
Abstract
Peatlands play a disproportionate role in the global carbon cycle. However, many peatlands have been ditched to lower the water table and converted into agriculture, which contributes to anthropogenic greenhouse gas emissions. Hydrologic restoration of drained peatlands could offset greenhouse gas emissions from these actions, but field examples that consider various greenhouse gases are still rare. Here, we examined emissions of carbon dioxide (CO2), methane (CH4), and nitrous oxide (N2O) from soils in drained shrub bogs in North Carolina, USA, before and after hydrologic restoration. We used static chamber methods and a before-and-after, control-impact (BACI) experimental design. We found that hydrologic manipulation (akin to restoration) increased water table levels by 65%, even with the impact of two hurricanes before and one after hydrologic manipulation. Increased water table levels led to a 58% decrease in CO2 fluxes, and an increase in CH4 (251%) and N2O fluxes (85%). Water table depth and soil temperature explained 43% of variation in CO2, while water table depth explained 25% and 18% of variation in CH4 and N2O fluxes, respectively. Despite the increases in CH4 and N2O, the higher magnitude of fluxes and large decline in CO2 lead to an overall lowering of greenhouse gas emissions after hydrologic restoration. Our results suggest that raising the water table in this shrub bog peatland decreased overall greenhouse gas emissions, illustrating that hydrologic restoration of peatlands can be a valuable climate mitigation practice.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Peatlands play a disproportionate role in the global carbon cycle, storing 20–25% of the soil carbon in less than 3% of the land area (Reddy and DeLaune 2008). Peatlands store large amounts of carbon that has been identified as “irrecoverable”, meaning that if released to the atmosphere we will not be able to recover it in the time available to prevent the worst impacts of climate change (Goldstein et al. 2020; Noon et al. 2021). However, many peatlands have been ditched and drained and converted into agriculture (Leifeld and Menichetti 2018). Ditching and draining leads to lowering of water tables, facilitating oxidation, and leading to release of stored carbon (Neubauer and Megonigal 2021). It is estimated that 3% of all anthropogenic greenhouse gas emissions come from drained peatlands (Le Quéré et al. 2018; Evans et al. 2021). Efforts to preserve undrained peatlands or re-wet drained systems are intensifying in Europe (Leifeld and Menichetti 2018), but knowledge gaps and lack of adequate financial incentives have hindered similar efforts in the United States, particularly in the Southeast.
Peatlands once covered over 1.5 million hectares across the Southeastern U.S. coastal plain, from Virginia to northern Florida (Richardson 2003). Peatlands in the Southeast include pocosins, bogs, Carolina bays, and other forested wetlands (Batzer and Sharitz 2014). Large-scale drainage of peatlands in the Southeast for silviculture followed the example of Hofmann Forest, North Carolina, where large trials showed that pine tree productivity increased when ditches were installed (Terry and Hughes 1975; Fox et al. 2007). Subsequently, the demand for food and fiber resulted in widespread conversion via artificial drainage in the latter half of the twentieth century. It is estimated that by the mid-1980s drainage impacted approximately 1 million ha in the Southeastern coastal plain (Skaggs et al. 2016). Restoration of these systems began with government programs, like those at Pocosin Lakes National Wildlife Refuge, which used water control structures to raise water tables in an effort to recover the hydrology to pre-drainage conditions (Wang et al. 2015). Recently, there has been interest in peatland restoration as a way to decrease greenhouse gas emissions, and therefore serve as a climate change mitigation tool (Sleeter et al. 2017; National Academy of Sciences 2019).
