
Abstract
Peatlands are responsible for the majority of methane (CH4) emission from wetlands globally. Hydrological changes induced by climatic and anthropogenic disturbance may substantially alter CH4 emission in peatlands. Here we measured CH4 emission monthly for 1.5 years in natural, drained and restored shrub bogs in North Carolina, USA. Methane emissions from all sites were consistently low (< 0.05 mg CH4 m− 2 h− 1). We occasionally detected markedly higher CH4 emissions (> 1 mg CH4 m− 2 h− 1) at sites where the water level remained close to the ground surface for 2–3 months, suggesting that surface litter mostly, not deep peat, contributes to CH4 emission. We verified this inference by incubating 2-cm sections of peat sliced from intact soil cores for 6 months. Only the saturated surface litter emitted CH4, which indicated a 5-cm threshold of ground water level for CH4 emission in our shrub bogs. During a wet year, water levels in the wet sites (natural and restored) remained at least 5 cm below soil surface for about 90 % of the days. We thus demonstrate the CH4 emission is negligible from these shrub bogs. This study also indicates that restoration through a non-inundated rewetting would not stimulate CH4 emission in drained/degraded low-latitude shrub bogs, such as pocosins.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Methane, as the most abundant greenhouse gas after CO2, is responsible for a large amount of anthropogenically-mediated warming due to its high global warming potential (IPCC 2006). Methanogens produce CH4 efficiently under strictly anaerobic conditions. Only fully or partially saturated wetlands could emit substantial amounts of CH4 that accounts for the largest natural source of atmospheric CH4 (Bridgham et al. 2013). Peatlands, a type of wetland characterized by highly organic soils, store about one third of global soil carbon (Limpens et al. 2008; Joosten 2010). A comprehensive synthesis including 19,000 instantaneous measurements mostly from boreal, temperate, and subtropical peatlands reported that the mean CH4 emission from these peatlands was about 4.0 mg m− 2 h− 1 ranging from − 0.8 to 238.4 mg m− 2 h− 1 (Turetsky et al. 2014). Based on the mean CH4 emission, the near-natural peatlands (> 3 million km2) worldwide could release about 105 Tg CH4 yr− 1, which represents two thirds of CH4 emitted from global wetlands (Bridgham et al. 2013).
Although temperature, water table, and vegetation have been identified as the primary controls on CH4 emission, the high spatiotemporal variations by several orders of magnitude (Bridgham et al. 2013; Turetsky et al. 2014) raise uncertainty in predicting CH4 emission from peatlands. More interestingly, among the 19,000 measurements, the highest CH4 emissions (238.4 mg m− 2 h− 1) occurred in bogs under optimal water levels around 25 cm below peat surface (Turetsky et al. 2014), which indicates that bogs with low concentrations of inorganic alternative electron acceptors like NO3−, Mn (III,V), Fe(III) and SO42− actually could have a more important contribution to global atmospheric CH4 if the hydrology shifts toward more methanogen-prone conditions. Recent studies also found that humic substances in peat can serve as organic alternative electron acceptors, constraining CH4 production (Keller et al. 2009; Klupfel et al. 2014). The role of humic substances could partially explain why highly recalcitrant deep peat plays only a minor role in CH4 production in many studies (Chanton et al. 1995; Charman et al. 1999; Couwenberg 2009; Strack and Zuback 2013), making the development of peatland CH4 emission factors even more complicated. Most of the bog data from Turetsky et al. (2014) were collected from boreal regions, where Sphagnum mosses dominate. Given low phenolics and humic substance in fresh Sphagnum (Wang et al. 2015), we deduce a possible reason for the maximum CH4 emission occurred in the upper surfaces of groundwater at around 25 cm (Turetsky et al. 2014). Living and marginally decomposed Sphagnum mosses provide sufficient fresh carbon to anoxic layers just below 25-cm layers for the anaerobically active methanogens, but are not yet limited by the organic electron acceptor—dissolved humic substance (Keller et al. 2009; Klupfel et al. 2014) transported from the highly humified catotelm.
