
Abstract
Beneficial symbionts residing in our gut have positive therapeutic effects on several metabolic disorders including diabetes. Oral administration of probiotic and prebiotic foods strengthens the beneficial symbiont populations in the gut and may prevent immune-mediated destruction of pancreatic β-cells. The present study was designed to elucidate the gut microbiome of diabetic rats supplemented with a Lactobacillus probiotic and a Saccharomyces cerevisiae (SC) cell wall prebiotic. Diabetes mellitus was induced in male Wistar rats with allaxon monohydrate (150 mg/kg). The rats were fed chow maintenance diet (control and diabetic control groups) or the same diet supplemented with a SC prebiotic (1 %), probiotic (multispecies Lactobacillus @108 CFU), or synbiotic. On d30, DNA was extracted from colon digesta for 16S ribosomal RNA (rRNA) gene sequencing. Serum was obtained to estimate total oxidant and anti-oxidant concentrations. A distinct clustering pattern (Unifrac distances, analysis of similarities (ANOSIM) P = 0.0361) was observed for the different treatment groups, with the main distinction consisting of the separation between the control and the diabetic control groups. Distinct bacterial clades dominated different treatment groups, particularly for the control and the diabetic control groups, though several bacterial groups overlapped, demonstrating a core microbiota dominated mainly by Firmicutes and Bacteroides. A trend of dysbiosis, characterized by low species richness, was observed in the diabetic rats, albeit not statistically significant. Serum oxidant and anti-oxidant concentrations were not different (P > 0.05) among different treatment groups. No significant effects of supplementations of prebiotic, probiotic, and synbiotic were observed on species richness or clustering pattern of the microbiome.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Type 1 diabetes mellitus (T1DM) is a metabolic disorder that results from a myriad of factors. The disease is usually characterized by oxidative and/or T cell-mediated autoimmune destruction of pancreatic beta cells, leading to partial or complete loss of insulin production [1, 2]. Although limited information is available about the pathogenesis of the disorder, it is generally believed that in genetically susceptible individuals, a chronic inflammatory disease of the gut triggers the primary insult, leading to redox destruction of β-islet cells [3–5]. Furthermore, recent studies have suggested that the risk of development of diabetes is also triggered by the gastrointestinal tract (GIT) microbiome [6] and that the disease is associated with microbial dysbiosis in the GIT [7].
The GIT microbiota is considered important for host health, nutrition, and immunity and can be affected by age, stressors, disease, and diet [8]. Several microbes living in the gut are known for their beneficial effects (e.g., anti-allergic, anti-carcinogenic, anti-diabetic, and cholesterol lowering) on host health [3, 9]. Microbes living in the intestine produce numerous unknown metabolites that are absorbed from the gut and influence host health and immunity and alter the gut–brain barrier [10].
Consumption of fermented foods augments gut microbiome and delays the progression of streptozotocin-induced diabetes in a rat model [11]. These functional foods can lower plasma glucose and delay the progression of experimentally induced diabetes in rats [11–13]. Recently, Park, Ahn [14] has elucidated that oral administration of probiotic Lactobacillus spp. suppressed insulin resistance, reduced glucose and cholesterol concentrations, reduced reactive oxygen species, and decreased blood pressure. Probiotic Lactobacillus can prevent onset of insulin-dependent diabetes mellitus in mice by enhancing the number of beneficial symbionts in the host gut [15]. Also, probiotics are known for reducing predisposing factors for diabetes like obesity, allergies, and autoimmune disorders. Similarly, prebiotics such as xylo-oligosaccharides and fructo-oligosaccharides have also been reported to ameliorate the metabolic abnormalities associated with diabetes such as hyperglycemia, hypercholesterolemia, glucosuria, proteinuria, and diabetic nephropathy [16]. Particularly, supplementation with the prebiotic cell wall extract from Saccharomyces cerevisiae for 12 weeks has been shown to decrease blood pressure and improve glycemic indices in patients with T2D [17, 18].
