
Abstract
LATS2 (Large tumor suppressor) has been reported to be dys-regulated in several cancer types. However, its function in non-small cell lung cancer (NSCLC) remains poorly understood. Here, it was found that the expression level of LATS2 was decreased in NSCLC tissues. Moreover, forced expression of LATS2 in NSCLC cells inhibited cell growth and migration, while knockdown of the expression of LATS2 promoted the tumorigenicity of NSCLC cells. Mechanistically, LATS2 was found to negatively regulate NF-κB signaling in NSCLC cells. Taken together, our study suggested that down-regulation of LATS2 was very important in the progression of NSCLC, and restoring the function of LATS2 might be a promising therapeutic strategy for NSCLC.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Lung cancer remains the most common cause of cancer-related death worldwide, and the incidence of lung cancer is increasing [1]. Non-small cell lung cancer (NSCLC) is the primary histological classification of lung cancer [2]. There are three histologic subtypes of NSCLC: squamous cell carcinoma (SCC), adenocarcinoma (AC), and large-cell carcinoma (LCC) [3]. Overall, 5-year survival of NSCLC has remained at 15 % for the last two decades and the prognosis of patients with NSCLC principally correlates with tumor invasion and metastasis [4]. The invasiveness of lung cancer is characterized by multiple alterations of gene expression, which lead to abnormal changes in signaling pathways and biological behaviors [5, 6]. Tumor invasion and metastasis greatly limit treatment options and account for 90 % of NSCLCs [7]. Thus, it is important to search for new therapeutic targets involved in the regulation of lung cancer cell growth and invasion.
Large tumor suppressor (LATS), which encodes a putative serine/threonine kinase, has been identified as a tumor suppressor gene in Drosophila [8]. Disruption of the lats gene function results in the promotion of cell proliferation and tumor formation in Drosophila [8]. Two mammalian homologues of the Drosophila lats (lats1 and lats2) have been identified [9]. LATS1 and LATS2 are central players within the conserved Hippo signaling pathway [10]. Components of this pathway include the adaptor proteins WW45 and MOB which bring LATS1/2 in contact with the kinase Mst1/2 which phosphorylates and activates LATS1/2 [11].
Besides Hippo-LATS network, the molecular biology studies have showed that LATS is coupled to several well-established cell signaling pathways, in particular, the Ras, Rac/Cdc42 GTPases, p53 pathway [12–14]. Firstly, constitutively active Ras induces LATS2 expression and loss of LATS2 cooperates with oncogenic Ras to promote tumorigenesis. Secondly, the interaction with LIMK1 implicates LATS1 in the maintenance of genetic stability. Alternatively, this interaction could potentially link LATS1 to Rac/Cdc42-mediated actin dynamics at the leading edge of cells, which is a possible mechanism of LATS-inhibited cell migration. Thirdly, significant evidence links LATS to p53, including its role in mediating LATS effects on the G1 checkpoint as well as the identification of overlapping transcriptional targets between LATS and p53, such as p21 and TP53INP1 [14].
Overexpression of LATS1 can inhibit cell proliferation by inducing G2/M arrest [15], whereas LATS2 has been reported to inhibit cell growth by causing G1/S or G2/M cell cycle arrest [16]. Dysregulation of LATS1 and/or LATS2 has been implicated in soft tissue sarcomas, leukemia, astrocytoma, and in cancers of the breast, prostate, lung, liver, and esophagus [17–23]. Decreased expression of LATS2 is seen in prostate cancer tissues [23, 24]. Previous studies have found that LATS1 inhibited the proliferation and invasion of NSCLC cells [24]. However, the function of LATS2 in NSCLC has not been well studied. In this study, it was found that the expression of LATS2 was down-regulated in NSCLC clinical samples and cell lines. Overexpression of LATS2 inhibited the growth and migration of NSCLC cells, while knockdown of the expression of LATS2 promoted the growth, migration and metastasis of NSCLC cells. Mechanistically, LATS2 was found to inhibit NF-κB signaling in NSCLC cells. Collectively, our study revealed the tumor suppressive role of LATS2 in NSCLC.
