
Abstract
In 2005–2008, we radio-tracked 17 foxes in rural areas of Southern Germany. The mean home range size was 76.6 ha (95% MCP) or 138.9 ha (95% fixed kernel), and the built-up area formed an integral part of the home range. Home ranges of juvenile foxes were significantly smaller than home ranges of adult foxes. Gender-specific differences among adult foxes were not established. A minimum population density of 2.7 foxes per km2 and summer densities of up to 13.4 foxes per km2 were calculated. Therefore, the fox density was three to eight times higher than that of strictly rural foxes. Daytime resting sites of foxes were mostly found in forests (62.2%) and reedbed areas (20.6%). Of the resting sites, 14.8% were situated inside settlements, in fallow gardens or gardens of residents. During the day, foxes exhibited habitat preferences for forests and reedbed areas. A habitat structure that offers plenty of cover or dense vegetation is essential for its selection as a safe resting site. If this basic requirement is fulfilled, foxes also choose resting sites within settlements, and are not disturbed by human presence.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
A characteristic of the red fox (Vulpes vulpes) is its ability to thrive in a wide spectrum of habitats, in essence due to its omnivorous diet and high adaptability (Macdonald 1983; Kidawa and Kowalczyk 2011). This wide spectrum is particularly exhibited in the size of its home range, which correlates with the habitat (Macdonald 1981). According to Burt’s definition (1943), the home range is “that area traversed by the individual in its normal activities of food gathering, mating, and carrying for young”. This concept has been criticized, discussed and redefined, but remains unchanged in its central ideas (Millspaugh and Marzluff 2001). Home range size is thus delimited by the spatially available resources or by energetic and/or biophysical limitations (Macdonald 1981, 1983; Kelt and Van Vuren 2001). The size also depends on balancing energy expenditure and cost–benefit analysis (Davies and Houston 1984): larger territories may contain more resources, but they are more difficult to control and defend (Macdonald 1983). Home range size is thus determined by the habitat or the availability of resources within it. Generally, foxes in rural areas in Europe occupy home ranges of >200 ha (Reynolds and Tapper 1995; Cavallini and Lovari 1994; Berghout 2000). In urban habitats, home ranges of <50 ha and fox population densities of >10 animals per km2 have been estimated (Harris 1980; Harris and Smith 1987; König 2005; Marks and Bloomfield 2006). In rural parts of Germany, densities of 0.43 foxes per km2 have been established in Oberlausitz (Ansorge 1991), and 0.74 foxes per km2 in the district of Garmisch Partenkirchen (Vos 1995). In urban areas, fox population densities are generally higher than in rural areas (Harris and Rayner 1986; Trewhella et al. 1988).
Only patchy scientific data are available for a habitat which, by the terms of the definitions, is neither completely urban nor 100% rural. For villages and small towns with <10,000 inhabitants, few studies on the spatial behaviour of red foxes are available (Janko 2003; Thoma 2008; Janko and König 2011). The aim of this study was to investigate the home range size and population density of red foxes living nearby villages and small towns. Additionally, habitat selection and habitat preferences for resting sites were analysed to establish the conditions under which foxes choose resting sites outside and inside rural settlements.
Material and methods
Study area
The study area is in the district of Starnberg, southwest of Munich, and is situated in the state of Bavaria. The study was carried out in the communes of Herrsching am Ammersee (48°00′ N, 11°10′ E, 568 m above sea level; area, 20.9 km2), Seefeld (48°02′ N, 11°14′ E, 570 m above sea level; area, 34.9 km2) and Andechs (47°58′ N, 11°11′ E, 690 m above sea level; area, 40.4 km2). The settlements are situated in a rurally structured landscape. This varies from cultivated agricultural areas to grasslands and forests. A great number of ponds with reedbeds are scattered across the countryside. Larger reedbed areas lie to the north of Herrsching and north of the Pilsensee. The small town of Herrsching (8,606 inhabitants) covers a developed area of 3.0 km2. Seefeld covers an area of 1.1 km2 and is home to around 4,300 people.
