower than the phenotypic variance for the traits studied. With regard to heritability, relatively high broad sense heritability was recorded for ERS and moderate value was observed for CY and extremely less strong for SY.
High heritability values in traits suggest good repeatability for the traits across environments and is a reliable way to practice simple selection based on phenotypic performance (Gravois and Milligan [12]). The very low heritability for SY indicated low repeatability of this trait and the difficulty for the improvement of this trait by simple phenotypic selection. Chaudhary and Pandey et al. [7, 24] found similar result for sugar yield.
Conclusions
The results indicated that the performance of sugarcane genotypes for the traits estimable recoverable sugar, cane yield and sugar yield was highly influenced by genotype by environment interaction (GEI) effects. The GEI variation was a substantial proportion of the total variation in all the three traits studied. The very low heritability for sugar yield indicated low repeatability of this trait and the difficulty for the improvement of this trait by simple phenotypic selection. The GGE biplot analysis indicated that testing genotypes across locations gives more reliable information than testing for ratoonng ability. Metehara being the most discriminating and representative location for both cane and sugar yield is preferred as the ideal environment for genotype evaluation. In the future, more number of multi environment trials involving the current studied locations should be performed to validate the ideal environment. With their higher performance in terms of cane and sugar yield and stable across the test locations, genotypes G1 (PSR-9784) and G7 (VMC95-252) were found to be the ideal genotypes. The GGE biplot also identified specifically adapted genotypes for cane and sugar yield; accordingly, G22 (FGO6-750) and G14 (VMC-9647) found to be specifically adapted at Metehara and Fincha, respectively. These genotypes could be commercially grown at the studied locations or similar agro-ecologies.
References
Akinwale RO, Fakorede MAB, Badu-Apraku B, Oluwaranti A (2014) Assessing the usefulness of GGE biplot as a statistical tool for plant breeders and agronomists. Cereal Res Commun 42:534–546. https://doi.org/10.1556/crc.42.2014.3.16
Alam MN, Nath UK, Karim KMR, Ahmed MM, Mitul RY (2017) Genetic variability of exotic sugarcane genotypes. Scientifica. https://doi.org/10.1155/2017/5202913
Allard RW (1999) Principles of plant breeding, 2nd edn. Wiley, New York
Barreto FZ, Balsalobre TW, Chapola RG et al (2021) Genetic variability, correlation among agronomic traits, and genetic progress in a sugarcane diversity panel. Agriculture 11:533. https://doi.org/10.3390/agriculture11060533
Blanche SB, Myers GO (2006) Identifying discriminating locations for cultivar selection in Louisiana. Crop Sci 46:946–949. https://doi.org/10.2135/cropsci2005.0279
Chang GL, Chai S-X (2010) Application of GGE biplot in spring wheat yield stability analysis in rainfed areas of China. Chin J Eco-Agric 18:988–994. https://doi.org/10.3724/sp.j.1011.2010.00988
Chaudhary RR (2001) Genetic variability and heritability in sugarcane. Nepal Agric Res J 4:56–59
Esayas T (2014) Exploration and collection, characterization, genetic diversity analysis and association of traits for yield and yield components of sugarcane (Saccharum spp.) in Ethiopia. Dissertation, Haramaya University, Ethiopia
Falconer DS (1952) Selection for large and small size in mice. J Genet 51:470–501
Falconer DS, Mackay TFC (1996) Introduction to quantitative genetics, 4th edn. Addison Wesley Longman, Harlow
Ftwi M, Mekibib F, Tesfa M (2017) Maturity classification of sugarcane (Saccharum officinarum L) genotypes grown under different production environments of Ethiopia. Adv Crop Sci Technol. https://doi.org/10.4172/2329-8863.1000304
Gravois KA, Milligan SB (1992) Genetic Relationships between Fibre and Sugarcane Yield Components. Crop Sci 32:62-67. https://doi.org/10.2135/cropsci1992.0011183X003200010014x
Guilly S, Dumont T, Thong-Chane A et al (2017) Analysis of multienvironment trials (met) in the sugarcane breeding program of Réunion Island. Euphytica. https://doi.org/10.1007/s10681-017-1994-1
Hartley HO (1950) The maximum F-ratio as a short-cut test for heterogeneity of variance. Biometrika 37:308. https://doi.org/10.2307/2332383
Hoarau M (1969) Sugar cane analysis by hydraulic press method. Int Sugar J 71:328–332
Holland JB, Nyquist WE, Cervantes-Martinez CT (2003) Estimating and interpreting heritability for plant breeding: an update. Plant Breed Rev 22:11–112
Kang MS (1993) Simultaneous selection for yield and stability in crop performance trials: consequences for growers. Agron J 85:754–757. https://doi.org/10.2134/agronj1993.00021962008500030042x
Kerby T, Burgess J, Bates M, Albers D, Lege K (2000) Partitioning variety and environment contribution to variation in yield, plant growth, and fiber quality. In: Proceedings of the belt wide cotton conference, New Orleans. National Cotton Council of America, Memphis, pp 528–532
Khan IA, Seema N, Raza S, Yasmine S (2015) Comparative performance of sugarcane somaclones and exotic germplasm under agro-climatic conditions of Tando Jam. Pak J Bot 47(3):1161–1166
Kobishanov YM (1981) Aksum: political system, economics and culture, first to fourth century. In: Mokhtar G (ed.) General history of Africa II. Ancient civilizations of Africa. UNESCO International Scientific Committee for the Drafting of a General History of Africa. University of California Press.
