
Abstract
The increase in the production of tomato, Solanum lycopersicon Mill. (Solanaceae), has favored the proliferation of pests, especially Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). In this study, the development, reproduction, survival, and demographic parameters of T. absoluta reared on six commercial tomato cultivars (Cherry, Cordilheira, Giuliana, Nemoneta, Paron, and Santa Clara) were evaluated. Tuta absoluta completed its development in all tomato cultivars. Development from newly hatched caterpillar to newly emerged adult varied between 24.8 and 28.2 days. Female fecundity ranged from 126.3 to 166.9 eggs, with fertility from 54.2 to 84.1%. Mortality during egg-adult development varied between 21.4 and 46.4% for insects reared on cultivars Cherry and Giuliana, respectively. The cultivars Cordilheira, Giuliana, and Santa Clara are promising options to tomato producers in order to decrease the attack and proliferation of T. absoluta. However, the development and population growth of T. absoluta is faster on the tomato cultivar Cherry.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The tomato leafminer, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae), is an olyphagous microlepidopteran pest that attacks species of the family Solanaceae, especially tomatoes. Larvae penetrate the leaves and feed on the mesophyll, affecting the photosynthetic capacity of the plant (Fernandez & Montagne 1990, Uchôa-Fernandes et al 1995, Pereyra & Sánchez 2006). In addition, injury caused directly to fruits can result in severe yield losses (Colomo & Berta 1995).
Tuta absoluta is a major pest of tomato in South America (Torres et al 2001) and worldwide (Desneux et al 2011). More recently, it has been reported in Spain and several other countries of Europe and the Middle East (Viggiani et al 2009, Desneux et al 2010, Kiliç 2010). An estimated 27.2% of tomatoes produced in the world are infested by T. absoluta (Desneux et al 2011).
Insect growth and reproduction depend on the quality and quantity of food resources available (Hagen et al 1984). To assess the effect of a host on the population growth of an insect, population parameters can be used, such as intrinsic rate of increase (R m), net reproductive rate (R 0), and generation time (T) based on survival, fertility, development time, and sex ratio (Sauvion et al 2005). Also, interactions with plants are important factors in the development of herbivorous insects (Jaenike 1978, Pizzamiglio 1991), while the susceptibility of plants to insects depends on their chemical and morphological characteristics, as well as their ecological environment and protective compounds (Han et al 2014, 2015, Larbat et al 2016). This information on resistance and susceptibility of cultivars (Maluf et al 1997, Ecole et al 2000, 2001, Medeiros et al 2006) and biological data on pest populations reared on different commercial cultivars can then be used in the development of pest management strategies. In this study, the development, reproduction, and survival of T. absoluta were evaluated on six commercial tomato cultivars. In addition, fertility tables were constructed to compare demographic parameters of this species reared on these genotypes.
Material and Methods
Insect rearing
Tuta absoluta rearing started in March 2010 with caterpillars and pupae collected in an organic commercial field of Cherry tomatoes in the municipality of Colombo, metropolitan area of Curitiba, state of Paraná (25°17′31″S, 49°13′26″W), Brazil (Krechemer & Foerster 2015). The insects were reared for 2 years on the tomato cultivar Santa Clara, and field-collected larvae were continuously added to the laboratory stock population.
In order to standardize the tests, insects were reared on the cultivar Carmen for three generations before the experiments were conducted. The rearing procedures followed Krechemer & Foerster (2015).
Development and survival of Tuta absoluta on different tomato cultivars
The development and survival of immature stages of T. absoluta were evaluated on six tomato cultivars (Cherry, Cordilheira, Giuliana, Nemoneta, Paron, and Santa Clara) under controlled climatic conditions (20 ± 2°C, 70 ± 10% RH, and 12 h photophase). One hundred forty newly hatched caterpillars for each tomato cultivar were individualized in polyethylene containers (7 × 4 cm). Each individual received one tomato foliole washed in 1% (v/v) sodium hypochlorite with its petiole wrapped in cotton moistened in water to keep leaves turgid.
Tomato leaves were replaced when caterpillars consumed approximately 70% of its area or when the leaves began to wither to allow larvae to move into them. Pupae were removed from leaves, sexed, and maintained in polyethylene containers until emergence of adults. Adults were randomly separated into couples based on emergence date (next section).