Materials and methods
Cell culture
Human normal lung bronchus epithelial cell line BEAS-2B and NSCLC cell lines A549, H1299, H520, and H23 were purchased from ATCC (American Type Culture Collection) and cultured in Dulbecco’s modified Eagle’s medium (Invitrogen) supplemented with 10 % fetal bovine serum (FBS; PAA Laboratories, Pasching, Austria), 10 units/ml penicillin-G, and 10 mg/ml streptomycin. All cells were incubated at 37 °C in a humidified atmosphere containing 5 % CO2.
Clinical samples
Fifty-six NSCLC tissues (28AC, 22SCC, and 6LCC; 37 male patients and 19 female patients) were collected from patients who received surgery for NSCLC at Shanghai Chest Hospital which was affiliated to Shanghai Jiaotong University for this study. All of the patients have given informed consent. Dissected samples were frozen immediately after surgery and stored at −80 °C until needed.
Plasmid construction and transfection
To generate the LATS2 expression vector, the open reading frame of human LATS2 complementary DNA (cDNA) was cloned into the expression vector pcDNA3.1. The LATS2 expression vector and empty pcDNA3.1 were transfected into H23 and H520 cells using Lipofectamine 2000 reagent (Invitrogen). The transfected cells were selected in the presence of 600 μg/ml G418, and resistant cells were pooled and further confirmed the expression of exogenous LATS2 by Western blot.
RNA extraction and real-time PCR (RT-PCR) analysis
Total RNA was isolated from NSCLC tissues and matched normal tissues of NSCLC patients after their informed consent using TRIzol reagent (Invitrogen) according to the manufacturer’s instructions. The RNA samples were separated in 2 % agarose gels containing ethidium bromide, and their quality was then determined by visibility of 18 and 28S RNA bands under UV light. Two micrograms of total RNA with high quality (light absorbance A260/280 = 2.0) was processed directly to cDNA with the reverse transcription kit (Promega, Madison, WI), following the manufacturer’s instructions, in a total volume of 25 μl. The cDNA was diluted before used (1:20). The primer pair used for amplification of the human LATS2 gene was as follows: forward primer, 5′-AGCTGGACTCTGTGAAGCTG-3′, and reverse primer, 5′-TGTCCACCTTACAAGCAAGG-3′. As an internal standard, a fragment of human beta-actin was amplified by PCR using the following primers: forward primer, 5′-GATCATTGCTCCTCCTGAGC-3′, and reverse primer, 5′-ACTCCT GCTTGCTGATCCAC-3′. Amplification reactions were performed in a 20 μl volume of the LightCycler-DNA Master SYBR Green I mixture from Roche Applied Science as follows: with 10 pmol of primer, 2 mM MgCl2, 200 μM dNTP mixture, 0.5 units of Taq DNA polymerase and universal buffer. All of the reactions were performed in triplicate in an iCycler iQ System (Bio-Rad), and the thermal cycling conditions were as follows: 95 °C for 3 min; 40 cycles of 95 °C for 30s, 58 °C for 20s, and 72 °C for 30s; 72 °C for 10 min. To confirm specificity of amplification, the PCR products from each primer pair were subjected to a melting curve analysis and electrophoresis in 2 % agarose gel. In each reaction, the threshold cycle number (CT) was determined for both the target (LATS2) and the control gene (beta-actin) with the iCycler software and the mean CT for three reactions were calculated. The△CT or the CT of the housekeeping gene subtracted from the CT of the target gene was plotted for each samples.
