
Abstract
Understanding the food habits of mammalian carnivores is crucial for the comprehension of the role of apex-predators in maintaining healthy ecosystems. The puma is currently the most widespread top predator and the carnivore most frequently involved in conflicts with humans in the Americas. We analyzed puma diet in the South American temperate region, a vast area largely modified by humans, to assess the importance of native prey with respect to livestock and exotic species. We reviewed 18 studies published between 1991 and 2020 to which we added 19 feces of puma from a rangeland area of central Argentina. “Undetermined small rodent,” plains vizcacha, European hare, and wild boar were the most frequent species in the small area of the Argentinean Espinal. In the southern temperate region, exotic (European hare and wild boar) and native species (guanaco and armadillos) were the most frequent wild prey species, while sheep was the most frequent livestock followed by cattle. Exotic species had a greater frequency of occurrence than native and livestock species. Livestock frequency of occurrence was greater inside protected areas than outside. These findings reveal that, although the puma is considered in this large area as conflictive, wild prey (exotic and native) are its main food sources. Given the potentially crucial role exotic species can play in the ecosystem, understanding the role of pumas in controlling their populations is a challenge for future research.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
After centuries of persecution (Stolzenburg 2008), large carnivores are increasingly recovering their social reputation thanks to the recognition of their major role in maintaining ecosystem processes and biodiversity (Terborgh et al. 1999; Ray et al. 2005). As apex predators, large carnivores have important impacts on densities and behavior of both prey and mesopredator species, generating top-down effects across ecosystem trophic webs (Terborgh and Estes 2013; Ripple et al. 2014). However, in human-modified landscapes, carnivore persecution and alteration on prey abundances (du Toit et al. 2017) can drastically affect the potential of apex predators in regulating ecosystem dynamics (Newsome and Ripple 2015; Kuijper et al. 2016). Mostly due to the global habitat loss and fragmentation, as well as the human conflict provoked by livestock predation, large carnivores are among the world’s most threatened species (Treves and Karanth 2003; Ripple et al. 2014). Describing large carnivores’ food habits contributes to understand their ecological importance in preserving ecosystem health, and simultaneously provides valuable information on the status of wild prey populations. This is particularly relevant in rangelands which are the most extensive land-use globally (Lund 2007; Briske 2017).
The puma Puma concolor is one of the most widely distributed and highly adaptable large carnivores in the Americas (Elbroch and Quigley 2019). Although puma diet predominantly relies on large prey (70 to 165 kg; Carbone et al. 1999), this felid feeds on diverse terrestrial and semiterrestrial vertebrates (Ruth and Murphy 2009). Because of predation on livestock, the puma is broadly considered a “conflictive” species (Inskip and Zimmermann 2009; Murphy and Macdonald 2010), even if less than 10% of feces composition is typically attributable to domestic prey (Ruth and Murphy 2009).
Puma food habits have been largely studied across its geographic distribution (e.g., Rau and Jiménez 2002; Novack et al. 2005; Elbroch and Wittmer 2013; Gómez-Ortiz et al. 2015). North American studies reported diets mainly based on large prey, particularly cervids, such as elk Cervus elaphus (Husseman et al. 2003), mule deer Odocoileus hemionus (Smith et al. 2016), and white-tailed deer Odocoileus virginianus (Cassaigne et al. 2016). In Central and South America, where large prey is less common, puma diet comprises a greater variety of species, spanning from large (guanaco Lama guanicoe, Pia 2013; Vicugna Vicugna vicugna, Pacheco et al. 2004, collared peccary Tayassu tajaco, Rueda et al. 2013; common rhea Rhea americana, Pessino et al. 2001) to medium and small prey (e.g., Pudu Pudu puda, Rau et al. 1991; nine-banded armadillo Dasypus novemcinctus, Gómez-Ortiz et al. 2015). Because of their generalistic feeding behavior, pumas also readily prey on exotic species, including the European hare Lepus europaeus, wild boar Sus scrofa, and European rabbit Oryctolagus cuniculus (Buenavista and Palomares 2018).
Temperate zones are the most extensively human-altered regions in the world: in these areas, many species have been entirely extirpated, while others are constrained to fragmented and highly modified remnants of natural ecosystems (Baldi et al. 2006; Wilcove et al. 1986). The distribution range of P. concolor covers, approximately, 1,837,630.675 km2 of the South American temperate region (from latitudes greater than 35° S). In this region, the nearly continuous puma’s distribution (with the exception of Tierra del Fuego province; Nielsen et al. 2015) overlaps with scarcely preserved landscapes. Only 13.1% of this large territory is under legal protection, where poorly connected protected areas are concentrated, mainly, along the Andean range. Large native ungulates (guanaco, Pampas deer Ozotoceros bezoarticus, Patagonian huemul Hippocamelus bisulcus), in this region, have been heavily hunted and/or still compete with livestock herds (Baldi et al. 2001; Dellafiore et al. 2003; Flueck and Smith-Flueck 2006; FAO 2010), as well as with several introduced mammal species (Buenavista and Palomares 2018).
In this study, we analyzed puma diet in a rangeland area of the Espinal ecoregion and reviewed literature available from South American temperate regions. Given native large prey depletion, increasing exotic prey availability (Buenavista and Palomares 2018), and widespread puma-livestock conflict (Kissling et al. 2009; Llanos et al. 2014; Llanos and Travaini 2020; Guerisoli et al. 2017; Iriarte-Walton et al. 2016), we expected that in this region: (1) puma diet would primarily rely on exotic prey (Buenavista and Palomares 2018), and (2) livestock would represents a common food source. Finally, since protected areas (PAs) main goal is long-term wildlife conservation (Day et al. 2012), we also expected that (3) pumas would preferentially prey on wild species inside PAs.