Capturing and radio-tracking
From July 2005 to August 2008, foxes were captured within the settlement or on the settlement edge (≤250 m) using live traps. Twenty-five wooden box traps (160 × 43 × 26 cm) were set up inside the study area (6.1 traps/km2) in residents’ gardens, fallow gardens, along railway lines or in patches of forest. Traps were baited with dog food (Frolic).
A total of 22 foxes were caught and radio-collared: 12 animals in gardens within the settlement, and 10 on the settlement edge. One adult male and one juvenile vixen were caught twice. Of the 22 foxes, 8 died during the course of the 3-year study. Four were killed by traffic, three were shot and one fox was found dead under a garden shed (Table 1). Captured foxes were immobilised with an intramuscular dart administered by blowpipe and containing the sedative “Hellabrunner Mischung” (400 ml ketamine, 500 mg xylazine) (Wiesner 1998). Depending on body weight, the administered dose ranged between 0.4 and 0.6 ml (0.08–0.1 ml/kg). After immobilisation, the animals did not display signs of trauma or behavioural abnormalities (Janko et al. 2011). The age (juvenile and adult) was determined by examining tooth wear (Harris 1978). For data evaluation, young foxes were classified from 31st of January of the following year as adult animals, in accordance with Stiebling (2000). The animals were fitted with VHF collars (Biotrack, GB, TW-3). The system consisted of a receiver (Televilt RX-98, Sweden; Telonics TR-4, USA), H-antenna and headphones. The telemetric measurements were made on foot, by car or a combination of the two. At night, bearings from the animals were recorded over several hours at intervals ≤15 min by triangulation (Harris et al. 1990). The distance to the fox was seldom more than 300 m, so that serious errors with the bearings can be ruled out almost entirely. Within the settlement, the distance to the fox was shorter—often under 50 m. In the course of the study, it became clear that good orientation on the ground and the minimized distance to the foxes resulted in a high accuracy of bearings (Riley et al. 2002). During the day, foxes were localised within their daytime resting sites 2–4 days per week. The inactivity of the foxes was established by the stable signal strength of the radio transmitter (Kowalczyk and Zalewski 2011) over more than 2–4 min.
Calculation of home range, habitat selection and population density
To evaluate the normal distribution of the data set, we used the Kolmogorov–Smirnov test. The data of 17 of 22 captured and radio-tagged foxes were used for further analysis, with a total of 4,371 bearings. All bearings were entered in ArcGIS 8.1, and subsequently, home ranges sizes were calculated using the program Biotas 1.0.1 alpha (Ecological Software Solutions, Switzerland). Home ranges were calculated using the minimum convex polygon (MCP) method (Mohr 1947; Kenward 1987). As a second home range estimator, fixed kernel density estimation (KDE) (Worton 1989) was applied. For MCP and KDE, we applied 95% of all bearings taken for each fox to represent the foxes’ home range sizes (95MCP and 95KDE). Age- and gender-specific differences were established using the Mann–Whitney U test (SPSS 17.0).
The daytime resting sites and the habitat availability of foxes were differentiated according to the following categories: forest, reed, settlement, arable land and grassland. In total, 825 bearings of foxes were taken, and the location of 99 daytime resting sites could be analysed. A preference ranking of habitat classes for daytime resting sites was calculated as developed by Aebischer et al. (1993), whereby the use of resting sites is related to habitat availability within the 95MCP home range. The habitat availability for 17 foxes within the 95MCP range was calculated using the program Biotas 1.0.1 alpha. Forest was used as the reference habitat class (Aitchison 1982).
Population densities were calculated on the basis of the mean home range sizes of adults and juveniles and assuming that there was no gender-specific overlapping (Herr et al. 2009), and on the assumption that a minimum of two foxes (male and female) live in one home range (Macdonald 1983). According to Macdonald (1983), the home range will reliably support a pair of foxes and may sometimes support additional foxes because group sizes are limited by the availability of prey. This assumption allows us to estimate a “minimum population density” for the small town habitat.