Various environmental drivers affect the emissions of greenhouse gases from peatlands (Leifeld et al. 2019). Soil temperature and soil water table levels are usually the major drivers of greenhouse gas emissions in peatlands (Reddy and DeLaune 2008). Higher temperatures lead to higher emissions of greenhouse gases (primarily CO2 and CH4 (Updegraff et al. 2001; Bridgham et al. 2008). Higher water table levels increase CH4 fluxes from peatlands (Turetsky et al. 2014). Drainage usually leads to increased emissions of CO2, and modeling suggests that the global peatland biome went from carbon sink to a source in 1960, due to widespread hydrologic alterations (Leifeld and Menichetti 2018). Restoration of water levels in northern hemisphere peatlands could decrease greenhouse gas emissions by 0.3–3.4 Gt CO2eq (Huang et al. 2021). On the other hand, continued water table drawdown due to human activities and climate drying in the coming decades could increase CO2 emissions by 0.86 Gt CO2eq yr−1 (Leifeld and Menichetti 2018). In Pocosin Lakes National Wildlife Refuge, long-term drainage of shrub bogs led to changes in plant community structure and increased greenhouse gas emissions (Wang et al. 2015, 2021). Raising the water table and establishing a more natural hydrologic regime in these shrub bogs led to decreased CO2 fluxes, without large increases in CH4 (Wang et al. 2021). Nitrous oxide (N2O) is a powerful greenhouse gas that can also be controlled by changes in soil water content (Morse et al. 2012b), and has been shown to be higher in degraded peatlands (Liu et al. 2020). Research in a restored coastal plain wetland found that N2O fluxes increased in response to higher soil moisture (Morse and Bernhardt 2013), suggesting that raising water tables in peatlands could enhance N2O fluxes. However, the relative changes of all three gases in response to hydrologic manipulations have not been explored.
We examined the changes in the soil fluxes of CO2, CH4, and N2O from shrub bog peatlands before and after hydrologic restoration in Pocosin Lakes National Wildlife Refuge. We hypothesized that raising the water table (i.e. bringing the water table closer to the soil surface) would decrease CO2 emissions from manipulated sites by decreasing microbial oxidation of soil and plant derived carbon. We also hypothesized that raising the water table would increase CH4 and N2O emissions, but the increase in both gases would be smaller than the decrease in CO2 fluxes, leading to an overall decrease in greenhouse gases from restored sites. Finally, we hypothesized that water table depth and soil temperature would be important drivers of fluxes of all three gases.
Methods
Site Description
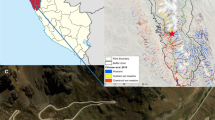
The study was conducted in the Pocosin Lakes National Wildlife Refuge (PLNWR) located in the Albemarle-Pamlico peninsula of North Carolina (35.7510°N, 76.5102°W, Fig. 1). The refuge contains more than 44,000 ha of shrub bog wetlands, also called pocosins (Richardson 1983). These shrub bog wetlands have deep organic soils (1–3 m depth), which tend to be nutrient poor and acidic (Richardson 2003). Average annual rainfall is approximately 1300 mm and mean annual temperature is 16.8 °C. Due to high temperatures and evapotranspiration, water table depths are usually more than 20 cm below the soil surface (Wang et al. 2015). The refuge was established in areas that had been drained for agriculture, and in 1990, management began blocking drainage canals to restore water levels. Vegetation in this area includes mature trees of pond pine (Pinus serotina Michx.), loblolly bay (Gordonia lasianthus (L.) Ellis), and swamp bay [Persea palustris (Raf.) Sarg.]. Common shrubs in this are include inkberry [Ilex glabra (L.) A. Gray], large gallberry [Ilex coriacea (Pursh) Chapm.], honeycup [Zenobia pulverulenta (W. Bartram ex. Willd.) Pollard], and fetterbush lyonia [Lyonia lucida (Lam.) K. Koch]. The western brakenfern [Pteridium aquilinum (L.) Kuhn] is common groundcover in in this area.

A Map of study sites in Pocosin Lakes National Wildlife Refuge (image from Google Earth, earth.google.com/web/). Inset shows location of the refuge in North Carolina. Restored sites are within red lines and non-manipulated controls are within blue lines. Blue circles denote location of sampling plots. Photos of water control structures (B, and C, photo credit E. Soderholm)
Our work was conducted in the Clayton Blocks Pocosin Restoration Project within PLNWR (Fig. 1). The project seeks to restore a more natural hydrologic regime to 536 ha of pocosin wetland, without impacting adjoining land uses. In order to hydrologically isolate the restoration area from adjacent drained private agricultural land, a new 4 km berm was built using borrow material from the excavation of new southern and western perimeter ditches just inside the berm. Three new water control structures (Fig. 1B and C) were then installed to raise the water table to pre-drainage levels, and allow for more natural seasonal variation of water level. The water control structures in the new ditches use flashboard risers to provide drainage control in the interior areas (Fig. 1). The structures also allow excess water to leave the project area and enter the downgradient drainage network, as would occur via sheetflow in intact pocosin wetlands. This is particularly important after large rainfall events, such as hurricanes, to reduce the potential of long-term inundation conditions that cannot be tolerated by pocosin plant communities. The berm, new ditch, and new water control structures were installed from July 2016 to March 2017, and water tables were raised in April 2017 (Fig. 2).

