
Abstract
In recent decades, invasive shrubs have replaced herbaceous wetlands in many parts of the world. In Florida, the native shrub Salix caroliniana Michx. (Carolina willow) expanded its distribution throughout the upper St. Johns River, replacing herbaceous marshes with willow swamps. To identify ways to prevent its expansion, we experimentally tested the effects of watering regime, temperature, substrate, and seed source on willow germination and seedling survival. In growth chamber experiments, germination and survival were most affected by watering regime and were greatest in saturated, organic soils. Survival decreased with soil inundation and on drier, sandy soils. Variable texture and nutrient content in native soils had no differential effect on germination or survivability of willow. Time of seed production, seed source, and delay in watering significantly affected germination. Seed germination occurred quickly after being sown. However, seed viability declined just as quickly. Whenever a soil held sufficient water, especially through capillarity, seeds of Carolina willow germinated and survived well. Seasonal manipulation of water levels to flood marshes during seed-fall and to inundate willow seedlings provides managers with an effective strategy for reducing establishment of Carolina willow.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Despite a national policy of no net loss of wetlands, the conterminous USA lost 260,700 ha (644,000 acres) of wetlands between 1986 and 1997 (Dahl 2000). Largely overlooked within Dahl’s (2000) status and trends report was an increase in freshwater shrub wetlands and a concomitant decline in freshwater emergent marshes. These two trends are causally linked wherever freshwater shrubs displace herbaceous marsh vegetation. Wetland plants are among the most notorious invasive species (Zedler and Kercher 2004), and exotic shrubs such as Melaleuca quinquenervia (melaleuca), Mimosa pigra (catclaw mimosa), Schinus terebinthifolius (Brazilian peppertree), and Tamarix ramosissima (salt cedar) are listed among the world’s “100 Worst Invasive Alien Species” (Lowe et al. 2004). Replacement of herbaceous marshes by near-monocultures of invasive shrubs generally decreases biodiversity, alters nutrient retention and sedimentation patterns, lowers the water table, reduces surface water flows and stream braiding, and may have allelopathic effects on native flora (Gordon 1998). However, these negative ecosystem consequences of wetland invasion are not limited to exotic shrubs; in central Florida, USA, the native Carolina willow (Salix caroliniana Michx.) has become invasive where human activities altered marsh nutrients, hydrology, and disturbance regimes.
During the last five decades, Carolina willow encroached into areas within the upper St. Johns River basin (USJRB) of central Florida that historically were herbaceous marsh (Lowe et al. 1984; Hall 1987; Kinser et al. 1997; Miller et al. 1998), rapidly colonizing newly created mudflats (Rodgers 1989; Southall et al. 2003; Ponzio et al. 2006). Coincidentally, an extensive network of levees and canals facilitated agricultural production and flood control (SJRWMD 2007), which by 1984 eliminated approximately 38 % of the USJRB floodplain wetlands (Lowe et al. 1984). Willow expansion transformed herbaceous marshes, wet prairies, and sloughs into nearly monospecific, willow shrub swamps (Ponzio et al. 2006). While the exact conditions favoring willow invasions are unknown, they have been attributed to altered hydroperiods, reduced fire frequency, nutrient increases, and disturbance caused by agriculture and development (Zukowski and Gawne 2006; Ponzio et al. 2006; Quintana-Ascencio et al. 2013).
Like many invasive wetland shrubs, Carolina willow has a combination of life-history traits, including both sexual and vegetative reproduction, which enables rapid colonization of disturbed areas (Young and Clements 2003). Carolina willow produces very large numbers of small, hairy seeds that disperse by wind and running water, do not exhibit dormancy, and germinate rapidly (Niiyama 1990; Titus 1991; Karrenberg et al. 2002; Hanselman et al. 2005). Saturated soils in exposed drawdown zones provide a favorable environment for its germination and vegetative expansion (Tallent-Halsell and Walker 2002; Kuzovkina and Quigley 2005). Combined with its ability to withstand flooding (once established) by producing adventitious roots and lenticels (Reyes 2012), these life-history traits allow Carolina willow to rapidly colonize and persist in wetland ecosystems.
Effectively controlling invasive wetland shrubs requires an understanding of the factors affecting their early life-history stages, particularly water availability, soil characteristics and nutrients, temperature, and genetic variation (e.g., Taylor and McDaniel 1998; Sher et al. 2002; Ainouche et al. 2009). We explored the importance of these factors to germination and early survival of Carolina willow in three experiments conducted in a growth chamber. In Experiment 1, we manipulated watering regime; in Experiment 2, we tested the effect of different soils and seed source; and in Experiment 3, we assessed the influence of temperature regime, production timing, delay in watering, and seed source. In all three experiments, we quantified the effects of treatments on germination and early survival of Carolina willow. Our objective was to identify conditions limiting willow recruitment and establishment and to provide information for willow management and marsh restoration, both in the USJRB, and elsewhere in the world where Salix species are invasive. For example, Salix babylonica, S. cinerea, and S. humboldtiana are listed as invasive alien species in Australia (GISD 2013), where a national task force was established to control invasive willows.