Western blot analysis
Cells were plated into 35-mm dishes and cultured to 80 % confluence. The cells were then scraped and lysed in RIPA buffer, and cell lysates were centrifuged at 10,000 g (4 °C for 20 min). Protein concentrations were determined using Bradford reagent (Sigma) according to the manufacturer’s instructions. Equal amounts of total cellular protein were mixed with loading buffer (62.5 mM Tris–HCl, pH 6.8, 10 % glycerol, 2 % SDS, 2 % beta-mercaptoethanol and bromophenol blue), boiled for 5 min, and subjected to 10 % SDS-PAGE. Proteins were transferred to polyvinylidene difluoride membranes (Millipore, Bedford, MA). The membranes were blocked with Tris-buffered saline containing 0.05 % Tween 20 (TBST) and 5 % fat-free dry milk for 1 h at room temperature and incubated overnight with primary antibodies in TBST with 1 % bovine serum albumin. After washing with TBST, the membranes were further incubated for 1 h at room temperature with corresponding horseradish peroxidase-conjugated secondary antibody in appropriate dilution and then washed five times with the same buffer. The immunoreactive protein bands were visualized by ECL kit (Pierce). Antibodies to P65 (#4765, 1:1000), IKKs (#9966, 1:1000), IκB (#9240, 1:1000), LATS2 (#13646, 1:1000), phosphorylated IKKs (#9958, 1:1000), and IκB (#9246, 1:1000) were purchased from Cell Signaling Technology. Antibody to GAPDH (SC-365062, 1:3000) was purchased from Santa Cruz Biotechnology.
RNAi-mediated knockdown of LATS2
In our experiments, FG12 lentiviral vector, which has an independent open reading frame of green fluorescence protein (GFP), was used to produce double-stranded small interfering RNA (siRNA) to inhibit target gene expression in A549 and H23 cells. To construct the hairpin siRNA expression cassette, complementary DNA oligonucleotides for siRNA of LATS2 (si LATS2) or mutated sequence as control (si con) were synthesized, annealed, and inserted into FG12. Two LATS2 siRNA constructs were used as follows: LATS2 siRNA 1# (highlighted sequence was the complementary sequence with LATS2 mRNA), 5′-ACCG CTGGACTCTGTGAAGCTGTTCAAGAGACAGCTTCACAGAGTCCAGTTTTTTGGATCCC-3′ and 5′-TCGAGGGATCCAAAAAGAGCTGGACTCTGTGAAGCTGTCTCTTGAACAGCTTCACAGAGTCCAG-3′; LATS2 siRNA 2# (highlighted sequence was the complementary sequence with LATS2 mRNA), 5′-ACCGGGACATCCTGGCCGAGGCTTCAAGAGAGCCTCGGCCAGGATGTCCTTTTTTGGATCCC-3′ and 5′-TCGAGGGATCCAA AAAGGGACATCCTGGCCGAGGCTCTCTTGAAGCCTCGGCCAGGATGTCC-3′; si con vector (highlighted sequence was the random sequence as control that was not related to LATS2 mRNA), 5′-ACCGGTACATAGGGACGTAACGTTCAA GAGACGTTACGTCCCTATGTACCTTTTTGGATCCC-3′ and 5′-TCGAGGGATCCAAAAAGGTACATAGGGACGTAACGTCTCTTGAACGTTACGTCCCTATGTAC-3′. FG12 vector with si LATS2 or si con was transfected into HEK293T, and the virus with LATS2 siRNA or si con was harvested from culture medium. The harvested virus was purified by centrifugation at 25,000 g (4 °C, 150 min), and appropriate amounts of virus were used to infect A549 and H23 cells. After 3 days of infection, the GFP-positive cells were sorted by flow cytometry (BD Biosciences), which all stably expressed si LATS2 or si con.
Immunohistochemistry
NSCLC tissues were fixed in formalin, embedded in paraffin, and 5-μm-thick consecutive sections were cut and mounted on glass slides. After deparaffin and antigen recovery (in sodium citrate solution, pH 6.0, 20 min, 98 °C), the sections were washed thrice in 0.01 mol/l PBS (8 mmol/l Na2HPO4, 2 mmol/l NaH2PO4, and 150 mmol/l NaCl) for 5 min each, blocked for 1 h in 0.01 mol/l PBS supplemented with 0.3 % Triton X-100 and 5 % normal goat serum, followed by addition of anti-LATS2 (1:100) antibody at 4 °C overnight. After brief washes in 0.01 mol/l PBS, sections were exposed for 2 h to 0.01 mol/l PBS containing horseradish peroxidase-conjugated rabbit anti-goat IgG (1:500), followed by development with 0.003 % H2O2 and 0.03 % 3,30-diaminobenzidine in 0.05 mol/l Tris–HCl (pH 7.6). Immunohistochemistry for each sample was performed at least three separate times, and all sections were counterstained with hematoxylin.