Water table level shown as meter below the soil surface in control and manipulated sites (A). Mean monthly water table depth in control sites (C) and hydrologically restored sites (M) before (Pre) and after (Post) hydrologic restoration (B). Data recorded hourly by continuous loggers. Dashed vertical line in A) indicates date of water table manipulation. Dashed horizontal line in A) and B) denotes soil surface
We sampled four plots within the Clayton Blocks: two plots within block D16 that served as non-manipulated controls (drained), one study plot each within blocks C13 and C14 that served as the manipulated sites (Fig. 1). We installed two wells (10 cm PVC, 1.2 m deep) in each site that were instrumented with water level recorders (model Level Troll 500, In-Situ Inc., Fort Collins CO, USA) to measure water table level. Loggers were set up to record water level hourly and were downloaded once a month. Hurricanes Hermine (September 2016), Matthew (October 2016), and Harvey (August 2017) delivered large amounts of rain to our sites during our sampling period. To record soil temperature, we installed soil temperature sensors (model Hobo Pro V2, Bourne MA, USA) approximately 2 m away from the wells at 10 cm depth to capture root zone dynamics. Soil temperature sensors recorded temperature hourly and were downloaded monthly.
Greenhouse Gas Sampling
We used the static chamber technique to measure fluxes of CO2, CH4, and N2O from soils (Livingston and Hutchinson 1995). In January of 2016, we installed four bases for chambers in each of the four sites to a depth of 5 cm. The bases remained in place for duration of the study. Chamber tops were made of a white, opaque polycarbonate material (29.4 × 29.4 × 30.5 cm) and lined with aluminum foil to restrict light transmission, and were fitted with septa for gas sample extraction (Krauss and Whitbeck 2011). To create an air tight seal during sampling, each base was designed with a 2 cm (W) × 2 cm (D) channel which was filled with water. Tops were installed just prior to sampling and removed immediately after. Prior to placing the chamber top, vegetation inside the chamber base was estimated (percent cover), and if necessary either cut to 9-inch height or bent over (dependent upon density). Plant growth within the chambers was categorized as present/absent, action taken (none, bent or cut) and amount of plant coverage [none, low (> 20%), medium (20–50%), high(> 50%)].
We sampled gases monthly (or every other month, depending on weather conditions) before (April, May, June, August, September, and October 2016) and after hydrologic restoration (April, June, July, September, and October 2017). Field sampling occurred between 10:00 and 15:00 each day of sampling. Gas samples were transferred from the headspace of the static chamber using a 10-mL plastic syringe/hypodermic needle at intervals of 0, 30, 60 min (2016) and intervals of 0, 20, 40, 60 min (2017). We added an extra sampling interval in 2017 to try better capture CH4 flux dynamics. We did not observe CO2 concentrations to level out during the 60-min incubations, suggesting that the incubations were not long enough to change air pressure. During each sampling, we transferred the individual 10 mL gas samples into 3.7 mL Labco Exetainers fitted with double septa caps (Lampeter, United Kingdom) and stored in Whirl Pak bags inside of Ziploc bags for transport. CO2, CH4 and N2O concentrations of the gas samples were analyzed using a gas chromatograph (model Shimadzu GC2014, Durham, North Carolina, USA). CH4 and CO2 were measured using a flame ionization detector (FID) and N2O using an electron capture detector (ECD). Gas samples were analyzed within 72 h of collection. Fluxes were calculated from the linear change in gas concentrations as a function of time, chamber volume, collar area, and air temperature. Individual chamber flux calculations with an R2 less than 0.85 were not included in statistical analyses. We averaged fluxes for each of the four chambers in each sampling site and date.