Materials and methods
The Espinal ecoregion, samples collection, and processing
This ecoregion is mostly flat, and natural vegetation is mainly characterized by xerophytic species (dominated by species of the genus Prosopis), which depending on their relative density create different habitats (woodlands, grasslands with shrubs, and grasslands; Distel 2016; Oyarzabal et al. 2018). However, because of agriculture and ranching expansion, natural habitats suffered heavy human modification (Nanni et al. 2020), especially in the southern portion of this region (Patagones and Villarino counties, Buenos Aires Province; Appendix 1). Here, extensive livestock (cattle Bos taurus and sheep Ovis aries) breeding is one of the major sources of economic income (SENASA 2015).
A previous camera-trapping study (sampling effort: 10,621 camera-trap days) completed in this area, revealed that pumas occurred in 35 out of 86 sampling sites (Guerisoli et al. 2019). Mammals species presented in this region include the guanaco, Patagonian mara Dolichotis patagonum, plains vizcacha Lagostomus maximus, armadillos (large hairy armadillo Chaetophractus villosus, screaming hairy armadillo C. vellerosus, and pichi Zaedyus pichiy), and small rodents (mainly Caviidae and Cricetidae families) (Arturi et al. 2005). In this area are also present large to medium-size bird species, particularly the common rhea and partridges (such as the elegant crested tinamou Eudromia elegans, spotted nothura Nothura maculosa, Darwin’s nothura Nothura darwinii, brushland tinamou Nothoprocta cinerascens, and red-winged tinamou Rhynchotus rufescens). Because of human persecution and livestock presence, guanacos and plains vizcachas are rare and spatially localized, whereas introduced species (wild boars and European hares) are relatively common (Arturi et al. 2005). In this study area, it was estimated, trough line transects, low densities of wild prey (e.g., range: 0.1–1.3 ind/km2; Olla 2016) compared to livestock densities (22.9 sheep/km2 and 54.2 cattle/km2; SENASA 2015). Thus, we suggest that livestock largely outweighed wild prey in most of the study area.
Between 2014 and 2016, puma feces were collected in a rangeland area in the southern part of the Espinal (Latitude 38.4–4° S, Longitude 63.4–62° W; Appendix 1, Fig. 1). We completed 15 transects stratified per habitat (woodlands, grasslands, and grassland with shrubs). Each transect included 2–3 operators and a scat-detection dog (Border collie breed trained by Matías Piedrabuena; “Got Scat?” Project, https://www.facebook.com/gotscat/). The dog was specifically trained to search the scent of puma feces’ and to avoid the scents of the other carnivores present in the area (Geoffroy’s cat Leopardus geoffroyi, Pampas cat Leopardus colocolo, Pampas fox Lycalopex gymnocercus, and dog Canis lupus domesticus). The length of each transect ranged from 1 to 7 km and averaged 3.9 km. Each transect was surveyed five to six times, achieving a total sampling effort of approximately 350 km. The identification of the pumas feces was based on the color, shape, texture, diameter, and macroscopic content (Yáñez et al. 1986). For each sample, we recorded the geographic coordinates. The feces were stored in paper bags, at room temperature, until further analyses.

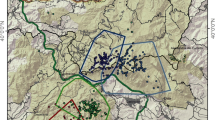
Location of the study areas of puma (Puma concolor) diet reviewed literature from the South American temperate region (area below the 35° S parallel; black line). Black lines show puma distribution range (IUCN, Nielsen et al. 2015) and gray polygons represent the PAs (WDPA). Map also includes the study area of our fieldwork-based data (#23). 1: Branch et al. 1996, 2: Elbroch and Wittmer 2013, 3: Fernández and Baldi 2014, 4: Ferreyra et al. 2010, 5: Gelin et al. 2017, 6: Iriarte et el. 1991, 7: Novaro et al. 2000, 8: Pessino et al. 2001, 9: Rau and Jiménez 2002, 10: Rau and Jiménez 2002, 11: Rau and Jiménez 2002, 12: Rau and Jiménez 2002, 13: Rau and Jiménez 2002, 14: Rau et al. 1991, 15: Skewes et al. 2012, 16: Zanón-Martínez et al. 2012, 17: Zanón-Martínez et al. 2012, 18: Zanón-Martínez et al. 2012, 19: Zanón-Martínez et al. 2012, 20: Zanón-Martínez et al. 2012, 21: Sarasola et al. 2016 and Zanón Martínez et al. 2016, 22: Zúñiga et al. 2005 and Zúñiga and Pedreros 2014, 23: our study, 24: Palacios, 25: LLanos and Travaini 2020
To assess the feces content, we followed the point-frame method, described in Ciucci et al. (2004). Feces were first volume-weighted, then soaked in water and soap for 24–48 h, disaggregated, and then thoroughly filtrated (0.7–0.5 mesh size) under running water. Once the samples were dried, we mixed and distributed each feces component in a 10 × 10 cm grid. Then, through a systematic sampling, we extracted 100 items (hairs) from each sample for identification. All the solid materials (e.g., bones, claws), of each feces, were identified through macroscopic identification (Pearson 1995; Rau and Jiménez 2002). To avoid inter-observer bias, only one trained observer (MMG) performed the items identification. Prey remains were identified to the lowest possible taxonomical level. The hairs were identified using an optical microscope and following the cuticle and medullar patterns for local mammals described in both Chehébar and Martín (1989) and Bonzano and Dellafiore (2018).
The prey consumption was expressed as the frequency of occurrence (FO; number of feces containing a given item divided by the total number of feces) and the relative biomass (B) for each detected item (i). For biomass estimates, we used the Ackerman’s linear correction factor (Ackerman et al. 1984; Bi) applying the following equations:
where Bi = prey biomass (kg)/feces and BMi = body weight of prey (item “i”) in kg.