Results
Home ranges
The mean home range size was 76.6 ± 44.1 (±SD) ha for 95MCP and 138.9 ± 85.8 ha for 95KDE. For adult foxes (n = 11), the 95MCP mean home range size was 99.0 ± 36.8 ha and 176.5 ± 81.1 ha using the 95KDE (Fig. 1). Mean home ranges of juvenile foxes (n = 6) were smaller, at 35.3 ± 19.0 ha 95MCP or 69.9 ± 39.4 ha 95KDE. Age-specific differences in home range sizes were significant, both within the 95MCP (p = 0.003) and within the 95KDE (p = 0.007). No gender-specific differences for adult animals could be found (95MCP, p = 0.662; 95KDE, p = 0.715).

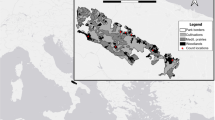
95%MCP home ranges (polygonal shapes) and daytime resting sites (crossed circles) of red foxes in the small town of Herrsching
Daytime resting sites
During the day, the resting sites of foxes were mostly found in forests (62.2%) and reedbed areas (20.6%) (n = 826 bearings). Forests are of fundamental importance for foxes; 16 of the 17 foxes used at least one resting site in the forest. Resting sites were concentrated on forested slopes in the vicinity of the village, in dense spruce or deciduous thickets and in reedbed areas. In 14.8% of cases, foxes rested within villages and small towns and to 2.4% within grain fields. Four of the 17 foxes spent the day in fallow gardens and in residents’ gardens within the urban area (under garden sheds, in dense bushes, behind stacks of wood). Foxes used between one and ten resting sites within their home range (n = 99, mean = 3.85). Significant habitat preferences are exhibited for forests and reedbed areas (Table 2).
Population density
Based on the mean home range size of 76.6 ha (95MCP), a minimum population density of 2.7 foxes per km2 was calculated. Differentiating between adult and juvenile foxes and assuming that vixen in central Europe bear four to five cubs (Keuling et al. 2010), the following summer density was calculated: given four cubs per litter, the density was 11.3 juvenile foxes per km2. For adult foxes, there was a density of 2.1 animals per km2. Added together, the summer density was thus 13.4 foxes (adult and juvenile) per km2.
Discussion
A comparison with urban and rural foxes shows that the estimated home range is intermediate in size. Rural foxes use home ranges larger than 200 ha in size (Cavallini and Lovari 1994; Reynolds and Tapper 1995; Weber and Meia 1996; Dekker et al. 2001). Urban foxes establish smaller home ranges, between 12 and 63 ha (Harris 1980; Macdonald 1981; Adkins and Stott 1998; White et al. 2006; Marks and Bloomfield 2006). In contrast to this, village foxes use both the open countryside and the settlement area. Here, residents’ gardens are well frequented (Janko and König 2011). One reason for the concentration on gardens is the high food availability (Macdonald 1981; Harris 1981; Janko 2003; Contesse et al. 2004). The availability of resources is a key factor in home range delimitation (Macdonald 1983; Kelt and Van Vuren 2001). The estimated home range size is in accordance with the tendency for increasing urbanization of the red fox to be linked to a decrease in home range size. This effect could also be observed for other omnivorous habitat generalists such as for coyotes (Atwood et al. 2004) or raccoon dogs (Prange et al. 2004). The fact that there are no gender-specific differences but that there are age-related differences in fox’ home range sizes is in accordance with other studies (Cavallini and Lovari 1994; Poulle et al. 1994).