Statistical Analyses
We used a replicated before-after, control-impact design (BACI), taking gas flux measurements before and after hydrologic restoration in both manipulated and non-manipulated (control) sites. We used one way ANOVAs to compare soil temperature and water level across sites, and before and after manipulation. Because gas fluxes were not normally distributed, we used non-parametric Kruskal–Wallis test to compare fluxes pre and post manipulation in manipulated and control sites. We predicted that raising the water table has a strong effect on fluxes, therefore, we would observe differences in the manipulated plots and not the control plots. We also used simple linear regression models on log transformed fluxes (and we added 100 to all fluxes to remove negative values) to examine environmental controls (soil temperature, water table depth, and their interaction) on fluxes of all three gases. We used a cut off for p > 0.05. To examine if hydrologic manipulation increased emissions of all three gases, we calculated cumulative fluxes before and after hydrologic restoration. For CH4 and N2O we used the sustained flux global warming potential (Neubauer and Megonigal 2015) to estimate CO2 equivalents.
Results
Water table depth (m below soil surface) fluctuated in response to hurricanes and increased after hydrologic manipulation (Fig. 2). Water levels in the two manipulated sites (C13 and C14) were lowest in 2016 (-1.0 m) and were highest after restoration (0.06 and 0.16 m). Mean water levels increased (meaning water table was closer to the soil surface) by 65% after hydrologic restoration in the manipulated sites (F1, 51 = 35.50, p < 0.001), but did not change in the control sites (D16A and B, Fig. 2). Mean soil temperature ranged from 20 to 22 °C, but did not differ across sites. Soil temperature was higher in 2017 (22.4 °C) than 2016 (21.05 °C, F1,51 = 3.95, p = 0.05).
The majority of the flux calculations for CO2 fluxes met requirements for including into statistical analyses (94% of incubations had regressions with R2 > 0.85). Mean CO2 fluxes in control sites were similar (270.2 ± 38 and 279.1 ± 40 mg CO2 m−2 h−1 in pre and post manipulation sampling periods, respectively; Fig. 3A and D). Mean CO2 fluxes in manipulated sites declined from 793.2 ± 171 mg CO2 m−2 h−1 to 327 ± 170 mg CO2 m−2 h−1 after manipulation (58% decline, χ2 = 3.86, p = 0.04, Fig. 3A and D). There was no difference in CO2 fluxes across cover of plants, or action taken (none, bent, or cut), however low power might have prevented us from detecting a difference (majority of measurements had high cover plants and vegetation was bent). A linear model including average daily water table depth and soil temperature explained 43% of the variation in CO2 fluxes (Table 1), with CO2 fluxes increasing with higher water table depth and higher temperatures (Fig. 4A, B).

Boxplots (A, B, and C) and time series (D, E, and F) of CO2 (A and D), CH4 (B and E), and N2O (C and F) fluxes in control plots (C-) and hydrologically manipulated (M-), before (Pre) and after (Post) restoration. For the time series (D, E, and F) the black line denotes the post-manipulation period

Linear regressions of log transformed fluxes of CO2 (A, B), CH4 (C, D) and N2O (E, F) versus water table depth (meters, A, C, E) and soil temperature (B, D, F). Table 1 reports model parameters
Many of the flux calculations for CH4 did not meet our minimum requirements, only 37% of flux calculations had an R2 > 0.85. Mean CH4 fluxes in the control sites were not significantly different: 28.3 ± 23 and 6.90 ± 24 μg CH4 m−2 h−1 in pre and post manipulation sampling periods, respectively (Fig. 3B and E). Mean CH4 fluxes in the manipulated sites were 25.8 ± 30 and 90.7 ± 32 μg CH4 m−2 h−1 in pre and post manipulation sampling periods respectively, a 251% increase in CH4 fluxes (χ2 = 3.52, p = 0.06). However, this increase in CH4 fluxes was mainly driven by a single date in October 2017, which was much higher than all other measurement (433 μg CH4 m−2 h−1, Fig. 3E). If we exclude that measurement, the mean CH4 flux for the manipulated sites in the post-manipulation period is 47.8 ± 15 μg CH4 m−2 h−1, which is still higher, but the difference is smaller (85% increase, χ2 = 2.5, p = 0.10). The CH4 fluxes were negatively associated with water table depth and the interaction of water table depth and soil temperature (R2 = 0.25, Table 1, Fig. 4).