Methods
Seed Collection
We collected mature catkins of Carolina willow at the peak of its March-April reproductive season in the USJRB (Fig. 1). We pulled whole seeds covered with woolly hairs from opened catkins, and placed them into a plastic bag. We inflated, closed, and shook the bag to remove the woolly hairs, and sorted the released seeds. We selected only green seeds for all experiments because rough, brown seeds were non-viable (L. Castro-Morales, pers. obs.). Seeds were sown within 24 h of collection except as indicated below.

Sites of gray up pointing triangle seed and black circle soil sampling within the upper St. Johns River basin, Florida, USA: Blue Cypress Marsh Conservation Area (BC), River Lakes Conservation Area (RL), and St. Johns Marsh Conservation Area (SJ)
Experiment 1: Watering Regime
The USJRB has a humid subtropical climate, with a distinct dry season in winter and a wet season in late spring to early fall. Water levels fluctuate dramatically and expose willow seeds to the entire range of drought to flood conditions. In certain areas, water level can be manipulated for restoration and management (Quintana-Ascencio et al. 2013). We, therefore, evaluated the effect of water regimes on germination and seedling survival of Carolina willow. Experiment 1 had six hydrologic treatments that were replicated six times. Hydrologic treatments were: 1) continuous flooding, with water maintained 5 cm above the soil surface; 2) saturated soil continuously kept moist by capillarity, by keeping the outside container always filled with 2 cm of water; 3) soil watered daily; 4) soil watered once every 3 d; 5) soil watered once every 5 d; and 6) sandy soil watered once every 8 d. These six treatments represented the range of hydrologic conditions that willow seeds experience in the field, from inundated soils to extreme drought.
On March 7, 2009, we filled 420 ml plastic pots with ~90 g of commercial potting soil (Earthgrow potting soil ®, Scotts Company, Marysville OH, USA) or with a native, sandy soil collected from St. Johns Marsh Conservation Area. We then moistened pots to field capacity with tap water and placed each pot into a separate 946 ml plastic container. We scattered 20 seeds of Carolina willow, collected on March 6th, onto the soil surface of each pot. Immediately after sowing, we applied one of the six hydrologic treatments. We established six replicates per treatment (total N = 36) and kept pots in a CMP 4030 plant growth chamber (CONVIRON, Canada). We rearranged pots weekly to minimize spatial bias. We set temperature conditions to simulate early field germination season using winter temperature data and daily light schedules (with variable fluorescent 30 W bulbs and incandescent 60 W bulbs) from Archbold Biological Station, Lake Placid, Florida. We recorded germination and survival daily for 30 d. Germination was defined as appearance of cotyledons.
Experiment 2: Soil and Seed Source
The USJRB includes a wide range of soil types, from deep, organic muck to inorganic, sandy soils (USDA 1974). Agriculture and human development have contributed excess nutrients to some sites, forming soils with different combinations of texture and nutrients. The USJRB also spans 125 km from north to south, creating the potential for local variation in willow demographic parameters. We therefore evaluated germination responses of Carolina willow to soil type/nutrient concentrations and seed source.
Experiment 2 was also implemented in the CMP 4030 growth chamber and had a 6 × 4 factorial design that was replicated 5 times. Treatments were all possible combinations of six soils crossed with four seed sources (i.e., 24 treatment combinations and five replicates). Soils originated from one of three different sites (Fig. 1): Gator muck (Gator Soil Series, USDA 1989: bulk density 0.04 g/cm3, organic matter 91–93 %), a very poorly drained, thick loamy soil from Blue Cypress Marsh Conservation Area (BC); Montverde peat (Montverde Soil Series, USDA 1974: bulk density ~0.6 g/cm3 [approximated value obtained for Micco peat, a similar soil in the same area], organic matter 70 %), a poorly drained, thick organic soil from River Lakes Conservation Area (RL); and Floridana sand (Floridana Soil Series, USDA 1990; bulk density 1.1 g/cm3, organic matter 2 %), a poorly drained, sandy soil from St. Johns Marsh Conservation Area (SJ).
