
Abstract
Background:
Infant adipose-derived mesenchymal stem cells (ADSCs) collected from excised polydactyly fat tissue, which was surgical waste, could be cultured and expanded in vitro in this study. In addition, the collecting process would not cause pain in the host. In this study, the proliferation, reduction of senescence, anti-oxidative ability, and differentiation potential in the infant ADSCs were compared with those in the adult ADSCs harvested from thigh liposuction to determine the availability of infant ADSCs.
Methods:
Proliferation was determined by detecting the fold changes in cell numbers and doubling time periods. Senescence was analyzed by investigating the age-related gene expression levels and the replicative stress. The superoxide dismutase (SOD) gene expression, adipogenic, neurogenic, osteogenic, and tenogenic differentiation were compared by RT-qPCR. The chondrogenic differentiation efficiency was also determined using RT-qPCR and immunohistochemical staining.
Results:
The proliferation, SOD (SOD1, SOD2 and SOD3) gene expression, the stemness-related gene (c-MYC) and telomerase reverse transcriptase of the infant ADSCs at early passages were enhanced compared with those of the adults'. Cellular senescence related genes, including p16, p21 and p53, and replicative stress were reduced in the infant ADSCs. The adipogenic genes (PPARγ and LPL) and neurogenic genes (MAP2 and NEFH) of the infant ADSC differentiated cells were significantly higher than those of the adults’ while the expression of the osteogenic genes (OCN and RUNX) and tenogenic genes (TNC and COL3A1) of both demonstrated opposite results. The chondrogenic markers (SOX9, COL2 and COL10) were enhanced in the infant ADSC differentiated chondrogenic pellets, and the expression levels of SODs were decreased during the differentiation process.
Conclusion:
Cultured infant ADSCs demonstrate less cellular senescence and replicative stress, higher proliferation rates, better antioxidant defense activity, and higher potential of chondrogenic, adipogenic and neurogenic differentiation.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
To use osteochondral grafts to reconstruct injured sites of articular cartilage remains challenging because there are limited sources of donor cartilage, and surgical scar formation may inhibit regeneration and cause morbidity to donor sites. To address the issues, mesenchymal stem cell (MSC) regeneration studies have been provided as an alternative.
Human MSCs isolated from bone marrow can be propagated in culture and differentiate into a variety of mesenchymal phenotypes, such as chondrocytes, osteoblasts, and adipocytes [1,2,3]. MSCs have become a promising tool for clinical applications in cartilage regeneration because they can be readily isolated and expanded in culture and have a limited tendency to form tumors [4]. In particular, adipose-derived mesenchymal stem cells (ADSCs) can be utilized as an alternative for bone and cartilage engineering and are easily and numerously available in adipose tissue [5, 6]. Compared with bone marrow, adipose tissue is a better source for MSC isolation because of less invasiveness, high cell yield, enhanced proliferation and better maintenance of stemness [7, 8]. Chondrogenic differentiation of ADSCs has exhibited treatment potential for osteoarthritis in a sheep model [9].
Chondrocytes and MSCs are the most general sources of cartilage tissue engineering [10, 11]. The proliferation of chondrocytes from surgical waste of polydactyly cartilage in children was compared with that in adults, and the result demonstrated that the cells from the waste of polydactyly cartilage had significantly higher growth rates than those from adults [12]. Up to the present, comparisons of adult ADSCs and infant ADSCs in chondrogenic potential have been less documented. Some studies revealed that using autologous ADSCs from elderly patients for potential therapeutic purposes towards chondrogenic regeneration might be restricted [13]. The degrees of proliferation and differentiation of infant ADSCs should be higher than those of adult ADSCs. ADSCs isolated from younger donors could inspire the host's self-healing capabilities and be feasibly applied in therapeutic treatment for skin regeneration [14]. Base on the aforementioned results, we hypothesize that infant ADSCs may have better chondrogenic differentiation potential than adult ADSCs in vitro. The positive results of this study will offer a new source of ADSCs that can be largely expanded in vitro for cell therapy.
The occurrence of polydactyly is about 1.6–10.9 per 1000 in general population and 0.3–3.6 per 1000 in live births, and it often happens to males twice as many in numbers as to females [15, 16]. The excised polydactyly fat tissue is generally considered as surgical waste. In fact, the precious cell source for cell engineering can be extracted from this kind of sample [12]. Adult ADSCs can be collected by liposuction, which is less invasive. In this study, we found the infant ADSCs could be derived from the polydactyly fat tissue; furthermore, the extracted infant ADSCs could be cultured and expanded to generate a large enough number of cells for experiments. Therefore, we are interested to find out if infant ADSCs increase the differentiation potential after in vitro expansion. If the infant ADSCs from surgical waste of polydactyly fat tissue are proved to preserve more stemness and better proliferation, they can be an ideal source for cell therapy. In addition, superoxide dismutase (SOD1, SOD2 and SOD3) can influence the therapeutic effects and differentiation abilities of MSCs [17,18,19]. SOD1, SOD2 and SOD3 may also play important roles in chondrogenic differentiation of infant ADSCs. The purposes of these experiments were aimed to investigate whether infant ADSCs increased the potential of chondrogenic differentiation in vitro and the roles of SOD1, SOD2 and SOD3 during chondrogenic differentiation of infant ADSCs.
2 Materials and methods
2.1 Isolation and expansion of the adult and infant ADSCs
The adult ADSCs (n = 3) collected from lipoaspirate were obtained from donors submitted to thigh liposuction under local anesthesia. The protocol and procedure were approved by the Institutional Review Board (IRB). Once the soft tissues were infiltrated with a solution of saline plus epinephrine (1:1,000,000), the subcutaneous fat was suctioned (− 500–− 700 mmHg) using a conventional liposuction machine equipped with a 2.5-mm (inner diameter) cannula. For the infant ADSCs (n = 3), the fat tissue was obtained from excised redundant thumbs after surgical reconstruction for children with polydactyly. The procedures were IRB-approved.