The majority (71%) of flux calculations for N2O were included in analyses. Mean N2O fluxes in control sites were not significantly different: 74.6 ± 15 and 41.7 ± 14 μg N2O m−2 h−1 for pre and post manipulation sampling periods, respectively (Fig. 3C and F). Mean N2O fluxes in manipulated sites were 112.8 ± 72 and 221.0 ± 70 μg N2O m−2 h−1 for pre and post-manipulation sampling periods, which is a 96% increase and not statistically significant (χ2 = 2.1, p = 0.14, Fig. 3C and F). Lower water table levels were related to higher N2O fluxes (R2 = 0.18, Table 1, Fig. 4E and F).
Cumulative fluxes of all three gases expressed as CO2 equivalents were similar in the control plots pre and post restoration (Fig. 5). Cumulative fluxes were highest in the manipulated sites before hydrologic restoration, and decreased after restoration (Fig. 5). Even though CH4 and N2O increased slightly, this was not enough to compensate for the large decline in CO2 fluxes (Fig. 5).

Cumulative greenhouse gases fluxes (in CO2 equivalents) in control (C-) and manipulated (M-) plots before (Pre) and after (Post) restoration for all three gases (A) and just CH4 and N2O (B)
Discussion
Hydrologic restoration of pocosin peatlands through water control structures was successful and increased water levels by 65% (Fig. 2). In support of the first hypothesis, CO2 emissions in the manipulated sites declined by 58% after hydrologic restoration (Fig. 3A). In support of the second hypothesis, both CH4 (85–251%) and N2O (96%) fluxes increased after restoration in the manipulated sites (Fig. 3B and C), though the differences were not significant. While we cannot fully attribute the changes in fluxes to hydrologic restoration, two lines of evidence suggest that increasing water levels led to the changes we observed. First, the control plots did not exhibit any significant changes in fluxes in any of the three gases between the pre and post manipulation period (Fig. 3). Second, the strong correlation of all three gases with water table depth suggests that raising the water table was an important driver of fluxes in the manipulated sites (Fig. 4). We also found that the overall decrease in CO2 overcame the increase in CH4 and N2O even while accounting for their different global warming potentials (Fig. 5), suggesting that hydrologic restoration can lead to an overall reduction in the climate forcing of these drained peatlands.
CO2Emission Dynamics
The range of CO2 fluxes measured in the two years of this study (85–2500 mg CO2 m−2 h−1) were similar to rates reported in nearby mature forested wetlands and a restored wetland (150–2000 mg CO2 m−2 h−1, Morse et al. 2012a). Our fluxes were also similar to those reported in previous studies in similar systems: in natural, drained, and restored pocosin sites in Pocosin Lakes Wildlife Refuge (100–1200 mg CO2 m−2 h−1, Wang et al. 2015), and peatland sites in Great Dismal Swamp National Wildlife Refuge (20–2000 mg CO2 m−2 h−1, Gutenberg et al. 2019). The highest fluxes we observed were in the manipulated sites before restoration (Fig. 3A), which were the sites with the lowest water levels (Fig. 2A). This agrees with results in a Canadian peatland where CO2 fluxes were highest when water table depth was lowest (70 cm below soil surface, Moore and Knowles 1989). These results suggest that hydrologic restoration of pocosins may be most effective in areas where water tables are more than 70 cm below the surface, which will likely provide the largest reduction in CO2 fluxes.
It is interesting that the CO2 fluxes in the control sites did not increase between the pre and post manipulation period, even though 2017 (the post manipulation period) was on average one degree warmer. The slope of the linear regression model suggests that CO2 should have increased on average by 5 mg CO2 m−2 h−1 (Table 1), which could have been missed with our temporal sampling regime. The decrease in CO2 from the manipulated sites after hydrologic restoration suggest that CO2 fluxes from these systems are more sensitive to water table depth than temperature, which agrees with findings from peatlands in Europe (Evans et al. 2021). Future changes in hydrologic regimes might have stronger impacts on emissions from peatlands than warming.