The synthesis by Turetsky et al. (2014) brings up questions whether broadly-distributed lower-latitude peatlands emit comparable CH4 as these subtropical bogs typically experience a seasonal lowering of groundwater often well below the ground surface and are under warmer temperatures (Wang et al. 2015). While the response of CH4 emission to water level in these low-latitude wooded peatlands might differ from boreal Sphagnum peatlands because of the lower water levels, higher phenolics, and higher humic substances in the low-latitude peatlands (Wang et al. 2015; Hodgkins et al. 2018). In terms of restoring degraded low-latitude peatlands and expanded wooded plants in current boreal Sphagnum peatlands due to climate change, a key question that needs to be resolved is what is the optimum water level or hydrological manipulation to mitigate CH4 emission from these peatlands. To address the question, we monitored CH4 emission and associated environmental parameters in natural, restored, and drained sites in a low-latitude shrub bog (pocosin) from August 2011 to January 2013. In addition, we randomly measured CH4 emission from selected shallow ditches and their riparian areas to identify potential hotspots in the shrub bog.
Methods
Site Description
Pocosins are naturally-occurring freshwater evergreen shrub bogs and are found along the southeastern coastal plain from Virginia to Florida in the USA (Richardson 2012). Natural pocosins covered over 900,000 hectares in North Carolina alone before the 1970s, after which, about 70 % of pocosins were ditched and drained for farming or forestry (Richardson 1991). In the 1990s, the Pocosin Lakes National Wildlife Refuge (PLNWR) administration started blocking the drainage canals to restore water levels. Our study sites are located in the PLNWR in the east coast of North Carolina, USA. The site is dominated by dense broadleaf evergreen shrubs (< 2 m high) with peat depths of 1 ~ 3 m. The mean annual temperature is 16.8 °C, and precipitation is about 1230 mm. Because of high evapotranspiration, the groundwater levels rarely rise above the ground surface and often stay > 20 cm below the ground surface (Wang et al. 2015).
We established this study in 5 blocks (1600 × 900 m each) with different hydrologic regimes—natural, drained, and restored (rewetting) as treatments (Fig. 1). Each treatment had been in place for > 20 years. The natural site (RF T2) is close to Pungo Lake and has been rarely disturbed, with water levels of 0 ~ − 60 cm (negative values of water level indicate water levels were below ground surface and hereinafter) in winter and often < − 100 cm in summer. It is covered by mature trees, including loblolly bay [Gordonia lasianthus (L.) Ellis], pond pine (Pinus serotina Michx.), and swamp bay [Persea palustris (Raf.) Sarg.]. For the restored sites (B7, C2T2, and D11), the water level is about − 20 ~ − 30 cm, where native shrubs dominate, including inkberry [Ilex glabra (L.) A. Gray], large gallberry [Ilex coriacea (Pursh) Chapm.], honeycup [Zenobia pulverulenta (W. Bartram ex. Willd.) Pollard], fetterbush lyonia [Lyonia lucida (Lam.) K. Koch] and laurel greenbrier (Smilax laurifolia L.) with some smaller trees of pond pine, and loblolly bay. The drained sites (C14T1, C14T2 and C14T3,) have the lowest water levels, mostly below − 50 cm depth and lower than − 150 cm in summer. The ground cover is occupied by western brakenfern [Pteridium aquilinum (L.) Kuhn] with scattered woody shrubs like winged sumac (Rhus copallinum L.), wax myrtle [Morella cerifera (L.) Small] and titi (Cyrilla racemiflora L.).

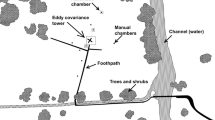
Sampling sites in the Pocosin Lakes National Wildlife Refuge on the southeastern coastal plains of the USA (image from Google Earth)
Field Measurements
A static chamber technique was used to measure CH4 emission monthly from August 2011 to January 2013. We initially set up triplicate plots for each treatment. Black bears repeatedly destroyed on site equipment at two of the natural site plots; therefore, only one plot was kept there. The static chamber contains two parts, an anchored bottom soil collar and a matched and portable top chamber. In May 2011, we permanently installed triplicate soil collars (28-cm diameter, 13-cm height) at each plot. Each collar is fashioned with a circular groove (2 cm wide by 1.5 cm deep) on the upper rim to receive the corresponding upper chamber. When the soil collar was submerged in > 5 cm standing water, the upper chamber was modified to act as a floating buoy. Plants (aboveground) inside the collar were removed. A temperature probe and a battery-driven fan for air mixing were installed inside the upper chamber (28-cm diameter, 21-cm height). At each plot, we recorded water levels every 30 min by an automated datalogger (Solinst Levelogger model 3001) since 2010.