Although some studies [19, 20] have reported that microbial dysbiosis is a contributing factor in the disease pathogenesis, limited literature is available investigating the gut microbiome of diabetic rats fed probiotics and/or prebiotics. Therefore, the present study was designed to study the effects of a prebiotic cell wall extract from S. cerevisiae and probiotic Lactobacillus spp. on the gut microbiome of diabetic rats using 16S ribosomal RNA (rRNA) gene sequencing.
Materials and methods
Animals, diets, and study design
Thirty adult male Wistar rats of the same weight (200 ± 20 g) were divided into five treatment groups. The rats were housed under standard management conditions (two rats/cage, 24 ± 2 °C, 12-h light/12-h dark cycle) with free access to food and water. After 1-week acclimatization period, rats were injected intraperitoneally with 150 mg/kg allaxon monohydrate (Sigma-Aldrich, UK) as a 5 % solution in normal saline (diabetic groups) or the same volume of normal saline (control group). Post injection, 5 % glucose solution was provided for 48 h to prevent initial drug-induced hypoglycemic mortality. Ninety-six hours post injection, blood (Aviva Accu-Chek, Roche Diagnostics) and urine (Benedict’s qualitative test) glucose was tested to confirm diabetes in rats injected with allaxon. Only rats with blood glucose level above 180 mg/dL were used in the analysis. The rest of the animals which did not demonstrate the inclusion criteria of hyperglycemia were excluded.
Rats were divided into the following groups: control group fed chow maintenance diet (CMD), diabetic control group fed CMD, diabetic prebiotic group fed CMD supplemented with 1 % S. cerevisiae yeast cell wall extract, diabetic probiotic group fed CMD supplemented with a multispecies probiotic of Lactobacillus spp. (˜108 CFU), and diabetic synbiotic group fed CMD supplemented with a combination of both 1 % prebiotic and 108 CFU of the probiotic Lactobacillus spp. The CMD was composed of 54 % corn starch, 21 % casein, 10 % refined soybean oil, 10 % cane sugar, and 5 % vitamin–mineral premix (National Research Council). Dietary treatments were continued from day 0 to day 30, and blood glucose levels were measured on a weekly basis. At the end of the study period, rats were killed by decapitation, and trunk blood and colon digesta were collected.
Serum oxidant and anti-oxidant analysis
Blood was centrifuged at 1500×g at 4 °C for 15 min for serum extraction. Serum total oxidant concentrations (TOC; μm of H2O2 equivalent/L) were measured using a colorimetric method based on the oxidation of ferrous ion to ferric ion in the presence of various oxidant species [21]. The procedure was calibrated with hydrogen peroxide. Total anti-oxidant concentrations (TAC; mM Eq. of vitamin C/L) were measured using a novel automated colorimetric method using odianisidine dihydrochloride as the substrate as described by Erel [22]. The data obtained for serum TOC and TAC were analyzed using ANOVA to evaluate treatment effects.
Microbiome analysis
Colon digesta was subjected to DNA extraction using the BiOstic® FFPE Tissue DNA Isolation Kit (MoBio Laboratories, Carlsbad, CA), following the manufacturer’s protocol. The V4 region of the 16S rRNA gene was amplified with primers 515F (5′-GTGCCAGCMGCCGCGGTAA-3′) and 806R (5′-GGACTACVSGGGTATCTAAT-3′) at the MR DNA Laboratory (Shallowater, TX, USA). PCR amplification products were verified on 2 % agarose gels, and samples were purified using calibrated Ampure XP beads. The Illumina TruSeq DNA Library was used to prepare a DNA library and sequenced at MR DNA on an Illumina MiSeq instrument, as described earlier [23]. Raw sequence data were screened, trimmed, denoised, filtered, chimera-depleted, and clustered as operational taxonomic units (OTUs) at 97 % similarity, using QIIME (V1.7) default settings. The sequences obtained in this study were submitted to NCBI Short Read Archive as FASTQ files with accession number SRR1613115.