We followed standard procedures for soil collection and handling and analyzed soil porewater nutrients of these three soils following EPA guidelines (O’Dell 1993 a,b,c; Martin et al. 1994). Deionized water was added to soil samples with a dilution factor of 10:1 for peat and 2:1 for sand, respectively. Soil/water slurries were shaken for 1 h and centrifuged for 10 min at 6,000 rpm to extract diluted porewater samples. The supernatant was filtered using a 0.45 mm membrane filter with a vacuum filtration apparatus. Porewater samples were preserved, as appropriate, and sent to the laboratory for analysis. Nutrient concentrations in the porewater of these soils were: 0.095 + 0.005 mg/L NOx- (mean + s.d., n = 3), 0.047 + 0.023 mg/L NH4+, 0.105 + 0.012 mg/L PO4-, and 0.326+ 0.016 mg/L K+ for pure BC soil; 0.064 + 0.006 mg/L NOx-, 0.148 + 0.031 mg/L NH4+, 0.086 + 0.011 mg/L PO4-, and 0.105 + 0.014 mg/L K+ for pure RL soil; and 0.055 + 0.004 mg/L N0x-, 0.249 + 0.06 mg/L NH4+, 0.114 + 0.011 mg/L PO4-, and 0.230 + 0.034 mg/L K+ for pure SJ soil. We created three more soil treatments using 1:1 mixtures of the three original soils, to explore willow responses along the gradient of soil textures and nutrients.
On March 27, 2009, we filled pots with soils that varied in texture and nutrient concentration. We moistened all pots to capacity with tap water and planted 30 seeds of Carolina willow collected on March 25, 2009 from one of four sites in the USJRB (Fig. 1). As in Experiment 1, we rearranged pots weekly to minimize spatial bias and used identical temperature conditions. We watered as needed to maintain the outside containers with tap water 2 cm deep, thereby watering soil interstices by capillarity. This was the best germination condition in Experiment 1. We again recorded germination and survival daily for 30 d.
Experiment 3: Temperature, Seed Source, Production Timing, and Delay in Watering (or Initiation of Watering)
Our third growth chamber experiment investigated fine-scale variation in the timing of willow germination. Detailed information about timing is vital for controlling willows, which often release seeds during a small window of conditions favorable for germination (e.g., Stromberg et al. 2007). In Central Florida, Carolina willow has a short flowering season, ranging from late winter (late February) to early spring (April). We, therefore, assessed the effect of temperature regime, seed source, time of production, and time to watering (loss of viability, see below) on germination success. A complete factorial design this size was impractical, so Experiment 3 had a nested design, where outer treatments were one of two temperatures (simulated northern and southern temperature regimes), and seeds were collected at one of three times (early, middle, and late flowering seasons). Nested within these outer factors, we crossed two seed sources (northern and southern locations) with six times to watering (0, 2, 4, 6, 8 or 10 d after collection). Each of these 12 inner factors were replicated three times and the total number of Petri dishes was 216.
We collected seeds of Carolina willow from the southern (State Route [SR] 60) and northern (SR 50) regions of the USJRB (Fig. 1) during the 2010 reproductive season. Because reproduction begins earlier in the south, we collected seeds on an overlapping schedule: southern region, early (March 11), middle (March 19), and late (April 8); and northern region, early (March 19), middle (April 8) and late (April 22). On each collection date, we gathered seeds from three different willows within each region. We pooled seeds within regions and placed 30 seeds onto filter paper in plastic Petri dishes (55 mm diameter × 15 mm depth) within the CMP 4030 growth chamber and rearranged them weekly to minimize spatial bias. We randomly distributed Petri dishes into two chambers, which had two different temperature regimes that represented the spring temperature of the USJRB in either the northern (Titusville, FL) or southern region (Lake Wales, FL). We then applied one of six different watering treatments: seeds were watered (with tap water) the same day they were collected, or 2, 4, 6, 8 or 10 d after collection. We then collected germination data daily for 16 d.
Data Analysis
In Experiments 1 and 2, we compared minimum time elapsed for germination (Min lag), mean time elapsed for germination (Mean lag), time elapsed for 50 % of seeds to germinate (Response time T50), maximum number of seeds germinated in a day (Max germination), and total proportion germinating (G) among treatments (e.g., Barnea et al. 1991; Shipley and Parent 1991; Ponzio 1998; Traveset et al. 2001). We used survival analysis models with constant (exponential) and variable (Weibull) hazards to evaluate germination and survival schedules (Crawley 2007). One-way ANOVA was performed to test for differences in survival. We used Tukey’s HSD as a post hoc test to identify which means differed significantly. We inspected model residuals and found no evidence of significant departures from homoscedasticity and normal error distributions. The Type I error rate for all tests was α = 0.05. Analyses were performed using R (R Development Core Team 2011).