When collecting gas samples, we placed the lower edge of the top chamber into the water-filled groove to create a good seal. Four gas samples were taken at 10–15 min interval, depending on season, from the headspace through polytetrafluoroethylene tubes into 100 mL gas sampling bags (multilayer polymer with aluminum foil) by syringe. Simultaneously, air temperature, soil temperature, and soil moisture (0–5 cm) were recorded, and the surface soil (0–5 cm) was collected for physicochemical analyses at each gas sampling (see details later). Methane concentrations were determined within 3 d after sampling by a gas chromatograph (GC, Varian 450, California, USA) equipped with a flame ionization detector with a methanizer. The analytical accuracy was maintained by calibrating the GC against two standard gas mixtures in every 8 samples. The concentration of the CH4 standard was 2 ppm. The CH4 emission rate was calculated from the linear change of its concentrations in the chamber as a function of time, base area, chamber volume, and the molar volume of CH4 at air temperature inside of chamber. Emissions with an r2 < 0.8 in a linear regression of concentration change over time were considered erroneous and excluded. Based on the gas sampling interval and our GC sensitivity, the lowest reliable CH4 emission rate were about 0.01–0.03 mg CH4 m− 2 h− 1. About 76 and 42 % of emission rates were higher than 0.01 and 0.03 mg CH4 m− 2 h− 1, respectively.
Microcosm Experiment
As we observed clear differences in CH4 emissions across replicate collars located only 2–3 m apart during the 2 to 3-month flooding period in some natural and restored sites, the variations of water levels near the soil surface seemed to contribute to the emission patterns. We collected three soil cores (30–45 cm deep) from the natural and restored sites, including RFT2 at the north of Pungo Lake, C2T2, and Pungo East at the east edge of Pungo Lake. Pungo East, dominated by shrubs, is also a natural site without major disturbance for > 30 years, more like the restored C2T2 than the natural RFT2 in terms of vegetation. Each soil core was sliced into 2-cm Sec. (54 in total) on site and transported back to our lab and stored in a cold room at 4 °C. About 8–10 g fresh soil (~ 2.5 g dry soil) from each section was placed in a 60-mL vial with a silicone stopper. A 20 mL DI water was then added to a fully saturated condition at a water level about 2–3 cm above the soil surface, similar to the field conditions when we observed high CH4 emissions. We used Parafilm M® Laboratory film, which is air permeable but water-resistant, to seal the top during the non-sampling period. Hence, the incubation with oxygen presence in the headspace could, to a large extent, mimic the field surface-saturation conditions. After a one-week equilibration, we collected gas samples with a syringe from the vial headspace at the beginning and end of 48-hour sealed incubation. Gas samples were analyzed by a GC (Varian 450, California, USA) for CH4 concentration. The CH4 emission was calculated based on the elevated CH4 concentration, time, air volume and temperature (25 °C) in the vial and the dry weight of peat soil. We took samples twice a week initially, then progressed to weekly, and finally biweekly after the third month. We did not detect any CH4 emission until the end of the third month.
Soil Chemistry
Deionized-water extraction was used to determine dissolved organic carbon (DOC) and dissolved inorganic carbon (DIC) by a total C analyzer (Shimadzu 5000 A, Kyoto, Japan), whereas soluble phenolics were determined by following the Folin–Ciocalteu procedure (Lowe 1993). Inorganic nitrogen (NH4+–N and [NO3− + NO2−]–N) extracted with 2 M KCl was analyzed colorimetrically on a flow–injection analyzer (Lachat QuikChem 8000, Wisconsin, USA). A combustion CN analyzer (ThermoQuest Flash EA1112, Milan, Italy) was used to measure total carbon and total nitrogen in soil.
Statistical Analysis
A Shapiro-Wilk normality test was run first for CH4 emission at each site. The NPAR1WAY procedure performs nonparametric tests to compare the difference in CH4 emission among sites. Pearson’s correlation was employed to detect the association between environmental factors and CH4 emission using the mean CH4 emissions from triplicated plots at each site. Standard error was calculated for means and error bars. Unless otherwise noted, significant difference was set at a probability of 0.05. All analyses were conducted with SAS 9.4 software (SAS Institute Inc., Cary, NC, USA).