During the daytime, only a few foxes rest inside the settlement area, under garden sheds or in fallow gardens. These quarters are characteristic resting sites of urban foxes (Harris 1977; König 2005). There is however a clear preference for natural habitats, which is typical of rural foxes (Cavallini and Lovari 1994; Reynolds and Tapper 1995). The forest was the most important daytime retreat for foxes in the study area. The preference for reedbeds occurs in large, connected areas, and in small, fragmented areas. Resting sites in reedbeds should therefore not be considered as compensation for a lack of forests. They represent an independent and important resting area. Resting sites in thickets, on fallow gardens or in reedbeds show that habitat-related factors play a key role. The selection of the resting site is primarily regulated by a high level of cover or vegetation. This provides the foxes with the safety they require. If these prerequisites are met, foxes rest in the vicinity of settlements, hiking trails, forest roads or even within settlements. Human proximity does not lead to foxes avoiding these structures. For village foxes, the proximity of resting sites to the settlement allows rapid access to lucrative food sources after the resting period. In cities, omnivorous carnivores tend to choose natural, quiet habitats during the day (Riley et al. 2003; Adams and Lindsey 2010). A habitat structure that offers plenty of cover is thus crucial for the selection of safe resting sites (Reynolds and Tapper 1995; Marks and Bloomfield 2006), and it is a limiting factor for fox populations in urban areas (White et al. 2006).
Seen in context, the reduction in home range size has the effect of raising population density (Harris 1980). In rural areas, there are between 0.34 and 1.06 foxes per km2 (Ansorge 1991; Vos 1995; Heydon et al. 2000; Keuling et al. 2010). Fox density in villages (2.7 foxes/km2) is thus raised by a factor of 3 to 8. To make precise statements about the population density, the size of a family group has to be known. In this study, precise family sizes were not established. Therefore, the calculated minimum density was definitely the lowest population density level. This density could be increased by subadult foxes (Reynolds and Tapper 1995) or by floaters (Gloor 2002). The increased fox population density in villages and small towns fits in with the investigations on population dynamics conducted by Trewhella et al. (1988). It is postulated that villages and small towns offer enough resources to effect a reduction in home range size, but not enough to build up autonomous urban fox population. Small town foxes are moderately adapted to settlements (or humans), and at the present time, it is unclear at what point a transition to an autonomous urban fox population becomes possible. This transition point is largely dependent on the size of the urban area as this affects the availability of the three essential resources (food, dens, daytime resting sites). Important co-factors are the housing density (Harris and Rayner 1986), attitudes towards wild animals (König 2008) and the migration and mortality pattern of the red fox (Krebs 1985).
References
Adams CE, Lindsey KJ (2010) Urban wildlife management. CRC Press, Boca Raton
Adkins CA, Stott P (1998) Home ranges, movements and habitat associations of red foxes Vulpes vulpes in suburban Toronto, Ontario. J Zool 244:335–346
Aebischer NJ, Robertson PA, Kenward RF (1993) Compositional analysis of habitat use from animal radio-tracking data. Ecology 74:1313–1325
Aitchison J (1982) The statistical analysis of compositional data. J R Stat Soc 44:139–177
Ansorge H (1991) Die Ernährungsökologie des Rotfuchses, Vulpes vulpes, in der Oberlausitz während des Winterhalbjahres. Abhandlungen und Berichte des Naturkundemuseums Görlitz 65:1–24
Atwood TC, Weeks HP, Gehring TM (2004) Spatial ecology of coyotes along a suburban-to-rural gradient. J Wildl Manag 68:1000–1009
