
Abstract
Stormwater ponds (SP) are increasingly used for water management along roads and in urban environments. How these infrastructures compare to natural wetlands in terms of biodiversity remains unclear, however. Studies to date have evaluated the subject in general terms, without considering the different zones in SP and wetlands (from aquatic, at the pond edge, to terrestrial, at the upper bank) or other local and regional factors. In this project, we aimed to compare the taxonomic diversity and composition of plant communities established in four different zones of SP with that in either roadside or remote natural wetlands. We also evaluated the effect of various local and regional factors on those communities. Our results show that, globally, the species composition of the lower, wetter zones was similar between SP and natural wetlands, especially roadside wetlands, while higher, drier zones showed significant differentiation. Factors explaining observed differences between SP and both roadside and remote natural wetlands were water level fluctuations, road proximity, slope, and age of the SP. Stormwater ponds also exhibited lower beta diversity than both types of wetlands. Nonetheless, our study suggests that with some modifications in their design, SP have the potential to harbour more wetland plant communities.
Résumé
Les bassins de rétention (BR) sont de plus en plus utilisés pour la gestion de l’eau le long des autoroutes et des zones urbaines. Cependant, nous ne savons pas comment ces infrastructures se comparent aux milieux humides (MH) naturels en termes de biodiversité. Les études sur le sujet font une évaluation générale qui ne tient pas compte des différentes zones des BR et des MH (du plus humide en bordure de mare, jusqu’au plus terrestre en haut de talus), ou encore des facteurs locaux ou régionaux. Ce projet vise à comparer la diversité taxonomique et la composition des communautés végétales établies dans quatre zones différentes des BR avec des milieux humides naturels, soit de bord de route, soi isolés. Nous avons également évalué l’effet de plusieurs facteurs locaux ou régionaux sur ces communautés. Nos résultats montrent que globalement, la composition spécifique des zones plus humides était similaire entre les BR et les MH naturels, en particulier ceux situés près d’une route, alors que les zones les plus hautes étaient significativement différentes entre elles. Les facteurs expliquant les différences observées entre les BR et les MH naturels de bord de route et isolés étaient les fluctuations du niveau d’eau, la proximité de la route, la pente et l’âge des BR. De plus, les BR montraient une diversité bêta plus faible que les deux types de milieux humides. Notre étude suggère néanmoins que, moyennant certaines modifications dans leur conception, les BR ont le potentiel d’abriter davantage de communautés végétales de MH.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Wetlands are one of the most biodiverse habitats on Earth (Costanza et al. 1997; Dodds et al. 2008), displaying crucial ecological functions such as water level regulation, water filtration, and carbon sequestration (Temmink et al. 2022). However, these unique ecosystems are rapidly declining globally (Zedler and Kercher 2005; Davidson 2014; Dixon et al. 2016) due to land-use changes, with global net losses estimated at 21% or more since 1700 (Fluet-Chouinard et al. 2023). The St. Lawrence River basin is the fourth most disturbed one worldwide, with losses estimated to reach slightly more than 50% in the last two centuries, the most drastic changes happening in the last decades (Poulin and Pellerin 2013; Fluet-Chouinard et al. 2023). To manage the surplus surface water runoff resulting from both the loss of wetlands and the increase in impervious surfaces, the construction of stormwater ponds (SP) is becoming widespread in urban and peri-urban environments (Tixier et al. 2011; Birch et al. 2022).
In addition to their primary functions of water regulation and pollutant reduction (Starzec et al. 2005; Mitchell et al. 2007), SP often constitute habitats for plants and wildlife, (Marsalek et al. 2005), increasing species connectivity in human-dominated landscapes (Birch et al. 2023), although they are often of poor quality due to pollution (McKercher et al. 2024). In turn, the presence of plants further contributes to the ecosystem functioning of SP, increasing the services provided such as pollutant removal (McKercher et al. 2024) and peak flow attenuation through increased evapotranspiration (Natarajan and Davis 2015). However, to what extent plant communities in these infrastructures mirror those of wetlands (i.e., natural wetlands in similar environmental contexts) remains to be explored.
Studies assessing SP plant diversity show that SP are less diverse than natural wetlands (Reinartz and Warne 1993; Kuntz et al. 2018). Although some SP are planted or seeded with a set of species following construction, they are often rapidly colonized by invasive species that tend to exclude others, including native wetland species. In some cases, total species richness may be comparable between SP and wetlands; however, upland as well as invasive wetland species prevalent in SP such as Typha angustifolia and Phragmites australis subsp. Australis tend to replace native, wetland obligate species (Rooney et al. 2015). Thus, the richness of wetland obligate and facultative species has been found to be lower in SP than in natural wetlands, leading to lower ecological value (Rooney et al. 2015; Perron and Pick 2020).
Stormwater ponds are often required by urban planning, especially when there is a loss of natural, permeable surfaces (Rivard et al. 2014). Roads are an example of such infrastructure for which the implementation of SP is mandatory in order to decrease the risk of flooding and avoid contaminant dispersion in the immediate environment (Birch et al. 2006; Hwang et al. 2019). However, since SP are often built close to roads, their plant communities are often exposed to high levels of contaminants (nitrogen, metals, hydrocarbons, de-icing salts, etc.) that affect plant colonization, survival, and reproduction (Truscott et al. 2005; Akbar et al. 2006; Gülser and Erdogan 2008; Skultety and Matthews 2018). Roads have also been shown to have a detrimental impact on the plant community composition and diversity of natural wetlands (Findlay and Houlahan 1997; Findlay and Bourdages 2000; Avon et al. 2010), especially within 50 m of either side (Fakayode and Olu-Owolabi 2003; Gülser and Erdogan 2008; Khalid et al. 2018).
Evaluating restored or constructed sites can be done by using unmanaged reference ecosystems as a standard of comparison (SERI 2004; Moreno-Mateos et al. 2012). This makes it possible to determine whether the attributes of the former (e.g., composition, diversity, richness, strata, functions) follow the expected trajectory towards restoration or construction objectives (Zedler and Callaway 1999; Campbell et al. 2002; Balcombe et al. 2005; Matthews et al. 2009). In this study, and even though SP are not built to emulate wetlands (as is the case with constructed wetlands), we used natural wetlands to look for similarities with SP in terms of plant community composition and to estimate whether SP could be refugia for wetland plant species, despite their primary function of water management.
In this study, we aimed to determine whether roadside SP and natural wetlands in a North American context have similar plant communities. We wanted to evaluate if and how SP could be modified in order to achieve certain ecological goals, such as harbouring wetland plant communities. We focussed on the different zones often found within such constructed environments: aquatic, wet littoral, dry littoral, and upper bank, comparing roadside SP to both roadside and remote natural wetlands. We explored the influence of local and regional factors on plant community diversity and composition. We predicted that (1) SP show higher alpha diversity and lower beta diversity than natural wetlands, due to the presence of drier, more terrestrial zones; and that (2) SP resemble roadside wetlands more than remote ones.