Results
The soil physicochemical parameters have been described in detail previously (Supplementary Table 3 in Wang et al. 2015). Briefly, the soil in the natural site (pH = 3.3) was more acidic than that in other sites (pH: 4 ~ 5). The soils in all sites contained similar soil carbon (53–56 %). The drained sites showed much higher N content of nitrate plus nitrite, about twice as much as that in the natural and restored sites. The annual precipitation in 2011 was 1150 mm, and the year of 2012 was a little wetter (1205 mm) with 70 % of its annual precipitation in the growing season. The mean air temperatures in 2011 and 2012 were similar. The mean water levels in 2012 was − 25 ± 17 cm at the natural site (RFT2). Mean water levels were − 16 ± 7, − 38 ± 10, and − 55 ± 14 cm at the restored sites (B7, C2T2, and D11T1, respectively) and − 68 ± 8, − 70 ± 10 and − 84 ± 11 cm at the drained sites (C14T1, C14T2 and C14T3, respectively). As the synthesis study (Turetsky et al. 2014) showed the optimal water level for CH4 emission in northern bogs is about 25 cm below ground surface. To compare with their study we counted the total days of water levels at − 20 ~ − 30 cm, > −5 cm and > − 10 cm in 2012 (Table 1). There were 34, 113 and 80 days in 2012 at RFT2, B7 and C2T2, respectively, when water levels stayed within − 20 ~ − 30 cm. We observed water levels above − 5 cm for 44 days, and above − 10 cm for 76 days at the natural site (RFT2). The water levels in the drained sites (C14T1, C14T2, C14T3) were nearly always below − 50 cm.
Methane emissions and their temporal variations at each site are presented in Figs. 2 and 3A. The median emission at the natural site (RFT2) was 0.128 mg CH4 m− 2 h− 1, which was much higher than at the restored sites (0.024, 0.018 and 0.015 mg CH4 m− 2 h− 1 at B7, C2T2 and D11T1, respectively) and the drained sites (0.005, 0.005 and 0.008 mg CH4 m− 2 h− 1 at C14T1, C14T2 and C14T3, respectively). Generally, the mean emissions at each site were about 10 times higher than the median. The emissions of CH4 from the drained pocosin were always small, often below the detection limit and negative (CH4 oxidation was higher than its production) at times. In the whole observation period, we only detected high emission (> 1 mg CH4 m− 2 h− 1) at RFT2 and B7 two months after the summer storm that happened in late June 2012 (Fig. 3A and B), in which the emissions reached 5.8 mg CH4 m− 2 h− 1 at B7 and 45.6 mg CH4 m− 2 h− 1 at RFT2 in September and November, respectively. However, during the peak emission period we observed exceptional high spatial variation of CH4 emission within the triplicate plots that were only 2–3 m apart at the same site (Fig. 3A). We dug a small hole next to each chamber to measure the water level temporally below the ground surface or measured the standing water depth by a ruler at sites RFT2 and B7, and we found an exponential increase of CH4 emission with water levels when the water level rose above − 5 cm (Fig. 4A and B) and the emission reached its maximum when the water level was close to the ground surface. We also examined what other physicochemical parameters might affect the emission by running Pearson’s correlation using all data regardless of treatments. These parameters included soil and air temperatures, soil moisture (0–5 cm), ammonium, nitrate + nitrite, soluble phenolics, DOC, soil total nitrogen and carbon. Other than water level (r = 0.311, p = 0.001), only soil moisture (r = 0.566, p < 0.0001) was significantly related to CH4 emissions, which actually was partially controlled by the water level (r = 0.202, p = 0.04).

Methane emission from the natural (RFT2), restored (B7, C2T2 and D11T1) and drained (C14T1, C14T2 and C14T3) sites in pocosins. All measurements at triplicated plots are included in this box chart. Same letters above the box indicates nonsignificant differences of CH4 emission among sites, CI = confidence interval

Temporal variations of CH4 emission (A, mean ± 1 SD, n = 3) and water level (B) in 7 sites in pocosins. The dash line at water level of − 5 cm, is identified as a threshold of CH4 emission in the shrub bogs

Spatial variation of CH4 emission caused by microtopography-induced water level differences at B7 restored (A) and RFT2 natural (B). The same symbol in each figure represents the emissions from the triplicate soil collars located only 2–3 m apart at one site
As methanogens require strictly anaerobic conditions, we expected even higher CH4 emission in locations with higher standing water. However, in the shallow flooded ditch in the pocosin the CH4 emissions were low, only about 0.05 mg CH4 m− 2 h− 1 on the same day in November 2012 when we observed the peak emission at RFT2. In October 2019, we tested CH4 emissions on the edge of a small shallow ditch in pocosins. Again, the highest emission reached 128 mg CH4 m− 2 h− 1 with the water level close to the ground surface, while only 1 m away from this hotspot the higher elevation (about 30 cm above) plot emitted CH4 only at a rate of 2.7 mg CH4 m− 2 h− 1.