In Experiment 3, we used nested ANOVA to test the effects of seed source, production timing, temperature regime, and initiation of watering regimes on germination. Temperature regime in the growth chamber and production timing were the between-effects in this analysis; the two factors nested within were seed source and initiation of watering (Table 3).
Results
Experiment 1: Watering Regime
Most seeds germinated 3 – 5 d after the start of the experiment (Tables 1 and 2; Figs. 3 and 4). Percent germination on commercial potting soil and native sand was moderate (range 25–63 %) in all treatments (Table 1). The variable hazard Weibull model better represented the germination schedule than the exponential model because most germination occurred rapidly, generally during the first 5 d (Fig. 2). Mean lag time, response time T50, and maximum germination rate did not differ significantly among treatments (Table 1). Across all treatments, mean lag time was 4.6 ± 0.17 d (hereinafter, mean ± 1 SE), response time T50 was 3.8 ± 0.12 d, and maximum germination rate was 5.2 ± 0.41 seeds/d (out of 20 seeds). However, the total proportion of seeds germinating (0.63 ± 0.09) was significantly higher for seeds sown in the saturated treatment on commercial potting soil than for those on sand watered every 8 d. The proportion of seedlings that survived 30 d in the saturated treatment (0.89 ± 0.03; Fig. 2b) was significantly higher than all other treatments (all Z ≥ 2.88, p ≤ 0.004). No seedlings survived to 30 d in the driest treatment: sand with 8 d between watering.

Mean (± 1 SD) proportion of Carolina willow germinating (a) and proportion of germinated seeds that survived (b) under different watering regimes in Experiment 1. * indicates a treatment that differed significantly (p < 0.05) from all others
Experiment 2: Soil and Seed Source
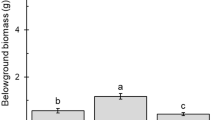
The total proportion of seeds germinating on soils collected from natural sources was higher (0.90 – 1.00) than on the commercial potting soil used in Experiment 1. Germination occurred rapidly and a Weibull model again best described the germination schedule (Fig. 3a). Minimum lag time, mean lag time, response time T50, maximum germination rate, and total proportion germinating did not differ significantly among soil types (all F5,18 < 0.96, p > 0.49), but mean lag time and total proportion germinating varied among seed sources (Table 2). Mean lag time ranged from 3.4 to 4.8 d, mean T50 from 3.0 to 3.5 d, and maximum germination rate from 18.8 to 23.8 seeds/d (total seeds = 30; Fig. 3a) among seed sources. Mean lag time was shorter and total proportion germinating was higher for seeds from northern sources (SR 46 and SR 50; Table 2; Fig. 3b). There was no support for overall differences among soils or seed sources in total proportion of seedlings surviving (both F < 1.01, p > 0.45), which averaged 0.84 ± 0.12.

Germination of Carolina willow among different soils (a) and seed source locations (b). Blue Cypress Marsh Conservation Area (BC); River Lakes Conservation Area (RL); St. Johns Marsh Conservation Area (SJ). Mixed soils were 1:1 ratios by volume of all possible combinations: BC/RL, BC/SJ and SJ/RL
Experiment 3: Temperature Regime, Seed Source, Collection Season, and Initiation of Watering
We did not find significant effects of production timing or temperature regime (Table 3 and Fig. 4). However, we did find a decrease in germination with delay in watering. We also found an interaction between watering delay and source of seeds with longer viability for those seeds from the southern region (Fig. 4c). The total proportion of seeds germinating was significantly greater when they originated from the southern region (0.52 vs. 0.46 respectively).

Mean and 95 % CI germination of Carolina willow as a function of collection time (a), temperature regime (b), seed source (c) and days until watering. Confidence intervals are asymmetrical because they were back-calculated from the angularly-transformed data used for hypothesis testing. Collection time was early, middle, or late in the flowering season at each location
Discussion
Exotic wetland shrubs usually become invasive because they are superior competitors than the native species they displace (e.g., Gordon 1998). For example, Tamarix spp. tolerate soil salinity better than native riparian plants, which gives tamarisks a competitive advantage and the potential to spread throughout the entire southwestern USA (Morisette et al. 2006) and arid central Australia (Griffin et al. 1989). In contrast, invasions of native wetland shrubs usually occur following anthropogenic disturbances of herbaceous wetlands altering the context of their interactions. Agriculture disturbance, altered tidal regimes and increasing nutrient levels and sedimentation have been hypothesized as causes of the replacement of saltmarsh by mangroves in south-east Australia (Saintilan and Williams 1999). Worldwide, expansion of willow species is associated with reduced water fluctuation, fire suppression, nutrient increases, and expansion of farming and urbanization (Zukowski and Gawne 2006; Ponzio et al. 2006; Quintana-Ascencio et al. 2013). Identifying cost-effective ways to halt and eventually reverse invasions of exotic or native shrubs into herbaceous wetlands is vital for preserving native biodiversity and ecological services, as well as economic functions and social values.