A total of 1,200,168 sequences were recovered from all samples. To standardize sequence depth, further data analysis was performed on an even sample depth of 57,546. Alpha rarefaction (species richness per sample) and beta diversity (microbiome similarity between samples) were measured and plotted using QIIME. The analysis of similarities (ANOSIM; P = 0.05) was performed on unweighted UniFrac distances to compare the microbiome among the different treatment groups. Relative clustering pattern of the microbiome among different treatment groups was analyzed using Pearson’s correlation test, and a dendrogram was constructed using default settings in METAGENassist [24]. The Kolmogorov–Smirnov test was used to evaluate the normal distribution of the data. Further, non-parametric Kruskal–Wallis H test was also applied on the data and resulting p values were adjusted for multiple comparisons using Benjamini and Hochberg’s false discovery test.
Results
Cecal microbiome
Sequencing of 16S rRNA genes was performed to explore the phylogenetic composition of the microbiome. Approximately, 1,200,168 (minimum 57,546, maximum 153,060, median 71,981, and standard deviation 29,044) chimera-depleted good-quality 16S rRNA gene sequences were retrieved from all the samples. These sequences corresponded to 23 phyla, 39 classes, 70 orders, 138 families, and 291 genera. Regardless of the high bacterial diversity, only three phyla (Firmicutes, Bacteroidetes, and Proteobacteria) accounted for more than 99 % of all the obtained sequences (Fig. 1).

Composition of colon microbiome of the control, diabetic control, diabetic probiotic, diabetic prebiotic, and diabetic synbiotic rats at the phylum level. The bars represent median percentage of sequences
A cluster hierarchy dendrogram based on the Pearson correlation test using default parameters was constructed in Fig. 2. The vertical axis of the dendrogram represents the dissimilarity between clusters. The horizontal axis represents the samples. The dendrogram revealed that the control and probiotic-supplemented diabetic groups had significantly different (dissimilarity ≥1.0) communities compared to the other treatment groups.

Pearson correlation hierarchical clustering dendrogram of 16S rRNA-based sequences
Principal coordinate analysis (PCoA) plots for unweighted UniFrac distances were constructed to evaluate microbiome variation between different treatment groups (Fig. 3). We applied the statistical analysis ANOSIM on the unweighted UniFrac distances and observed a significant clustering pattern in the PCoA plots (ANOSIM with 43,259 permutation, P = 0.0361), demonstrating significant differences in microbiomes of different treatment groups. Pairwise ANOSIM analysis revealed that only the control and the diabetic control groups had significantly different (P = 0.031) clustering pattern. All the other treatment groups had non-significantly different (P > 0.05) clustering pattern.

Alpha diversity measures at 57,546 sequences per sample in the different treatment groups. Bacterial diversity and richness index graphs (Chao 1 (a), observed species (b), and Shannon–Weaver (c)) obtained from colon microbiome samples. Control (red), diabetic control (green), diabetic probiotic (blue), diabetic prebiotic (purple), and diabetic synbiotic (orange) (Color figure online)
The alpha rarefaction analysis (Chao 1, observed species, and Shannon), at an even sample depth of 57,546 sequences per sample, revealed high inter-individual variability in rats from the different treatment groups (Fig. 4). Species richness was highest (P > 0.05) in the control group and lowest (P > 0.05) in the diabetic control group. However, alpha rarefaction indices were non-significantly different among the different treatment groups.

Principal coordinate analysis (PCoA) plots of the unweighted Unifrac distance matrix. The plots show each combination of the first three principal coordinates. Control (red, square), diabetic control (red, triangular), diabetic prebiotic (blue, triangular), diabetic probiotic (aqua, circular), and diabetic synbiotic (green, triangular) (Color figure online)
We further tested our finds by subjecting our data to Kruskal–Wallis one-way analysis of variance and false discovery tests. These tests reveal no significant differences in bacterial taxa on various phylogenetic levels among different treatment groups (Table S1).
Serum biochemistry
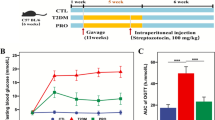
We measured serum glucose concentrations on day 30 of the study (Fig. S1). Control group had relatively low (P > 0.05) serum glucose concentrations compared with the diabetic rats. The serum oxidant concentrations and anti-oxidant activities were also determined at the termination of the experiment. Perhaps, no significant differences were observed among the different treatment groups (Fig. S2).