The Ackerman’s correction factor (Ackerman et al. 1984) was designed especially for pumas, and we believe that, although it is a linear factor, it is the most suitable way to estimate prey biomass consumption in the temperate southern region. We expressed Bi as percentage, dividing the contribution of each item by the sum of all items contributions to the total biomass consumed. Since Bi estimation depends strongly on the prey body weight used for calculation, we identified livestock most predated age classes based on information obtained through interviews to ranchers and direct observations of puma killing sites from the field (see Guerisoli et al. 2017 for more details). We then used the average adult body weight for O. aries and the average under 12 months old calves and foals weights for B. taurus and Equus caballus, respectively. Finally, we extracted the remaining prey weights from different sources, specified in Table 1.
To understand if the sample size was representative of prey species presented in the area we applied a species accumulation curve (Foster et al. 2010) using the specaccum function of the Vegan R package (RStudio Team 2020).
Puma diet in South American temperate region: search criteria and data extraction for literature review
South American temperate region is a large area (1,570,150 km2) characterized by temperate climate (Köppen-Geiger climate classification; Beck et al. 2018) and comprehending portions of both Argentine and Chilean territories below the 35° S parallel (Fig. 1). We performed a systematic search of puma food habits studies with Scopus (Scopus 2019) and Google Scholar (Google 2019), using “key words” (e.g., Puma concolor + diet + feces). We included all the articles published until the year 2020 and located in the South American temperate region. The results obtained in the Espinal study area were also included in the review.
From each literature, we extracted the frequency of occurrence (FO) of each item (expressed as the number of feces containing a given item divided by the total number of feces), and when authors did not reported this value (e.g., Gelin et al. 2017), we calculated it based on the samples size and the number of occurrences of each item. We considered only the FO because provides an objective information about how often a given item is eaten (Zabala and Zuberogoitia 2003), and represents the most cited parameter in carnivore diet studies (e.g., Díaz-Ruiz et al. 2013; Soe et al. 2017). We used an overall FO values for multiple year sampling of a same study area (e.g., Zúñiga et al. 2005 and Zúñiga and Pedreros 2014; Appendix 2), while when a study presented multiple study areas results, we consider an FO per area (e.g., Rau and Jiménez 2002; Appendix 2). We discarded the “P. concolor” item since we assumed that could be derived from a grooming behavior.
Definition of prey categories and conservation areas for comparative analyses
Following Scognamillo et al. (2003), we categorized prey in three body size groups: small (< 1 kg), medium (1–15 kg), and large (> 15 kg). Additionally, we categorized food items into native, exotic (wild boar and European hare) and livestock (e.g., cattle, sheep). Because livestock breeding is not completely absent inside PAs, for both Argentina (e.g., APN 2019) and Chile (Praus et al. 2011), and following the initial hypothesis of a difference in puma feeding behavior depending on the conservation status of each study area, we categorized the researches inside or outside the PAs. Finally, we checked for significant differences between the mean FO of each prey category (prey size and exotic, native, livestock), and between inside and outside PAs, performing a Z-test. To overcome potential bias due to variation in sample size, we generated 95% confidence intervals (CI) around each prey category value through non-simultaneous bootstrap considering 1000 iterations with the library boot of RStudio Team (Manly 2018).
Results
Diet composition in a rangeland area of southernmost Espinal ecoregion
We collected 19 puma feces (corresponding to an encounter rate of 0.05 feces/km) with a feces mean volume of 45.8 (± 35 SD) ml. We detected 2.5 (± 1.4) food items per feces, and identified 14 different food items (11 to the species and/or genus level and three unidentified categories [armadillo, small rodents, and medium/small birds]; Table 1). The species accumulation curve did not reach a plateau, indicating that the number of samples was insufficient for a complete description of puma diet in this area (Appendix 3). The mean body weight of ingested prey was 31.5 (± 42.6, range: 0.4–100) kg. The most frequent prey species were “undetermined small rodent,” followed by plains vizcacha, European hare, and wild boar (Table 1). Although we detected livestock in approximately 16% of the samples, each single livestock species had a FO slightly more than 5% (Table 1). The Bi estimates showed a greater relative biomass contribution for large and medium prey than small prey, indicating that only four species (plains vizcacha, European hare, guanaco, and wild boar) exceeded 10% of biomass consumed (Table 1). Overall, native species represented the group with the largest ingested biomass, followed in descending order, by exotics and livestock (Table 1).
Literature reviewed and puma diet description in South American temperate region
We reviewed 18 papers containing relevant information on puma diet for the region, published between 1991 and 2020, and corresponding to 25 different study areas (Fig. 1; Appendix 2). Two papers described puma food habits in more than one study area, two involved the same area and the remaining were one-site surveys (Appendix 2). Counting also our field-based study, we found more information from Argentina (npub = 12, nstudyareas = 16) than Chile (npub = 6, nstudyareas = 10; Fig. 1). Most studies characterized puma diet through feces analysis, one through kill site inspections, and one with both stomach and intestine content (Appendix 2).
Hare (L. europaeus; 88.4%, n = 23, of the 26 study areas), guanaco (53.8%, n = 14 study areas), armadillos (Z. pichi and C. villosus; 50%, n = 13 study areas), and wild boar (30.7%, n = 8 study areas) were the most recorded wild species in puma diet (Table 2). Among livestock, sheep was the most common item (40% of the study areas), followed by cattle (Table 2). The FO varied across study areas, with the lowest FO for wild boar (0.1%) to the greatest for guanaco (83.5%) and hares (86%; Table 2). Although plains vizcacha and pudu were found in only 4 and 6 study areas, respectively, they presented high values of FO (12.8–87.1% and 8.3–64%, respectively; Table 2). The ranges of the FO of the other species are presented in Table 2.