Collectively, these findings suggest that only the surface soil layer, after months of saturation, significantly contributed to pocosin CH4 emissions. This assumption is verified by our incubation experiment (Fig. 5). In this experiment, we did not find any CH4 emission within the first three months although the water levels were always about 2–3 cm above the soil surface. In the fourth month high CH4 emissions occurred only in the surface soil [(0–4 cm) in RFT2 and Pungo East and (0- 6 cm) in C2T2], and sharply dropped to almost undetectable below 4–6 cm, hence, we suggest the-water level threshold for CH4 emissions in pocosins is about 5 cm below the peat surface. Based on the bulk density of 0.03 g cm− 3 (Richardson 2003), the potential CH4 emission at 25 °C from the top 5-cm soil in RFT2 natural site was 0.104 mg CH4 m− 2 h− 1.

Methane emission along soil depths at 2 cm intervals of soil cores in pocosin after 4 months of lab incubation under a saturated condition
Discussion
Optimal Water Levels, Potential Hotspot and Hot Periods for CH4 Emission in Shrub Bogs
Our study shows that water levels play an overarching role in controlling CH4 emission in pocosin shrub bogs. When water level was below a − 5 cm threshold, the CH4 emission was low and could be ignored. We did not observe any relationships between CH4 emission and soil or air temperature although the soil temperature varied in from 2 to 30 °C. Low-latitude peatlands differ from boreal peatlands, where temperature is more of a limiting factor than in pocosins. Also, the soil temperature tended to be higher when water levels were lower in pocosins; thus, it is difficult to determine the full temperature effect.
As expected, the optimal water levels for increasing CH4 emissions in the pocosin shrub bogs are much higher, i.e., close to the ground surface, which differs from the Sphagnum peatlands where it is at about − 25 cm (Turetsky et al. 2014). The acrotelm of Sphagnum-dominated boreal peatlands consists of substantial living, or marginally decomposed mosses, that are generally thicker (> 25 cm) than in wooded peatlands. These living Sphagnum mosses maintain high soil moisture and release fresh carbon in abundance, which is the primary substrate for methanogenic Archaea (Segers 1998; Lai 2009). However, this substrate contains little humic substance (Hodgkins et al. 2018) that can act as alternative electron acceptors to constrain CH4 production (Keller et al. 2009; Klupfel et al. 2014). Although the old recalcitrant peat in our deeper layers is saturated, the lower fresh carbon and higher humic substance result in very low rates of CH4 production, a finding supported by our incubation experiment (Fig. 5) and also consistent with other studies (Chanton et al. 1995; Charman et al. 1999; Clymo and Bryant 2008). In addition, the water level of − 25 cm in northern Sphagnum peatlands could be a sweet-spot layer of anoxia for methanogens with plenty of fresh carbon and lower humic substances. For example, Bogs at Mer Bleue in Canada, carbohydrate content in the surface layer (0 ~ − 25 cm) was about twice as high as that in deeper layers (below − 25 cm) and it showed a sharp decrease at depths of round − 25 ~ − 30 cm (Hodgkins et al. 2018). However, compared to Sphagnum peatlands, the fresh litter layer in our shrub peatlands is much thinner, and in pocosins soil color turns to dark black below − 3 ~ − 5 cm with a sharp decrease in carbohydrate content (Hodgkins et al. 2018), which indicates that the humification increases from the surface to soils below − 5 cm. Higher peat humics content in deeper soil might be the reason why we only detected high CH4 emission when the water level rose above − 5 cm. Importantly, the water levels of > − 5 cm in our wettest site appeared in less than 15 % of the days in a wet year, thus the Archaea might be dormant most of the time. After a heavy storm, methanogens also take a long time to regain a fully active metabolism to break their dormancy while the saturation partially suppresses methanotrophs (Taubner et al. 2015). Our laboratory incubation experiment with oxygen present in the headspace differed from a fully-anaerobic incubation but was close to the field conditions. Thus, the results in the incubation were in line with our field observations, that is, it took 2–3 months for the saturated surface layer to emit CH4 at a high rate. As we sampled monthly, we may have missed the peak event while still showing exceptional burst after the heavy storm in 2012, which did not happen in a drier 2011 after a light storm. Detailed sampling research is needed to determine the dormancy time of methanogens in pocosins that have high phenolic content. The high emissions we observed follow the range that was reported in peatlands globally (Turetsky et al. 2014). Therefore, the surface saturated sites are potential hotspots of CH4 emission in the shrub bogs, while only occurring after months of saturation.