Discussion
Type 1 diabetes is an idiopathic syndrome characterized by destruction of insulin-producing beta cells of the pancreas. Though the exact link is not well established, it is thought that microbial dysbiosis in the gut can aggravate the immune system, thereby bolstering disease pathogenesis [6]. Therefore, present research work is focused on understanding gut microbiome characteristics and its potential role in host health and disease. Similarly, several studies are being conducted to explore the beneficial effects of probiotics and prebiotics for the management of diabetes [11–13]. To the best of our knowledge, only few studies [19, 20] have been conducted so far to investigate the dynamics of the gut microbiome in diabetic rats, supplemented with prebiotics and/or probiotics.
Phylogenetic data presented here were analyzed simultaneously at various taxonomic levels using QIIME 1.7 to classify the microbiomes of control and diabetic rats. Three phyla (Firmicutes, Bacteroidetes, and Proteobacteria) dominated the gut microbiome of all rats, and no significant differences were observed among different treatment groups at the phylum level. Similarly, no significant differences were observed at class- and order-level phylogeny. These findings are in agreement with the earlier work of Qin, Li [7], who found no significant bacterial dysbiosis in diabetic subjects compared to controls. However, our findings differ from some earlier reports as well. Giongo, Gano [20] found a decrease in Firmicutes and increase in Bacteroidetes in murine T1D model. Similarly, a microbial dysbiosis, characterized by a decline in relative abundance of Firmicutes and an increase in proportions of Bacteroidetes and Proteobacteria, was reported by [25].
In the present study, higher species richness and diversity, as indicated by alpha rarefaction (Fig. 3), were observed in the control group compared to the other treatment groups. We also observed that the first two coordinates of PCoA plots explained 22.76 % variation in the microbiome, suggesting that independent variables (diabetes and supplementations) were responsible for the observed variation between the samples [26]. A significant difference (pairwise ANOSIM P = 0.031) in clustering pattern revealed that only diabetes had an influence on the gut microbiome of rats. These findings are in agreement with the previously published Finnish children autoimmune T1D work [20]. In a leptin-resistant obesity model of mice diabetes, Everard, Lazarevic [27] demonstrated that modulation of gut microbiome by prebiotic diet can improve glucose and lipid homeostasis, leptin sensitivity, and activity of targeted enteroendocrine cells in diabetes. The study also showed a positive correlation between prebiotic-induced modulation of gut microbiome and these metabolic parameters, particularly the overrepresentation of Firmicutes in prebiotic-fed groups.
It is well known through earlier experiments and clinical studies that oxidative stress plays a major role in the pathogenesis of diabetes [28]. Oral administration of yogurt fortified with Lactobacillus spp. has been shown to suppress streptozotocin-induced oxidative damage [11] and improve anti-oxidant status and contribute to better management of T2D [12]. Furthermore, some earlier studies have also reported that the supplementation with S. cerevisiae cell wall improved gut microbial diversity [29] and increased the numbers of Bifidobacterium spp., Faecalibacterium spp., and Ruminococcus spp. in the colon [30]. These microbes generally augment production of anti-inflammatory and immune modulatory factors [31]. Further, probiotic and prebiotic supplements route multiple mechanism, which are not yet fully explored, to augment diabetes-associated suffering. Although previous studies have reported that these supplements can reduce plasma triglyceride levels, muscle lipid infiltration, adipose tissue mass, and oxidative stress [27], antithetical to the previous studies, we could not find significant effects of supplementation with either prebiotic, probiotic, or synbiotic on the gut microbiome of diabetic rats. In addition, serum TOC and TAC concentrations were also found unaltered. Though the exact reason for differences in findings is not known, perhaps, it may be proposed that different disease model, dosage (prebiotic 5 vs 100 g/kg, probiotic ~108 vs 738 CFU/g), duration of supplementations (30 days vs 6 week), microbiome analysis procedure (16S rRNA sequencing vs culture plate), and the number of animals used in the study have attributed to these controversies.