Methodology
Study area
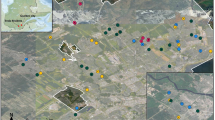
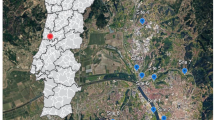
We conducted our study along four roads located in southeastern Québec, Canada (Fig. 1) classified as intra-provincial highways with two to four lanes, depending on the segment. The annual average daily traffic ranged between 7000 cars (Highways 175 and 85) and 15 000 cars (Highways 367 and 73; MTQ 2018). The study area spread both north and south of the St. Lawrence River and encompassed four bioclimatic domains as classified by the provincial forest agency (balsam fir-yellow birch, sugar maple-yellow birch, sugar maple-basswood, and balsam fir-white birch; MRNF 2022). This ecological classification of Québec territory uses plant communities to define homogenous regions in terms of climate and precipitation. For sites located north of the St. Lawrence River (Highways 175 and 367; n = 36), mean annual temperatures vary from 0.5 °C to 4.7 °C (depending on altitude), with mean annual precipitation of 1288 mm, of which 24% falls as snow. For sites located south of the St. Lawrence River, along the Highway 73 area (n = 12), the mean annual temperature is 4.5 °C, with mean annual precipitation of 1117 mm, of which 20% falls as snow. Further east, for sites along Highway 85 (n = 12), the mean annual temperature is 3.4 °C, with mean annual precipitation of 1029 mm, of which 26% falls as snow (MELCC 2021).

Location of the study area and the 60 study sites in southeastern Québec (Canada). Studied roads are indicated by numbers
Site Selection
We first selected 174 stormwater pond (SP) sites from a database provided by the provincial transport agency, the Ministère des transports du QuébecFootnote 1. We only included permanent ponds (i.e., filled with water year-round). We discarded the more recent SP (i.e., those built in the last three years), to ensure sufficient colonization by the local flora, as well as those located in dense constructed areas of urban centers, due to the absence of urban natural wetlands. From the remaining SP (28), we selected those situated at least 500 m apart from each other, to limit spatial correlation (Houlahan et al. 2006). A total of 20 SP met these criteria, ranging in size from 0.14 to 1 ha (mean area: 0.46 ha). All 20 selected SP consisted of a circular or elongated body of open water surrounded by steep slopes (7 to 26%). Their bottom substrate consisted mainly of rockfill without an impervious membrane, and their shoreline had been enriched with 10 to 40 cm of topsoil of unknown origin. Banks of all 20 SP had been systematically hydroseeded with a mix of grasses, and some sites had been planted with herbs and shrubs (16 out of 18 sites for which information was available; see Appendix A for list of species).
We paired two natural wetlands with each SP for comparison purposes, using a database of natural wetlands provided by Ducks Unlimited and the provincial environment agency (Ministère de l’Environnement et de la Lutte aux Changements Climatiques), completing this selection with a visual analysis of satellite images (ESRI 2020) to identify ponds, which were then validated on site. The selected sites were small ponds: marshes surrounding an open-water area. We selected the first one to be situated 50 m or less from a paved road (hereafter roadside wetlands; RoW), and the other within 50 m to 1 km from a paved road (hereafter remote wetlands; ReW). Including both RoW and ReW allowed us to account for the effect of road disturbance on plant community composition and turnover between SP and natural wetlands. We selected wetlands ranging in size from 0.04 to 0.78 ha (mean area: 0.32 ha), an area comparable to the selected SP in order to avoid species area bias. Overall, we sampled 60 sites: 20 from each type (SP, RoW, and ReW).
Vegetation Sampling
We conducted plant surveys from June to August 2019, except for eight sites (located throughout the study region) that were visited at the end of July 2020. We surveyed sites once, except for those surveyed before mid-July of 2019, which were visited a second time during the season to validate species identification, since most species had not yet flowered at the first visit. We divided the sites into three sections as homogenous as possible in terms of vegetation and noted the proportion of the site represented by each section (Fig. 2). We placed a transect in the center of each section, perpendicular to the shore and extending from the aquatic zone to the upper bank. Along each transect, we sampled one plot from each of the four delineated zones, namely: aquatic, wet littoral, dry littoral, and upper bank. We considered the aquatic zone to be the lowest vegetation growing in the water (maximum water depth: 1 m), the wet littoral, the transitional community found right below the water surface, the dry littoral, the transitional community found right above the water surface, and the upper bank, the community extending to the upper bank or the forest edge. We used rectangular plots of 5 m2, and adapted length and width to the spatial structure of each zone (plots measured either 1 m x 5 m or 2 m x 2,5 m), placing them in representative areas of each zone. Considering that the water level fluctuates throughout the season (though not much, with an average of 19 cm), we avoided surveying the transitional communities too close to the water line, using the narrower plot dimensions to survey the wet and dry littorals (Fig. 2). This way, we could be consistent with zone delineation from one site to another. We identified all species within the plots and visually estimated their cover using six classes: <1%, 1–5%, 6–25%, 26–50%, 51–75%, and 76–100%. For each zone on every site, we calculated mean species cover by using the median of each class, weighted using the estimated spatial proportion of each of the three sections. For site-level analyses, we averaged these four weighted mean cover values together.

Schematic view of the sampling design. Along each transect, the four plots represent Q1: aquatic zone, Q2: wet littoral, Q3: dry littoral, and Q4: upper bank
Since SP upper banks were mowed during the summer, it was sometimes impossible to identify some grass species. In such cases, we identified the observed plants as “grass”, encompassing the three species used for hydroseeding: Agrostis gigantea, Festuca rubra, and Poa pratensis. Consequently, we pooled all occurrences of these three species together in this category (“grass”) for all sites.
Local and Regional Factors
We measured the slope (%) of each transect with a clinometer and then averaged the values for the entire site. We also measured water conductivity (µS/cm) and pH on a water sample composed of three sub-samples, each taken in a different location across the pond surface, to reduce variability in physicochemical properties due to water inflow. We assessed landscape composition within a 1 km radius using shapefiles with delineated polygons for forest, crops, and wetlands in ArcGIS software (ESRI 2020). Geographical data was provided by the provincial forest agency (forest areas; MRNF 2017), Financière Agricole du Québec (crop areas; FADQ 2020), as well as Ducks Unlimited and the provincial environment agency (wetland areas; MDDEP 2011; CIC and MELCC 2020). Variables used for analyses are presented in Table 1.