Each sample was digested in 0.1% collagenase (Wako, Osaka, Japan) Hank’s balanced salt solution (HBSS) by incubation in a shaking water bath (37 °C, 30 min). To remove the collagenase, stromal vascular fraction (SVF) cells had been washed with PBS three times [20, 21]. After filtration, the pellet was resuspended with an appropriate amount of the medium for cell counting and seeding in this time period (on a ~ 2–3 × 106/10cm2 dish). SVF cells were seeded in the Dulbecco’s Modified Eagle Media (DMEM), supplemented with 10% fetal bovine serum (FBS), and antibiotic–antimycotic solution (Corning life science, New York, NY, USA). The culture medium had been changed every two days and propagated at 1:5 dilution every four days when the cell confluence achieved 80%. The cells were cultured in 75% N2, 5% CO2 and 20% O2 for culture in the Forma Series II Water Jacketed CO2 Incubator (Thermo Fisher Scientific, Waltham, MA, USA).
2.2 Flow cytometric analysis
To analyze the cell surface expression of typical marker proteins, the ADSCs were harvested in 5 mmol/l EDTA in PBS. The cells were incubated with the following anti-human antibodies: CD29–phycoerythrin (PE), CD73–Allophycocyanin (APC), CD90–fluorescein isothiocyanate (FITC, Becton Dickinson, San Jose, CA, USA), CD105–cyanine (Cy5.5), CD44–PE, CD14–PE, CD79a–PE, CD11b–PE, CD19–PE, CD34–PE, CD45–PE, and HLA-DR-PE. Mouse isotype antibodies (Becton Dickinson and Beckman Coulter) were used as the controls. Ten thousand labeled cells were acquired and analyzed using the FACS CantoII Cytometer System running Diva software (Becton Dickinson).
2.3 Cell proliferation analysis
The adult and infant ADSCs were seeded at 160 cells/cm2 and cultured in 75% N2, 5% CO2 and 20% O2. The cells were reseeded at the same density every 9–12 days per passage and cultured under the same conditions. Fold increases in cell numbers and population doubling time periods of each passage were detected in triplicate cultures [22].
2.4 Senescence associated β-galactosidase (SA-β-gal) assays
To determine the cell senescence of the adult and infant ADSCs, a senescence β-galactosidase cell staining kit (Cell Signaling Technology, Danvers, MA, USA) was used. Briefly, the adult and infant ADSCs at passage 7 were seeded at a density of 1 × 105/ well on a 6-well plate with the culture medium and incubated for five days at 37 °C, 5% CO2, 20% O2. The cells were then fixed and stained following the manufacturer’s instructions. The senescent cells were stained in blue under a microscope.
2.5 γH2AX immunostaining
The adult and infant ADSCs at passage 3 were permeabilized with a permeabilization buffer (0.1% Triton X-100 in PBS) on glass slides, fixed with 4% paraformaldehyde, and then stained with primary antibodies against γ-H2AX (Taiclone Biotech Corp., Taipei, Taiwan) at an appropriate dilution. The DyLight 488-conjugated goat anti-rabbit IgG secondary antibodies (GeneTex, Irvine, CA, USA) were then added on the slides and incubated. The samples were also counterstained with 4, 6-diamidino-2-phenylindole (DAPI; Sigma-Aldrich, St. Louis, MO, USA). The total fluorescence intensity in arbitrary units from 60 to 100 cells was measured by the Image-Pro Plus (v4.5.0.29, Media Cybernetics, Silver Spring, MD, USA) and normalized with cell numbers.
2.6 Multilineage differentiation
The adipogenic inducing medium (AIM) comprised a culture medium supplemented with 10% FBS, 50 μg/ml ascorbic acid-2 phosphate, 0.1 μM dexamethasone, 50 μM indomethacin (Sigma-Aldrich), 45 μM 3-isobutyl-1-methylxanthine (Sigma-Aldrich), and 1 μg/ml insulin (Sigma-Aldrich), all of which had been used to induce adipogenic differentiation. The osteogenic inducing medium (OIM) included a culture medium supplemented with 10% FBS, 50 μg/ml ascorbic acid-2 phosphate (Nacalai, Kyoto, Japan), 0.01 μM dexamethasone (Sigma-Aldrich), and 1 mM b-glycerol phosphate (Sigma-Aldrich), all of which had been used to induce osteogenic differentiation. For the neuronal differentiation, the cells with 80% confluence were induced by a neurogenic inducing medium (NIM) supplemented with 50 μM All-trans retinoic acid (RA; Sigma-Aldrich) and 1 mM 2-Mercaptoethanol (β-ME; Sigma-Aldrich), both of which had been placed in a 37 °C, 5% CO2 and 20% O2 incubator for 24 h. Then the cells were maintained with DMEM solution containing 1% FBS for seven days [1]. The tenogenic inducing medium (TIM) was the culture medium with Glutamax (Thermo Fisher Scientific) supplemented of 0.1 μg Primocin (InvivoGen, San Diego, CA, USA), 50 μg/ml Ascorbic acid (AA; Sigma Aldrich), 50 ng/ml BMP-12, 100 ng/ml CTGF, and 10 ng/ml TGF-β3 (all from PeptroTech, London, UK). All the mediums had been changed every two days for a period of seven days. The cells cultured in a medium without any further supplementation were used as the controls [23].
2.7 Chondrogenic differentiation in vitro
2.5 × 105 adult or infant ADSCs at passages 3 to 5 were induced in an induction medium contained the serum-free and high-glucose (4.5 g/l) Dulbecco’s modified Eagle’s medium (HG-DMEM) (Gibco, Grand Island, NY, USA) with 50 mg/ml ITS plus Premix (BD Biosciences, San Jose, CA, USA), 6.25 mg/ml insulin, 6.25 mg/ml transferrin, 6.25 mg/ml selenious acid, 1.25 mg/ml bovine serum albumin, and 5.35 mg/ml linoleic acid. 10–7 M dexamethasone (Sigma-Aldrich) and 50 mg/ml ascobate-2-phosphate (Sigma-Aldrich) were added in presence of 10 ng/ml TGF-β1 (R&D Systems, Minneapolis, MN, USA) and cultured in 75% N2, 5% CO2 and 20% O2. The medium had been changed every three days, and the pellets had been observed and harvested on day 7, day 14, and day 21 during the induction [24].