We conducted three experiments in growth chambers over 2 years using Carolina willow seeds and soils from several locations in the upper St. Johns River basin. The results consistently demonstrated that germination and early seedling survival of Carolina willow were strongly affected by the time delay between seed collection and watering, watering regime, and seed source. These environmental factors can be manipulated to reduce willow expansion into riparian zones that historically were herbaceous marshes, and to restore and maintain these marshes after mature willows are removed. This is a cost-effective means of controlling willows and mirrors the hydroperiod manipulations used to return arid streams dominated by invasive salt cedars back to the native communities of cottonwoods (Populus sp.) and black willow (S. nigra; e.g., Taylor and McDaniel 1998).
Only green seeds of Carolina willow were viable (L. Castro-Morales, pers. obs.) and successful germination was strongly influenced by water availability. More than 90 % germination occurred under saturated conditions, and germination success was greatest when seeds were watered immediately after collection. Carolina willow seeds started losing viability just 5 d after seeds were released. Upon release, willow seeds are covered by woolly hairs contributing to their dispersal by wind or buoyancy on the water surface (Douglas 1995; Liotta 2001; Karrenberg et al. 2002). Similar to those of Carolina willow, once seeds of other Salix spp. lose their woolly hairs and contact water or wet soil, germination occurs rapidly, usually within 24 h (Siegel and Brock 1990; Douglas 1995; Karrenberg et al. 2002; Seiwa et al. 2008). Although seeds of Carolina willow did not show dormancy in our experiments, germination was possible for several months for seeds held without moisture or light in a refrigerator, suggesting that viability may be longer than 10 d in more northern latitudes. However, most seeds we collected and held for more than a month under low temperature germinated, but none survived after a few days (L. Castro-Morales, pers. obs.). Monitoring willow flowering and limiting availability of moist, open substrates when seeds are dispersing (especially on organic soils) is one way to curtail colonization of Carolina willow into open herbaceous marshes. This is the reverse case to exploiting willow’s hydrological window to facilitate its establishment over other riparian zone species (e.g., Stromberg et al. 2007).
Soil moisture was fundamental for survival and establishment of Carolina willow. While some willow species (e.g., S. viminalis: Splunder et al. 1995) are drought tolerant, seeds and seedlings of Carolina willow and other species die after just 8 – 9 d without watering or precipitation (McLeod and McPherson 1973; Sacchi and Price 1992; Johnson et al. 2007; Quintana-Ascencio et al. 2013). In our experiments, most willow seeds sown onto dry, sandy soil and held without water for 8 d failed to survive, but seeds with access to water had high mean germination and survival rates. However, germination and seedling survival still depended on relatively subtle, centimeter-scale differences in water levels. Seeds in saturated soils kept moist by capillarity had the highest germination rates and consistently had the highest seedling survival. Willows in this treatment had access to unlimited water but did not experience anaerobic substrates. Flooded seeds had germination rates similar to seeds on saturated soils, but seedling survival was lower. Inundation conditions with higher water levels than those evaluated in our study (5 cm depth) may decrease survival further. Seedlings and saplings flooded to or above the soil surface show adverse responses ranging from diminished growth and photosynthesis to mortality (Jones et al. 2006; Quintana-Ascencio et al. 2013). Flooding also decreases seed viability by preventing the emergent radicle of floating seedlings from contacting the substrate (Titus 1991; Taiz and Zeiger 2002). Flooding, therefore, is a potential management strategy for reducing willow seedlings in an invaded wetland. We demonstrated its efficacy in two field experiments (Quintana-Ascencio et al. 2013), but we caution that once willows become small saplings (> 20 cm tall), they must be completely inundated for months to cause mortality (Quintana-Ascencio et al. 2013).
Seed source also explained significant variation in germination and seedling survival of Carolina willow. However, aside from longer viability, seed sources had unpredictable effects that varied from one experiment to the next. From a management perspective, and given Carolina willow’s short reproductive season, it will be more valuable to monitor seeding schedules, and to better understand spatial variation on seed amount, quality, and dispersal patterns. However, genetic considerations often are important in invasions of other wetland species, where specific genotypes or hybrids are invasive and others are not (e.g., Spartina sp. in Europe and the USA; Ainouche et al. 2009).