We obtained information about SP construction from the provincial transport agency, but some data were missing. Available data included the age (available for n = 20 SP), pond bottom substrate (n = 11), organic soil thickness at the shoreline (n = 13), and a list of planted or sown species (n = 18), which consisted of herbaceous and shrub species ranging from wetland obligate to upland status. During SP construction, planting had been carried out on a small scale, covering only small portions of the SP and apart from the number of species planted, more precise information (e.g., the number and location of plants) was unavailable. As the information on pond substrate was missing for 45% of the SP, this factor could not be included in our analyses. Finally, we obtained water level fluctuation data using a time-lapse camera placed in front of a ruler installed in the open water on each site, taking a photograph every 24 h between May 22 and September 9, 2019. Water level fluctuation corresponded to the difference between maximum and minimum water levels observed.
Statistical Analyses
Taxonomic Richness
We computed total taxonomic richness at each site as the number of distinct taxa found in all plots. Similarly, we assessed taxonomic richness for each zone individually (aquatic, wet littoral, dry littoral, and upper bank) on each site. Performing an ANOVA on a linear model, we tested whether taxonomic richness differed between site types (SP, RoW, ReW), first, taking SP as a whole and second, testing each specific zone separately. We verified that model residuals followed a normal distribution and displayed homogeneous variance (Shapiro-Wilk and Bartlett’s tests). When we obtained significant results, we conducted post hoc multiple comparisons using least significant difference (LSD) tests. We adjusted p-values using Holm’s correction for multiple testing.
In addition, we evaluated whether the proportion of obligate wetland, facultative wetland, non-native, and invasive taxa (% of the total cover) differed between site types using Kruskal-Wallis nonparametric tests and performed multiple comparisons with Wilcoxon tests. We analyzed non-native and invasive species separately, but all invasives were also non-native species. Wetland indicator status followed Bazoge et al. (2014) and the Plant List of Attributes, Names, Taxonomy, and Symbols (PLANTS) Database (USDA and NRCS 2020), while species origin in Québec (native or non-native) followed the Database of Vascular Plants of Canada (VASCAN; Brouillet et al. 2020).
Beta Diversity and Species Composition
We calculated beta diversity for each type of site (SP, RoW, ReW) using the mean Hellinger distance of the sites to their respective group centroid. Specifically, we computed three site-by-species matrices using mean cover data and built site-by-site distance matrices using the Hellinger distance. We also analyzed differences in beta diversity between site types, using a distance-based permutation test for homogeneity of multivariate dispersions on a site-by-site distance matrix including all 60 sites (PERMDISP; Anderson et al. 2006) with 9 999 permutations. Finally, we conducted post hoc multiple comparisons using LSD tests. We did this for sites as a whole, as well as for each of the four zones separately.
To detect a shift in species composition, we tested for location differences between centroids obtained with PERMDISP, using PERMANOVA with pseudo-F ratios (9 999 permutations). We made pairwise comparisons with Bonferroni corrections on site types to identify which ones differed. Because this test is sensitive to differences in multivariate dispersions, we used data visualization to support the interpretation of the statistical test. We illustrated the differences in multivariate dispersion and composition in principal coordinates analysis ordinations (PCoA).
Impact of Local and Regional Factors on Plant Community Composition
To assess differences in local variables between site types, we analyzed the slope, pH, and conductivity data by performing an analysis of variance (ANOVA) on a linear model. We then determined the factors associated with vegetation composition through a redundancy analysis (RDA) of the vegetation data constrained by all local and regional (landscape composition) variables, for all three site types. For the RDA, we removed rare species from the vegetation matrix (< 20%; 79 taxa remained) and applied Hellinger’s transformation. We standardized all explanatory variables, and assessed the significance of the model using a permutation test with 9 999 randomized runs (Legendre and Legendre 2012).
To evaluate the specific influence of SP design on taxonomic composition, we only used data from the 20 SP for further analyses. Design variables included slope, water level fluctuation, SP age, thickness of organic soil, and number of sown species (Table 1). We calculated the mean ecological distance of each zone of every SP to the corresponding zones in the natural wetlands, RoW and ReW combined, using the site-by-site distance matrix from the beta diversity analyses. We then calculated the correlations and associated adjusted R2 values between these distances and each of the aforementioned design variable.
We performed all statistical analyses using R 4.0.1 (R Core Team 2019). We used the Anova function from the “car” package (Fox et al. 2020) for analyses of variance. We used the shapiro.test function for Shapiro-Wilk tests, bartlett.test for Bartlett tests, and wilcox.test for Wilcoxon tests, all from the “stats” package (R Core Team 2019). We conducted LSD tests using the LSD.test function and Kruskal-Wallis tests with the Kruskal function, both from the “agricolae” package (Mendiburu 2020). We obtained Hellinger distances with the vegdist function, did multivariate dispersion analyses with betadisper, measured centroid position with the adonis2 function, did Hellinger transformations and standardizations with decostand, redundancy analyses with rda, and permutational tests with the anova.cca function, all from the “vegan” package (Oksanen et al. 2020).
Results
Taxonomic Richness
We identified a total of 319 taxa on the 60 study sites: 207 in stormwater ponds (SP), 223 in roadside wetlands (RoW), and 200 in remote wetlands (ReW). Of these 319 taxa, 97 were wetland obligate and 79 were wetland facultative species, together representing 55% of the sampled flora. Moreover, 69 taxa (22%) were non-native species, among which 20 (6%) were also considered invasive (Appendix B). In SP, four of the ten most common taxa were non-native, none were invasive, and three were wetland indicators (Table 2). However, in both types of natural wetlands, the ten most frequent taxa were all native and with the exception of Abies balsamea, Fragaria virginiana, and Euthamia graminifolia, were wetland indicator species.
The mean taxa richness of SP sites was significantly higher than that of both types of natural wetlands, between which it was similar (Fig. 3a). This higher richness of SP was mainly linked to the high richness of the SP dry littoral zone (Fig. 3b). For other zones, taxa richness was similar between site types (Fig. 3b).

Species richness for all zones combined (a) or presented separately (b) for the three types of sites (SP, RoW and ReW). Mean (diamond), median (line), 25–75% quartiles (boxes) and ranges (whiskers) are provided. The model was tested with ANOVA. Means with different letters (within a same comparison group) differ significantly (LSD test, α < 0.05). SP: Stormwater ponds, RoW: Roadside wetlands; ReW: Remote wetlands
Beta Diversity and Species Composition
Taxonomic beta diversity differed significantly between SP and natural wetlands (F = 15.19; p < 0.001; Fig. 4). It was statistically higher in both types of natural wetlands, meaning that plant communities of SP were more homogeneous from site to site than those of natural wetlands. At the zone level, beta diversity was similar between wetland types for the aquatic zone but, again, was lower in SP than in natural wetlands for the three others (Fig. 5).
