
Abstract
The prevalence of obesity has increased rapidly during recent years and has reached epidemic proportions. As a result, the scientific community is interested in active biomolecules which are naturally present in plants and foodstuffs and may be useful in body weight management. In recent years, polyphenols have made up one of the most frequently studied groups among these molecules. Numerous studies have been carried out on animals to analyse the potential anti-obesity effects of resveratrol, a non-flavonoid polyphenol, and a general consensus concerning the body-fat-lowering effect of this compound exists. By contrast, studies in humans have been few so far. Moreover, in these studies, the effectiveness of resveratrol is low. The aims of the present review are to summarize the results reported so far on this topic and to justify the differences observed between animals and humans. It seems that the reduced response to resveratrol in humans cannot be attributed to the use of lower doses in humans because the doses that induce body-fat-lowering effects in rodents are in the same range as those used in human studies. With regard to the experimental period length, treatments were longer in animal studies than in human studies. This can be one of the reasons contributing to the reduced responses observed in humans. Moreover, animals used in the reported studies are young while volunteers participating in human studies are adults, suggesting that resveratrol may be more efficient in young individuals. In addition to differences in the experimental designs, metabolic differences between animals and human cannot be discarded.
Similar content being viewed by others

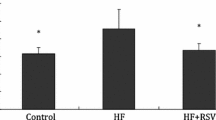
Avoid common mistakes on your manuscript.
Introduction
The prevalence of obesity has increased rapidly during recent years, reaching epidemic proportions. An important issue to be emphasized is that nowadays, obesity is not only an adult problem as children are also increasingly affected. According to WHO data, in 2014, approximately 1.9 billion adults, aged 18 and older, were overweight. Of these, over 600 million were obese. As far as children are concerned, 41 million children under the age of 5 were overweight or obese in 2014 [41]. Obesity is associated with increased risk of development of numerous adverse health conditions, such as type 2 diabetes, dyslipidemia, hypertension and other cardiovascular diseases and cancer, among others [18].
Energy restriction and physical activity are the commonly used tools in overweight and obesity treatment. However, due to social and economic reasons among others, compliance with this treatment protocol is often very poor, especially in the long term. Furthermore, the number of drugs approved for obesity treatment remains limited as a consequence of their side effects, and as a result, some have been withdrawn from the market in recent years.
In this context, the scientific community is interested in active biomolecules which are naturally present in plants and foodstuffs and which may be useful in body weight management. In fact, plants have been an important source of drugs; many currently available drugs have been derived directly or indirectly from them. Moreover, natural products have been used as traditional medicines around the world to treat various diseases for thousands of years. Among these molecules, polyphenols, a wide and heterogeneous family of phytochemicals present in different plant species, make up one of the most frequently studied groups in recent years.
One of the more widely studied polyphenols in the field of obesity is resveratrol (3,5,4′-trihydroxy-trans-stilbene), a non-flavonoid polyphenol that shows hydrogen atoms available for transfer to reactive species and interrupt oxidative cascades (Fig. 1). It is produced naturally in several plants in response to injury or fungal attack [25]. It was first detected in the roots of white hellebore (Veratrum grandiflorum) and is found in various foodstuffs such as grapes, berries, red wine and nuts [10]. Although the molecule exists in two isoforms, trans-resveratrol and cis-resveratrol (Fig. 1), the trans form, which is the preferred steric form in nature, is relatively stable [8]. The glucose-bound form of resveratrol is piceid, predominant in food sources and converted to trans-resveratrol by hydrolysis (Fig. 1) [9].

Structure of piceid and trans-resveratrol
A general consensus concerning the body-fat-lowering effect of resveratrol in mice and rats exists [1, 37]. The present review focuses on the studies devoted to analysing the potential anti-obesity effects of resveratrol in animal models and human beings (for a review of the usefulness of many other phytochemicals for the prevention and treatment of obesity, please refer to [20]). As this review focuses its interest on resveratrol anti-obesity properties, many of the studies with other aims have not been included in the present review although they include data concerning body weight or body fat following resveratrol treatment.
Studies in animals
Many studies have been addressed using animal models and various experimental approaches to analyse the potential anti-obesity effect of resveratrol. The most commonly used species are mice (Table 1) and rats (Table 2).