Roles of Plants in CH4 Emission in Peatlands
In this study, the CH4 emissions from the natural sites were much higher than those in the restored site even when the surface litter in both sites was under saturated conditions (Fig. 4). The field observation was reaffirmed in the incubation experiment (Fig. 5) in which we had mimicked the surface-saturation field conditions only. The incubated vials allowed oxygen in the headspace which means the CH4 emission we observed represent a difference between CH4 production and oxidation. Therefore, the different on site CH4 emissions between the natural and restored sites likely stemmed from distinct substrates for methanogens, rather than from the time we took measurements, i.e., whether we caught the right moments. Phenolics have shown to constrain CH4 emission (Miao et al. 2012; Ye et al. 2012; Ward et al. 2013). Compared to high-phenolic shrubs (Wang et al. 2015), we noticed that the mature trees at the natural sites contained less phenolics, and thus allowed for methanogen more accessible to fresh carbon. Also, in comparison with the potential emission in the incubation, we found that the highest on site emissions were over 400 times higher, likely caused by diminishing fresh carbon availability in the absence of living plants during laboratory incubation. Hence the quality of fresh litter or exudates from the living plants may play important roles in regulating CH4 emission when the water level is optimum. Finally, as noted earlier, the natural site has existed next to Pungo Lake for thousands of years and thus the hydrologic flooding conditions have greatly influenced the plant and microbial communities at this site. This suggests the pocosin soil near the lake may be a hotspot for CH4 emissions compared to the domed peatlands found away from the lakes.
Implications for Peatland Restoration and Unravelling of CH4 Emission Puzzle
Rewetting drained or degraded peatlands as a climate-mitigation measure can effectively reduce aerobic CO2 emission. However, addressing CH4 emission after rewetting is still a challenge. Our research, together with other studies (Turetsky et al. 2014), suggests that the varied water-level threshold for CH4 emission in peatlands might be the reason why CH4 emission responses to rewetting differ among types of wetland and among samples within the same wetland. Only when the water levels exceed a depth threshold do the methanogens become active and/or more CH4 is produced by methanogens than consumed by methanotrophs. In our shrub bogs the threshold is about − 5 cm, and we also observed the soil moisture (0 ~ − 5 cm) significantly impacted the emissions, which is consistent with the observations in the Great Dismal Swamp (Gutenberg et al. 2019). The pocosin site in the Great Dismal Swamp, with comparable plant communities to our natural site, emitted CH4 as high as 75 mg CH4 m− 2 h− 1 when water level stayed above − 5 cm. Thus water level thresholds for CH4 emissions likely are controlled by the downward infiltration of fresh carbon/substrate that are affected by plant community, root depth, root exudates and the upward moving of dissolved humic substance (Blodau and Deppe 2012) as an alternative electron acceptor when water level rises. More studies are needed to detect the water-level thresholds in different peatlands, to determine the optimal water level for CH4 abatement, which can be site-specific. We suggest that the future restoration and hydrological management in peatlands should keep the water level below the threshold for CH4 emission. Otherwise, the higher CH4 emission after rewetting (Mahmood and Strack 2011; Strack et al. 2014; Knox et al. 2015; Christen et al. 2016) may balance out the reduced CO2 emission as the radiative forcing of CH4 is about 32–45 times more than CO2 in a century on a mass basis (Neubauer and Megonigal 2015). Alternatively, as only the saturated litter layer produces substantial CH4 (Fig. 5), the top litter layer above the threshold could be removed prior rewetting to reduce CH4 emission at the beginning of rewetting. Removal of surface litter has been shown to reduce CH4 emissions by factors of 30–400 in a temperate rewetted bog (Huth et al. 2020), and over 1000 times from rewetting former agricultural peatlands (Harpenslager et al. 2015).
The exceptionally high spatiotemporal variations, by several orders of magnitude, seriously limit the predictability of CH4 emission from peatlands. Our studies suggest that the threshold of water level and the time lag of CH4 production after rewetting or storm might be two keys to more precisely predicting and managing CH4 emission in peatlands. Only when the water levels are above the water-level threshold for a period of time can methanogens be fully reactivated from their dormancy, and the peatlands start emitting CH4. Therefore, after the site-specific or peatland type-specific threshold of water level and the time lag for methanogens are identified, the rainfall amount, intensity, duration, and frequency, can be used to develop water management practices to determine whether and when the peatlands become CH4 sources. Once the water level and time lag requirements are met, the magnitude of CH4 emissions could be further explained by temperature, soil moisture, and substrate quality associated with plant communities, as well as organic and inorganic electron acceptors (Bridgham et al. 2013).