Taxonomic composition (i.e., distances between centroids; see Methods section) also differed significantly between SP and natural wetlands (F = 4.424; p < 0.001; Fig. 4). The two types of natural wetlands had similar species composition (overlapping ellipses and proximity of their two centroids). At the zone level, plant species composition of the aquatic zone of SP was similar to that of natural wetlands (similar centroids for the three groups; Fig. 5). Wet littoral species composition of the SP was significantly different from that of ReW, but not of RoW, which had an intermediate species composition. For the dry littoral and the upper bank, SP were clearly different from both RoW and ReW, which in turn had similar dry littoral but different upper bank plant species composition.

Taxonomic beta diversity. Multivariate dispersion of species composition for the three types of sites (SP, RoW and ReW). Beta diversity is measured as the average distance of sites to their group centroid, here represented on the first two axes of a PCoA and using a boxplot (mean, median and quartiles) of the sites-to-centroid distance. Ellipses illustrate standard deviation. Model was tested with permutation tests. Letters are used to indicate significant differences in species composition (replacement) and mean distance to group centroid (turnover; LSD test; α ≤ 0.05)
Obligate species cover was significantly lower in SP compared to RoW, but was similar to that of ReW (Fig. 6). Wetland facultative cover was significantly lower in SP than in both types of natural wetlands. Non-native and invasive species covers were both higher in SP than in natural wetlands. Even though the difference is not significant, RoW had mean non-native and invasive species covers at least double those of ReW.

Plant beta diversity per zone for the three types of sites. PCoAs represent species composition, with ellipses representing the standard deviation for each site type. Boxplots (mean, median and quartiles) represent site distance to their group centroid. Letters are used to indicate significant differences in species composition (replacement) and mean distance to group centroid (turnover) of the three site types (LSD test; α ≤ 0.05). SP: Stormwater ponds, RoW: Roadside wetlands, ReW: Remote wetlands

Proportion of total cover of wetland obligate, wetland facultative, non-native, and invasive species for all study sites. Mean richness (diamond), median (line), 25–75% quartiles (boxes) and ranges (whiskers) are provided. The model was tested with Kruskal-Wallis test. Means with different letters differ significantly (Wilcoxon’s test). SP: Stormwater ponds, RoW: Roadside wetlands; ReW: Remote wetlands
Impact of Local and Regional Factors on Plant Community Composition
Variables included in the RDA analyses accounted for 21% of the total variance (Fig. 7). The first axis was strongly associated with local variables and clearly separated SP from natural wetlands. Stormwater ponds were characterized by higher pH and conductivity as well as steeper slopes (Fig. 8) and were associated with species identified as “grass” (Agrostis gigantea, Festuca rubra, and/or Poa pratensis), as well as Vicia cracca and Trifolium hybridum (Fig. 7). The second axis of the RDA was mostly related to landscape composition, and while significant, did not give any clear pattern. This axis only slightly separates RoW and ReW, therefore explaining a limited portion of the total variance.

Redundancy analysis (RDA) for the effects of local (slope, pH, conductivity, water level fluctuation) and regional factors (cover of forest, crops, wetlands) on species composition (adjusted R2 = 0.12). Grass: Agrostis gigantea, Festuca rubra and/or Poa pratensis

Slope (%), pH values, and conductivity (µS/cm) for study sites (SP, RoW and ReW). Mean (diamond), median (line), 25–75% quartiles (boxes) and range (whiskers) are provided. The model was tested with ANOVA. Means with different letters (within a same comparison group) differ significantly (LSD test, α < 0.05). SP: Stormwater ponds, RoW: Roadside wetlands; ReW: Remote wetlands
Mean ecological distance of SP to natural wetlands was correlated with three design variables, namely water level fluctuation, slope, and age (Fig. 9). Water level fluctuation was positively correlated to ecological distance in the aquatic zone, meaning that SP with the least variation in water level across the season had aquatic communities more similar to those of natural wetlands. The slope was positively correlated to ecological distance in the wet littoral, indicating that gentler slopes supported wet littoral communities closer to those of natural wetlands. Finally, the age of the SP was negatively correlated to ecological distance in the dry littoral, indicating that dry littoral communities of SP became more similar to those of natural wetlands with time.

Correlations between mean Hellinger distance of SP to natural wetlands and design variables (water level fluctuation, slope and SP age)
Discussion
This study was designed to assess whether stormwater ponds (SP) are similar to wetlands in terms of taxonomic alpha and beta diversity, as well as community composition, and to evaluate the impact of local and regional factors on SP plant communities. Results revealed similarities between SP and wetlands, especially in the lower, wetter zones. Moreover, water level fluctuation, slope, and age of SP influenced SP plant community composition.
Examining SP at a finer scale, with emphasis on specific zones, revealed some similarities with wetlands, whereas previous studies mostly found differences (Reinartz and Warne 1993; Rooney et al. 2015; Kuntz et al. 2018; Perron and Pick 2020). Of the four zones identified in this study (aquatic, wet littoral, dry littoral, and upper bank), the two wetter zones in SP exhibited a species composition similar to that in natural wetlands, especially roadside wetlands. The more terrestrial zones were, however, quite different in terms of richness and composition. Stormwater ponds surveyed in this study could therefore be considered as ponds surrounded by a limited marsh area and an extensive bank with more terrestrial characteristics. They can thus be considered to be of ecological interest, since both natural and man-made ponds have a strong potential for conservation (Céréghino et al. 2014).
The similarities between the aquatic and wet littoral zones in SP and natural wetlands were correlated with two factors, water level fluctuations and slope. Higher fluctuations of the water level in SP increased their mean ecological distance from natural wetlands. Extended flooding can influence communities by two contrasting mechanisms: either by reducing the prevalence of upland species and promoting the diversity of wetland plants (Gathman et al. 2005; Drinkard et al. 2011), or by causing the loss of specialist wetland plants that require a specific hydroperiod or an intermediate water depth (Greenway et al. 2007). This provides space for colonization by invasive wetland plants that can withstand these fluctuations (Wei and Chow-Fraser 2006; Zhang et al. 2015; Sun et al. 2019; Bedell et al. 2021). The invasive P. australis subsp. australis, not yet widespread in the study region, can indeed withstand extreme fluctuations in water level (up to 120 cm; Hanslin and Saebo 2017). In the studied SP, water levels were relatively stable: mean water level fluctuation (10.8 cm) was comparable to that of natural wetlands (9.2 cm). On the other hand, slope was strongly correlated to ecological distance between SP and natural wetlands in the wet littoral. Therefore, the steep slopes contributed to the differentiation of SP and natural wetlands. The slopes of SP were indeed much steeper (7 to 26%, 18% on average) than those of natural wetlands (0 to 17%, 6% on average). While weaker slopes have sometimes been thought to favor invasive species establishment, no such relationship between invasive plants and slope was observed in our study (results not shown). This could also be due to the study region, where P. australis subsp. australis invasions are not widespread (Jodoin et al. 2008).