CH4Emission Dynamics
CH4 fluxes in this study were lower (-29 to 433 μg CH4 m−2 h−1) compared to nearby forested wetlands (0 to 8500 μg CH4 m−2 h−1, Morse et al. 2012a). Our CH4 fluxes were also lower than those measured in pocosin wetlands in Great Dismal Swamp (-5.18 to 3000 μg CH4 m−2 h−1), but similar to fluxes measured in nearby pocosin sites (median 5 to 150 μg CH4 m−2 h−1, Wang et al. 2021). However, the Wang et al. (2021) did report fluxes as high as 45,000 μg CH4 m−2 h−1, which occurred in undrained pocosins when water levels were less than 5 cm from the soil surface, during the summer. The elevated value of CH4 flux we observed in the post restoration period (433 μg CH4 m−2 h−1), and the low success rates of our incubations to estimate CH4 fluxes (37%) also agree with previous literature that CH4 fluxes can be highly variable in space and time (Turetsky et al. 2014; Wang et al. 2021). Our results support the Wang et al. (2021) suggestion that rewetting pocosin peatlands will not lead to significant CH4 fluxes, as long as water tables are maintained 5–10 cm below the soil surface.
N2O Emission Dynamics
N2O fluxes are hard to forecast, because they are the product of various processes, mainly nitrification and denitrification (Morse et al. 2012a; Morse and Bernhardt 2013). N2O fluxes in our sites (-16 to 840 μg N2O m−2 h−1) were also similar to two forested wetlands and a restored wetland (0–1000 μg N2O m−2 h−1, Morse et al. 2012a). The highest fluxes of N2O observed occurred in September 2016 (450 μg N2O m−2 h−1) and September 2017 (840 μg N2O m−2 h−1) in the manipulated sites, which were months when the water table was close to the soil surface due to a hurricane in 2016 and manipulations in 2017 (Fig. 2A). The fact that the highest N2O fluxes occurred when the water table was close to the surface, but not above it, could be explained because N2O is the product of incomplete anaerobic (denitrification) and aerobic (nitrification) processes (Morse and Bernhardt 2013), so it is important to have variable redox conditions. Finally, N2O fluxes were higher and contributed more to the overall warming potential of these systems than CH4 (Fig. 5), which surprised us. Previous studies have reported low levels of CH4 in these pocosin peatlands, which has been attributed to high levels of phenolics due to the vegetation (Wang et al. 2021). The high phenolics can inhibit methanogenesis and serve as alternate electron acceptors (Keller and Bridgham 2007). There are various possible explanations for the relatively high N2O fluxes: 1) drainage led to increased N mineralization (Wang et al. 2015); 2) legacy of past agricultural use in these pocosin peatlands; and/or 3) high N deposition from the adjacent farm. These three possible explanations are not mutually exclusive, and illustrate that N2O fluxes may be considerations for future projects when planning restoration of peatlands as a climate mitigation tool.
Conclusions
Our results show that manipulating water levels in drained peatlands in the Southeast can decrease overall greenhouse gas emissions. We estimate that restoration led to decreased emissions by 9.6 tons of CO2 equivalents per acre over the first year after restoration. It is estimated that restoration of drained peatlands in Virginia, North Carolina, and South Carolina could avoid up to 1.5 Tg CO2 emissions over the coming decades (Fargione et al. 2018). Our results illustrate that these CO2 reductions outweigh potential negative impacts from increased CH4 and N2O emissions, which is similar to what has been found in northern peatlands (Liu et al. 2020; Evans et al. 2021). Increasing water table levels in drained peatlands also reduces the probability of catastrophic wildfire (Poulter et al. 2006). These changes to hydrology can also help change vegetation over longer periods of time (Wang et al. 2015), which can provide other ecosystem benefits, such as habitat for enhancing biodiversity. Given the large extent of peatland drainage in the Southeast, hydrologic restoration can provide multiple benefits and be a valuable tool in mitigating climate change.
Data Availability
The datasets generated during and analyzed during the current study are available from the corresponding author on reasonable request.