2.8 Reverse transcription (RT) and real-time polymerase chain reaction (PCR) analysis
Total RNA was prepared using the Trizol reagent (Invitrogen, Mulgrave, Australia). For the first strand cDNA synthesis, random sequence primers were used to prime reverse transcription reactions, and synthesis was carried out by a SuperScript™ III Reverse Transcriptase (Invitrogen). cDNA was synthesized from the total RNA by means of the M-MuLV reverse transcriptase. The polymerase chain reaction (PCR) was performed with cDNA as the template in a 30-μl reaction mixture containing specific primer pairs which were listed in Table 1. Negative controls were used for each primer by replacing the RNA templates with RNA-free distilled water. PCR was performed by using a Taq DNA polymerase Recombinant (Invitrogen), and each cycle consisted of the following steps: denaturation for 45 s at 94 °C, annealing for 1 min at 51–58 °C, and 90 s of elongation at 72 °C. The β-actin expression was used to normalize the input template cDNA to analyze relative gene expression. The reaction products were resolved by electrophoresis on a 1.5% agarose gel and visualized with ethidium bromide. Real-time amplification of the genes was performed using the ABI Assays on Demand primers© and SYBR green® universal PCR master mix on the ABI 7500 real-time PCR machine according to the manufacturer’s instructions (Applied Biosystems, Foster City, CA, USA). To check the efficiency of the PCR amplification and cDNA synthesis, glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as an internal control. Analysis of the results was carried out using the software supplied with the machine using the delta CT method.
2.9 Immunohistochemistry staining and quantification of the chondrogenic pellets
The equivalent diameters (the diameter of the whole pellet) of the chondrogenic pellets formed by the ADSCs on day 21 after induction were measured. The pellets had been washed twice in PBS, fixed in 4% paraformaldehyde for three hours and prepared for paraffin embedding. Proteoglycan components on the sects. (4 mm thick) were stained with Alcian blue (ScyTek Laboratories, Logan, UT,, USA) combined with nuclear fast red staining (ScyTek). The sections were deparaffinized, hydrated, and treated with 0.4 mg/ml proteinase K in Tris–HCl (pH = 7) for 15 min, and endogenous peroxidase was blocked with 3% hydrogen peroxide (Sigma-Aldrich). After having been washed with PBS and incubated with the blocking serum for 5 min, the sections were exposed to primary antibodies against collagen type 2 (COL2; Abcam, Cambridge, UK) or collagen type 10 (COL10; Abcam) overnight at 4 °C. Then the sections were incubated with the secondary anti-rabbit polymer-horseradish peroxidase (HRP; Abcam) for 30 min. Staining was revealed with the DAB substrate and hematoxylin. The percentages of the stained areas in the whole sections were detected with the image analysis software (Image Pro Plus 4.0, Media Cybernetics Inc., Rockville, MD, USA).
2.10 Statistical analysis
Prism (version 5.03, GraphPad, La Jolla, CA, USA) was used to perform the quantitative analysis of the results. Data were presented in mean ± standard error. Statistical significance of the experimental results was determined by the Mann–Whitney U test or the one-way ANOVA analysis, and a result was significant when the p value was less than 0.05.
3 Results
3.1 Cell proliferation of the adult and infant human ADSCs
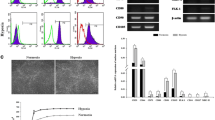
The cell-surface protein profiles of ADSCs were confirmed by flow cytometry. Both the adult and infant ADSCs were positive for CD29, CD44, CD73, CD90, and CD105 and negative for CD11b, CD14, CD19, CD34, CD45, CD79a, and HLA-DR (Fig. 1A). The cell morphologies of the adult and infant ADSCs at passage 3 were not noticeably different (Fig. 1B). The fold increases in the cell numbers of the infant ADSCs at passages 1 to 10 were significantly higher than those of the adult ADSCs (Fig. 1C), and the cell doubling time periods of the infant ADSCs were kept for one day through the passages that were significantly shorter than those of the adult ADSCs (Fig. 1D). Although the proliferation of the infant ADSCs might decline after five passages, the results indicated that the proliferation of the ADSCs collected from the infants were better-preserved.

Characterization of the adult and infant adipose-derived mesenchymal stem cells (ADSCs). A The isolated human adult and infant ADSCs detected with antibodies, which were represented by solid lines, against CD29, CD44, CD73, CD90 and CD105 were positive, and those against CD11b, CD19, CD34, CD45, CD14b, CD79a, and HLA-DR were negative. All the detections were determined using the FACS CantoII Cytometer System. The dashed lines represented isotype control staining. B The morphologies of the adult and infant ADSCs at passage 3 were imaged using phase contrast microscopy. (magnification × 40; the scale bar = 200 μm; magnification × 200; the scale bar = 200 μm) C The cell numbers of every passage, which were counted before reseeding, were divided by the initial cell number to calculate fold increases. The fold increases in the cell numbers and D cell doubling time periods of the adult and infant ADSCs were calculated from passages 1 (P1) to 10 (P10). Mean ± SD with three experimental replicates were expressed. Statistical significance of comparing the adult and infant ADSCs was determined by the unpaired t test. “*” represented p < 0.05. “**” represented p < 0.01. “***” represented p < 0.001
3.2 Senescence and replicative stress in the adult and infant ADSCs
The expression of aging-related genes, including p16 (Fig. 2A), p21 (Fig. 2B), and p53 (Fig. 2C), was significantly downregulated in the infant ADSCs at early passages compared with that in the adult ADSCs. However, the p53 gene was significantly upregulated in infant ADSCs at late passages compared with that at early passages. The stemness-related gene (c-MYC) and telomerase reverse transcriptase (TERT), which is consistent with the telomerase activity, were significantly upregulated in the infant ADSCs at early passages compared with those in the adult ADSCs. Nevertheless, they were downregulated in the infant ADSCs at late passages compared with those at early passages (Figs. 2D, E). SA-β-gal assays showed that the adult ADSCs at passage 7 were more positive than the infant ones (Fig. 2F). Furthermore, the immunofluorescent signals of γH2AX, as markers of replicative stress, in the nuclei of the infant ADSCs were showed to be less positive than those in the adult ones (Fig. 2G).