The results concerning water level fluctuation and slope highlight the importance of SP design for the establishment of wetland plant communities. Design choices made before SP construction have a direct impact on plant communities that establish on its shores. Biodiversity goals could be achieved for SP if their design were modified, but this may be antagonistic to engineering objectives. Indeed, stormwater ponds are usually designed to have maximum water holding capacity for a given area, which calls for steep slopes (Rivard et al. 2014). The SP that were visited for this study had slopes that were actually steeper than what is recommended in the provincial guidelines (6%; Rivard et al. 2014), which makes it possible to store more water in a given area, but minimizes the potential for harbouring wetland plants. Therefore, it seems a compromise between water-holding capacity and habitat for wetland plant communities is reachable.
In addition to differences in plant communities attributable to design factors, proximity to a road also exerted an influence on taxonomic composition of SP, more specifically in the wet littoral zone. This was underlined by the fact that SP plant communities were similar to those of roadside wetlands, but not remote wetlands. Roads can impact wetlands through pollution as well as non-native and/or invasive species propagule pressure. Here, it seems that stormwater ponds were more vulnerable to the establishment of such species than roadside wetlands. Soil disturbance and low plant biomass during SP construction and time of establishment are known to favor invasion by non-native species (Davis and Froend 1999; Pearson et al. 2018). Moreover, many non-native species produce more seeds, and do so earlier in the season than native species, causing strong priority effects (i.e., outcompeting species whose seeds arrive later; Wilsey et al. 2015). This might explain their higher prevalence in such disturbed environments as SP. However, we must note that P. australis subsp. Australis produces seeds in late autumn and reproduces mostly vegetatively in the study region (Gervais et al. 1993; but see also Brisson et al. 2010).
The plant communities of the dry littoral and upper bank of SP were clearly different from those of the natural wetlands. Most of the dry area of SP was a terrestrial habitat unsuitable for wetland plant communities. However, there was a correlation between the age of the SP and the dry littoral plant communities: with time, dry littoral plant communities from SP became more similar to those of natural wetlands. Considering that it may take decades for plant communities to completely regenerate after restoration (Lebrija-Trejos and Bongers 2008; Bullock et al. 2011; Moreno-Mateos et al. 2012), succession processes are likely still taking place. Compared to SP, natural wetlands showed dry littoral communities with a higher cover of shrubs (37 ± 5 vs. 22 ± 5%) and trees (5 ± 1 vs. 3 ± 2%), and a lower cover of vines (3 ± 1 vs. 15 ± 3%) and forbs (48 ± 6 vs. 113 ± 12%).
The drier zones of SP were characterized by grass species, sown to rapidly cover the soil and stabilize the banks, but still exhibited a higher proportion of non-native, as well as invasive plants. Planting shrubs or trees could support the establishment of such taller plant species, which are found in natural wetlands, and bring heterogeneity, offering different microhabitats and eventually increasing beta diversity, as well as controlling for invasives (Lelong et al. 2009). The establishment of tall species and the control of invasive species could also be achieved by reducing mowing operations (Baoyin et al. 2015; Wigginton and Meyerson 2018). Rather than being vegetated mainly with grass, this diversification of plant structure of the upper zones could provide ecosystem services that extend beyond those of wetlands. As such, the two higher zones could for example support pollinators through the introduction of flowering plants. They could also offer further connectivity for the fauna (see for example Smith et al. 2019).
This study highlights the homogeneity of SP in terms of taxonomic composition, which mirrors findings in other studies (Reinartz and Warne 1993; Rooney et al. 2015; Kuntz et al. 2018; Perron and Pick 2020; Sinclair et al. 2020). Sinclair et al. (2020) identified two main explanatory factors, namely the choice of only a few similar species for pond construction and limited dispersion ability of species in an urban context. In our case, the homogeneity of SP could also be due to other factors, such as their recent construction, design homogeneity, high maintenance, and propagule pressure by roads.
This calls for a set of actions, the first of which could be the differentiation of SP through a variety of designs. Instead of a single “ideal” design, we should aim to develop a catalogue of options that could be tailored to specific regional needs in terms of plant diversity, wildlife habitat, and ecosystem services related to water. Native vegetation should be established as fast as possible in order to pre-empt invasion by non-native species. Other strategies to increase richness include seeding native species following SP construction (Reinartz and Warne 1993; Balcombe et al. 2005; Mitsch et al. 2014), which would also limit colonization by unwanted species that use roads as dispersal corridors (Reinartz and Warne 1993), as well as increasing microtopographic heterogeneity by disking the soil (Moser et al. 2007). Finally, zones that were found to be equivalent to natural wetlands were the two aquatic ones, which have a limited extent due to the strong slopes (Fig. 10). As such, we suggest that the perimeter of the littoral be increased by favoring more complex shorelines, making SP with higher perimeter-area ratios. Wetland perimeter length, more so than area, has an impact on species richness (Reinartz and Warne 1993).

An illustration of the transition zone in a SP (a) and a remote natural wetland (b). The transition zone is much narrower along the steep slope of the SP, and much wider along the almost null slope of the wetland
Conclusion
Our study characterized SP to a finer scale than usually reported in the literature: the lower zones in SP are indeed similar in some ways to those in natural wetlands. Factors that could explain differences between the two types of sites are linked with design characteristics inherent to the primary function of SP. Moreover, we identified management improvements to foster the establishment of more natural communities. Although SP show extensive terrestrial banks, the lower zones should merit increased consideration by policy makers if they wish to increase their biotic value. Keeping water level fluctuations to a minimum, lowering the slopes, enhancing the upper bank by promoting heterogeneity (by planting trees and shrubs) are the key strategies that we identified. Complexifying the shoreline, keeping maintenance to a minimum, as well as promoting a diversity of SP designs, would be further means of enhancing beta diversity between SP.
Our study brings nuances to the existing literature: even if the surveyed SP were not comparable as a whole to remote natural wetlands, they did share similarities, having the potential to harbor wetland plant communities without being dominated by invasive or non-native species. These findings should encourage further research into how these green infrastructures could be valued, especially considering their ever-growing presence, and their importance in urban landscapes. It is important to remember, however, that this study was conducted outside of dense urban areas, and we believe that the higher disturbance experienced by urban SP could impact species composition of their wetter zones.