References
Batzer DP, Sharitz RR (2014) Ecology of Freshwater and Estuarine Wetlands, 2nd Editio. University of California Press, Oakland
Bridgham SD, Pastor J, Dewey B et al (2008) Rapid carbon response of peatlands to climate change. Ecology 89:3041–3048
Evans CD, Peacock M, Baird AJ et al (2021) Overriding water table control on managed peatland greenhouse gas emissions. Nature 593:548–552. https://doi.org/10.1038/s41586-021-03523-1
Fargione JE, Bassett S, Boucher T et al (2018) Natural climate solutions for the United States. Science Advances 4:1–15. https://doi.org/10.1126/sciadv.aat1869
Fox TR, Jokela EJ, Allen HL (2007) The development of pine plantation silviculture in the Southern United States. Journal of Forestry 105:337–347. https://doi.org/10.1093/jof/105.7.337
Goldstein A, Turner WR, Spawn SA et al (2020) Protecting irrecoverable carbon in Earth’s ecosystems. Nature Climate Change 10:287–295. https://doi.org/10.1038/s41558-020-0738-8
Gutenberg L, Krauss KW, Qu JJ et al (2019) Carbon Dioxide Emissions and Methane Flux from Forested Wetland Soils of the Great Dismal Swamp, USA. Environmental Management 64:190–200. https://doi.org/10.1007/s00267-019-01177-4
Huang Y, Ciais P, Luo Y et al (2021) Tradeoff of CO2 and CH4 emissions from global peatlands under water-table drawdown. Nature Climate Change 11:618–622. https://doi.org/10.1038/s41558-021-01059-w
Keller JK, Bridgham SD (2007) Pathways of anaerobic carbon cycling across an ombrotrophic-minerotrophic peatland gradient. Limnology and Oceanograhy 52:96–107
Krauss KW, Whitbeck JL (2011) Soil Greenhouse Gas Fluxes during Wetland Forest Retreat along the Lower Savannah River, Georgia (USA). Wetlands 32:73–81. https://doi.org/10.1007/s13157-011-0246-8
Le Quéré C, Andrew RM, Friedlingstein P et al (2018) Global Carbon Budget 2017. Earth System Science Data 10:405–448. https://doi.org/10.5194/essd-10-405-2018
Leifeld J, Menichetti L (2018) The underappreciated potential of peatlands in global climate change mitigation strategies. Nature Communications 9:1071
Leifeld J, Wüst-Galley C, Page S (2019) Intact and managed peatland soils as a source and sink of GHGs from 1850 to 2100. Nature Climate Change 9:945–947. https://doi.org/10.1038/s41558-019-0615-5
Liu H, Wrage-Mönnig N, Lennartz B (2020) Rewetting strategies to reduce nitrous oxide emissions from European peatlands. Communications Earth and Environment 1:1–7. https://doi.org/10.1038/s43247-020-00017-2
Livingston GP, Hutchinson GL (1995) Enclosure based measurement of trace-gas exchange: applications and sources of error. In: Matson PA, Harriss RC (eds) Biogenic trace gases: measuring emissions from soil and water. Blackwell Science, Cambridge, Massachusetts, pp 14–51
Moore T, Knowles R (1989) The influence of water table levels on methane and carbon dioxide emissions from peatland soils. Canadian Journal of Soil Science 69:33–38. https://doi.org/10.4141/cjss89-004
Morse JL, Ardon M, Bernhardt ES (2012a) Greenhouse gas fluxes in southeastern US coastal plain wetlands under contrasting land uses. Ecological Applications 22:264–280
Morse JL, Ardón M, Bernhardt ES (2012b) Using environmental variables and soil processes to forecast denitrification potential and nitrous oxide fluxes in coastal plain wetlands across different land uses. Journal of Geophysical Research: Biogeosciences https://doi.org/10.1029/2011JG001923
Morse JL, Bernhardt ES (2013) Using 15N tracers to estimate N2O and N2 emissions from nitrification and denitrification in coastal plain wetlands under contrasting land-uses. Soil Biology & Biochemistry 57:635–643
National Academy of Sciences, Engineering, and Medicine (2019) Negative Emissions Technologies and Reliable Sequestration: A Research Agenda. National Academies Press, Washington D.C., USA https://doi.org/10.17226/25259.