After adults died, the right posterior tibia and wing were removed, and their lengths measured with a Wild Heerbrugg M3 stereo microscope fitted with an eyepiece reticle. Measurements were later calculated using conversion tables. Egg incubation time was recorded for adults reared for each tomato cultivar. The number of eggs produced per couple was considered a sample, with a total of 2147 (Cherry), 2061 (Cordilheira), 2892 (Giuliana), 2059 (Nemoneta), 2837 (Paron), and 2209 (Santa Clara) eggs. Following incubation, the duration of larval and pupal stages was determined.
Reproduction and longevity of Tuta absoluta on different tomato cultivars
Fecundity, fertility, and longevity of T. absoluta adults obtained from caterpillars reared on each cultivar were determined with 20 replications per cultivar. Each couple was formed based on emergence date and placed in 10 cm tall by 10 cm in diameter cages. A tomato leaf was used as stimulus for oviposition in each cage (Krechemer & Foerster 2015). Tuta absoluta adults received 10% (w/v) honey solution as food in moistened cotton placed in the plastic containers. Food and leaves for oviposition were replaced daily, when pre-oviposition, oviposition, fecundity, fertility, and longevity were evaluated.
Fertility life table
Fertility life tables for T. absoluta were constructed for the six cultivars. The life history data for T. absoluta were analyzed based on the age-stage, two-sex life table according to Chi & Liu (1985) and Chi (1988). The reproductive parameters (net reproductive rate (R 0), mean duration of one generation (T), intrinsic rate of increase (R m), and finite rate of increase (λ)) were calculated and compared using the TWOSEX-MSChart procedure (Chi 2009). The means, variances, and standard errors of the life table parameters were estimated using 10,000 bootstrap samples (Efron & Tibshirani 1993). The parameters were compared between treatments using paired bootstrap test.
Data analysis
All tests were conducted using a completely randomized design. The effects of tomato cultivars on the development and measurements of wing and tibia of T. absoluta were tested for homogeneity and normality with the Levene and Shapiro-Wilk tests, respectively. Only the datasets that did not conform to a normal distribution were transformed with the formulas log (x + 1) and (x + 0.5)1/2 and compared with an analysis of variance (ANOVA). Significant differences among treatments were compared with the Tukey test (p < 0.05). Mortality during larval and pupal stages and egg-to-adult development were analyzed with a chi-square test at 1% significance level. Survival curves were constructed with the Kaplan-Meier method (p < 0.05) (Kaplan & Meier 1958). All statistical analyses were performed using the software Statistica v. 7 (Statsoft 2004).
Results
Development and survival of Tuta absoluta on six tomato cultivars
Tuta absoluta completed its development on all tomato cultivars, but the genotype consumed during the larval stage affected the development period from egg to adult (F (5,379) = 13.89; p < 0.001). Insect development was shorter on the cultivar Santa Clara and longer on Cordilheira (28.2 days), which was similar to the values obtained for Nemoneta, Paron, and Santa Clara (Table 1). Egg incubation period was shorter on Cherry, Cordilheira, and Nemoneta (F (5,95) = 170.3; p < 0.001), and larval development was shortest on Cherry (14.9 days) (F (5,466) = 30.1; p < 0.001). The duration of the pupal stage duration was similar among treatments (F (5,375) = 2.01; p = 0.07) (Table 1).
Tomato cultivars also affected the size of T. absoluta adults. The tibia length of females fed the tomato cultivar Paron was longer than those fed Cordilheira (F (5,102) = 3.71; p < 0.001), but female wingspan was similar among treatments (F (5,107) = 0.46; p = 0.80) (Table 2). Tibia length of males was not significantly different (F (5,86) = 1.66; p = 1.53), but wingspan of insects raised on Santa Clara was longer than those fed Cherry, Cordilheira, and Giuliana (F (5,87) = 4.42; p < 0.001) (Table 2).
Survival during the life cycle of Tuta absoluta
Survival curves revealed the effect of tomato cultivars on T. absoluta (χ 2 (5) = 21.91; p < 0.001) (Fig 2). Larval mortality rate was lower than expected for insects fed the cultivar Cherry (Table 3), and it was 38.6, 31.4, and 10% when reared on Cordilheira, Giuliana, and Cherry, respectively. Pupal mortality was 5.7, 17.9, and 15% for insects reared on cultivars Cordilheira, Santa Clara, and Giuliana, respectively. Egg-adult mortality rate was lowest for insects reared on Cherry and highest on Giuliana (Table 3). Insects fed leaves of the latter that reached the adult stage had a longer lifespan of approximately 110 days, more than twice longer than those reared on Cordilheira, with less than 50 days (Fig 1).