Data Availability
The datasets generated during the current study are available from the corresponding author on reasonable request.
References
Akbar KF, Headley ADD, Hale WHG, Athar M (2006) A comparative study of de-icing salts (sodium chloride and calcium magnesium acetate) on the growth of some roadside plants of England. J Appl SCI Environ Manag 10:67–71
Anderson MJ, Ellingsen KE, McArdle BH (2006) Multivariate dispersion as a measure of beta diversity. Ecol Lett 9:683–693. https://doi.org/10.1111/j.1461-0248.2006.00926.x
Avon C, Bergès L, Dumas Y, Dupouey JL (2010) Does the effect of forest roads extend a few meters or more into the adjacent forest? A study on understory plant diversity in managed oak stands. Ecol Manage 259:1546–1555. https://doi.org/10.1016/j.foreco.2010.01.031
Balcombe CK, Anderson JT, Fortney RH, Rentch JS, Grafton WN, Kordek WS (2005) A comparison of plant communities in mitigation and reference wetlands in the mid-appalachians. Wetlands 25:130–142. https://doi.org/10.1672/0277-5212(2005)025[0130:acopci]2.0.co;2
Baoyin T, Li FY, Minggagud H, Bao Q, Zhong Y (2015) Mowing succession of species composition is determined by plant growth forms, not photosynthetic pathways in Leymus chinensis grassland of Inner Mongolia. Landsc Ecol 30:1795–1803. https://doi.org/10.1007/s10980-015-0249-6
Bazoge A, Lachance D, Villeneuve C (2014) Identification et délimitation des milieux humides du Québec méridional. Ministère Du Développement durable, de l’Environnement et de la lutte contre les Changements Climatiques. Direction de l’écologie et de la conservation et Direction des politiques de l’eau, Québec, Québec
Bedell J-P, Hechelski M, Saulais M, Lassabatere L (2021) Are acts of selective planting and maintenance drivers for vegetation change in stormwater systems? A case study of two infiltration basins. Ecol Eng 172:106400. https://doi.org/10.1016/j.ecoleng.2021.106400
Birch GF, Matthai C, Fazeli MS (2006) Efficiency of a retention/detention basin to removecontaminants from urban stormwater. Urban Water J 3:69–77. https://doi.org/10.1080/15730620600855894
Birch W, Drescher M, Pittman J, Rooney R (2022) Trends and predictors of wetland conversion in urbanizing environments. J Environ Manage 310:114723. https://doi.org/10.1016/j.jenvman.2022.114723
Birch WS, Drescher M, Rooney RC, Pittman J (2023) Influences of urban stormwater management ponds on wetlandscape connectivity. Can Water Resour J 1–16. https://doi.org/10.1080/07011784.2023.2224522
Brisson J, de Blois S, Lavoie C (2010) Roadside as Invasion Pathway for Common Reed (Phragmites australis). Invasive Plant Sci Manag 3:506–514. https://doi.org/10.1614/ipsm-09-050.1
Brouillet L, Coursol F, Meades SJ, Favreau M, Anions M, Bélisle P, Desmet P (2020) VASCAN, the Database of Vascular Plants of Canada. http://data.canadensys.net/vascan. Accessed September 2020
Bullock JM, Aronson J, Newton AC, Pywell RF, Rey-Benayas JM (2011) Restoration of ecosystem services and biodiversity: conflicts and opportunities. Trends Ecol Evol 26:541–549. https://doi.org/10.1016/j.tree.2011.06.011
Campbell DA, Cole CA, Brooks RP (2002) A comparison of created and natural wetlands in Pennsylvania, USA. Wetl Ecol Manag 10:41–49. https://doi.org/10.1023/A:1014335618914
Canards Illimités Canada (CIC), ministère de l’Environnement et Lutte contre les changements climatiques (MELCC) (2020) Cartographie détaillée des milieux humides des secteurs habités du sud du Québec – Données du projet global [ESRI Canada], Québec (Québec)
Céréghino R, Boix D, Cauchie HM, Martens K, Oertli B (2014) The ecological role of ponds in a changing world. Hydrobiologia 723:1–6. https://doi.org/10.1007/s10750-013-1719-y
Costanza R, d’Arge R, de Groot R et al (1997) The value of the world’s ecosystem services and natural capital. Nature 387:253–260. https://doi.org/10.1038/387253a0
Davidson NC (2014) How much wetland has the world lost? Long-term and recent trends in global wetland area. Mar Freshw Res 65:934–941. https://doi.org/10.1071/mf14173
Davis JA, Froend R (1999) Loss and degradation of wetlands in southwestern Australia: underlying causes, consequences and solutions. Wetl Ecol Manag 7:13–23. https://doi.org/10.1023/A:1008400404021
Dixon MJR, Loh J, Davidson NC, Beltrame C, Freeman R, Walpole M (2016) Tracking global change in ecosystem area: the Wetland Extent trends index. Biol Conserv 193:27–35. https://doi.org/10.1016/j.biocon.2015.10.023
Dodds WK, Wilson KC, Rehmeier RL, Knight GL, Wiggam S, Falke JA, Dalgleish HJ, Bertrand KN (2008) Comparing Ecosystem Goods and Services provided by restored and native lands. Bioscience 58:837–845. https://doi.org/10.1641/b580909
Drinkard MK, Kershner MW, Romito A, Nieset J, de Szalay FA (2011) Responses of plants and invertebrate assemblages to water-level fluctuation in headwater wetlands. J N Am Benthol Soc 30:981–996. https://doi.org/10.1899/10-099.1
ESRI (2020) ArcGIS Desktop: Release 10.8. Environmental Systems Research Institute. Redlands, CA
Fakayode SO, Olu-Owolabi BI (2003) Heavy metal contamination of roadside topsoil in Osogbo, Nigeria: its relationship to traffic density and proximity to highways. Environ Geol 44:150–157. https://doi.org/10.1007/s00254-002-0739-0
Financière Agricole du Québec (FADQ) (2020) Base de données de parcelles et productions agricoles déclarées 2019. https://www.fadq.qc.ca/documents/donnees/base-de-donnees-des-parcelles-et-productions-agricoles-declarees. Accessed February 2019
Findlay CS, Bourdages J (2000) Response time of wetland biodiversity to road construction on adjacent lands. Conserv Biol 14:86–94
Findlay CS, Houlahan J (1997) Anthropogenic correlates of species richness in southeastern Ontario wetlands. Conserv Biol 11:1000–1009. https://doi.org/10.1046/j.1523-1739.1997.96144.x
Fluet-Chouinard E, Stocker BD, Zhang Z et al (2023) Extensive global wetland loss over the past three centuries. Nature 614:281–286. https://doi.org/10.1038/s41586-022-05572-6