Prey categories and conservation areas
Large prey category was composed mainly by livestock and native species, while medium and small prey categories was largely represented by native prey (Table 2). Medium prey tended to present a higher mean FO (24.8%, SD = ± 27%, n = 71) than large prey (19%, SD = ± 21.9%, n = 51). However, we did not detect any significant difference between their values (Z = − 1.6, p = 0.09), and CIs widely overlapped (Fig. 2, a1). Large and medium prey categories presented greater mean FO values than small prey (8.9%, SD = ± 10.3%, n = 48; large vs small: Z = 2.9, p < 0.05; medium vs small: Z = 4.9, p < 0.05) and their CIs did not present overlaps (Fig. 2, a2).

Mean frequency of occurrence (FO, %) and 95% confidence intervals (non-simultaneous bootstrap considering 1000 iterations) of the prey categories reported by puma diet studies (18 reviewed papers and field-based results from this study) from South America temperate region.. A1: size (large, medium, small), A2: species (native, exotic, livestock), B1 and B2 compare the same categories than A2 for inside and outside protected areas (PAs), respectively
Exotic species (34%, SD = ± 26.9%, n = 37) presented a higher mean FO than native (15.7%, SD = ± 20.3%, n = 107; Z = 3.9, p < 0.05) and livestock species (8.3%, SD = ± 9.1%, n = 25; Z = 5.5, p < 0.05). While the mean FO of native preys was higher than livestock species (Z = − 2.7, p < 0.05). The CIs showed very small overlap between livestock and native species (Fig. 2, a2).
Nineteen study areas were located inside PAs, while 7 were located in private lands (outside PAs). Sampled studies PAs were mainly concentrated along the Andes Mountains, varying in surface extension, from 0.89 to 21,689 km2 (Fig. 1). Puma diet tended to rely less on native species inside PAs (mean FO = 14.5%, SD = ± 20.1%, n = 84) than outside (mean FO = 19.9%, SD = ± 21%, n = 23), even if this difference was not significant (Z = − 0.8, p = 0.4). The higher mean FO in puma diet of exotic species inside PAs (35.9%, SD = ± 30.2%, n = 25) than outside (29.9%, SD = ± 18.6%, n = 12) was also not significant (Z = 0.8, p = 0.4). Finally, livestock mean FO inside PAs (9.1%, SD = ± 10%, n = 19) was significantly (Z = 2, p < 0.05) greater than outside (5.4%, SD = ± 4.7%, n = 6). Regardless of the similar CIs extents between protected and non-protected areas, exotic species CIs clearly showed that the FO values were greater than native and domestic species inside PAs and did not overlap with these categories (Fig. 2, B). On the other hand, CIs largely overlapped between exotic and native species outside PAs, while domestic species had smaller FOs and their CIs did not overlap with any of the other categories (Fig. 2, B).
Discussion
Diet composition in a rangeland area of southern Espinal ecoregion
Sample size
Under this productive context, where pumas are protagonist of an intense conflict with ranchers because of livestock predation (Guerisoli et al. 2017), this feline was mainly concentrated in the consumption of two exotic preys: the European hare and wild boar. Surprisingly, livestock species were underrepresented in the samples, while native prey (plains vizcacha and “undetermined small rodents”) were also among the first items in its diet. We acknowledge that the sample size (n = 19) was low. A small encounter rate of feces can be associated, for example, to weather conditions or to specific behaviors of felids (e.g., burying their feces; Reed et al. 2011). Considering the sampling effort completed and the use of a scat detection dog, we think that perhaps a low density of pumas, along with an intense human persecution of this species, could have affected the encounter rate. Although puma density has never been estimated for this study area, Gallo et al. (n.d.) estimated, for an area with similar habitat, 0.23 pumas/100km2, a density lower than 0.30 individuals/100km2 estimated for an area in northeast Argentina where this felid is strongly persecuted (Paviolo et al. 2009).
The species accumulation curve did not reach a plateau, indicating that the sample size was insufficient to completely describe puma diet. This was further supported by the fact that we did not detect common rhea remains through samples processing, despite the discovering of a puma kill-site on this species during fieldwork activities (Guerisoli personal observation). However, and in spite of the rarity and restricted distribution of some species, we detected two (L. guanicoe and S. scrofa) of the three (L. guanicoe, S. scrofa, and Rhea americana) large wild prey present in the area and that were already reported for puma diet in other studies (Skewes et al. 2012; Gelin et al. 2017). Still, further studies are needed to reliably describe large prey relevance for this felid diet in this area.
Puma diet in South American temperate region
Our results suggest that the European hare is the most detected wild prey in the southern temperate region, followed by guanaco and wild boar. In accordance with Buenavista and Palomares (2018), in more than half of the studies, exotic species were the most recorded items, showing greater FO than native prey. Additionally, the exotic wild boar and European hare are also among the five species with the greatest FO in puma feces in this vast region. Medium preys were the most represented size category in puma diet. This is consistent with Monroy-Vilchis et al. (2009), who indicated that pumas typically tend to prey on large mammals, whereas, at the southern limits of its distribution, they mainly feed on medium-sized prey. However, it has to be underlined that native species, with a size similar to the European hare (i.e., Patagonian mara and plains vizcacha), were present only in a small number of areas (Patagonian mara, n = 4 study areas, plains vizcacha, n = 4). Thus, for this size category (medium prey), our findings support the hypothesis proposed by Novaro et al. (2000) for northern Patagonia, where native species could have largely been replaced by exotic species. The same authors reported high-density estimates (45.4 ± 5 individuals/km2; Novaro et al. 2000) of European hare in unprotected areas. Consistently, this species has been found to represent an important food resource also for many mesopredators of the South American temperate region (i.e., culpeo Lycalopex culpaeus, chilla Lycalopex griseus, Geoffroy’s cat, and Pampas cat; Buenavista and Palomares 2018).
Among large prey, guanacos were the most recorded item (n = 14, 78.2% of the study areas). Due to hunting pressure, natural grassland deterioration, caused mainly by the sheep overgrazing (Golluscio et al. 1998), and direct interspecific competition with livestock, this native camelid has experienced an abrupt decline since the 1800s (Baldi et al. 2010). Although recent findings reported an increase of guanaco populations in the Argentine Patagonia (Gavuzzo et al. 2015), and most of the studies reviewed found this item, guanacos could be facing negative consequences due to wild boar density, which has been estimated to be around 35 km2/year in the Argentinean Patagonia (Pescador et al. 2009).