Despite finer sediments retaining more water and providing better conditions for willow germination (Karrenberg et al. 2002), and recruitment being limited by moisture and water availability (Keddy and Constabel 1986), germination and establishment of Carolina willow did not differ significantly among the regional soil types we tested. We used soils that included a range from two pure organic soils to a sandy, inorganic soil, plus all of their 1:1 mixtures. Whenever a soil held sufficient water, especially through capillarity, seeds of Carolina willow germinated and survived well. We found similar responses in greenhouse experiments with willow seedlings and small saplings; strong responses to water and weak or no responses to NH4 +, PO4 −, K+ and micronutrient supplements (Fauth et al., in prep.). This contrasts strongly with invasions of cattail (Typha domingensis Pers.) into sawgrass (Cladium mariscus (L.) Pohl ssp. jamaicense (Crantz) Kük) communities in the nearby Everglades, which are facilitated by enhanced nutrients that strongly favor cattail (Newman et al. 1996). Because organic soils that occur near river margins in the USJRB are usually kept moist by capillarity, it is unlikely that managers can create conditions dry enough to reduce willow germination. Inundation during Carolina willow’s reproductive season may be a more effective management strategy than dry-down to reduce its establishment by seed. Manipulating water levels and flows is an effective technique for controlling some invasive wetland plants (e.g., Typha: Urban et al. 1993; Tamarix: Gladwin and Roelle 1998; but not Melaleuca: Serbesoff-King 2003) and illustrates how context-dependent interactions can be manipulated to restore herbaceous wetland communities.
References
Ainouche ML, Fortune PM, Salmon A, Parisod C, Grandbastien MA, Fukunaga K, Ricco M, Misset MT (2009) Hybridization, polyploidy and invasion: lessons from Spartina (Poaceae). Biological Invasions 11:1159–1173
Barnea A, Yom-Tov Y, Friedman J (1991) Does ingestion by birds affect seed germination? Functional Ecology 5:394–402
Crawley MJ (2007) The R book. John Wiley & Sons, Chichester
Dahl TE (2000) Status and trends of wetlands in the conterminous United States 1986 to 1997. U.S. Department of the Interior, Fish and Wildlife Service, Washington, D.C., 82 pp
Douglas DA (1995) Seed germination, seedling demography, and growth of Salix setchelliana on glacial river gravel bars in Alaska. Canadian Journal of Botany 73:673–679
Gladwin DN, Roelle JE (1998) Survival of plains cottonwood (Populus deltoides sub. sp. monilifera) and saltcedar (Tamarix ramosissima) in response to flooding. Wetlands 18:669–674
Global Invasive Species Database (GISD) (2013) Salix. Available from: http://www.issg.org/database/species/search.asp?sts=sss&st=sss&fr=1&x=0&y=0&sn=salix&rn=&hci=-1&ei=-1&lang=EN Accessed 14 January 2013
Gordon DR (1998) Effects of invasive, non-indigenous plant species on ecosystem processes: lessons from Florida. Ecological Applications 8:975–989
Griffin GF, Smith DS, Morton SR, Allan GE, Masters KA, Preece N (1989) Status and implications of the invasion of tamarisk (Tamarix aphylla) on the Finke River, Northern Territory, Australia. Journal of Environmental Management 29:297–315
Hall GB (1987) Establishment of minimum surface water requirements for the greater Lake Washington Basin. Technical Publication SJ87–3, St. Johns River Water Management District, Palatka, FL. 85 pp
Hanselman JA, Bush M, Lee MA (2005) Seed content and percent organic matter in surface sediments as indicators of wetland plant communities, Blue Cypress Marsh, Florida. Florida Scientist 68:250–260
Jones DT, Sah JP, Ross MS, Oberbauer SF, Hwang B, Jayachandran K (2006) Responses of twelve tree species common in Everglades tree islands to simulated hydrologic regimes. Wetlands 26:830–844
Johnson KG, Allen MS, Havens KE (2007) A review of littoral vegetation, fisheries, and wildlife responses to hydrologic variation at Lake Okeechobee. Wetlands 27:110–126
Karrenberg S, Edwards PJ, Kollmann J (2002) The life history of Salicaceae living in the active zone of floodplains. Freshwater Biology 47:733–748
Keddy PA, Constabel P (1986) Germination of ten shoreline plants in relation to seed size, soil particle size and water level: an experimental study. Journal of Ecology 74:133–141