In vivo metastasis assay
The A549-Luc stable cell line was established with G418 selection. Luciferase expression was determined by using luciferin (Xenogen) and an in vivo imaging system (Xenogen). The luciferase-expressing A549/si con cells and luciferase-expressing A549/si LATS2 cells (1 × 106 cells in 200 μl PBS) were injected into the left ventricle of the nude mice. The metastatic lesions were monitored every 8 days. Before mice were anesthetized with Forane (Abbott), an aqueous solution of luciferin (150 mg/kg intraperitoneally) was injected 10 min before imaging. The animals were placed into a light-tight chamber of the CCD camera system (Xenogen), and the photons emitted from the luciferase-expressing cells within the animal were quantified for 1 min, using the software program Living Image (Xenogen) as an overlay on Igor (Wavemetrics).
Crystal violet assay
For cell growth assay, equal number of cells were seeded in 6-well plates and cultured in medium supplemented with 10 % FBS for 7 days. Medium was changed every other day. Cell growth was stopped after 7 days in culture by removing the medium and adding 0.5 % crystal violet solution in 20 % methanol. After staining for 5 min, the fixed cells were washed with phosphate-buffered saline (PBS) and photographed.
Soft agar assay
For clonogenic assay, cells were plated into 6-well flat bottomed dishes using a two-layer soft agar system with 1.0 × 104 cells per well in a volume of 1 ml per well. After 14 days of incubation, the colonies were counted and measured. All of the experiments were done at least three times.
Boyden chamber assay
Boyden chambers (8-μm pore size polycarbonate membrane) were obtained from Neuroprobe Corporation, Bethesda, MD, USA. Cells (2 × 105) in 0.05-ml medium containing 1 % FBS were placed in the upper chamber, and the lower chamber was loaded with 0.152-ml medium containing 10 % FBS. After 8 h of incubation, cells migrated to the lower surface of filters were detected with traditional H&E staining, and five fields of each well were counted. Three wells were examined for each cell type, and the experiments were repeated for at least three times.
Results
LATS2 was down-regulated in NSCLC
Here, we first examined the messenger RNA (mRNA) level of LATS2 in 51 NSCLC clinical samples and paired normal tissues using real-time PCR analysis. Down-regulation of LATS2 mRNA level was found in about 80 % (40/51) NSCLC clinical samples compared with the matched normal tissues (Fig. 1a). These observations were further confirmed by the immunohistochemistry study, which demonstrating loss of LATS2 protein level in NSCLC tissues (Fig. 1b). In the next study, we used Western blot analysis to examine the protein level of LATS2 in NSCLC samples. Consistent with the immunohistochemistry study, decreased protein level of LATS2 was observed in six randomly chosen NSCLC tissues compared with the normal tissues (Fig. 1c). Moreover, the normal lung cells BEAS-2B and NSCLC cell lines (H23, H1299, H529, and A549) displayed differential expression of LATS2 with highest protein level of LATS2 in normal lung cell line BEAS-2B and the decreased protein level of LAST2 in a panel of NSCLC cell lines (Fig. 1d). Taken together, these observations indicated that the expression of LATS2 was down-regulated in NSCLC.