Fox J, Ellison S, Murdoch D et al (2020) Car: Companion to Applied Regression. R package version 3.0–10. https://cran.r-project.org/web/packages/car
Gathman JP, Albert DA, Burton TM (2005) Rapid plant community response to a water level peak in northern Lake Huron coastal wetlands. J Gt Lakes Res 31:160–170. https://doi.org/10.1016/s0380-1330(05)70296-3
Gervais C, Trahan R, Moreno D, Drolet AM (1993) Le Phragmites australis Au Québec: distribution géographique, nombres Chromosomiques Et reproduction. Can J Bot 71:1386–1393. https://doi.org/10.1139/b93-166
Greenway M, Jenkins G, Polson C (2007) Macrophyte zonation in stormwater wetlands: getting it right! A case study from subtropical Australia. Water Sci Technol 56:223–231. https://doi.org/10.2166/wst.2007.494
Gülser F, Erdogan E (2008) The effects of heavy metal pollution on enzyme activities and basal soil respiration of roadside soils. Environ Monit Assess 145:127–133. https://doi.org/10.1007/s10661-007-0022-7
Hanslin HM, Mæhlum T, Sæbø A (2017) The response of Phragmites to fluctuating subsurface water levels in constructed stormwater management systems. Ecol Eng 106:385–391. https://doi.org/10.1016/j.ecoleng.2017.06.019
Houlahan JE, Keddy PA, Makkay K, Findlay CS (2006) The effects of adjacent land use on wetland species richness and community composition. Wetlands 26:79–96. https://doi.org/10.1672/0277-5212(2006)26[79:TEOALU]2.0.CO;2
Hwang HM, Fiala MJ, Wade TL, Park D (2019) Review of pollutants in urban road dust: part II. Organic contaminants from vehicles and road management. Int J Urban Sci 23:445–463. https://doi.org/10.1080/12265934.2018.1538811
Jodoin Y, Lavoie C, Villeneuve P, Theriault M, Beaulieu J, Belzile F (2008) Highways as corridors and habitats for the invasive common reed Phragmites australis in Quebec, Canada. J Appl Ecol 45:459–466. https://doi.org/10.1111/j.1365-2664.2007.01362.x
Khalid N, Hussain M, Young HS, Boyce B, Aqeel M, Noman A (2018) Effects of road proximity on heavy metal concentrations in soils and common roadside plants in Southern California. Environ Sci Pollut Res 25:35257–35265. https://doi.org/10.1007/s11356-018-3218-1
Kuntz K, Kinlock NL, Tyler AC, Burkett MB (2018) Stormwater ponds as hotspots of Biodiversity and Biogeochemistry: a comparison of natural and created Wetland ponds. Abstract #B41E-2764, American Geophysical Union, Fall Meeting 2018 B41E-2764
Lebrija-Trejos E, Bongers F, Garcia EAP, Meave JA (2008) Successional change and resilience of a very dry tropical deciduous forest following shifting agriculture. Biotropica 40:422–431. https://doi.org/10.1111/j.1744-7429.2008.00398.x
Legendre P, Legendre L (2012) Numerical Ecology, 3rd English Edition. Elsevier, Amsterdam
Lelong B, Lavoie C, Thériault M (2009) Quels sont les facteurs qui facilitent l’implantation du roseau commun (Phragmites australis) le long des routes du Sud Du Québec? Écoscience 16:224–237. https://doi.org/10.2980/16-2-3237
Marsalek J, Rochfort Q, Grapentine L (2005) Aquatic habitat issues in urban stormwater management: challenges and potential solutions. Ecohydrol Hydrobiol 5:269–279
Matthews JW, Spyreas G, Endress AG (2009) Trajectories of vegetation-based indicators used to assess wetland restoration progress. Ecol Appl 19:2093–2107. https://doi.org/10.1890/08-1371.1
McKercher LJ, Kimball ME, Scaroni AE, White SA, Strosnider WHJ (2024) Stormwater ponds serve as variable quality habitat for diverse taxa. Wetlands Ecol Manage 32:109–131. https://doi.org/10.1007/s11273-023-09964-x
Mendiburu F (2020) Agricolae: Statistical procedures for Agricultural Research. R package version 1.3-3. https://cran.r-project.org/web/packages/agricolae/
Ministère des transports du Québec (MTQ) (2018) Débits de circulation entre les années 2000 et 2018 (fréquence de 2 ans) et carte interactive des données les plus récentes. http://transports.atlas.gouv.qc.ca/Infrastructures/InfrastructuresRoutier.asp. Accessed November 2020
Ministère du développement durable, de l’environnement et des parcs (MDDEP) (2011) Cartographie des milieux humides potentiels – Structure physique des données. Version du 26 avril 2011. https://www.donneesquebec.ca/recherche/dataset/milieux-humides-potentiels. Accessed February 2019
Ministère des ressources naturelles et forêts (MRNF) (2017) Carte écoforestière à jour, Données Québec. https://www.donneesquebec.ca/recherche/dataset/carte-ecoforestiere-avec-perturbations. Accessed February 2019
Ministère des ressources naturelles et forêts (MRNF) (2022) Zones de végétation et domaines bioclimatiques du Québec, Gouvernement du Québec F24-06-2211, Québec, Québec
Mitchell VG, Deletic A, Fletcher TD, Hatt BE, McCarthy DT (2007) Achieving multiple benefits from stormwater harvesting. Water Sci Technol 55:135–144. https://doi.org/10.2166/wst.2007.103
Mitsch WJ, Zhang L, Waletzko E, Bernal B (2014) Validation of the ecosystem services of created wetlands: two decades of plant succession, nutrient retention, and carbon sequestration in experimental riverine marshes. Ecol Eng 72:11–24. https://doi.org/10.1016/j.ecoleng.2014.09.108
Moreno-Mateos D, Power ME, Comin FA, Yockteng R (2012) Structural and functional loss in restored Wetland ecosystems. PLoS Biol 10:8. https://doi.org/10.1371/journal.pbio.1001247
Moser K, Ahn C, Noe G (2007) Characterization of microtopography and its influence on vegetation patterns in created wetlands. Wetlands 27:1081–1097. https://doi.org/10.1672/0277-5212(2007)27[1081:comaii]2.0.co;2
Natarajan P, Davis AP (2015) Hydrologic performance of a Transitioned Infiltration Basin Managing Highway Runoff. J Sustainable Water Built Environ 1(3):04015002. https://doi.org/10.1061/JSWBAY.0000797
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin RB, O’Hara GL, Simpson GL, Solymos P, Stevens MHH, Szoecs E, Wagner H (2020) Vegan: Community Ecology Package. R Package Version 2.5-7. https://cran.r-project.org/web/packages/vegan