Neubauer SC, Megonigal JP (2021) Biogeochemistry of wetland carbon preservation and flux. Wetland carbon and environmental management. In: Krauss KW, Zhu Z, Stagg CL (eds) Geophysical monograhp series. AGU. https://doi.org/10.1002/9781119639305.ch3
Neubauer SC, Megonigal JP (2015) Moving Beyond Global Warming Potentials to Quantify the Climatic Role of Ecosystems. Ecosystems 18:1000–1013. https://doi.org/10.1007/s10021-015-9879-4
Noon ML, Goldstein A, Ledezma JC et al (2021) Mapping the irrecoverable carbon in Earth’s ecosystems. Nature Sustainability. https://doi.org/10.1038/s41893-021-00803-6
Poulter B, Christensen NL, Halpin PN (2006) Carbon emissions from a temperate peat fire and its relevance to interannual variability of trace atmospheric greenhouse gases. Journal of Geophysical Research-Atmospheres D06301 https://doi.org/10.1029/2005jd006455
Reddy KR, DeLaune RD (2008) Biogeochemistry of wetlands: Science and applications. CRC Press, Boca Raton, FL
Richardson CJ (2003) Pocosins: Hydrologically isolated or integrated wetlands on the landscape? Wetlands 23:563–576
Richardson CJ (1983) Pocosins: Vanishing wastelands or valuable wetlands? Bioscience 33:626–633
Skaggs RW, Tian S, Chescheir GM et al (2016) Forest drainage. In: Amatya DM, Williams TM, Bren, L. Jong C de (eds) Forest Hydrology: processes, management and assessment. CABI Publisher, U.K. ebooks, p 124
Sleeter R, Sleeter BM, Williams B et al (2017) A carbon balance model for the great dismal swamp ecosystem. Carbon Balance Management 12:2. https://doi.org/10.1186/s13021-017-007-4
Terry TA, Hughes JH (1975) The effects of intensive management on planted loblolly pine (Pinus taeda L.) growth on poorly drained soils of the Atlantic Coastal Plain. Forest soils and forest land management, Proceeding of the fourth North American Soils Conference. Les Presses de l’Universite Laval, Quebec, Canada, pp 351–377
Turetsky M, Kotowska A, Bubier J et al (2014) A synthesis of methane emissions from 71 northern, temperate, and subtropical wetlands. Global Change Biology 20:2183–2197
Updegraff K, Bridgham SD, Pastor J et al (2001) Response of CO2 and CH4 emissions from peatlands to warming and water table manipulation. Ecological Applications 11:311–326
Wang H, Ho M, Flanagan N, Richardson CJ (2021) The Effects of Hydrological Management on Methane Emissions from Southeastern Shrub Bogs of the USA. Wetlands 41:87. https://doi.org/10.1007/s13157-021-01486-7
Wang H, Richardson CJ, Ho M (2015) Dual controls on carbon loss during drought in peatlands. Nature Climate Change 5:584–587. https://doi.org/10.1038/NCLIMATE2643
Acknowledgements and Funding
This work was supported by funding from The Nature Conservancy and NSF DEB-1713502. KWK, NC, and RFM were supported by the U.S. Geological Survey Land Carbon Program and Climate R&D Program.
Author information
Authors and Affiliations
Contributions
Christine Pickens, Ken W. Krauss, and Marcelo Ardón conceived and designed the study. Louise Armstrong, Ariane Peralta, Nicole Cormier, Rebecca F. Moss, Eric Soderholm, Aaron McCall, Christine Pickens and Marcelo Ardón participated in material preparation, data collection, and analyses. The first draft of the manuscript was written by Louise Armstrong and Marcelo Ardón, and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing Interests
Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the U.S. Government. The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Armstrong, L., Peralta, A., Krauss, K.W. et al. Hydrologic Restoration Decreases Greenhouse Gas Emissions from Shrub Bog Peatlands in Southeastern US. Wetlands 42, 81 (2022). https://doi.org/10.1007/s13157-022-01605-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13157-022-01605-y