Senescence and replicative stress in the adult and infant ADSCs. A P16, B P21, C P53, D c-MYC, and E TERT genes of the adult, infant ADSCs at passages 3–5 (early passages), and infant ADSCs at passages 9–10 (late passages) were detected by RT-qPCR, and the values were normalized to the expression of glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Mean ± SD were expressed. The values of the adult ADSCs were used as the controls. Statistical significance of comparing the adult and infant ADSCs was determined by the one-way ANOVA analysis. * p < 0.05. F The cell senescence of the differentiated adult and infant ADSCs at passage 7 was detected by the Senescence Detection Kit. The left panels showed the high cell density, and right ones showed the low cell density. Senescence-associated β-galactosidase (SA-β-gal) positive was stained and exhibited in blue. (magnification × 200; the scale bar = 100 μm). G The adult and infant ADSCs were immunostained with an anti-γH2AX antibody and DAPI. (magnification × 400; the scale bar = 20 μm) H The values (arbitrary unit) of the immunofluorescence intensity of γH2AX in the nuclei of the adult and infant ADSCs were quantified by the Image-Pro Plus v4.5.0.29 and normalized to the cell numbers. Mean ± SD with three experimental replicates were expressed in B, C, D, and F. Statistical significance of comparing the adult and infant ADSCs was determined by the Mann–Whitney U test. “*” represented p < 0.05. “**” represented p < 0.01
3.3 Comparison of the adult and infant ADSCs in multilineage differentiation
The adipogenic, osteogenic, neurogenic, and tenogenic differentiation in the adult and infant ADSCs was all determined by RT-qPCR. Peroxisome proliferator-activated receptor γ (PPARγ) and lipoprotein lipase (LPL) genes were detected to verify the adipogenic differentiation in the adult and infant ADSCs on day 14 (Figs. 3A, B). Osteocalcin (OCN) and runt-related transcription factor (RUNX) genes were used to determine the osteogenic differentiation of the adult and infant ADSCs (Fig. 3C, D). Tendon-related genes, including tenascin-C (TNC) and collagen type 3 alpha 1 chain (COL3A1), were used to determine the tenogenic differentiation (Fig. 3E, F), and microtubule-associated protein 2 (MAP2) and neurofilament heavy polypeptide (NEFH) were detected to determine the neurogenic differentiation (Figs. 3G, H). Adipogenic and neurogenic genes were highly expressed in the factor-induced differentiation of the infant ADSCs, which explained the fact that the differentiation potential of the infant ADSCs was significantly higher than that of the adult ones. However, the expression levels of the osteogenic and tenogenic genes in the differentiated infant ADSCs were significantly lower than those in the adult ones. In addition, the multilineage differentiation of the infant and adult ADSCs, including adipogenic, osteogenic, tenogenic, and neurogenic lineages, was confirmed by histological and immunofluorescence staining (Supplementary Figure S1).

Detection of the adipose-, osteocyte-, tendon- and neuron-related gene expression levels of the adult and infant ADSCs during differentiation. Adipose-related genes, including peroxisome proliferator-activated receptor-γ (PPARγ) A and lipoprotein lipase (LPL) B, osteocyte-related genes, including osteocalcin (OCN) C and runt-related transcription factor (RUNX) D, and tendon-related genes, including tenascin-C (TNC) E and collagen type 3 alpha 1 chain (COL3A1) F of the ADSCs after the factor-induced (AIM, OIM and TIM) differentiation for 14 days were detected by RT-qPCR. Neuron-related genes, including microtubule-associated protein 2 (MAP2) G and neurofilament heavy polypeptide (NEFH) H of the ADSCs after the factor-induced (NIM) differentiation for 7 days were detected. Each value of the gene expression was normalized to the expression of glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Relative fold changes of the values of the factor-induced and confluence-initiated differentiated ADSCs were compared with those of the controls (day 0) as one. The values were expressed as mean ± SEM. with three experimental replicates. Statistical significance was determined using the Mann–Whitney U test. “*” represented p < 0.05. “**” represented p < 0.01. “***” represented p < 0.001
3.4 Chondrogenic differentiation of the infant and adult human ADSCs in vitro
The infant ADSCs in the chondrogenic induction medium formed a larger pellet (Fig. 4A). The diameters of the pellets which were formed from the adult and infant ADSCs were measured on day 21. The mean diameter of the chondrogenic pellets from the adult ADSCs at passages 2 to 5 was 2 mm, and that of the pellets from the infant ADSCs was 2.5 mm, which was significantly longer than that from the adult ADSCs (Fig. 4B).

In vitro chondrogenic differentiation of the adult and infant ADSCs. A The chondrogenic pellets differentiated from P3-P5 of the adult and infant ADSCs (3 × 105 cells) on day 21. (The scale bar = 1 mm) B The diameters of the pellets were measured after differentiation on day 21. “*” represented p < 0.05
The SOX9 gene expression in the infant ADSC differentiated chondrogenic pellets on day 7 was significantly higher than that in the pellets formed from the adult ADSCs (Fig. 5A). The COL2 (Fig. 5B) and COL10 (Fig. 5C) expression levels were also significantly higher in the chondrogenic pellets formed from the infant ADSCs on day 21, compared with those formed from the adult ADSCs.