Taken together, these findings suggest a partial role of the gut microbiome in T1D. Though the sample size used here is too low to make a firm conclusion, these data suggest that T1D is associated with decreased diversity of the gut microbiome when compared with non-diabetic subjects. Although review of literature depicts positive role of prebiotic/probiotic supplements on diabetes-associated parameters, perhaps, we failed to modulate oxidative stress in the supplemented groups. Further investigations with higher dosages and longer durations of supplementations are necessary to ascertain better understanding of therapeutic effects of these functional foods. The present study can serve as springboard for future studies to delineate the pathophysiological role of the gut microbiome in diabetes.
References
Arrick DM, Mayhan WG. Cerebrovascular disease in type 1 diabetes: role of oxidative stress. studies in diabetes: Springer; 2014. p. 13-36.
Wen L, Ley RE, Volchkov PY, Stranges PB, Avanesyan L, Stonebraker AC, et al. Innate immunity and intestinal microbiota in the development of type 1 diabetes. Nature. 2008;455:1109–13.
Abbas Z, Jafri W. Yoghurt (dahi): a probiotic and therapeutic view. J Pak Med Assoc. 1992;42:221–4.
Bourlioux P, Koletzko B, Guarner F, Braesco V. The intestine and its microflora are partners for the protection of the host: report on the Danone Symposium “The Intelligent Intestine,” held in Paris, June 14, 2002. Am J Clin Nutr. 2003;78:675–83.
Watson JD. Type 2 diabetes as a redox disease. Lancet. 2014;383:841–3.
Musso G, Gambino R, Cassader M. Interactions between gut microbiota and host metabolism predisposing to obesity and diabetes. Annu Rev Med. 2011;62:361–80.
Qin J, Li Y, Cai Z, Li S, Zhu J, Zhang F, et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature. 2012;490:55–60.
Delzenne NM, Neyrinck AM, Bäckhed F, Cani PD. Targeting gut microbiota in obesity: effects of prebiotics and probiotics. Nat Rev Endocrinol. 2011;7:639–46.
Ghosh S, van Heel D, Playford RJ. Probiotics in inflammatory bowel disease: is it all gut flora modulation? Gut. 2004;53:620–2.
Schultz M, Linde HJ, Lehn N, Zimmermann K, Grossmann J, Falk W, et al. Immunomodulatory consequences of oral administration of Lactobacillus rhamnosus strain GG in healthy volunteers. J Dairy Res. 2003;70:165–73.
Yadav H, Jain S, Sinha PR. Oral administration of dahi containing probiotic Lactobacillus acidophilus and Lactobacillus casei delayed the progression of streptozotocin-induced diabetes in rats. J Dairy Res. 2008;75:189–95.
Ejtahed HS, Mohtadi-Nia J, Homayouni-Rad A, Niafar M, Asghari-Jafarabadi M, Mofid V. Probiotic yogurt improves antioxidant status in type 2 diabetic patients. Nutrition. 2012;28:539–43.
Roberfroid MB. Prebiotics and probiotics: are they functional foods? Am J Clin Nutr. 2000;71:1682s–7s.
Park DY, Ahn YT, Huh CS, McGregor RA, Choi MS. Dual probiotic strains suppress high fructose-induced metabolic syndrome. World J Gastroenterol. 2013;19:274–83.
Matsuzaki T, Nagata Y, Kado S, Uchida K, Kato I, Hashimoto S, et al. Prevention of onset in an insulin-dependent diabetes mellitus model, NOD mice, by oral feeding of Lactobacillus casei. APMIS. 1997;105:643–9.
Gobinath D, Madhu AN, Prashant G, Srinivasan K, Prapulla SG. Beneficial effect of xylo-oligosaccharides and fructo-oligosaccharides in streptozotocin-induced diabetic rats. Br J Nutr. 2010;104:40–7.
Hosseinzadeh P, Djazayery A, Mostafavi S-A, Javanbakht MH, Derakhshanian H, Rahimiforoushani A, et al. Brewer’s yeast improves blood pressure in type 2 diabetes mellitus. Iran J Public Health. 2013;42:602.