Data Availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Code Availability
Not applicable.
References
Blodau C, Deppe M (2012) Humic acid addition lowers methane release in peats of the Mer Bleue bog, Canada. Soil Biology and Biochemistry 52:96–98
Bridgham SD, Cadillo-Quiroz H, Keller JK, Zhuang Q (2013) Methane emissions from wetlands: biogeochemical, microbial, and modeling perspectives from local to global scales. Global Change Biology 19:1325–1346
Chanton JP, Bauer JE, Glaser PA, Siegel DI, Kelley CA, Tyler SC, Romanowicz EH, Lazrus A (1995) Radiocarbon evidence for the substrates supporting methane formation within northern Minnesota peatlands. Geochimica et Cosmochimica Acta 59:3663–3668
Charman DJ, Aravena R, Bryant CL, Harkness DD (1999) Carbon isotopes in peat, DOC, CO2, and CH4 in a Holocene peatland on Dartmoor, southwest England. Geology 27:539–542
Christen A, Jassal RS, Black TA, Grant NJ, Hawthorne I, Johnson MS, Lee S-C, Merkens M (2016) Summertime greenhouse gas fluxes from an urban bog undergoing restoration through rewetting. Mires Peat 17:1–24
Clymo R, Bryant C (2008) Diffusion and mass flow of dissolved carbon dioxide, methane, and dissolved organic carbon in a 7-m deep raised peat bog. Geochimica et Cosmochimica Acta 72:2048–2066
Couwenberg J (2009) Methane emissions from peat soils (organic soils, histosols): facts, MRV-ability, emission factors. Greifswald University, Ede
Gutenberg L, Krauss KW, Qu JJ, Ahn C, Hogan D, Zhu Z, Xu C (2019) Carbon Dioxide Emissions and Methane Flux from Forested Wetland Soils of the Great Dismal Swamp, USA. Environ Manage 64:190–200
Harpenslager SF, Van Den Elzen E, Kox MAR, Smolders AJP, Ettwig KF, Lamers LPM (2015) Rewetting former agricultural peatlands: Topsoil removal as a prerequisite to avoid strong nutrient and greenhouse gas emissions. Ecological Engineering 84:159–168
Hodgkins SB, Richardson CJ, Dommain R, Wang H, Glaser PH, Verbeke B, Winkler BR, Cobb AR, Rich VI, Missilmani M, Flanagan N, Ho M, Hoyt AM, Harvey CF, Vining SR, Hough MA, Moore TR, Richard PJH, De La Cruz FB, Toufaily J, Hamdan R, Cooper WT, Chanton JP (2018) Tropical peatland carbon storage linked to global latitudinal trends in peat recalcitrance. Nat Commun 9:3640. https://doi.org/10.1038/s41467-018-06050-2
Huth V, Günther A, Bartel A, Hofer B, Jacobs O, Jantz N, Meister M, Rosinski E, Urich T, Weil M, Zak D, Jurasinski G (2020) Topsoil removal reduced in-situ methane emissions in a temperate rewetted bog grassland by a hundredfold. Science of The Total Environment 721:137763
IPCC (2006) 2006 IPCC Guidelines for National Greenhouse Gas Inventories, Prepared by the NationalGreenhouse Gas Inventories Programme, Eggleston H.S., Buendia L., Miwa K., Ngara T. and Tanabe K. (eds). IGES, Japan
Joosten H (2010) The Global Peatland CO2 Picture. Peatland status and drainage related emissions in all countries of the world. Ede, Netherlands, p 36
Keller JK, Weisenhorn PB, Megonigal JP (2009) Humic acids as electron acceptors in wetland decomposition. Soil Biology and Biochemistry 41:1518–1522
Klupfel L, Piepenbrock A, Kappler A, Sander M (2014) Humic substances as fully regenerable electron acceptors in recurrently anoxic environments. Nature Geoscience 7:195–200
Knox SH, Sturtevant C, Matthes JH, Koteen L, Verfaillie J, Baldocchi D (2015) Agricultural peatland restoration: effects of land-use change on greenhouse gas (CO2 and CH4) fluxes in the Sacramento-San Joaquin Delta. Glob Chang Biol 21:750–765
Lai DYF (2009) Methane dynamics in northern peatlands: A review. Pedosphere 19:409–421
Limpens J, Berendse F, Blodau C, Canadell JG, Freeman C, Holden J, Roulet N, Rydin H, Schaepman-Strub G (2008) Peatlands and the carbon cycle: from local processes to global implications – a synthesis. Biogeosciences 5:1475–1491
Lowe L (1993) Water-soluble phenolic materials. Soil Sampling and Methods of Analysis. CRC Press, Boca Raton, pp 409–412