Berghout M (2000) The ecology of the red fox (Vulpes vulpes) in the central tablelands of New South Wales. Dissertation, University of Canberra
Burt WH (1943) Territoriality and home range concepts as applied mammals. J Mammal 24:346–352
Cavallini P, Lovari S (1994) Home range, habitat selection and activity of the red fox in a Mediterranean coastal ecotone. Acta Theriologica 39:279–287
Contesse P, Hegglin D, Gloor S, Bontadina F, Deplazes P (2004) The diet of urban foxes (Vulpes vulpes) and the availability of anthrophogenic food in the city of Zürich, Switzerland. Mamm Biol 69:81–95
Davies NB, Houston AI (1984) Territory economics. In: Krebs JR, Davies NB (eds) Behavioural ecology, an evolutionary approach, 2nd edn. Blackwell, Oxford, pp 148–169
Dekker J, Stein A, Heitkönig I (2001) A spatial analysis of a population of red fox (Vulpes vulpes) in a Dutch coastal dune area. J Zool 255:505–510
Gloor S (2002) The rise of urban foxes (Vulpes vulpes) in Switzerland and ecological and parasitological aspects of a fox population in the recently colonised city of Zürich. Dissertation, Universität Zürich
Harris S (1977) The food of suburban foxes (Vulpes vulpes), with special reference to London. Mamm Rev 11:151–168
Harris S (1978) Age determination in the red fox (Vulpes vulpes)—an evaluation of technique efficiency as applied to a sample of suburban foxes. J Zool 184:91–117
Harris S (1980) Home range and patterns of distribution of foxes (Vulpes vulpes) in an urban area, as revealed by radio tracking. In: Amlaner CJ, Macdonald DW (eds) A handbook on biotelemetry and radiotracking. Pergamon Press, Oxford, pp 685–690
Harris S (1981) The food of suburban foxes (Vulpes vulpes), with special reference to London. Mamm Rev 11:151–168
Harris S, Rayner JMV (1986) Urban fox (Vulpes vulpes) populations estimates and habitat requirements in several British cities. J Anim Ecol 55:575–591
Harris S, Smith GC (1987) Demography of two urban fox (Vulpes vulpes) populations. J Appl Ecol 24:75–86
Harris S, Cresswell WJ, Forrde PG, Trewhella WJ, Woolard T, Wray S (1990) Home range analysis using radio-tracking data—a review of problems and techniques particularly as applied to the study of mammals. Mamm Rev 20:97–123
Herr J, Schley L, Roper J (2009) Socio-spatial organization of urban stone martens. J Zool 277:54–62
Heydon MJ, Reynolds JC, Short MJ (2000) Variation in abundance of foxes (Vulpes vulpes) between three regions of rural Britain, in relation to landscape and other variables. J Zool 251:253–264
Janko C (2003) Habitatnutzung des Rotfuchses (Vulpes vulpes) in Dörfern und Kleinstädten. Diploma thesis, Universität Hohenheim
Janko C, König A (2011) Disappearance rate of praziquantel- containing baits around villages and small towns in southern Bavaria, Germany. J Wildlife Diseases 47:373–38
Janko C, Börner K, König A, Schneider R (2011) Fang, Immobilisation und Besenderung von Rotfüchsen (Vulpes vulpes L.) - ein Erfahrungsbericht aus der Praxis. Beiträge zur Jagd- und Wildforschung 36:265–275
Kelt DA, Van Vuren DH (2001) The ecology and macroecology of mammalian home range area. Am Nat 156:637–645
Keuling O, Greiser G, Grauer A, Strauß E, Bartel-Steinbach M, Klein R, Wenzelides L, Winter A (2010) The German wildlife information system (WILD): population densities and den use of red foxes (Vulpes vulpes) and badgers (Meles meles) during 2003–2007 in Germany. Eur J Wildl Res. doi:10.1007/s10344-010-0403-z
Kenward R (1987) Wildlife radio tagging: equipment, field techniques and data analysis. Academic, London
Kidawa D, Kowalczyk R (2011) The effects of sex, age, season and habitat on diet of the red fox Vulpes vulpes in northeastern, Poland. Acta Theriol 56:209–218
König A (2005) Neue Untersuchungsergebnisse zur Ausbreitung des Kleinen Fuchsbandwurms (Echinococcus multilocularis) im Großraum München. In: Bayerische Akademie d. Wissenschaften (ed), Rundgespräche der Kommission für Ökologie, Band 29: Zur Ökologie von Infektionskrankheiten: Borreliose, FSME und Fuchsbandwurm. Verlag Dr. Friedrich Pfeil, pp. 71–84