Survival curves of the life cycle of males and females of Tuta absoluta (Lepidoptera: Gelechiidae) reared on six commercial tomato cultivars.
Reproduction and longevity of Tuta absoluta reared on six commercial tomato cultivars
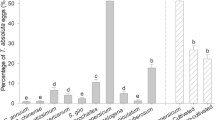
The tomato cultivars affected the pre-oviposition (F (5,94) = 4.08; p < 0.001), oviposition (F (5,94) = 4.09; p < 0.001), and fertility (F (5,94) = 9.66; p < 0.001) of T. absoluta. However, female fecundity (F (5,94) = 1.37; p = 0.24) was similar among treatments. The shortest pre-oviposition period was observed for insects fed the cultivar Cordilheira (1.3 days), being similar only to those fed Santa Clara (Table 4). Oviposition peaks were recorded until the third day for all cultivars (Fig 2). Females fed Nemoneta oviposited for 19.8 days, longer than that obtained for those reared on Cordilheira, with 11.3 days. The mean fecundity of T. absoluta ranged between 134.8 eggs for insects fed Santa Clara and 166.9 eggs for those reared on Paron, without differences among treatments (Table 4). The highest percentage of fertility was observed for eggs laid by females fed the Cordilheira, Cherry, and Nemoneta cultivars. The lowest fertility rates were observed for insects reared on Giuliana and Paron followed by Santa Clara (Table 4).

Mean number of eggs laid throughout the lifespan of Tuta absoluta (Lepidoptera: Gelechiidae) reared on six commercial tomato cultivars.
Longevity differed among T. absoluta females and males fed the six tomato cultivars (F (5,111) = 4.38, p < 0.001; F (5,111) = 2.18, p = 0.03). Females reared on Cherry, Giuliana, and Nemoneta had longer lifespans than those fed Cordilheira. Also, females had longer lifespans than males reared on Cherry, Cordilheira, and Nemoneta (F (1,34) = 4.97, p = 0.03; F (1,28) = 5.1, p = 0.03; F (1,30) = 6.05, p = 0.02, respectively). Longevity of females and males was similar for insects reared on Giuliana (F (1,42) = 0.61; p = 0.44), Paron (F (1,48) = 0.29; p = 0.59), and Santa Clara (F (1,40) = 1.24; p = 0.27).
Fertility table
The net reproductive rate (R 0) and finite rate of increase (λ) of T. absoluta were higher in insects reared on the cultivars Cherry and Nemoneta, but it differed only from Santa Clara. The longest generation time (T) was observed in insects reared on Santa Clara (40.3 days), while the shortest generation time was recorded on Cherry with 37 days. No differences were found between genotypes regarding the intrinsic rate of increase (R m) (Table 5).
Discussion
Currently, the control of T. absoluta is entirely conducted with broad-spectrum insecticides (Guedes & Siqueira 2012, Campos et al 2015). A high number of applications are required throughout the tomato growing season in Brazil (Guedes & Siqueira 2012), which have led to a steady increase in reports of insecticide resistance in T. absoluta. In South America, T. absoluta has been reported to be resistant to pyrethroids, organophosphates, abamectin, cartap, and chitin synthesis inhibitors (Siqueira et al 2000a, 2000b, Salazar & Araya 2001, Siqueira et al 2001, Lietti et al 2005, Silva et al 2011). Recent studies have also shown resistance of T. absoluta to insecticides containing spinosyns and diamide (Campos et al 2015, Roditakis et al 2013, 2015). In light of this context, alternative control methods to reduce insecticide applications need to be incorporated into the pest management of tomato crops. The first important aspect in pest management is the choice of a suitable tomato cultivar, which was examined in our study.