Chondrocyte-related gene expression in the adult and infant ADSC differentiated pellets. The expression levels of A SOX9 on day 7, B COL2, and C COL10 on day 21 of the adult and infant ADSC differentiated pellets had been detected by RT-qPCR during the differentiation from day 7 to day 21, and the values were normalized to the expression of glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Mean ± SD were expressed. The values of the adult ADSCs were used as the controls. Statistical significance of comparing the adult and infant ADSCs was determined by the Mann–Whitney U test. “*” represented p < 0.05
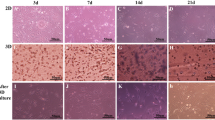
The glycosaminoglycan (GAG) content of the pellets generated from the adult and infant ADSCs was stained by Alcian blue (Figs. 6A, B). In addition, COL2 and COL10 protein expression levels were detected by immunohistochemistry staining (Figs. 6C–F). The GAG intensities of the infant ADSC generated chondrogenic pellets stained by Alcian blue were significantly higher than those of the adult ADSCs (Fig. 6G). Both COL2 and COL10 expression levels in the pellets formed from the infant ADSCs were significantly higher than those formed from the adult ADSCs (Figs. 6H, I).

Immunohistochemistry staining of the chondrogenic differentiated adult and infant ADSCs. A, B The pellets of the 21-day chondrogenic differentiated adult and infant ADSCs were sectioned and stained using Alcian blue. The protein expression levels of C, D collagen type II (COL2) and E, F collagen type X (COL10) were detected by immunostaining. (magnification × 40; the scale bar = 200 μm) G-I The densities of each staining were calculated and normalized with the whole backgrounds of the pellet sections. Mean ± SD with three experimental replicates were expressed. Statistical significance of comparing the adult and infant ADSCs was determined by the Mann–Whitney U test. “*” represented p < 0.05
3.5 SOD expression levels in the adult and infant ADSCs during the chondrogenic differentiation
The expression levels of the superoxide dismutase genes involving SOD1 (Fig. 7A), SOD2 (Fig. 7B), and SOD3 (Fig. 7C) in the infant ADSCs were all significantly higher than those in the adult ADSCs. The expression levels of these three SODs in the adult ADSCs and SOD1 in the infant ADSCs were all significantly increased on day14 and then dropped to the lowest on day 21 through the process of chondrogenic differentiation (Figs. 7D-G). The SOD2 expression in the infant ADSCs significantly rose on day 7 and also declined to the lowest on day 21 (Fig. 7H). The result of the SOD3 expression in the infant ADSCs was similar to that of SOD2, but it was not significantly different between different time periods (Fig. 7I).

Superoxide dismutase (SOD) expression levels in the adult and infant ADSCs. A SOD1, B SOD2, and C SOD3 genes of the adult and infant ADSCs (day 0) were detected by RT-qPCR, and the values were normalized to the expression of glyceraldehyde-3-phosphate dehydrogenase (GAPDH). The values of the adult ADSCs were used as the controls. Mean ± SD were expressed. Statistical significance of comparing the adult and infant ADSCs was determined by the Mann–Whitney U test. The levels of SOD1, SOD2 and SOD3 gene expression in the adult ADSCs D–F and infant ADSCs G–I during the time course of chondrogenic differentiation on day 0 (D0), day7 (D7), day14 (D14), and day 21 (D21) had been determined using RT-qPCR. The values of day 0 were used as the controls. Statistical significance of comparing the differences of different time periods was determined by the one-way ANOVA analysis. “*” represented p < 0.05. “**” represented p < 0.01. “***” represented p < 0.001
4 Discussion
The infant ADSCs were collected from the excised infant polydactyly fat tissue which was considered as surgical waste. However, it is a precious cell source of cell therapy [12]. In this study, the proliferation, anti-aging ability, antioxidation ability, and chondrogenic differentiation in the infant ADSCs were proved to be better than those in the adult ADSCs. The infant allogeneic ADSC culture is a useful strategy to significantly reduce cell senescence and advance for future chondrogenic differentiation and animal model studies.
ADSCs isolated from younger donors could inspire the host's self-healing capabilities and be feasibly applied in therapeutic treatment for skin regeneration [25]. Advancement of the donor age would diminish the function of stem cells in angiogenesis [26, 27]. Prolonged expansion caused MSC senescence, exhibited an enlarged flat cell morphology, increased SA-β-gal activity, demonstrated loss of cell properties, and increased the expression levels of senescence related genes, including p16, p21, and p53 [28]. MSC senescence is also a key factor that affects the potential of multi-lineage differentiation [29]. Senescent cell secreted factors might influence some fundamental processes, such as cell growth, cell migration, tissue construction, blood vessel formation, and differentiation. Part of the secreted factors are inflammatory cytokines that may cause chronic inflammation [30]. In addition, it has been reported that the pool size of MSCs reduces in aged animals [31]. ADSCs obtained from infant patients (less than one year old) with an end-stage liver disease during liver transplantation by subcutaneous abdominal surgery have more biological advantages than those obtained from older ones [32].
Senescence is started by the retinoblastoma protein (Rb) or p53 pathways, and then activates the cyclin-dependent kinase inhibitors p16 and p21, respectively. The function of each pathway can influence and collaborate to block the cell cycle and growth arrest and then cause senescence [33]. The p16, p21, and p53 have been found to be upregulated in long-term expansion of stem cells [34]. The p16 and p21 gene upregulations in aged tendon stem progenitor cells have been found to be in agreement with those in skeletal muscle aging. The local stem cells with dysregulation in the p16 signaling is related to the senescence of muscle tissue [35]. On the other hand, c-MYC is involved in controlling cell proliferation and differentiation, and TERT is consistent with telomerase activity and preserves the chromosome integrity [36]. In this study, we confirmed that the infant ADSCs indeed downregulated the expression levels of p16, p21, and p53, and enhanced the c-MYC and TERT expression levels at early passages. However, the levels of c-MYC and TERT expression would decline after long-term culture. MSCs from younger donors with longer telomeres have higher levels of telomerase activity [37]. The isolated MSCs from telomerase knock-out mice at early passages lost the ability of adipogenic and chondrogenic differentiation [38]. A study which compared the chondrogenic differentiation of rat BMSCs with ages of one week, 12 weeks, and one year showed that COL 2 and aggrecan expression levels declined with increasing ages [39]. In this study, the increase folds of cell numbers and the chondrogenic differentiation related genes and proteins were all significantly enhanced in the infant ADSCs and differentiated pellets compared with those in the adult ones. The results indicated that, under the same in vitro induction foundation, the ADSCs from infants with higher expression levels of telomerase genes had higher potential of proliferation and differentiation than those from adults. γH2AX, which is the histone variant with phosphorylation on the Ser-139 residue, acts as an early response to the DNA double-strand breaks [40], which will induce terminal cell cycle arrest and cellular senescence [41, 42]. The results of the SA-β-gal assays and the replicative stress, which were determined by detecting the expression of γH2AX, were less positive in the infant ADSCs; consequently, we confirmed the proliferation and replication abilities of the infant ADSCs were better than those of the adult ones.