Studies in mice
As far as mice are concerned, the pioneering studies on the anti-obesity effect of resveratrol were performed by Lagouge et al. in 2006 [23]. They fed C57BL/6J mice with a standard or a high-fat diet, with or without resveratrol at 0.4 g/100 g diet (0.4% w/w). This resveratrol supplementation resulted in an ingested daily dose varying from 200 to 400 mg/kg body weight/day depending on the evolution of body weight and food intake of the 4 to 8 week-old male mice fed these diets for 9 to 15 weeks. The effect of resveratrol in reducing weight gain was significant only under high-fat feeding conditions. The reduction in body mass gain was accounted for by a decrease in body fat. An unequivocal anti-obesity effect was attested by the lower weight of adipose depots. A histomorphological analysis of epididymal white adipose tissue showed smaller adipocytes upon resveratrol treatment. These beneficial effects were not due to decreased food intake. By contrast, basal energy expenditure was significantly increased in high-fat-fed mice treated with resveratrol. Resveratrol supplementation also enhanced the capacity for adaptive thermogenesis, since treated mice maintained their body at a higher temperature than that of non-treated when submitted to short-term cold exposure. Morphometric analysis of brown adipose tissue revealed larger mitochondrial structures in resveratrol-treated mice. Accordingly, resveratrol increased uncoupling protein 1 (UCP1) expression, heat production capacity and gene expression of a masterpiece of the thermogenic program: peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α), which is a potent inducer of mitochondrial biogenesis. Moreover, the ratio of acetylated to total nuclear PGC-1α protein was reduced in BAT and muscle, meaning that PGC-1α was activated, by a pathway dependent on the stimulation of the protein deacetylase sirtuin-1 (SIRT1).
Around the same date, another publication revealed a similar anti-obesity effect when the addition of 0.04% resveratrol to a high-fat diet (60% of calories from fat) was reported to limit the body weight of male C57BL/6NIA mice during the initial months of treatment [7]. However, the protocol of resveratrol treatment was different: the dietary dose was tenfold lower, and the mice were 1 year old at the beginning of supplementation. This resulted in an average resveratrol intake of 22.4 mg/kg body weight/day. This low dose still increased mitochondrial number, PGC-1α and AMP-activated protein kinase (AMPK) activities. However, at 18–24 months of age, when most of the analyses were performed, there was no pronounced difference in subcutaneous, abdominal and epididymal white adipose tissue or brown adipose tissue between the control and resveratrol-treated groups. The other effects of resveratrol consisted in increasing insulin sensitivity, locomotor function and survival and were therefore beneficial for limiting obesity- and ageing-related disorders.
Two other studies confirmed the capacity of resveratrol to alleviate the obesogenic properties and the harmful outcomes of high-fat feeding. One consisted in a supplementation of a high-fat diet with resveratrol at 0.4% during 10 weeks [21]. Those mice (C57BL/6J) supplemented with resveratrol showed reduced body weight gain and visceral fat pad weights when compared to mice fed the high-fat diet, although food intake was not affected. Histological analysis of epididymal adipose tissue also showed smaller adipocytes in resveratrol-treated mice. No differences were found between standard-fat-fed mice and resveratrol-treated mice in the values for these parameters. Significant reductions in triglyceride and cholesterol plasma levels were also observed.
In order to explain this anti-obesity effect, the authors analysed the expression of several genes in adipose tissue. They observed that resveratrol significantly reduced the expression of CCAAT-enhancer-binding protein α (C/EBPα), peroxisome proliferator-activated receptor γ 2 (PPARγ2), sterol regulatory element binding protein 1c (SREBP-1c) and liver X receptor (LXR), as well as their target genes fatty acid synthase (FAS), lipoprotein lipase (LPL), adipocyte protein 2 (aP2) and leptin. In addition, resveratrol-treated mice exhibited a reduced expression of adipogenic signalling molecules, such as galanin receptors 1 (GalR1), galanin receptors 2 (GalR2), protein kinase C delta (PKCσ), cyclin D (CyC-D) and E2F transcription factor 1 (E2F1), together with a limited phosphorylation of the extracellular signal-regulated kinases (p-ERK). Indeed, the level of gene expression was similar in mice fed the standard fat diet and the mice fed the high-fat diet treated with resveratrol. This study confirmed that resveratrol promoted a normalization of the disturbances induced by high-fat feeding and suggested that resveratrol-induced adipose tissue mass reduction was due, at least in part, to reduced adipogenesis and lipogenesis.
More confirmation was added by a study designed to compare the effects of two doses of resveratrol, using four experimental groups of male C57BL/6J mice: a group fed a standard diet, another fed a high-fat diet reaching 40% of calories as fat and two additional high-fat-diet-fed groups supplemented with 0.005 or 0.02% resveratrol for 10 weeks [13]. These treatments, which assumed that a resveratrol intake of 5 and 20 mg/kg body weight/day would be reached, reduced the weight of perirenal and mesenteric adipose tissue. However, only the lower dose reduced the weight of the epididymal and retroperitoneal depots. Morphological observations also indicated that epididymal adipocyte size was smaller in the resveratrol-supplemented mice. These reductions in fat deposition were not caused by a reduction in food intake. Resveratrol significantly decreased lipogenic enzyme activities, such as FAS and phosphatidate phosphohydrolase (PAP), in epididymal adipose tissue. Furthermore, mice fed the low dose of resveratrol showed a significant decrease in glucose-6-phosphate dehydrogenase (G6PDH) activity and a significant increase in fatty acid beta-oxidation. Taken as a whole, these results indicated that the body-fat-lowering effect of resveratrol was mediated by a reduction in lipogenesis, and in accordance with that observed for white adipose tissue weight, the effects of low resveratrol dose were greater than those of the high dose. Furthermore, Cho et al. observed that the low dose normalized the increase in plasma total cholesterol and apoB/apoA-I ratio induced by the high-fat diet, while these effects were not seen with the highest dose.