The development, survival, and reproduction of T. absoluta were affected by the tomato cultivars tested. Indices to assess the digestibility of plant nutrients and biomass conversion efficiency are difficult to measure in leafminers (Koricheva & Haukioja 1992). Thus, biological parameters of insects reared on different hosts are an important tool to compare the suitability of different hosts. A host that provides the conditions for a faster development time allows the occurrence of more generations of the pest in the field. Our results showed that the cultivar Cherry promoted the shortest development time for T. absoluta, indicating that population growth will be faster when larvae feed on this cultivar. On the other hand, the development of T. absoluta on Cordilheira was the longest among the evaluated cultivars. Mortality of immature stages was also higher, while the oviposition period was reduced as well as the longevity of adults. Longer development time and lower fecundity suggest that this host does not provide adequate nutrition for T. absoluta (Waters & Barfield 1989, Awmack & Leather 2002). Therefore, Cordilheira was the least susceptible cultivar to T. absoluta. Our findings regarding development time are in agreement with those reported previously on the biology of the tomato leafminer (Coelho & França 1987, Imenes et al 1990, Borgoni & Carvalho 2006), although some differences were observed. These studies do not mention the cultivars on which larvae were reared, but some of these variations may be associated to differences in temperature.
Wing and tibia length are parameters commonly used to measure the size of adult insects (Awmack & Leather 2002). Despite the differences found in the development, reproduction, and demographic parameters of T. absoluta reared on different cultivars, wing and tibia length of adults did not differ significantly. Thus, the latter are not suitable parameters to detect differences in the development of the tomato leafminer feeding on different hosts. Instead, pupal weight may be more appropriate to detect differences in the development.
The realized fecundity of T. absoluta females was not significantly different, but the fertility of insects reared on the cultivars Giuliana and Santa Clara was the lowest compared to those reared on the other genotypes. Fecundity is the total number of eggs produced by females, while fertility is the number of viable eggs produced. The potential fecundity is a measure of the reproductive capacity of an insect, defined by the number of eggs in the reproductive tract (Kazimirova 1996, Sequiera & Dixon 1996, Rossi et al 1999, Awmack & Leather 2002), whereas realized fecundity refers to the number of offspring produced during the insect’s life (Leather 1995). In many herbivorous species, especially among Lepidoptera, all eggs are present in the ovarioles before the adult molt (Leather 1994, Boggs & Ross 1993, Awmack & Leather 2002). Although these eggs may not be at the same stage of development, the quality of the host plant during larval development is the determining factor of the potential fecundity and fertility (Awmack & Leather 2002). The number of viable eggs obtained for insects reared on the cultivars Cherry, Cordilheira, and Nemoneta was similar to the higher values reported by Krechemer & Foerster (2015) at temperatures of 15 and 25°C, while the lowest fertility rates were observed for insects reared on the cultivars Giuliana, Paron, and Santa Clara. Nevertheless, the development time until adult stage differed only for insects reared on Cherry and Cordilheira. Thus, these hosts do not seem to strongly affect the development time of T. absoluta, but some negatively influence the viability of eggs, contributing to a lower population increase in the following generations. Moreover, our findings on the lifespan of males and females were similar to those of previous studies reporting that females live longer than males (Coelho & França 1987, Angel 1988, Haji et al 1988, Imenes et al 1990, Erdogan & Babaroglu 2014).
Demographic parameters of insects reared on different host plants under laboratory conditions are useful to assess their biotic potential when evaluating a new food source and the quality of the host plant (Sánchez & Pereyra 1995, Pereyra & Sánchez 2006). When R 0 is above 1 and R m positive, the population is growing on all cultivars (Carey 1993, Krebs 1994). In our study, highest intrinsic rate of increase (R m) and the second highest net reproductive rate (R 0) were observed for T. absoluta reared on the cultivar Cherry. These parameters are frequently used to measure species performance (Giske et al 1993), since they provide information on survival, development, and reproduction of a population (Chi & Yang 2003). The intrinsic rate of increase showed a stable age distribution, and this is considered an essential demographic parameter to predict and compare potential population growth (Andrewartha & Birch 1954, Varley & Gradwell 1970, Hulting et al 1990). Thus, the combined values of life table parameters indicate a high growth potential of the population of T. absoluta when reared on Cherry cultivar, since for each generation, the value of these parameters tend to increase. Furthermore, the mortality during the life cycle of T. absoluta reared on Cherry cultivar was lowest, while fertility rate was similar to those obtained for other cultivars. Therefore, these results indicate that Cherry is the most suitable cultivar for the development of T. absoluta and, consequently, the most susceptible.