Decreased expression level of LATS2 was detected in NSCLC. Relative mRNA level of LATS2 in human NSCLC samples and normal tissues. Real-time PCR was performed on 51 NSCLC samples and 51 normal tissues. The LATS2 expression was normalized to that of beta-actin. Data was calculated from triplicates. b The protein level of LATS2 in NSCLC samples and paired normal tissues was examined by immunohistochemistry. c The protein level of LATS2 in NSCLC tissues and paired normal tissues was examined by Western blot. d The protein level of LATS2 in lung bronchus epithelial cell line BEAS-2B and NSCLC cell lines (H1299, H23, H520, and A549)
Overexpression of LATS2 inhibited cell growth and migration
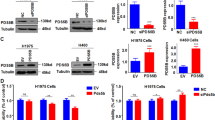
Down-regulation of LATS2 in the NSCLC samples promoted us to investigate the biological function of LATS2 in NSCLC. We first forced expression of LATS2 in H23 and H520 cells. Overexpression of LATS2 in H23 and H520 cells was confirmed by Western blot analysis (Fig. 2a). In the next study, we examined the function of LATS2 on the growth of H23 and H520 cells by crystal violet assay. Forced expression of LATS2 inhibited the growth of H23 and H520 cells dramatically (Fig. 2b). Furthermore, overexpression of LATS2 inhibited the anchorage-independent growth of H23 cells in the soft agar (Fig. 2c). Moreover, up-regulation of LATS2 impaired the motility of NSCLC cells shown by the Boyden Chamber assay (Fig. 2d). Taken together, these observations showed that forced expression of LATS2 inhibited cell growth, colony formation and cell migration.

Overexpression of LATS2 inhibited the growth and migration of H23 and H520 cells. a The H23 and H520 cells were stably transfected with either the pcDNA3.1 vector or the LATS2 expression vector. G418-resistant cells were pooled and confirmed the expression of exogenous LATS2 by Western blot analysis. b The effects of LATS2 on the growth of H23 and H520 cells were measured by crystal violet assay. c The effects of LATS2 on the anchorage-independent growth of H23 cells were measured by soft agar assay. Data shown was the representative results from three independent experiments. **P < 0.01 compared to the control group. d The effects of LATS2 on the migration of H23 and H520 cells. **P < 0.01
Knockdown of the expression of LATS2 promoted cell growth and migration
In the next study, we investigated whether endogenously expressed LAST2 played an important role in the growth and migration of NSCLC cells. Firstly, we down-regulated the basal expression of LATS2 in H23 and A549 cells using two independent siRNA sequences to exclude the off-target effects. Western blot analysis showed that these two independent siRNA sequences down-regulated the expression of LATS2 effectively (Fig. 3a). Consistent with the observations obtained from the cells overexpressing LATS2, knockdown of the expression of LATS2 promoted the growth of NSCLC cells both in liquid culture and on soft agar (Fig. 3b and c). Moreover, down-regulation of LATS2 enhanced the motility of NSCLC cells shown by the migration assay using Boyden Chamber (Fig. 3d). Collectively, these results revealed that down-regulation of LATS2 in NSCLC cells promoted cell growth and migration.

Knockdown of the expression of LATS2 promoted the growth and migration of A549 and H23 cells. a Knockdown of the expression of LATS2 in A549 and H23 cells. b Cell growth of LATS2 siRNA cells and control cells was measured by crystal violet assay. c Knockdown of the expression of LATS2 promoted the anchorage-independent growth of A549 cells measured by soft agar assay. Data shown was the representative results from three independent experiments. d Knockdown of the expression of LATS2 promoted the migration of A549 and H23 cells. *P < 0.05; **P < 0.01
LATS2 inhibited NF-κB signaling in NSCLC cells
In the next study, we investigated the underlying mechanism. As shown in Fig. 4a, overexpression of LATS2 significantly repressed the activity of an NF-κB-responsive promoter in H23 cells in a dose-dependent manner, while knockdown of the expression of LATS2 in A549 cells activated the NF-κB reporter (Fig. 4b). These observations indicated the negative regulation of NF-κB signaling by the expression of LATS2 in NSCLC cells. Moreover, overexpression of LATS2 in H23 cells decreased the accumulation of P65 in the nucleus shown by the immunostaining assay (Fig. 4c). Furthermore, up-regulation of LATS2 in H23 and H520 cells decreased the phosphorylation of IKKs, indicating LATS2 inhibited the activity of IKK IKKα/β/γ complex. Consistent with these results, the phosphorylated IκB was decreased upon the overexpression of LATS2 (Fig. 4d). Taken together, these studies revealed that LATS2 inhibited NF-κB signaling in NSCLC cells.