Some evidence exhibited that reactive oxygen species (ROS) could enhance the stemness of ADSCs [43] and initiate proliferation after quiescence exit [44]. Nevertheless, an excess amount of ROS could reduce the function of MSCs and cause cell death [45]. Copper/zinc-superoxide dismutase 1 (SOD1), which is a dimeric cytosolic enzyme that detoxifies superoxide anions to H2O2 [46], was shown to be highly neuroprotective against reperfusion injury after focal and global cerebral ischemia [47,48,49]. In addition, the ability of SOD1 has been studied so as to advance the therapeutic potential of ADSCs to resist ischemic damage [19]. The fact that Mn-superoxide dismutase 2 (SOD2) enhances the therapeutic effects of ADSCs has also been studied [17]. The expressions of SOD3 in BMSCs are influenced during adipogenic, chondrogenic, and osteogenic differentiation [18]. The antioxidant enzymes are among the major mechanisms by which cells counteract the deleterious effects of ROS [47]. However, it has not yet been determined whether SOD1, SOD2 and SOD3 play critical roles in enhancing chondrogenic differentiation of ADSCs from oxidative stress. Therefore, the purpose of this study was to determine the relationship between the expression levels of SODs (SOD1, SOD2 and SOD3) and chondrogenic differentiation of infant ADSCs. We have found that the expression levels of SODs in the infant ADSCs were higher than those in the adult ADSCs before chondrogenic differentiation. The expression levels of SODs of ADSCs declined following differentiation, and those of the infant ADSCs were even lower than those of the adult ones.
ROS has been found to be required during the in vitro induction of chondrogenic and adipogenic differentiation. The chondrogenic markers were upregulated by adding H2O2 during the differentiation, which was suppressed when the antioxidant N-acetylcysteine (NAC) was added [50]. TGF-β1, an inducer of chondrogenic differentiation, also caused a pro-oxidant effect in cartilage, but not cytotoxicity [51]. Besides, NADPH oxidase 2 and 4 (Nox2 and Nox4) generated ROS was increased during the early stage of chondrogenic differentiation [52]. All of these studies suggested that ROS played an important role in chondrogenic differentiation, and also explained the decline of the expression levels of SODs during the differentiation process.
An age dependent reduction in adipogenic differentiation of mouse ADSCs has been found, but not in osteogenic differentiation [53]. However, a result in rats demonstrated that an advanced age decreased the gene expression of adipogenic differentiation [54]. The propensity for adipogenic differentiation decreased in senescent BMSCs, whereas osteogenic differentiation increased [34]. According to a theoretical inverse relationship between osteogenic and adipogenic differentiation, differentiation towards an osteoblast phenotype exists at the expense of an adipocytic phenotype, or vice versa [55]. We also found that the infant ADSCs had the tendency toward chondrogenic, adipogenic, and neurogenic differentiation but had less osteogenic and tenogenic potential, and the adult ADSCs showed the opposite tendency. This result suggested that infant and adult ADSCs had different degrees of differentiation potential under the same differentiation induction.
There are some limitations in this study. The sample numbers of ADSC might not to be large enough to reduce the inter-individual variability. The detection time periods of different differentiation levels were diverse due to the distinct cell proliferation during the differentiation processes. The ADSCs would not survive for a long period in some differentiation processes.
In this study, we harvested the infant ADSCs from excised polydactyly fat tissue that resolved the problem of collection. The difficulty of collecting surgical specimen is still an issue for infant ADSCs to be used. However, infant ADSCs show better abilities of proliferation, less senescence, and more potential of differentiation. The expanded infant ADSCs cell line could still be wildly applied, even to treat child-related arthritis such as juvenile rheumatoid arthritis. To sum up, infant ADSCs collected from excised polydactyly fat tissue is a useful strategy to reduce cell senescence and advance for chondrogenic differentiation. Infant ADSCs show an advantageous application in cartilage regeneration and repair.
References
Hung SC, Cheng H, Pan CY, Tsai MJ, Kao LS, Ma HL. In vitro differentiation of size-sieved stem cells into electrically active neural cells. Stem Cells. 2002;20:522–9.
Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD, et al. Multilineage potential of adult human mesenchymal stem cells. Science. 1999;284:143–7.
Prockop DJ. Marrow stromal cells as stem cells for nonhematopoietic tissues. Science. 1997;276:71–4.
Tuan RS, Boland G, Tuli R. Adult mesenchymal stem cells and cell-based tissue engineering. Arthritis Res Ther. 2003;5:32–45.
Hildner F, Albrecht C, Gabriel C, Redl H, van Griensven M. State of the art and future perspectives of articular cartilage regeneration: A focus on adipose-derived stem cells and platelet-derived products. J Tissue Eng Regen Med. 2011;5:e36-51.
Ogura F, Wakao S, Kuroda Y, Tsuchiyama K, Bagheri M, Heneidi S, et al. Human adipose tissue possesses a unique population of pluripotent stem cells with nontumorigenic and low telomerase activities: potential implications in regenerative medicine. Stem Cells Dev. 2014;23:717–28.