The tomato cultivar Cherry, one of the most cultivated tomato genotype worldwide, is the most predisposed to the attack of T. absoluta, while Cordilheira, Giuliana, and Santa Clara were the least suitable ones for its development and reproduction. Therefore, these cultivars are the most recommended for commercial tomato crops to reduce the damage caused by the tomato leafminer.
References
Andrewartha HG, Birch LC (1954) The distribution and abundance of animals. University of Chicago Press, Chicago, p 793
Angel RV (1988) Reconicimento, identificacion y biologia de species de Gelechiidae (Lepidoptera) en plantas solanaceas del departamento de Antioquia: I. Scrobipalpula absoluta (Meyrick). Rev Col Entomol 14:25–32
Awmack CS, Leather SR (2002) Host plant quality and fecundity in herbivorous insects. Annu Rev Entomol 47:817–844
Boggs CL, Ross CL (1993) The effect of adult food limitation on life history traits in Speyera mormonia (Lepidoptera, Nymphalidae). Ecology 74:433–441
Borgoni PC, Carvalho GS (2006) Biologia de Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) em diferentes cultivares de Lycopersicon esculentum Mill. Boikos 20:49–61
Campos MR, Silva TBM, Silva WM, Silva JE, Siqueira HAA (2015) Spinosin resistance in the tomato borer Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). J Pest Sci 88:405–412
Carey JR (1993) Applied demography for biologists, with special emphasis on insects. Oxford University Press, New York, p 206
Chi H (1988) Life-table analysis incorporating both sexes and variable development rate among individuals. Environ Entomol 17:26–34
Chi H, Liu H (1985) Two new methods for the study of insect population ecology. Bull Inst Zool Academia Sinica 24:225–240
Chi H (2009) TWOSEX-MSChart: a computer program for the age-stage, two-sex life table analysis. http://140.120.197.173/Ecology/
Chi H, Yang TC (2003) Two-sex life table and predation rate of Propylaea japonica Thunberg (Coleoptera: Coccinellidae) fed on Myzus persicae (Sulzer) (Homoptera: Aphididae). Environ Entomol 32:327–333
Coelho MCF, França FH (1987) Biologia, quetotaxia da larva e descrição da pupa e adulto da traça-do-tomateiro. Pesq Agropec Bras 22:129–135
Colomo MV, Berta DC (1995) Fluctuación de la población de Scrobipalpula absoluta (Meyrick) (Lepidoptera, Gelechiidae) en plantaciones de tomate en el Departamento de Lules, Tucumán. Acta Zool Lilloana 43:165–177
Desneux N, Luna MG, Guillemaud T, Urbaneja A (2011) The invasive South American tomato pinworm, Tuta absoluta, continues to spread in Afro-Eurasia and beyond: the new threat to tomato world production. J Pest Sci 84:403–408
Desneux N, Wajnberg E, Wyckhuys AGK, Burgio G, Arpaia S, Narvaez-Vasquez CA, Gonzalez-Cabrera J, Catalan Ruescas D, Tabone E, Frandon J, Pizzol J, Poncet C, Urbaneja CA (2010) Biological invasion of European tomato crops by Tuta absoluta: ecology, geographic expansion and prospects for biological control. J Pest Sci 83:197–215
Ecole CC, Picanco MC, Guedes RNC, Brommonschenkel SH (2001) Effect of cropping season and possible compounds involved in the resistance of Lycopersicon hirsutum f. typicum to Tuta absoluta (Meyrick) (Lep., Gelechiidae). J Appl Entomol 125:193–200
Ecole CC, Picanco M, Moreira MD, Magalhaes STV (2000) Chemical components associated with resistance of Lycopersicon hirsutum f. typicum to Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). An Soc Entomol Bras 29:327–337
Efron B, Tibshirani RJ (1993) An introduction to the bootstrap. Chapman and Hall, New York, p 465
Erdogan P, Babaroglu NE (2014) Life table of the tomato leaf miner, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). JAFAG 31:80–89
Fernandez S, Montagne A (1990) Biología del minador del tomate, Scrobipalpula absoluta (Meyrick) (Lepidoptera: Gelechiidae). Bol Entomol Venez 5:89–99