Kroonenberg PM (1995) Introduction to biplots for G × E tables. Department of Mathematics. Research Report 51. Queensland, Australia: University of Queensland, Australia
Luo J, Pan Y-B, Que Y et al (2015) Biplot evaluation of test environments and identification of mega-environment for sugarcane cultivars in China. Sci Rep. https://doi.org/10.1038/srep15505
Munro-Hay S (1991) Aksumite overseas interests. Northeast Afr Stud 13(2/3):127–140
Pandey D, Singh SP, Jeena AS, Khan KA, Sum T, Negi A, Koujalagi D (2018) Study of genetic variability, heritability and genetic advance for various yield and quality traits in sugarcane genotypes (Saccharum Officinarum). Int J Curr Microbiol App Sci 7(4):1464–1472. https://doi.org/10.20546/ijcmas.2018.704.165
Rea R, De Sousa-Vieira O, Díaz A et al (2015) Genotype–environment interaction, megaenvironments and two-table coupling methods for sugarcane yield studies in Venezuela. Sugar Tech 18:354–364. https://doi.org/10.1007/s12355-015-0407-9
Regis JA, Andrade JA, Santos A et al (2018) Adaptability and phenotypic stability of sugarcane clones. Pesq Agrop Bras 53:42–52. https://doi.org/10.1590/s0100-204x2018000100005
Sandhu SK, Brar SS, Singh RS et al (2014) GGE biplot analysis for Cane and sugar yield from advanced-stage sugarcane trials in Subtropical India. J Crop Improv 28:641–659. https://doi.org/10.1080/15427528.2014.925025
Statistical Analysis System (SAS) Institute (2004) Version 9. SAS Institute Inc., Cary
Sun M, Jiang WM, Li HY, Liu Z (2010) Analysis of stable-production character by GGE biplot on sunflower hybrids. Heilongjiang Agri Sci 9:11–13
Surinder KS, Sawanpreet S, Brar RS, Singh PS, Bhagat I, Kang MS (2014) GGE biplot analysis for cane and sugar yield from advanced-stage sugarcane trials in subtropical India. J Crop Improv 28(5):641–659. https://doi.org/10.1080/15427528.2014.925025
VSN International (2015) Genstat for Windows 18th Edition. VSN International, Hemel Hempstead. Web page: Genstat.co.uk
Yan W, Holland JB (2009) A heritability-adjusted GGE biplot for test environment evaluation. Euphytica 171:355–369. https://doi.org/10.1007/s10681-009-0030-5
Yan W, Hunt LA (2001) Interpretation of genotype × environment interaction for winter wheat yield in Ontario. Crop Sci 41:19–25. https://doi.org/10.2135/cropsci2001.41119x
Yan W, Kang MS (2003) GGE biplot analysis: a graphical tool for breeders, geneticists, and agronomists. CRC Press, Boca Raton
Yan W, Kang MS, Ma B et al (2007) GGE biplot vs. ammi analysis of genotype-by-environment data. Crop Sci 47:643–653. https://doi.org/10.2135/cropsci2006.06.0374
Acknowledgements
The authors are grateful to the overall support of the Sugar Industry Group of Ethiopia.
Funding
No funding was received for conducting this study.
Author information
Authors and Affiliations
Contributions
Esayas Tena: Conceived and designed the experiments; Performed the experiments; Analyzed and interpreted the data; Wrote the paper. Feyissa Tadesse: Conceived and designed the experiments. Diribu Tesfaye and Feven Million: Performed the experiments.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no competing interests to declare that are relevant to the content of this article.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Tena, E., Tadesse, F., Tesfaye, D. et al. GGE Biplot Analysis of Exotic Sugarcane Genotypes in Major Sugarcane Producing Agro-ecologies of Ethiopia. Agric Res 13, 563–573 (2024). https://doi.org/10.1007/s40003-024-00721-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40003-024-00721-x