Zhu Y, Liu T, Song K, Fan X, Ma X, Cui Z. Adipose-derived stem cell: A better stem cell than BMSC. Cell Biochem Funct. 2008;26:664–75.
Sharath SS, Ramu J, Nair SV, Iyer S, Mony U, Rangasamy J. Human Adipose tissue derivatives as a potent native biomaterial for tissue regenerative therapies. Tissue Eng Regen Med. 2020;17:123–40.
Ude CC, Sulaiman SB, Min-Hwei N, Hui-Cheng C, Ahmad J, Yahaya NM, et al. Cartilage regeneration by chondrogenic induced adult stem cells in osteoarthritic sheep model. PLoS One. 2014;9:e98770.
Mehrabani D, Mojtahed Jaberi F, Zakerinia M, Hadianfard MJ, Jalli R, Tanideh N, et al. The healing effect of bone marrow-derived stem cells in knee osteoarthritis: A case report. World J Plast Surg. 2016;5:168–74.
Manafi A, Hamedi ZS, Manafi A, Rajabiani A, Rajaee A, Manafi F. Injectable cartilage shaving: An autologous and long lasting filler material for correction of minor contour deformities in rhinoplasty. World J Plast Surg. 2015;4:93–100.
Mortazavi F, Shafaei H, Soleimani Rad J, Rushangar L, Montaceri A, Jamshidi M. High quality of infant chondrocytes in comparison with adult chondrocytes for cartilage tissue engineering. World J Plast Surg. 2017;6:183–9.
Ye X, Liao C, Liu G, Xu Y, Tan J, Song Z. Age-Related Changes in the regenerative potential of adipose-derived stem cells isolated from the prominent fat pads in human lower eyelids. PLoS One. 2016;11:e0166590.
Ma N, Qiao C, Zhang W, Luo H, Zhang X, Liu D, et al. Original Research: Adipose-derived stem cells from younger donors, but not aging donors, inspire the host self-healing capability through its secreta. Exp Biol Med (Maywood). 2017;242:68–79.
Castilla E, Paz J, Mutchinick O, Muñoz E, Giorgiutti E, Gelman Z. Polydactyly: A genetic study in South America. Am J Hum Genet. 1973;25:405–12.
Umair M, Ahmad F, Bilal M, Ahmad W, Alfadhel M. Clinical genetics of polydactyly: An updated review. Front Genet. 2018;9:447.
Sen S, Domingues CC, Rouphael C, Chou C, Kim C, Yadava N. Genetic modification of human mesenchymal stem cells helps to reduce adiposity and improve glucose tolerance in an obese diabetic mouse model. Stem Cell Res Ther. 2015;6:242.
Nightingale H, Kemp K, Gray E, Hares K, Mallam E, Scolding N, et al. Changes in expression of the antioxidant enzyme SOD3 occur upon differentiation of human bone marrow-derived mesenchymal stem cells in vitro. Stem Cells Dev. 2012;21:2026–35.
Yoo DY, Kim DW, Chung JY, Jung HY, Kim JW, Yoon YS, et al. Cu, Zn-superoxide dismutase increases the therapeutic potential of adipose-derived mesenchymal stem cells by maintaining antioxidant enzyme levels. Neurochem Res. 2016;41:3300–7.
Yoshimura K, Shigeura T, Matsumoto D, Sato T, Takaki Y, Aiba-Kojima E, et al. Characterization of freshly isolated and cultured cells derived from the fatty and fluid portions of liposuction aspirates. J Cell Physiol. 2006;208:64–76.
Wu SH, Shirado T, Mashiko T, Feng J, Asahi R, Kanayama K, et al. Therapeutic effects of human adipose-derived products on impaired wound healing in irradiated tissue. Plast Reconstr Surg. 2018;142:383–91.
Tsai CC, Chen YJ, Yew TL, Chen LL, Wang JY, Chiu CH, et al. Hypoxia inhibits senescence and maintains mesenchymal stem cell properties through down-regulation of E2A–p21 by HIF-TWIST. Blood. 2011;117:459–69.
Stanco D, Caprara C, Ciardelli G, Mariotta L, Gola M, Minonzio G, et al. Tenogenic differentiation protocol in xenogenic-free media enhances tendon-related marker expression in ASCs. PLoS One. 2019;14:e0212192.
Hung SC, Chen NJ, Hsieh SL, Li H, Ma HL, Lo WH. Isolation and characterization of size-sieved stem cells from human bone marrow. Stem Cells. 2002;20:249–58.
Pfaller MA, Diekema DJ. Epidemiology of invasive candidiasis: A persistent public health problem. Clin Microbiol Rev. 2007;20:133–63.
Dimmeler S, Vasa-Nicotera M. Aging of progenitor cells: Limitation for regenerative capacity? J Am Coll Cardiol. 2003;42:2081–2.
Rivard A, Fabre JE, Silver M, Chen D, Murohara T, Kearney M, et al. Age-dependent impairment of angiogenesis. Circulation. 1999;99:111–20.
Fafian-Labora JA, Morente-López M, Arufe MC. Effect of aging on behaviour of mesenchymal stem cells. World J Stem Cells. 2019;11:337–46.
Drela K, Stanaszek L, Nowakowski A, Kuczynska Z, Lukomska B. Experimental strategies of mesenchymal stem cell propagation: Adverse events and potential risk of functional changes. Stem Cells Int. 2019;2019:7012692.
Davalos AR, Coppe JP, Campisi J, Desprez PY. Senescent cells as a source of inflammatory factors for tumor progression. Cancer Metastasis Rev. 2010;29:273–83.
Kasper G, Mao L, Geissler S, Draycheva A, Trippens J, Kühnisch J, et al. Insights into mesenchymal stem cell aging: Involvement of antioxidant defense and actin cytoskeleton. Stem Cells. 2009;27:1288–97.
Jin Y, Yang L, Zhang Y, Gao W, Yao Z, Song Y, et al. Effects of age on biological and functional characterization of adiposederived stem cells from patients with endstage liver disease. Mol Med Rep. 2017;16:3510–8.