Kinser P, Lee MA, Dambek G, Williams M, Ponzio KJ, Adamus C (1997) Expansion of Carolina willow in the Blue Cypress Marsh Conservation Area, Upper St. Johns River Basin. Professional Paper SJ97–PP1, St. Johns River Water Management District, Palatka, Florida. 13 pp
Kuzovkina YA, Quigley MF (2005) Willows beyond wetlands: uses of Salix L. species for environmental projects. Water, Air, & Soil Pollution 162:183–204
Liotta J (2001) Rasgos biológicos de Salix humboldtiana Willd. y régimen de pulsos de inundación. Interciencia 26:397–403
Lowe EF, Brooks JE, Fall CJ, Gerry LR, Hall GB (1984) U.S. EPA Clean Lake Program, Phase I Diagnostic-Feasibility study of the Upper St. Johns River Chain of Lakes, Volume 1 – Diagnostic Study. Technical Publication SJ 84–15. St. Johns River Water Management District, Palatka, FL
Lowe S, Browne M, Boudjelas S, De Poorter M (2004) 100 of the world’s worst invasive alien species: a selection from the Global Invasive Species Database. Published by The Invasive Species Specialist Group (ISSG), a specialist group of the Species Survival Commission (SSC) of the World Conservation Union (IUCN), 12 pp
Martin TD, Brockhoff CA, Creed JT, EMMC Methods Work Group (1994) Determination of metals and trace elements in water and wastes by inductively coupled plasma-atomic spectrometry. Method 200.7, Revision 4.4. Environmental Monitoring Systems Laboratory, Office of Research and Development, U.S. Environmental Protection Agency, Cincinnati, Ohio
McLeod KW, McPherson JK (1973) Factors limiting the distribution of Salix nigra. Bulletin of the Torrey Botanical Club 100:102–110
Miller SJ, Ponzio KJ, Lee MA, Keenan LW, Miller SR (1998) The use of fire in wetland preservation/restoration: are there risks? Pp. 127–139 in T.L. Pruden and L.A. Brennen, eds., Fire in Ecosystem Management: Shifting the Paradigm from Suppression to Prescription. Proceedings of the Tall Timbers Fire Ecology Conference, No. 20, May 22–25, 1996, Boise, Idaho
Morisette JT, Jarnevich CS, Ullah A, Cai W, Pedelty JA, Gentle JE, Stohlgren TJ, Schnase JL (2006) A tamarisk habitat suitability map for the continental United States. Frontiers in Ecology and the Environment 4:11–17
Newman S, Grace JB, Koebel JW (1996) Effects of nutrients and hydroperiod on Typha, Cladium and Eleocharis: implications for Everglades restoration. Ecological Applications 6:774–783
Niiyama K (1990) The role of seed dispersal and seedling traits in colonization and coexistence of Salix species in a seasonally flooded habitat. Ecological Research 5:317–331
O’Dell J (1993a) (ed.) Determination of Ammonia Nitrogen by semi-automated colorimetry. Method 350.1. Environmental Monitoring Systems Laboratory, Office of Research and Development, U.S. Environmental Protection Agency, Cincinnati, Ohio
O’Dell J (1993b) (ed.) Determination of Nitrate-Nitrite Nitrogen by automated colorimetry. Method 353.2. Environmental Monitoring Systems Laboratory, Office of Research and Development, U.S. Environmental Protection Agency, Cincinnati, Ohio
O’Dell J (1993c) (ed.) Determination of Phosphorus by semi-automated colorimetry. Method 365.1 Environmental Monitoring Systems Laboratory, Office of Research and Development, U.S. Environmental Protection Agency, Cincinnati, Ohio
Ponzio KJ (1998) Effects of various treatments on the germination of Sawgrass, Cladium jamaicense Cranz, seeds. Wetlands 18:51–58
Ponzio KJ, Miller SJ, Underwood E, Rowe SP, Voltolina DJ, Miller TD (2006) Responses of a willow (Salix caroliniana Michx.) community to roller-chopping. Natural Areas Journal 26:53–60
Quintana-Ascencio PF, Fauth JE, Castro-Morales LM, Ponzio KJ, Hall D, Snyder K (2013) Taming the beast: managing hydrology to control Carolina Willow (Salix caroliniana) seedlings and cuttings. Restoration Ecology 21:639–647
Reyes MJ (2012) Use of adventitious roots for the determination of hydroperiod in isolated wetlands. Doctoral dissertation. University of South Florida, Tampa, Florida
Rodgers JA (1989) Willowy species. Aquatics 11:4–8
Sacchi CF, Price PW (1992) The relative roles of abiotic and biotic factors in seedling demography of arroyo willow (Salix lasiolepis: Salicaceae). American Journal of Botany 79:395–405
Saintilan N, Williams RJ (1999) Mangrove transgression into saltmarsh environments in south-east Australia. Global Ecology and Biogeography 8:117–124
Serbesoff-King K (2003) Melaleuca in Florida: a literature review on the taxonomy, distribution, biology, ecology, economic importance and control measures. Journal of Aquatic Plant Management 41:98–112