Hosseinzadeh P, Javanbakht MH, Mostafavi S-A, Djalali M, Derakhshanian H, Hajianfar H, et al. Brewer’s yeast improves glycemic indices in type 2 diabetes mellitus. Int J Prev Med. 2013;4:1131.
de Goffau MC, Fuentes S, van den Bogert B, Honkanen H, de Vos WM, Welling GW, et al. Aberrant gut microbiota composition at the onset of type 1 diabetes in young children. Diabetologia. 2014;57:1569–77.
Giongo A, Gano KA, Crabb DB, Mukherjee N, Novelo LL, Casella G, et al. Toward defining the autoimmune microbiome for type 1 diabetes. ISME J. 2010;5:82–91.
Erel O. A new automated colorimetric method for measuring total oxidant status. Clin Biochem. 2005;38:1103–11.
Erel O. A novel automated direct measurement method for total antioxidant capacity using a new generation, more stable ABTS radical cation. Clin Biochem. 2004;37:277–85.
Klindworth A, Pruesse E, Schweer T, Peplies J, Quast C, Horn M, et al. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2012:gks808.
Arndt D, Xia J, Liu Y, Zhou Y, Guo AC, Cruz JA, et al. METAGENassist: a comprehensive web server for comparative metagenomics. Nucleic Acids Res. 2012;40:W88–95.
Larsen N, Vogensen FK, van den Berg FW, Nielsen DS, Andreasen AS, Pedersen BK, et al. Gut microbiota in human adults with type 2 diabetes differs from non-diabetic adults. PLoS One. 2010;5:e9085.
Lozupone C, Knight R. UniFrac: a new phylogenetic method for comparing microbial communities. Appl Environ Microbiol. 2005;71:8228–35.
Everard A, Lazarevic V, Derrien M, Girard M, Muccioli GG, Neyrinck AM, et al. Responses of gut microbiota and glucose and lipid metabolism to prebiotics in genetic obese and diet-induced leptin-resistant mice. Diabetes. 2011;60:2775–86.
Maritim A, Sanders R, Watkins JB. Diabetes, oxidative stress, and antioxidants: a review. J Biochem Mol Toxicol. 2003;17:24–38.
Pourabedin M, Xu Z, Baurhoo B, Chevaux E, Zhao X. Effects of mannan oligosaccharide and virginiamycin on the cecal microbial community and intestinal morphology of chickens raised under suboptimal conditions. Can J Microbiol. 2014;60:255–66.
Upadrasta A, O’Sullivan L, O’Sullivan O, Sexton N, Lawlor PG, Hill C, et al. The effect of dietary supplementation with spent cider yeast on the swine distal gut microbiome. PLoS One. 2013;8:e75714.
Peng J, Narasimhan S, Marchesi JR, Benson A, Wong FS, Wen L. Long term effect of gut microbiota transfer on diabetes development. J Autoimmun. 2014.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The present study was partially sponsored by the American Association for the Advancement of Science under AAAS–CSTSP International Activity for BMENA Scientific Exchange Program. MU Sohail visited Texas A&M University under AAAS-BMENA Scientific Exchange Program. Other than this, authors declare no conflict of interest.
All procedures and protocols were adopted under the guidelines of the Animal Care and Ethics Committee, Offices of Research, Innovation, and Commercialization, GC University Faisalabad, Pakistan.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Table S1
Differences in bacterial taxa on various phylogenetic levels among different treatment groups. (DOCX 32 kb)
Figure S1
Serum glucose concentrations (mg/dL) on d 30 of the study. (TIFF 71 kb)
Figure S2
Serum total oxidant (μm of H2O2 equivalent/L) concentrations and total anti-oxidant (mM Eq. of vitamin C/L) concentrations. (TIF 40 kb)
Rights and permissions
About this article

Cite this article
Sohail, M.U., Shabbir, M.Z., Steiner, J.M. et al. Molecular analysis of the gut microbiome of diabetic rats supplemented with prebiotic, probiotic, and synbiotic foods. Int J Diabetes Dev Ctries 37, 419–425 (2017). https://doi.org/10.1007/s13410-016-0502-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13410-016-0502-9