Regarding livestock, sheep were among all items, the third most common prey, but in general, the FO of livestock species was smaller than those of wild prey species. The puma is considered a conflictive species in the temperate region of South America, due to livestock predation (e.g., Novaro et al. 2000; Llanos et al. 2014; Guerisoli et al. 2017; Lucherini et al. 2018; Guerisoli et al. 2020). In this region, sheep prevalence in the diet, compared with cattle and goats, likely relates to the predominant abundance of this domestic species in Patagonia (FAO 2010), where additionally, pumas suffer heavy retaliation killing (Llanos et al. 2014). Food habits description represents an important tool/resource of information to complement data on pumas predation on livestock, which is recommended to be assessed also, with ranch monitoring and kill site surveys (Guerisoli et al. 2020), since livestock items can originate from opportunistic scavenging (Bauer et al. 2005).
Protected areas
Internationally recognized as one of the 37 biogeographic regions in the world which preserved its wild conditions (Mittermeier et al. 2003), the Patagonian ecoregion identifies almost the entire South American temperate region. Here, while most of the National Parks are located in proximity of the Andean mountain range (WDPA 2019), their number in eastern Patagonia (corresponding to the great plateau extending between the Andean mountain range towards the Atlantic coast; Steffen 1944) is almost null (WDPA 2019). Thus, the network of PAs in this territory is poorly conserving the most characteristic and widespread landscapes of the region: the Patagonian steppe. Most of the literature we reviewed involved protected areas, and yet 40% of them included O. aries in puma diet. Additionally, the FO of both exotic and livestock species were greater inside than outside PAs. More studies are needed to better understand the importance of conservation efforts in preserving this top predator’s ecological role inside PAs, as well as to more properly describe its trophic ecology outside PAs.
Based on the potential crucial role covered by exotic species as food resource, puma key role in prey populations control (Elbroch and Wittmer 2013; Barry et al. 2019) could be especially relevant for constraining the ongoing geographic expansion and population size increase of these species in the South American temperate region. Exotic species, including wild boar and European hare, can be responsible of serious conservation and management issues, including spread of diseases to livestock and people, vehicle collisions, reduction in native species abundance and richness (Jaksic 1998; Massei and Genov 2014). Thus, by predating on them, pumas not only control their populations, but also could provide an additional service by avoiding or reducing indirect issues that these species presence frequently cause (Barry et al. 2019). Conversely, exotic species could represent alternative prey to livestock, particularly when native species are less abundant, thus attenuating puma-human conflict. However, further research is required to properly understand predator-prey interactions in this vast region, where ecosystems and their vertebrate communities have been extensively altered by human action.
Data availability
We provide supplementary material.
References
Ackerman BB, Lindzey FG, Hemker TP (1984) Cougar food habits in southern Utah. J Wildl Manag 48:147–155
APN (2019) Administración de Parques Nacionales. Parque Nacional Los Alerces Plan de Gestión 2019-2029 (42 Pp). Argentina
Arturi M, Brown A, Martínez Ortíz U, Acerbi M, Corcuera J (2005) Ecorregión Espinal: Situación ambiental en la Ecorregión del Espinal. In: Situación Ambiental en Argentina. Fundación Vida Silvestre, Buenos Aires, pp 241–246
Baldi R, Albon S, Elston D (2001) Guanacos and sheep: evidence for continuing competition in arid Patagonia. Oecologia 129:561–570
Baldi G, Guerschman JP, Paruelo JM (2006) Characterizing fragmentation in temperate South America grasslands. Agric Ecosyst Environ 116:197–208
Baldi R, Novaro A, Funes M, Walker S, Ferrando P, Failla M, Carmanchahi P (2010) Guanaco management in Patagonian rangelands: a conservation opportunity on the brink of collapse. In: Du Toit JT, Kock R, Deutsch J (eds) Wild rangelands: conserving wildlife while maintaining livestock in semi-arid ecosystem, pp 266–290
Barry JM, Elbroch LM, Aiello-Lammens ME, Sarno RJ, Seelye L, Kusler A, Quigley HB, Grigione MM (2019) Pumas as ecosystem engineers: ungulate carcasses support beetle assemblages in the Greater Yellowstone Ecosystem. Oecologia 189:577–586
Bauer JW, Logan KA, Sweanor LL, Boyce WM (2005) Scavenging behavior in puma. Southwest Nat 50:466–472
Beck HE, Zimmermann NE, McVicar TR, Vergopolan N, Berg A, Wood EF (2018) Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci Data 5:180214
Bonzano M, Dellafiore CM (2018) Atlas para la identificación de pelos de mamíferos de Argentina. Fundación Bio Red, Buenos Aires
Branch LC, Pessino M, Villarreal D (1996) Response of Pumas to a Population Decline of the Plains Vizcacha. Journal of Mammalogy 77(4):1132–1140
Briske DD (2017) Rangeland systems: processes management and challenges. Springer, New York
Buenavista S, Palomares F (2018) The role of exotic mammals in the diet of native carnivores from South America. Mammal Rev 48:37–47