Along the same lines as the study reported by Cho et al., successive studies also revealed that the anti-obesity properties of resveratrol did not follow a classical dose-response curve. Very recently, the absence of a classical sigmoidal dose-response pattern for resveratrol in limiting body weight gain was reported in a study comparing the effects of the administration by oral gavage of resveratrol at 5, 22.5 and 45 mg/kg body weight/day for 12 weeks in high-fat-fed mice. All the treated groups exhibited a significant reduction in body weight compared with the high-fat control, but there was no difference between the three resveratrol-treated groups [16]. In another study, the anti-obesity effects of resveratrol were tested at 1, 10 and 30 mg/kg body weight/day for 10 weeks in high-fat-fed mice (45% of calories from fat) and the results showed that, without decreasing energy intake, resveratrol attenuated the high-fat diet-induced weight gain and protected against lipid deposition in white adipose tissue and liver. Although in some parameters, there was a dose-response pattern, for many parameters, the effect of a daily intake of 10 mg/kg was equivalent to or even tended to be greater than that of 30 mg/kg [12].
There are also studies reported in the literature that did not find the previously described resveratrol-induced positive effects. In the study published by Voigt et al. [40], high-fat-fed C57BL/6J male mice were treated for only 21 days with oral resveratrol (30 mg/kg/day). This treatment tended to reduce body weight gain and adipose tissue weight without reaching significant differences when compared to control. In this study, it may not have been the resveratrol dosage that caused the lack of effect, but rather the short treatment duration. Even a treatment duration of 4 weeks appeared insufficient to detect any anti-obesity effects of resveratrol. In a study aiming to compare the anti-obesity effect of resveratrol to that of its dimer ε-viniferin, C57BL/6J mice were fed a very high-fat diet (60% of total calories as lipids) with or without the respective stilbenes for 4 weeks. Supplementations with 0.2 and 0.4% resveratrol were not associated with significant reductions of body weight gain or adipose tissue weight when compared to control, though this took place in the case of ε-viniferin-containing supplementation [31].
Tauriainen et al. carried out a study in mice fed a high-fat diet (60% of energy as fat) supplemented with resveratrol at 0.2 and 0.4% [38]. While these treatments did not clearly decrease body or fat pad weights or modify body fat content, they partially protected against high-fat diet-induced fatty liver formation. In this case, there is no clear justification for the lack of effects of resveratrol administration. The ingested amounts of the polyphenol represented 135 and 282 mg/kg body weight/day, doses that are included in the wide range of doses tested by other research teams (5 to 400 mg/kg body weight/day). As far as the experiment period length is concerned, it was longer (15 weeks) than most of the “positive” studies that were performed with supplementation periods lasting between 8 and 11 weeks.
In addition to mice fed a high-fat diet, there were other studies performed under different feeding conditions. Thus, Andrade et al. [5] treated mice fed a standard diet with 30 mg/kg body weight/day of resveratrol. Under these conditions, the delipidating effect of resveratrol was also observed. After 2 months of treatment, these animals presented significantly decreased fat accumulation in adipose tissue, with diminished total cholesterol plasma levels when compared with the controls. Additionally, increased oxygen consumption was observed. Analyses of messenger RNA (mRNA) of thermogenesis-related genes showed significant increase in UCP1, SIRT1 and bone morphogenetic protein-7 (BMP-7) expression in brown adipose tissue, suggesting that increased thermogenesis could be involved in the reduction in body fat accumulation induced by the polyphenol.
Mendes et al. [27] fed FVB/N mice a high-fat diet or a high-protein diet with or without resveratrol (4 g/kg diet). Mice fed the high-protein diet supplemented with resveratrol showed decreased body weight, body adiposity, adipose tissue weight, adipocyte area and total cholesterol, as well as acetyl-CoA carboxylase (ACX) and FAS gene expressions. Again, such body-fat-lowering effect of resveratrol was mediated in part by a reduction in de novo lipogenesis. As far as mice fed the high-fat diet supplemented with resveratrol were concerned, only decreased concentration of total cholesterol was observed when compared with mice fed the high-fat diet not supplemented with the polyphenol.