Mahmood MS, Strack M (2011) Methane dynamics of recolonized cutover minerotrophic peatland: Implications for restoration. Ecological Engineering 37:1859–1868
Miao Y, Song C, Sun L, Wang X, Meng H, Mao R (2012) Growing season methane emission from a boreal peatland in the continuous permafrost zone of Northeast China: effects of active layer depth and vegetation. Biogeosciences 9:4455–4464
Neubauer SC, Megonigal JP (2015) Moving beyond global warming potentials to quantify the climatic role of ecosystems. Ecosystems 18:1000–1013
Richardson CJ (1991) Pocosins: An ecological perspective. Wetlands 11:335–354
Richardson CJ (2003) Pocosins: Hydrologically isolated or integrated wetlands on the landscape? Wetlands 23:563–576
Richardson CJ (2012) 14. Pocosins: Evergreen Shrub Bogs of The Southeast. In: Batzer DB and Baldwin AH. (ed) Wetland Habitats of North America. University of California Press, Berkeley, pp 189–202
Segers R (1998) Methane production and methane consumption: a review of processes underlying wetland methane fluxes. Biogeochemistry 41:23–51
Strack M, Zuback YCA (2013) Annual carbon balance of a peatland 10 year following restoration. Biogeosciences 10:2885–2896
Strack M, Keith AM, Xu B (2014) Growing season carbon dioxide and methane exchange at a restored peatland on the Western Boreal Plain. Ecological Engineering 64:231–239
Taubner R-S, Schleper C, Firneis MG, Rittmann SK-MR (2015) Assessing the ecophysiology of methanogens in the context of recent astrobiological and planetological studies. Life 5:1652–1686. https://doi.org/10.3390/life5041652
Turetsky MR, Kotowska A, Bubier J, Dise NB, Crill P, Hornibrook ERC, Minkkinen K, Moore TR, Myers-Smith IH, Nykänen H, Olefeldt D, Rinne J, Saarnio S, Shurpali N, Tuittila E-S, Waddington JM, White JR, Wickland KP, Wilmking M (2014) A synthesis of methane emissions from 71 northern, temperate, and subtropical wetlands. Global Change Biology 20:2183–2197
Wang H, Richardson CJ, Ho M (2015) Dual controls on carbon loss during drought in peatlands. Nature Climate Change 5:584–587
Ward S, Ostle N, Oakley S, Quirk H, Henrys P, Bardgett R (2013) Warming effects on greenhouse gas fluxes in peatlands are modulated by vegetation composition. Ecology Letters 16:1285–1293
Ye R, Jin Q, Bohannan B, Keller JK, McAllister SA, Bridgham SD (2012) pH controls over anaerobic carbon mineralization, the efficiency of methane production, and methanogenic pathways in peatlands across an ombrotrophic–minerotrophic gradient. Soil Biology and Biochemistry 54:36–47
Acknowledgements
We would like to thank Wes Willis, Jonathan Bills and Scott Winton for the field and lab measurements, and Dr. Randy Neighbarger for technical editing. Our thanks also go to Sara Ward and Dave Kitts, USFWS for helping with site selections and field assistance.
Funding
U.S. Fish and Wildlife Service, Region 4, The Nature Conservancy of North Carolina, US DOE Office of Science, Terrestrial Ecosystem Sciences (DE-SC0012272), and the Duke University Wetland Center Endowment provided financial support.
Author information
Authors and Affiliations
Contributions
HW and CJR designed and set up the field experiment. HW and MH collected and analyzed soil/gas samples, HW design and conduct the lab incubation. NF monitored water level in field. HW was a major contributor in writing the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Conflicts of Interest/Competing Interests
The authors have no conflicts of interest.
Ethics Approval
Not applicable.
Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article belongs to the Topical Collection: Peatlands.
Rights and permissions
About this article

Cite this article
Wang, H., Ho, M., Flanagan, N. et al. The Effects of Hydrological Management on Methane Emissions from Southeastern Shrub Bogs of the USA. Wetlands 41, 87 (2021). https://doi.org/10.1007/s13157-021-01486-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13157-021-01486-7