Giske J, Aksnes DL, Forland B (1993) Variable generation times and Darwinian fitness measures. Evol Ecol 7:233–239
Guedes RNC, Siqueira HAA (2012) The tomato borer Tuta absoluta: insecticide resistance and control failure. CAB Rev Perspect Agric Vet Sci Nutr Nat Resour 7:1–7
Hagen KS, Dadd RH, Reese J (1984) The food of insects. In: Huffaker CB, Rabb RL (eds) Ecological methodology. J. Wiley & Sons, New York, pp 79–112
Haji FNP, Parra JRP, Silva JP, Batista JGS (1988) Biologia da traça-do-tomateiro sob condições de laboratório. Pesq Agropec Bras 23:107–110
Han P, Bearez P, Adamowicz S, Lavoir AV, Amiens-Desneux E, Desneux N (2015) Nitrogen and water limitation in tomato plants triggers negative bottom-up effects on the omnivorous predator Macrolophus pygmaeus. J Pest Sci 88:685–691
Han P, Lavoir AV, Le Bot J, Amiens-Desneux E, Desneux N (2014) Nitrogen and water availability to tomato plants triggers bottom-up effects on the leafminer Tuta absoluta. Sci Rep 4:4455
Hulting FL, Orr DB, Obrycki JJ (1990) A computer program for calculation and statistical comparison of intrinsic rates of increase and associated life table parameters. Fla Entomol 73:601–612
Imenes SDL, Uchôa-Fernandes MA, Campos TB, Takematsu AP (1990) Aspectos biológicos e comportamentais da traça-do-tomateiro Scrobipalpula absoluta (Meyrick, 1917), (Lepidoptera-Gelechiidae). Arq Inst Biol 57:63–68
Jaenike J (1978) On optimal oviposition behavior in phytophagous insects. Theor Popul Biol 14:350–356
Kaplan EL, Meier P (1958) Nonparametric estimation from incomplete observations. J Amer Statist Assoc 53:457–481
Kazimirova M (1996) Influence of larval crowding and mating on lifespan and fecundity of Mamestra brassicae (Lepidoptera: Noctuidae). Eur J Entomol 93:45–52
Kiliç T (2010) First record of Tuta absoluta in Turkey. Phytoparasitica 38:243–244
Krebs CJ (1994) Ecology: the experimental analysis of distribution and abundance. Harper Collins, New York
Koricheva J, Haukioja E (1992) Effects of air pollution on host plant quality, individual performance and population density of Eriocrania miners (Kepidoptera: Eriocraniidae). Environ Entomol 21:1386–1392
Krechemer FS, Foerster LA (2015) Tuta absoluta (Lepidoptera: Gelechiidae): thermal requirements and effect of temperature on development, survival, reproduction and longevity. Eur J Entomol 112:658–663
Larbat R, Adamowicz S, Robin C, Han P, Desneux N, Le Bot J (2016) Interrelated responses of tomato plants and the leafminer Tuta absoluta to nitrogen supply. Plant Biol 18:495–504
Leather SR (1994) Life history traits of insect herbivores in relation to host quality, p. 175–207. In: Bernays EA (ed) Insect-plant interactions, v 5. CRC Press, Boca Raton, p 305
Leather SR (1995) Factors affecting fecundity, fertility, oviposition and larviposition in insects. In: Leather SR, Hardie RJ (eds) Insect reproduction. CRC Press, Boca Raton, FL, pp 143–174
Lietti MMM, Botto E, Alzogaray RA (2005) Insecticide resistance in Argentine populations of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Neotrop Entomol 34:113–119
Maluf WR, Barbosa LV, Santa-Cecilia LVC (1997) 2-Tridecanone-mediated mechanisms of resistance to the South American tomato pinworm Scrobipalpuloides absoluta (Meyrick, 1917) (Lepidoptera-Gelechiidae) in Lycopersicon spp. Euphytica 93:189–194
Medeiros MA, Vilela NJ, Franca FH (2006) Technical and economic efficiency of biological control of the South American tomato pinworm in protected environment. Hort Bras 24:180–184
Pereyra PC, Sánchez NE (2006) Effect of two solanaceous plants of development and population parameters of the tomato leaf miner, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Neotrop Entomol 35:672–675