Carbone C, Mace GM, Roberts SC, Macdonald DW (1999) Energetic constraints on the diet of terrestrial carnivores. Nature 402:286–288
Cassaigne I, Medellín RA, Thompson RW, Culver M, Ochoa A, Vargas K, Torres-Gómez A (2016) Diet of pumas (Puma concolor) in Sonora, Mexico, as determined by GPS kill sites and molecular identified scat, with comments on jaguar (Panthera onca) diet. Southwest Nat 61:125–133
Chehébar C, Martín S (1989) Guía para el reconocimiento microscópico de los pelos de los mamíferos de la Patagonia. Doñana Acta Vertebrata 16:247–291
Ciucci P, Tosoni E, Boitani L (2004) Assessment of the point-frame method to quantify wolf Canis lupus diet by scat analysis. Wildl Biol 10:149–154
Day J, Dudley N, Hockings M, Holmes G, Laffoley D, Stolton S, Wells S (2012) Guidelines for applying the IUCN Protected Area Management Categories to Marine Protected Areas. Gland, Switzerland: IUCN. Pp: 36
Dellafiore CM, Demaría M, Maceira N, Bucher E (2003) Distribution and abundance of the pampas deer in San Luis province, Argentina. Mastozool Neotrop 10:41–47
Díaz-Ruiz F, Delibes-Mateos M, García-Moreno JL, María López-Martín J, Ferreira C, Ferreras P (2013) Biogeographical patterns in the diet of an opportunistic predator: the red fox Vulpes vulpes in the Iberian Peninsula. Mammal Rev 43:59–70
Distel RA (2016) Grazing ecology and theconservation of the Caldenal rangelands, Argentina. J Arid Environ 134:49–55
du Toit JT, Cross PC, Valeix M (2017) Managing the livestock–wildlife interface on rangelands. In: Briske DD (ed) Rangeland systems. Springer, Cham, pp 395–350
Elbroch LM, Quigley H (2019) Age-specific foraging strategies among pumas, and its implications for aiding ungulate populations through carnivore control. Cons Sci 1:e23
Elbroch LM, Wittmer HU (2013) The effects of puma prey selection and specialization on less abundant prey in Patagonia. J Mammal 94:259–268
FAO (2010) Food and agriculture organization of the United Nations statistical databases. See http://faostat.fao.org/
Fernández CS, Baldi R (2014) Feeding habits of the puma (Puma concolor) and the incidence of predation on guanaco (Lama guanicoe) mortality in northeastern Patagonia. Mastozoología Neotropical 21:331–338
Ferreyra N, Aprile G, Vila AR (2010) Dieta del Puma y Zorro Colorado en el Valle del Río de las Vueltas, Provincia de Santa Cruz. Informe preliminar 2010, pp. 14
Flueck WT, Smith-Flueck JM (2006) Why the Patagonian huemul deer in Argentina fails to recover: an ecological hypothesis. Proceedings of the 6th International Deer Biology Congress; 7-11 august, Prague, Czech Republic
Foster RJ, Harmsen BJ, Doncaster CP (2010) Sample-size effects on diet analysis from scats of jaguars and pumas. Mammalia 74:317–321
Gallo O, Castillo DF, Godinho R, Mac Allister ME, Fernández GP, Failla M, Casanave EB (n.d.) Molecular data revealed a structured puma (Puma concolor) population in northern Patagonia, Argentina. Mamm Biol MAMB-D-20-00047. In review
Gavuzzo AB, Gáspero P, Bernardos J, Pedrana J, De Lamo D, Von Thungen J (2015) Distribución y densidad de guanacos (Lama guanicoe) en la Patagonia, 1st edn. Bariloche, Río Negro
Gelin SML, Branch LC, Thornton DH, Novaro AJ, Gould MJ, Caragiulo A (2017) Response of pumas (Puma concolor) to migration of their primary prey in Patagonia. PLoS One 12:12
Golluscio RA, Deregibus VA, Paruelo JM (1998) Sustainability and range management in the Patagonia steppes. Ecol Austral 8:265–284
Gómez-Ortiz Y, Monroy-Vilchis O, Mendoza-Martínez GD (2015) Feeding interactions in an assemblage of terrestrial carnivores in central Mexico. Zool Stud 54:16
Guerisoli MDLM, Luengos Vidal E, Franchini M, Caruso N, Casanave EB, Lucherini M (2017) Characterization of puma–livestock conflicts in rangelands of central Argentina. R Soc Open Sci 4:170852
Guerisoli MDLM, Caruso N, Luengos Vidal EM, Lucherini M (2019) Habitat use and activity patterns of Puma concolor in a human-dominated landscape of central Argentina. J Mammal 100:202–211
Guerisoli M, Luengos Vidal E, Caruso N, Giordano A, Lucherini M (2020) Puma–livestock conflicts in the Americas: a review of the evidence. Mammal Rev. https://doi.org/10.1111/mam.12224
Husseman JS, Murray DL, Power G, Mack C, Wenger CR, Quigley H (2003) Assessing differential prey selection patterns between two sympatric large carnivores. Oikos 101:591–601
Inskip C, Zimmermann A (2009) Human-felid conflict: a review of patterns and priorities worldwide. Oryx 43:18–34
Iriarte JA, Johnson WE, Franklin WL (1991) Feeding ecology of the Patagonia puma in southernmost Chile. Revista Chilena de Historia Natural 64:145–156
Iriarte-Walton A, Sepúlveda C, Villalobos R, Lagos N (2016) El puma y el conflicto con la ganadería en Chile. In: Castaño-Uribe C, Lasso CA, Hoogesteijn R, Díaz-Pulido A, Payán E (eds) Conflictos humanos-felinos América Latina. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Colombia
Jaksic FM (1998) Vertebrate invaders and their ecological impacts in Chile. Biodivers Conserv 7:1427–1445
Jones KE, Bielby J, Cardillo M, Fritz SA, O’Dell J, Orme CDL, Connolly C (2009) PanTHERIA: a species-level database of life history, ecology, and geography of extant and recently extinct mammals. Ecology 90:2648–2648
Kissling DW, Fernández N, Paruelo JM (2009) Spatial risk assessment of livestock exposure to pumas in Patagonia, Argentina. Ecography 32:807–817