LATS2 regulated the activity of NF-κB signaling. a Overexpression of LATS2 down-regulated the activity of NF-κB reporter. b Knockdown of LATS2 up-regulated the activity of NF-κB reporter. c Overexpression of LATS2 inhibited the accumulation of P65 in NSCLC cells. d Overexpression of LATS2 inhibited the phosphorylation of IKKs complex and IκB
Knockdown of the expression of LATS2 promoted the metastasis of NSCLC cells in vivo
Our in vitro studies suggested that knockdown of LATS2 promoted the migration of NSCLC cells. Therefore, we evaluated whether knockdown of the expression of LATS2 in NSCLC cells could promote the metastasis of NSCLC cells in vivo by utilizing a tumor metastasis mouse model. A549-luc cells knocking down the expression of LATS2 were injected into SCID mice through tail vein. Knockdown of the expression of LATS2 increased the number of metastasis foci compared with control group (Fig. 5a), which was confirmed by the luciferase signal assay (Fig. 5b). Therefore, down-regulation of LATS2 enhanced the metastasis of NSCLC cells in vivo.

Knockdown of the expression of LATS2 promoted the metastasis of A549 cells in vivo. a Monitoring metastasis of bioluminescent A549/si con and A549/ si LATS2 cells. Images were obtained at the indicated time point after injection. b Mean photon counts of each group of mice were quantified and were displayed over time. Each point represented the mean ± SD. *P < 0.05; **P < 0.01
Discussion
Although the previous study has shown that LATS2 was down-regulated in NSCLC and induced the apoptosis of NSCLC cells through down-regulation of Bcl2 and Bcl-XL [25], the detail mechanisms remained poorly understood. In this study, we performed series of studies combining the clinical samples, cell cultures, and animal model to evaluate the function and the mechanisms of LATS2 in NSCLC tumorigenesis. Our data demonstrated that the expression of LATS2 was down-regulated in NSCLC samples. We provided evidence for the migration and metastasis suppression of LATS2 in the NSCLC cells and animal models. Importantly, we discovered that overexpression of LATS2 decreases the migration and metastasis of NSCLC cells via, in part, inhibition of NF-κB signaling.
Among the frequently activated pathways in lung cancer is the NF-κB transcription factor pathway. NF-κB activation typically occurs by nuclear translocation following inducible phosphorylation of inhibitory IκB proteins by the IKKα/β/γ (IκB kinase) complex. Importantly, recent studies in mice have defined a key role for NF-κB in KRAS-induced lung cancer [26]. Although the function of NF-κB in lung cancer has been extensively studied in mice, yet little is known about NF-κB function in human lung cancer [27, 28]. NF-kB is a transcription factor that is activated in response to various stimuli including growth factors and inflammatory molecules and is responsible for inducing gene expression associated with cell proliferation, apoptosis, angiogenesis, and migration [29]. C-myc and MMP9, two target genes of NF-κB pathway [30], have shown to promote the growth and migration of NSCLC cells [31, 32]. Also, our study showed that LATS2 inhibited the NF-κB pathway in NSCLC cells, indicating LATS2 might regulate the growth and motility of NSCLC cells through modulating the expression of NF-κB target genes.
Previous studies have shown that LATS2 was coupled to several well-established pathways, such as P53 signaling, YAP signaling [33, 34]. In this study, we have demonstrated that LATS2 coupled NF-κB pathway in NSCLC cells. Moreover, in our study, LATS2 inhibited the phosphorylation of IKKs as well as IκB and decreased the nuclear accumulation of P65, suggesting LATS2 involved in the NF-κB signal pathway upstream of IKKs complex. It has been reported that genes encoding IKKβ and IKKγ underwent frequent amplification in the tumorigenesis [35], indicating IKKs complex was not an ideal therapeutic target. Considering the fact that LATS2 cross-talked with multiple pathways, LATS2 might be a therapeutic target in the cancer treatment.
Although our data are suggestive, elucidation of the exact role and underlying mechanism of action for LATS2 in the development and progression of NSCLC will require additional investigations using LATS2 knockout mice model. Also, additional studies and validation of LATS2 as a prognostic marker in malignant NSCLC are needed.