Pizzamiglio JRP (1991) Ecologia das interações entre inseto/planta. In: Panizzi AR, Parra JRP (eds) Ecologia nutricional de insetos e suas implicações no manejo de pragas. São Paulo, Manole, pp 101–129 412 p
Roditakis E, Vasakis E, Grispou M, Stavrakaki M, Nauen R, Gravouil M, Bassi A (2015) First report of Tuta absoluta resistance to diamide inseticides. J Pest Sci 88:9–16
Roditakis E, Skarmoutsou C, Stavrakaki M, del Rosario M-AM, García-Vidal L, Bielza P, Haddi K, Rapisarda C, Rison J-L, Bassi A, Teixeira LA (2013) Determination of baseline susceptibility of European populations of Tuta absoluta (Meyrick) to indoxacarb and chlorantraniliprole using a novel dip bioassay method. Pest Manag Sci 69:217–227
Rossi AM, Stiling P, Cattell MV, Bowdish TI (1999) Evidence for host-associated races in a gall-forming midge: trade-offs in potential fecundity. Ecol Entomol 24:95–102
Salazar ER, Araya JE (2001) Respuesta de la polilla del tomate, Tuta absoluta (Meyrick), a insecticidas en Arica. Agric Tech 61:429–435
Sánchez NE, Pereyra PC (1995) Life table of the soybean looper Rachiplusia nu (Lepidoptera: Noctuidae) in the laboratory. Rev Soc Entomol Argent 54:89–96
Sauvion S, Mauriello V, Renard B, Boissot N (2005) Impact of melon accessions resistant to aphids on the demographic potential of silverleaf whitefly. J Econ Entomol 52:557–567
Sequiera R, Dixon AFG (1996) Life history responses to host quality changes and competition in the Turkey-oak aphid. Eur J Entomol 93:53–58
Silva GA, Picanço MC, Bacci L, Crespo ALB, Rosado JF, Guedes RNC (2011) Control failure likelihood and spatial dependence of insecticide resistance in the tomato pinworm, Tuta absoluta. Pest Manag Sci 67:913–920
Siqueira HAA, Guedes RNC, Fragoso DB, Magalhaes LC (2001) Abamectin resistance and synergism in Brazilian populations of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Int J Pest Manag 47:247–251
Siqueira HAA, Guedes RNC, Picanço MC (2000a) Cartap resistance and synergism in populations of Tuta absoluta (Lep., Gelechiidae). J Appl Entomol 124:233–238
Siqueira HAA, Guedes RNC, Picançoo MC (2000b) Insecticide resistance in populations of Tuta absoluta (Lepidoptera: Gelechiidae). Agric Forest Entomol 2:147–153
Statsoft, Inc. (2004) Software Statistica 7.0, Tulsa, OK, USA, Statsoft Inc.
Torres JB, Faria CA, Evangelista Junior WS, Pratissoli D (2001) Within-plant distribution of the leaf miner Tuta absoluta (Meyrick) immatures in processing tomatoes, with notes on plant phenology. Int J Pest Manage 47:173–178
Uchôa-Fernandes MA, Della Lucia TMC, Vilela EF (1995) Mating, oviposition and pupation of Scrobipalpuloides absoluta (Meyr.) (Lepidoptera: Gelechiidae). An Soc Entomol Bras 24:159–164
Varley GC, Gradwell GR (1970) Recent advance in insect population dynamics. Annu Rev Entomol 15:1–24
Viggiani G, Filella F, Delrio G, Ramassini W, Foxi C (2009) Tuta absoluta, a new Lepidoptera now reported in Italy. Informatore Agrario 65:66–68
Waters DG, Barfield CS (1989) Larval development and consumption by Anticarsia gemmatalis (Lepidoptera: Noctuidae) fed various legume species. Environ Entomol 18:1006–1010
Acknowledgments
The first author thanks the National Council for Scientific and Technological Development (CNPq) for scholarships.
Author information
Authors and Affiliations
Corresponding author
Additional information
Edited by Leandro P Ribeiro – EPAGRI
Rights and permissions
About this article

Cite this article
Krechemer, F.S., Foerster, L.A. Development, Reproduction, Survival, and Demographic Patterns of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) on Different Commercial Tomato Cultivars. Neotrop Entomol 46, 694–700 (2017). https://doi.org/10.1007/s13744-017-0511-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13744-017-0511-5