König A (2008) Fears, attitudes and opinions of suburban residents with regards to their urban foxes. Eur J Wildl Res 54:101–109
Kowalczyk R, Zalewski A (2011) Adaption to cold and predation—shelter use by invasive raccoon dogs Nyctereutes procyonoides in Bialowieza primeval forest (Poland). Eur J Wildl Res 57:133–142
Krebs CJ (1985) Ecology: the experimental analysis of distribution and abundance. Harper and Row, New York
Macdonald DW (1981) Resource dispersion and the social organization of the red fox (Vulpes vulpes). In: Worldwide Furbearer Conference 2, pp. 918–949
Macdonald DW (1983) The ecology of carnivore social behaviour. Nature 301:379–384
Marks CA, Bloomfield TE (2006) Home-range size and selection of natal den and diurnal shelter sites by urban red foxes (Vulpes vulpes) in Melbourne. Wildl Res 33:339–347
Millspaugh JJ, Marzluff JM (2001) Radio tracking and animal populations. Academic, London
Mohr CO (1947) Table of equivalent populations of North American small mammals. Am Midl Nat 37:223–249
Poulle ML, Artois M, Roeder JJ (1994) Dynamics of spatial relationships among members of a fox group (Vulpes vulpes: Mammalia: Carnivora). J Zool 233:93–106
Prange S, Gehrt SD, Wiggers EP (2004) Influences of anthropogenic resources on raccoon (Procyon lotor) movements and spatial distribution. J Mammal 85:483–490
Reynolds JC, Tapper SC (1995) The ecology of the red fox Vulpes vulpes in relation to small game in rural southern England. Wildl Biol 1:105–119
Riley SPD, Sauvajot RM, Fuller TK, York EC, Kamradt DA, Bromley C, Wayne RK (2002) Effects of urbanization and habitat fragmentation on bobcats and coyotes in Southern California. Conserv Biol 17:566–576
Stiebling U (2000) Untersuchungen zur Habitatnutzung des Rotfuchses, Vulpes vulpes (L. 1758), in der Agrarlandschaft als Grundlage für die Entwicklung von Strategien des Natur- und Artenschutzes sowie der Tierseuchenbekämpfung. Dissertation, Humboldt- Universität Berlin
Thoma D (2008) Untersuchungen zum urbanen Übertragungszyklus von Echinococcus multilocularis. Dissertation, Universität Hohenheim
Trewhella WJ, Harris S, McAllister FE (1988) Dispersal distance, home-range size and population density in the red fox (Vulpes vulpes): a quantitative analysis. J Appl Ecol 25:423–434
Vos A (1995) Population dynamics of the red fox (Vulpes vulpes) after the disappearance of rabies in county Garmisch Partenkirchen, Germany, 1987–1992. Ann Zool Fennici 32:93–97
Weber JM, Meia JS (1996) Habitat use by the red fox Vulpes vulpes in a mountainous area. Ethol Ecol Evol 8:223–232
White JG, Gubiani R, Smallman N, Snell K, Morton A (2006) Home range, habitat selection and diet of foxes (Vulpes vulpes) in a semi-urban riparian environment. Wildl Res 33:175–180
Wiesner H (1998) Tierschutzrelevante Neuentwicklungen zur Optimierung der Distanzimmobilisation. Tierärztliche Praxis 26:225–233
Worton BJ (1989) Kernel methods for estimating the utilization distribution in home-range studies. Ecology 70:164–168
Acknowledgements
We would like to thank all collaborators, field helpers and the communities of Andechs, Herrsching and Seefeld. Special thanks to the Konrad Adenauer Stiftung for furthering this work with a personal scholarship. Additionally, we would like to thank several companies for financial contributions that enabled us to purchase telemetry equipment and collars.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by: Karol Zub
Rights and permissions
About this article
Cite this article
Janko, C., Schröder, W., Linke, S. et al. Space use and resting site selection of red foxes (Vulpes vulpes) living near villages and small towns in Southern Germany. Acta Theriol 57, 245–250 (2012). https://doi.org/10.1007/s13364-012-0074-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13364-012-0074-0