Pearson DE, Ortega YK, Villarreal D, Lekberg Y, Cock MC, Eren Ö, Hierro JL (2018) The fluctuating resource hypothesis explains invasibility, but not exotic advantage following disturbance. Ecology 99:1296–1305. https://doi.org/10.1002/ecy.2235
Pellerin S, Poulin M (2013) Analyse de la situation des milieux humides Au Québec Et recommandations à des fins de conservation et de gestion durable. Ministère du Développement Durable, de l’Environnement, de la Faune et des Parcs (MDDEFP), Québec, Québec
Perron MAC, Pick FR (2020) Stormwater ponds as habitat for Odonata in urban areas: the importance of obligate wetland plant species. Biodivers Conserv 29:913–931. https://doi.org/10.1007/s10531-019-01917-2
R Core Team (2019) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Reinartz JA, Warne EL (1993) Development of vegetation in small created wetlands in southeastern Wisconsin. Wetlands 13:153–164. https://doi.org/10.1007/BF03160876
Rivard G (2014) Guide de gestion des eaux pluviales. Ministère De l’Environnement et de la lutte contre les Changements Climatiques (MELCC). Québec, Québec
Rooney RC, Foote L, Krogman N, Pattison JK, Wilson MJ, Bayley SE (2015) Replacing natural wetlands with stormwater management facilities: Biophysical and perceived social values. Water Res 73:17–28. https://doi.org/10.1016/j.watres.2014.12.035
Sinclair JS, Adams CR, Reisinger AJ, Bean E, Reisinger LS, Holmes AL, Iannone BV (2020) High similarity and management-driven differences in the traits of a diverse pool of invasive stormwater pond plants. Landsc Urban Plan 201:16. https://doi.org/10.1016/j.landurbplan.2020.103839
Skultety D, Matthews JW (2018) Human land use as a driver of plant community composition in wetlands of the Chicago metropolitan region. Urban Ecosyst 21:447–458. https://doi.org/10.1007/s11252-018-0730-5
Smith L, Subalusky A, Atkinson C, Earl J, Mushet D, Scott D, Lance S, Johnson S (2019) Biological connectivity of seasonally ponded wetlands across spatial and temporal scales. J Am Water Resour Assoc 55:334–353. https://doi.org/10.1111/1752-1688.12682
Society for Ecological Restoration International Science and Policy Working Group (SERI) (2004) The SER International primer on ecological restoration. www.ser.org & Society for Ecological Restoration International, Tucson, Arizona
Starzec P, Lind BOB, Lanngren A, Lindgren A, Svenson T (2005) Technical and environmental functioning of detention ponds for the treatment of highway and road runoff. Water Air Soil Pollut 163:153–167. https://doi.org/10.1007/s11270-005-0216-y
Sun ZH, Sokolova E, Brittain JE, Saltveit SJ, Rauch S, Meland S (2019) Impact of environmental factors on aquatic biodiversity in roadside stormwater ponds. Sci Rep 9:13. https://doi.org/10.1038/s41598-019-42497-z
Temmink RJM, Lamers LPM, Angelini C et al (2022) Recovering wetland biogeomorphic feedbacks to restore the world’s biotic carbon hotspots. Science 376:594–. https://doi.org/10.1126/science.abn1479
Tixier G, Lafont M, Grapentine L, Rochfort Q, Marsalek J (2011) Ecological risk assessment of urban stormwater ponds: literature review and proposal of a new conceptual approach providing ecological quality goals and the associated bioassessment tools. Ecol Indic 11:1497–1506. https://doi.org/10.1016/j.ecolind.2011.03.027
Truscott AM, Palmer SCF, McGowan GM, Cape JN, Smart S (2005) Vegetation composition of roadside verges in Scotland: the effects of nitrogen deposition, disturbance and management. Environ Pollut 136:109–118. https://doi.org/10.1016/j.envpol.2004.12.009
USDA NCRS (2020) The PLANTS Database. National Plant Data Team, Greensboro, NC. Accessed September 2020 http://plants.usda.gov
Wei AH, Chow-Fraser P (2006) Synergistic impact of water level fluctuation and invasion of Glyceria on Typha in a freshwater marsh of Lake Ontario. Aquat Bot 84:63–69. https://doi.org/10.1016/j.aquabot.2005.07.012
Wigginton SK, Meyerson LA (2018) Passive Roadside Restoration reduces management costs and fosters native Habitat. Ecol Restor 36:1. https://doi.org/10.3368/er.36.1.41
Wilsey BJ, Barber K, Martin LM (2015) Exotic grassland species have stronger priority effects than natives regardless of whether they are cultivated or wild genotypes. New Phytol 205:928–937. https://doi.org/10.1111/nph.13028
Zedler JB, Callaway JC (1999) Tracking wetland restoration: do mitigation sites follow desired trajectories? Restor Ecol 7:69–73. https://doi.org/10.1046/j.1526-100X.1999.07108.x
Zedler JB, Kercher S (2005) Wetland resources: Status, trends, ecosystem services, and restorability. Annual Review of Environment and resources. Annual Reviews, Palo Alto, pp 39–74
Zhang HJ, Wang RQ, Wang X, Du N, Ge XL, Du YD, Liu J (2015) Recurrent water level fluctuation alleviates the effects of Submergence stress on the Invasive Riparian Plant Alternanthera philoxeroides. PLoS ONE 10:15. https://doi.org/10.1371/journal.pone.0129549
Acknowledgements
We would like to thank the Ministère des transports du Québec for their financial support, É. Bergeron de Pani, M. Mazerolle and M. Hayes for scientific advice and field assistance. We thank K. Grislis for linguistic revision and D. Bastien for taxonomic identifications.
Funding
This work was supported by the Ministère des Transports du Québec (Grant number R803.1).
Author information
Authors and Affiliations
Contributions
Conceptualization and methodology: P.-A. Bergeron D’Aoust, S. Pellerin, and M. Poulin; Formal analysis and investigation: P.-A. Bergeron D’Aoust and M. Vaillancourt; Writing – original draft: P.-A. Bergeron D’Aoust and M. Vaillancourt; Writing – review & editing: S. Pellerin and M. Poulin.
Corresponding author
Ethics declarations
Competing Interests
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic Supplementary Material
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
D’Aoust, PA.B., Vaillancourt, M., Pellerin, S. et al. Are Plant Communities of Roadside Stormwater Ponds Similar to those Found in Natural Wetlands?. Wetlands 44, 99 (2024). https://doi.org/10.1007/s13157-024-01846-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13157-024-01846-z