Kuijper DPJ, Sahlén E, Elmhagen B, Chamaillé-Jammes S, Sand H, Lone K, Cromsigt JPGM (2016) Paws without claws? Ecological effects of large carnivores in anthropogenic landscapes. P Roy Soc B-Biol Sci 283:20161625
Llanos R, Travaini A (2020) Diet of puma (Puma concolor) in sheep ranches of central Patagonia (Argentina). J Arid Environ 177:104145
Llanos R, Travaini A, Montanelli S, Crespo E (2014) Age structure of cougars (Puma concolor) hunted under the bounty system in Patagonia. Selective or opportunistic capture? Ecol Austral 24:311–319
Lucherini M, Guerisoli M, Luengos VE (2018) Surplus killing in pumas: rumors and facts. Mammal Rev 48:277–283
Lund HG (2007) Accounting for the world’s rangelands. Rangelands 29:3–10
Manly BF (2018) Randomization, bootstrap and Monte Carlo methods in biology. Chapman and Hall/CRC
Massei G, Genov P (2014) Preliminary analysis of food availability and habitat use by the wild boar in a Mediterranean area. J Mt Sci 3:168–170
Mittermeier RA, Mittermeier CG, Brooks TM, Pilgrim JD, Konstant WR, Da Fonseca GA, Kormos C (2003) Wilderness and biodiversity conservation. Proc Natl Acad Sci U S A 100:10309–10313
Monroy-Vilchis O, Urios V, Zarco-González M, Rodríguez-Soto C (2009) Cougar and jaguar habitat use and activity patterns in central Mexico. Anim Biol J 59:145–157
Murphy T, Macdonald DW (2010) Pumas and people: lessons in the landscape of tolerance from a widely distributed felid. In: Macdonald DW, Loveridge AJ (eds) Biology and conservation of wild felids. Oxford University Press, UK, pp 431–451
Nanni AS, Rodríguez MP, Rodríguez D, Regueiro MN, Periago ME, Aguiar S, Ballari S, Blundo C, Derlindati E, Di Blanco Y et al (2020) Presiones sobre la conservación asociadas al uso de la tierra en las ecorregiones terrestres de la Argentina. Ecol Austral 30:304–320
Newsome TM, Ripple WJ (2015) A continental scale trophic cascade from wolves through coyotes to foxes. J Anim Ecol 84:49–59
Nielsen C, Thompson D, Kelly M, Lopez-Gonzalez CA (2015) Puma concolor. The IUCN Red List of Threatened Species 2015-4. Downloaded on August 2015
Novack AJ, Main MB, Sunquist ME, Labisky RF (2005) Foraging ecology of jaguar (Panthera onca) and puma (Puma concolor) in hunted and non-hunted sites within the Maya Biosphere Reserve, Guatemala. J Zool 267:167–178
Novaro AJ, Funes MC, Walker RS (2000) Ecological extinction of native prey of a carnivore assemblage in Argentine Patagonia. Biol Conserv 92:25–33
Olla V (2016) Analisi della disponibilità trofica per la conservazione del puma Puma concolor nell’Espinal argentino (100 pp.). Master thesis, Universitá di Bologna, Bologna, Italy
Oyarzabal M, Clavijo JR, Oakley LJ, Biganzoli F, Tognetti PM, Barberis IM et al (2018) Unidades de vegetación de la Argentina. Ecol Austral 28:40–63
Pacheco LF, Lucero A, Villca M (2004) Dieta del puma (Puma concolor) en el Parque Nacional Sajama, Bolivia y su conflicto con la ganadería. Ecol Bolivia 39:75–83
Paviolo A, Di Blanco YE, De Angelo CD, Di Bitetti MS (2009) Protection affects the abundance and activity patterns of pumas in the Atlantic Forest. J Mammal 90:926–934
Pearson OP (1995) Annotated keys for identifying small mammals living in or near Nahuel Huapi National Park or Lanin National Park, southern Argentina. Mastozool Neotrop 2:99–148
Pescador M, Sanguinetti J, Pastore H, Peris S (2009) Expansion of the introduced wild boar (Sus scrofa) in the Andean region, Argentinean Patagonia. Galemys 21:121–132
Pessino ME, Sarasola JH, Wander C, Besoky N (2001) Respuesta a largo plazo del puma (Puma concolor) a una declinación poblacional de la vizcacha (Lagostomus maximus) en el desierto del Monte, Argentina. Ecol Austral 11:61–67
Pia MV (2013) Trophic interactions between puma and endemic culpeo fox after livestock removal in the high mountains of central Argentina. Mammalia 77:273–283
Praus S, Palma M, Domínguez R (2011) La situación jurídica de las actuales áreas protegidas de Chile (481 Pp). Santiago de Chile, Chile
Rau JR, Jiménez JE (2002) Diet of puma (Puma concolor, Carnivora: Felidae) in coastal and Andean ranges of southern Chile. Stud Neotropical Fauna Environ 37:201–205
Rau J, Tillería MS, Martínez DR, Muñoz AH (1991) Dieta de Felis concolor (Carnivora: Felidae) en áreas silvestres protegidas del sur de Chile. Rev Chil Hist Nat 64:139–144
Ray JC, Luke H, Zigouris J (2005) Setting conservation and research priorities for larger African carnivores. Vol. 24. New York: Wildlife Conservation Society, US
Reed SE, Bidlack AL, Hurt A, Getz WM (2011) Detection distance and environmental factors in conservation detection dog surveys. J Wildl Manag 75:243–251
Ripple WJ, Estes JA, Beschta RL, Wilmers CC, Ritchie EG, Hebblewhite M, Schmitz OJ (2014) Status and ecological effects of the world’s largest carnivores. Science 343:1241484
RStudio Team (2020) RStudio: integrated development for R. RStudio, PBC, Boston, URL http://www.rstudio.com/
Rueda P, Mendoza GD, Martínez D, Rosas-Rosas OC (2013) Determination of the jaguar (Panthera onca) and puma (Puma concolor) diet in a tropical forest in San Luis Potosi, Mexico. J Appl Anim Res 41:484–489
Ruth TK, Murphy K (2009) Cougar-prey relationships. In: Cougar: ecology and conservation. The Univ. of Chicago Press, Chicago, pp 138–155