References
Siegel R, Naishadham D, Jemal A. Cancer statistics, 2013. CA Cancer J Clin. 2013;63(1):11–30.
Anagnostou VK, Dimou AT, Botsis T, et al. Molecular classification of nonsmall cell lung cancer using a 4-protein quantitative assay. Cancer. 2012;118(6):1607–18.
Lazar V, Suo C, Orear C, et al. Integrated molecular portrait of non-small cell lung cancers. BMC Med Genom. 2012;6:53.
Goodgame B, Viswanathan A, Zoole J, et al. Risk of recurrence of resected stage I non-small cell lung cancer in elderly patients as compared with younger patients. J Thorac Oncol. 2009;4(11):1370–4.
Broet P, Camilleri-Broet S, Zhang S, et al. Prediction of clinical outcome in multiple lung cancer cohorts by integrative genomics: implications for chemotherapy selection. Cancer Res. 2009;69(3):1055–62.
Campbell JM, Lockwood WW, Buys TP, et al. Integrative genomic and gene expression analysis of chromosome 7 identified novel oncogene loci in non-small cell lung cancer. Genome. 2008;51(12):1032–9.
Kelsey CR, Fornili M, Ambrogi F, et al. Metastasis dynamics for non-small-cell lung cancer: effect of patient and tumor-related factors. Clin Lung Cancer. 2012;14(4):425–32.
Xu T, Wang W, Zhang S, Stewart RA, Yu W. Identifying tumor suppressors in genetic mosaics: the Drosophila lats gene encodes a putative protein kinase. Development. 1995;121(4):1053–63.
Tao W, Zhang S, Turenchalk GS, et al. Human homologue of the Drosophila melanogaster lats tumour suppressor modulates CDC2 activity. Nat Genet. 1999;21(2):177–81.
Hergovich A, Hemmings BA. Mammalian NDR/LATS protein kinases in hippo tumor suppressor signaling. Biofactors. 2009;35(4):338–45.
Nishioka N, Inoue K, Adachi K, et al. The Hippo signaling pathway components Lats and Yap pattern Tead4 activity to distinguish mouse trophectoderm from inner cell mass. Dev Cell. 2009;16(3):398–410.
Muller-Taubenberger A, Kastner PM, Schleicher M, Bolourani P, Weeks G. Regulation of a LATS-homolog by Ras GTPases is important for the control of cell division. BMC Cell Biol. 2012;15(1):25.
Cai H, Xu Y. The role of LPA and YAP signaling in long-term migration of human ovarian cancer cells. Cell Commun Signal. 2012;11(1):31.
Colombani J, Polesello C, Josue F, Tapon N. Dmp53 activates the Hippo pathway to promote cell death in response to DNA damage. Curr Biol. 2006;16(14):1453–8.
Xia H, Qi H, Li Y, et al. LATS1 tumor suppressor regulates G2/M transition and apoptosis. Oncogene. 2002;21(8):1233–41.
Liang Y, Li Y, Li Z, et al. Mechanism of folate deficiency-induced apoptosis in mouse embryonic stem cells: cell cycle arrest/apoptosis in G1/G0 mediated by microRNA-302a and tumor suppressor gene Lats2. Int J Biochem Cell Biol. 2012;44(11):1750–60.
St John MA, Tao W, Fei X, et al. Mice deficient of Lats1 develop soft-tissue sarcomas, ovarian tumours and pituitary dysfunction. Nat Genet. 1999;21(2):182–6.
Hergovich A. YAP-Hippo signalling downstream of leukemia inhibitory factor receptor: implications for breast cancer. Breast Cancer Res. 2012;14(6):326.
Mojca S, Vid M, Damjan G. LATS2 tumour specific mutations and down-regulation of the gene in non-small cell carcinoma. Lung Cancer. 2009;64:25–262.
Lignitto L, Arcella A, Sepe M, et al. Proteolysis of MOB1 by the ubiquitin ligase praja2 attenuates Hippo signalling and supports glioblastoma growth. Nat Commun. 2012;4:1822.