Beauséjour CM, Krtolica A, Galimi F, Narita M, Lowe SW, Yaswen P, et al. Reversal of human cellular senescence: Roles of the p53 and p16 pathways. EMBO J. 2003;22:4212–22.
Cheng H, Qiu L, Ma J, Zhang H, Cheng M, Li W, et al. Replicative senescence of human bone marrow and umbilical cord derived mesenchymal stem cells and their differentiation to adipocytes and osteoblasts. Mol Biol Rep. 2011;38:5161–8.
Yan Z, Yin H, Brochhausen C, Pfeifer CG, Alt V, Docheva D. Aged tendon stem/progenitor cells are less competent to form 3D tendon organoids due to cell autonomous and matrix production deficits. Front Bioeng Biotechnol. 2020;8:406.
Wu KJ, Grandori C, Amacker M, Simon-Vermot N, Polack A, Lingner J, et al. Direct activation of TERT transcription by c-MYC. Nat Genet. 1999;21:220–4.
Wright WE, Shay JW. Historical claims and current interpretations of replicative aging. Nat Biotechnol. 2002;20:682–8.
Liu L, DiGirolamo CM, Navarro PA, Blasco MA, Keefe DL. Telomerase deficiency impairs differentiation of mesenchymal stem cells. Exp Cell Res. 2004;294:1–8.
Zheng H, Martin JA, Duwayri Y, Falcon G, Buckwalter JA. Impact of aging on rat bone marrow-derived stem cell chondrogenesis. J Gerontol A Biol Sci Med Sci. 2007;62:136–48.
Mah LJ, El-Osta A, Karagiannis TC. γH2AX: A sensitive molecular marker of DNA damage and repair. Leukemia. 2010;24:679–86.
Li H, Mitchell JR, Hasty P. DNA double-strand breaks: A potential causative factor for mammalian aging? Mech Ageing Dev. 2008;129:416–24.
Mah LJ, El-Osta A, Karagiannis TC. γH2AX as a molecular marker of aging and disease. Epigenetics. 2010;5:129–36.
Cheng NC, Hsieh TY, Lai HS, Young TH. High glucose-induced reactive oxygen species generation promotes stemness in human adipose-derived stem cells. Cytotherapy. 2016;18:371–83.
Lyublinskaya OG, Borisov YG, Pugovkina NA, Smirnova IS, Obidina JV, Ivanova JS, et al. Reactive oxygen species are required for human mesenchymal stem cells to initiate proliferation after the quiescence exit. Oxid Med Cell Longev. 2015;2015:502105.
Chang W, Song BW, Moon JY, Cha MJ, Ham O, Lee SY, et al. Anti-death strategies against oxidative stress in grafted mesenchymal stem cells. Histol Histopathol. 2013;28:1529–36.
McCord JM, Fridovich I. Superoxide dismutase. An enzymic function for erythrocuprein (hemocuprein). J Biol Chem. 1969;244:6049–55.
Chan PH. Reactive oxygen radicals in signaling and damage in the ischemic brain. J Cereb Blood Flow Metab. 2001;21:2–14.
Kinouchi H, Epstein CJ, Mizui T, Carlson E, Chen SF, Chan PH. Attenuation of focal cerebral ischemic injury in transgenic mice overexpressing CuZn superoxide dismutase. Proc Natl Acad Sci U S A. 1991;88:11158–62.
Yang G, Chan PH, Chen J, Carlson E, Chen SF, Weinstein P, et al. Human copper-zinc superoxide dismutase transgenic mice are highly resistant to reperfusion injury after focal cerebral ischemia. Stroke. 1994;25:165–70.
Heywood HK, Lee DA. Bioenergetic reprogramming of articular chondrocytes by exposure to exogenous and endogenous reactive oxygen species and its role in the anabolic response to low oxygen. J Tissue Eng Regen Med. 2017;11:2286–94.
Jallali N, Ridha H, Thrasivoulou C, Butler P, Cowen T. Modulation of intracellular reactive oxygen species level in chondrocytes by IGF-1, FGF, and TGF-beta1. Connect Tissue Res. 2007;48:149–58.
Kim KS, Choi HW, Yoon HE, Kim IY. Reactive oxygen species generated by NADPH oxidase 2 and 4 are required for chondrogenic differentiation. J Biol Chem. 2010;285:40294–302.
Shi YY, Nacamuli RP, Salim A, Longaker MT. The osteogenic potential of adipose-derived mesenchymal cells is maintained with aging. Plast Reconstr Surg. 2005;116:1686–96.
Kirkland JL, Hollenberg CH, Gillon WS. Ageing, differentiation, and gene expression in rat epididymal preadipocytes. Biochem Cell Biol. 1993;71:556–61.
James AW. Review of signaling pathways governing MSC osteogenic and adipogenic differentiation. Scientifica (Cairo). 2013;2013:684736.
Acknowledgements
We thank the work that was assisted in part by the Division of the Experimental Surgery of the Department of Surgery, Taipei Veterans General Hospital. This study was supported in part by grants from the Taipei Veterans General Hospital (V107C-108), the Ministry of Science and Technology (MOST 106–2314-B-075–060), the Taoyuan General Hospital, Ministry of Health and Welfare (PTH110024), and the National Yang Ming Chiao Tung University Hospital (RD0021-008).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no competing interests.
Ethical statement
The collection of human ADSCs was approved by the Institutional Review Board (IRB) of the Taipei Veterans General Hospital (IRB No. 2020–06-010B), and the adipose tissue was collected with patient informed consent. In addition, the procedures involving animals were approved by the Institutional Animal Care and Use Committee of the Taipei Veterans General Hospital.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Wu, SH., Yu, JH., Liao, YT. et al. Comparison of the Infant and Adult Adipose-Derived Mesenchymal Stem Cells in Proliferation, Senescence, Anti-oxidative Ability and Differentiation Potential. Tissue Eng Regen Med 19, 589–601 (2022). https://doi.org/10.1007/s13770-022-00431-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13770-022-00431-x