Sarasola JH, Zanón-Martínez JI, Costán AS, Ripple WJ (2016) Hypercarnivorous apex predator could provide ecosystem services by dispersing seeds. Scientific Reports 6
Scognamillo D, Maxit I, Sunquist M, Polisar J (2003) Coexistence of jaguar (Panthera onca) and puma (Puma concolor) in a mosaic landscape in the Venezuelan llanos. J Zool 259:269–279
Scopus (2019) https://www.scopus.com/home.uri
SENASA (2015) Informes y estadísticas. http://www.senasa.gob.ar/cadena-animal/bovinos-y-bubalinos/informacion/informes-y-estadisticas
Skewes O, Moraga CA, Arriagada P, Rau JR (2012) El jabalí europeo (Sus scrofa): Un invasor biológico como presa reciente del puma (Puma concolor) en el sur de Chile. Rev Chil Hist Nat 85:227–232
Smith JA, Wang Y, Wilmers CC (2016) Spatial characteristics of residential development shift large carnivore prey habits. J Wildl Manag 80:1040–1048
Soe E, Davison J, Süld K, Valdmann H, Laurimaa L, Saarma U (2017) Europe-wide biogeographical patterns in the diet of an ecologically and epidemiologically important mesopredator, the red fox Vulpes vulpes: a quantitative review. Mammal Rev 47:198–211
Steffen H (1944) Patagonia occidental. Las Cordilleras Patagónicas y sus Regiones Circundantes. Ediciones de la Universidad de Chile. Santiago de Chile, Chile
Stolzenburg W (2008) Where the wild things were: life, death, and ecological wreckage in a land of vanishing predators. Bloomsbury, New York
Terborgh J, Estes JA (eds) (2013) Trophic cascades: predators, prey, and the changing dynamics of nature. Island press, Washington DC
Terborgh J, Estes JA, Paquet P, Ralls K, Boyd-Heigher D, Miller BJ, Noss RF (1999) The role of top carnivores in regulating terrestrial ecosystems. In: Soulé M, Terborgh J (eds) Continental conservation: scientific foundations of regional reserve networks. Island Press, Washington DC, pp 39–64
Treves A, Karanth KU (2003) Human-carnivore conflict and perspectives on carnivore management worldwide. Conserv Biol 17:1491–1499
WDPA (2019) World database on protected areas. IUCN. https://www.protectedplanet.net/
Wilcove DS, McLellan CH, Dobson AP (1986) Habitat fragmentation in the temperate zone. Conserv Biol 6:237–256
Yáñez JL, Cárdenas JC, Gezelle P, Jaksić FM (1986) Food habits of the southernmost mountain lions (Felis concolor) in South America: natural versus livestocked ranges. J Mammal 67:604–606
Zabala J, Zuberogoitia I (2003) Badger, Meles meles (Mustelidae, Carnivora), diet assessed through scat-analysis: a comparison and critique of different methods. Folia Zool 52:23–30
Zanón-Martínez JI, Kelly MJ, Mesa-Cruz JB, Sarasola JH, DeHart C, Travaini A (2016) Density and activity patterns of pumas in hunted and non-hunted areas in central Argentina. Wildlife Research 43:449
Zanón Martínez JI, Travaini A, Zapata S, Procopio D, Santillán MA (2012) The ecological role of native and introduced species in the diet of the puma in southern Patagonia. Oryx 46:106–111
Zúñiga AH, Pedreros AM (2014) Hábitos alimentarios de Puma concolor (Carnivora, Felidae) en bosques fragmentados del sur de Chile. Mastozool Neotrop 21:157–161
Zúñiga A, Quintana V, Fierro A (2005) Relaciones tróficas entre depredadores en un ambiente frag-mentado del sur de Chile. Gestión Ambiental 11:31–42
Acknowledgments
We thank the ranch owners, all the volunteers, all the students of the Master’s in International Ecology, Université de Sherbrooke and the staff of Instituto Nacional de Tecnología Agropecuaria. We are grateful to DVM G. Bravo for supplying the optical microscope, to Dra. E. Casanave for facilitating access to several resources, and to Dr. P. Teta for the help in the identification of rodent bone remains. Also, we want to acknowledge the revisors, the Associate Editor and Editor-in-Chief, which have improved the manuscript with their comments and suggestions. M.M.G. and O.G were funded by scholarships from the Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET). S.M. was funded by Agencia Nacional de Promoción Científica y Tecnológica (ANPCYT). This work was supported by Proyectos de Investigación Plurianuales (CONICET); Secretaría de Ciencia y Técnica-Universidad Nacional del Sur (PGI 24B/198); Wild Felid Research and Management Association; Conservation, Research and Education Opportunities International (CREOi, 2014–2015); Sacramento Zoo and Felidae Conservation Fund.
Funding
This work was supported by Proyectos de Investigación Plurianuales 2014–2016 (CONICET); Secretaría de Ciencia y Técnica-Universidad Nacional del Sur (PGI 24B/198); Wild Felid Research and Management Association; Conservation, Research and Education Opportunities International (CREOi, 2014–2015); Sacramento Zoo and Felidae Conservation Fund.
Author information
Authors and Affiliations
Contributions
Not applicable.
Corresponding author
Ethics declarations
Conflict of interest
Not applicable.
Ethics approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Yes, we consent.
Code availability
Not applicable.
Additional information
Communicated by: Krzysztof Schmidt
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Guerisoli, M.M., Gallo, O., Martinez, S. et al. Native, exotic, and livestock prey: assessment of puma Puma concolor diet in South American temperate region. Mamm Res 66, 33–43 (2021). https://doi.org/10.1007/s13364-020-00549-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13364-020-00549-0