Li Y, Pei J, Xia H, Ke H, Wang H, Tao W. Lats2, a putative tumor suppressor, inhibits G1/S transition. Oncogene. 2003;22(28):4398–405.
Ishizaki K, Fujimoto J, Kumimoto H, et al. Frequent polymorphic changes but rare tumor specific mutations of the LATS2 gene on 13q11-12 in esophageal squamous cell carcinoma. Int J Oncol. 2002;21(5):1053–7.
Li W, Wang L, Katoh H, Liu R, Zheng P, Liu Y. Identification of a tumor suppressor relay between the FOXP3 and the Hippo pathways in breast and prostate cancers. Cancer Res. 2012;71(6):2162–71.
Lin XY, Zhang XP, Wu JH, Qiu XS, Wang EH. Expression of LATS1 contributes to good prognosis and can negatively regulate YAP oncoprotein in non-small-cell lung cancer. Tumour Biol. 2012;35(7):6435–43.
Ke H, Pei J, Ni Z, et al. Putative tumor suppressor Lats2 induces apoptosis through downregulation of Bcl-2 and Bcl-x(L). Exp Cell Res. 2004;298(2):329–38.
Starczynowski DT, Lockwood WW, Delehouzee S, et al. TRAF6 is an amplified oncogene bridging the RAS and NF-kappaB pathways in human lung cancer. J Clin Invest. 2012;121(10):4095–105.
Nair VS, Gevaert O, Davidzon G, Plevritis SK, West R. NF-kappaB protein expression associates with (18)F-FDG PET tumor uptake in non-small cell lung cancer: a radiogenomics validation study to understand tumor metabolism. Lung Cancer. 2012;83(2):189–96.
Wang LH, Yang JY, Yang SN, et al. Suppression of NF-kappaB signaling and P-glycoprotein function by gambogic acid synergistically potentiates adriamycin-induced apoptosis in lung cancer. Curr Cancer Drug Targets. 2012;14(1):91–103.
Samykutty A, Shetty AV, Dakshinamoorthy G, et al. Piperine, a bioactive component of pepper spice exerts therapeutic effects on androgen dependent and androgen independent prostate cancer cells. PLoS One. 2012;8(6):e65889.
Suh J, Rabson AB. NF-kappaB activation in human prostate cancer: important mediator or epiphenomenon? J Cell Biochem. 2009;91(1):100–17.
Chen X, Su Y, Fingleton B, et al. Increased plasma MMP9 in integrin alpha1-null mice enhances lung metastasis of colon carcinoma cells. Int J Cancer. 2005;116(1):52–61.
Little CD, Nau MM, Carney DN, Gazdar AF, Minna JD. Amplification and expression of the c-myc oncogene in human lung cancer cell lines. Nature. 1983;306(5939):194–6.
Aylon Y, Michael D, Shmueli A, Yabuta N, Nojima H, Oren M. A positive feedback loop between the p53 and Lats2 tumor suppressors prevents tetraploidization. Genes Dev. 2006;20(19):2687–700.
Zhang J, Smolen GA, Haber DA. Negative regulation of YAP by LATS1 underscores evolutionary conservation of the Drosophila Hippo pathway. Cancer Res. 2008;68(8):2789–94.
Guang W, Ding H, Czinn SJ, Kim KC, Blanchard TG, Lillehoj EP. Muc1 cell surface mucin attenuates epithelial inflammation in response to a common mucosal pathogen. J Biol Chem. 2012;285(27):20547–57.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (81201840), the Natural Science Foundation of Shanghai (13ZR1461300), the Health Bureau Foundation of Shanghai (20124Y152), and Chenxing Young Scholarship of Shanghai Jiaotong University.
Conflicts of interest
None
Author information
Authors and Affiliations
Corresponding author
Additional information
Feng Yao and Hongcheng Liu contributed equally to this work.
Rights and permissions
About this article

Cite this article
Yao, F., Liu, H., Li, Z. et al. Down-regulation of LATS2 in non-small cell lung cancer promoted the growth and motility of cancer cells. Tumor Biol. 36, 2049–2057 (2015). https://doi.org/10.1007/s13277-014-2812-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13277-014-2812-1