Seiwa K, Tozawa M, Ueno N, Kimura M, Yamasaki M, Maruyama K (2008) Roles of cottony hairs in directed seed dispersal in riparian willows. Plant Ecology 198:27–35
Sher AA, Marshall DL, Taylor JP (2002) Establishment patterns of native Populus and Salix in the presence of invasive nonnative Tamarix. Ecological Applications 12:760–772
Siegel RS, Brock JH (1990) Germination requirements of key southwestern woody riparian species. Desert Plants 10:3–8
Shipley B, Parent M (1991) Germination responses of 64 wetland species in relation to seed size, minimum time to reproduction and seedling relative growth rate. Functional Ecology 5:111–118
Southall EJ, Dale MP, Kent M (2003) Floristic variation and willow carr development within a southwest England wetland. Applied Vegetation Science 6:63–72
Splunder IV, Coops H, Voesenek LACJ, Blom CWPM (1995) Establishment of alluvial forest species in floodplains: the role of dispersal timing, germination characteristics and water level fluctuations. Acta Botanica Neerlandica 44:269–278
St. Johns River Water Management District (SJRWMD) (2007) Upper St. Johns River Basin, Surface Water Improvement and Management Plan. St. Johns River Water Management District, Palatka, 39 pp
Stromberg JC, Lite SJ, Marler R, Paradzick C, Shafroth PB, Shorrock D, White JM, White MS (2007) Altered stream-flow regimes and invasive plant species: the Tamarix case. Global Ecology and Biogeography 16:381–393
R Development Core Team (2011) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, http://www.R-project.org
Taiz L, Zeiger E (2002) Plant physiology, 3rd edn. Sinauer Associates, Sunderland
Tallent-Halsell NG, Walker LR (2002) Responses of Salix gooddingii and Tamarix ramosissima. Wetlands 22(2):776–785
Taylor JP, McDaniel KC (1998) Restoration of saltcedar (Tamarix sp.)-infested floodplains on the Bosque del Apache National Wildlife Refuge. Weed Technology 12:345–352
Titus JH (1991) Seed bank of a hardwood floodplain swamp in Florida. Castanea 56:117–127
Traveset A, Riera N, Mas RE (2001) Passage through bird guts causes interspecific differences in seed germination characteristics. Functional Ecology 15:669–675
United States Department of Agriculture. Soil Conservation Service (1974) Soil Survey of Brevard County, Florida. http://ufdc.ufl.edu/UF00026071/00001. Accessed November 27, 2013
United States Department of Agriculture. Soil Conservation Service (1989) Soil Survey of Orange County, Florida. http://ufdc.ufl.edu/UF00025733/00001/. Accessed November 27, 2013
United States Department of Agriculture. Soil Conservation Service (1990) Soil Survey of Seminole County, Florida. http://ufdc.ufl.edu/UF00025738/00001/. Accessed November 27, 2013
Urban NH, Davis SM, Aumen NG (1993) Fluctuations in sawgrass and cattail densities in Everglades Water Conservation Area 2A under varying nutrient, hydrologic and fire regimes. Aquatic Botany 46:203–223
Young JA, Clements CD (2003) Seed germination of willow species from a desert riparian ecosystem. Journal of Range Management 56:496–500
Zedler JB, Kercher (2004) Causes and consequences of invasive plants in wetlands: opportunities, opportunists, and outcomes. Critical Reviews in Plant Sciences 23: 431–452.
Zukowski S, Gawne B (2006) Potential Effects of Willow (Salix spp.) Removal on FreshwaterEcosystem Dynamics. A Literature Review. Report for the North East Catchment Management Authority. Murray-Darling Freshwater Research Centre, Wodonga
Acknowledgments
We thank the staff of the St. Johns River Water Management District for providing access to sites. We thank M. Ferrer, B. Stephens, Jennifer Navarra, and graduate students from the Restoration Ecology class: C. Brescacin, S. Haynes, M. Gordon, K. Grablow, L. Souto, J. Waddell, A. Weiler-Lazarz and L. Sánchez for help during the initial project stages and with soil and seed sampling. Lisa McCauley assisted with GIS and L. Pearman from the U. S. Geological Survey provided temperature data. This project was supported by the St. Johns River Water Management District.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Castro-Morales, L.M., Quintana-Ascencio, P.F., Fauth, J.E. et al. Environmental Factors Affecting Germination and Seedling Survival of Carolina Willow (Salix Caroliniana). Wetlands 34, 469–478 (2014). https://doi.org/10.1007/s13157-014-0513-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13157-014-0513-6