Likewise, Jeon et al. used homozygous apoE-deficient mice, a model of hypercholesterolemia that mimics the initiation and progression of human atherosclerosis [19]. After 12 weeks of atherogenic diet supplemented with 0.02% resveratrol, a significant reduction in body weight and epididymal white adipose tissue weight was observed, but not that of perirenal fat depot. No changes were observed in food intake. Resveratrol-treated mice exhibited increased plasma levels of HDL-cholesterol and lower apoB/apoA-I ratio. However, no change was detected in the activity of FAS, G6PDH or malic enzyme (ME) in epididymal white adipose tissue between control and treated animals. Nevertheless, resveratrol supplementation resulted in significantly elevated beta-oxidation and lowered activity of the lipogenic enzyme PAP in white adipose tissue.
Lastly, it may be noted that Kunming mice, widely studied in China, also exhibited lower body and visceral white adipose tissue weights when they were fed a high-fat diet (50% energy as fat) supplemented with resveratrol at a dose of 200 mg/kg body weight/day for 12 weeks [33]. The polyphenol improved the gut microbiota dysbiosis induced by the high-fat diet, by increasing the Bacteroidetes-to-Firmicutes ratios, inhibiting the growth of Enterococcus faecalis and increasing the growth of Lactobacillus and Bifidobacterium. Pearson’s correlation coefficient showed that there was a negative correlation between body weight and the ratios of Bacteroidetes to Firmicutes, as well as body weight and Lactobacillus and Bifidobacterium. By contrast, a positive correlation was found between body weight and E. faecalis. In addition, resveratrol significantly decreased mRNA expression of genes related to fatty acid synthesis, adipogenesis and lipogenesis (LPL, ACX, FAS, stearoyl-coenzyme A desaturase 1 (SCD-1), PPARγ) in epididymal adipose tissue. Thus, in this study, resveratrol seemed to induce body fat reduction through the inhibition of metabolic pathways by driving fat accumulation and also by modifying gut microbiota.
Taken as a whole, almost all the experiments performed on mouse models converge in demonstrating that oral administration of resveratrol ranging from 5 to 400 mg/kg body weight/day exerts anti-obesity actions that are especially detectable after 8 to 12 weeks of supplementation, when using simple morphologic indexes such as body mass and adiposity. Although the administration of the higher tested doses of resveratrol is hardly transposable to humans, the different studies have shown that resveratrol use is safe and that diverse complications of obesity can be alleviated owing to its supplementation, via (though not exclusively) SIRT1 and AMPK activation.
Studies in rats
Other studies have been performed in rats. In general terms, although mice are highly responsive to different treatments, resveratrol treatment also shows interesting effects on rats. In a study from our group [26], we analysed the effects of three doses of resveratrol (6, 30 and 60 mg/kg/day), administered for 6 weeks to male rats fed a high-fat high-sucrose diet (40% of energy as fat). Although a clear dose-response was not observed, important differences were found among the three experimental doses. While the lowest dose (6 mg/kg/day) did not show a body-fat-reducing effect, the intermediate dose (30 mg/kg/day) reduced all the adipose depots measured (subcutaneous, epididymal, perirenal and mesenteric). The highest dose (60 mg/kg/day) led to a significant reduction in mesenteric and subcutaneous depots but not in epididymal and perirenal tissues. No significant changes were observed in food intake. In good accordance with the results found in other studies performed in mice [13], the lower dose of resveratrol within the range of effective doses (30–60 mg/kg/day) was more effective than the higher one in this study. Serum parameters, cholesterol, triacylglycerols, free fatty acids and glucose, were not affected by resveratrol intake.
In other study designed to assess the effects of a combination of resveratrol and quercetin using a similar experimental design in terms of diet and experimental period length (high-fat high-sucrose diet, 6 weeks of treatment), we observed that resveratrol, at a dose of 15 mg/kg/day, was unable to reduce body weight gain or adipose tissue weight in rats [6]. These results showed that, under the experimental conditions used in these two studies, resveratrol was ineffective in the range of 6 to 15 mg/kg/day.
When we analysed the potential mechanisms of action underlying the body-fat-lowering effect of resveratrol in the cohort of animals treated with 30 mg/kg/day of this polyphenol [3], we observed a significant reduction in the activity of heparin-releasable LPL, the enzyme which allows adipose tissue to uptake fatty acids from triacylglycerol transported by chylomicrons and very low-density lipoproteins, as well in the activity of lipogenic enzymes, such as ACX, FAS and G6PDH in epididymal adipose tissue. These data suggest that the effect of resveratrol on adipose tissue was mediated, at least in part, by a reduction in fatty acid uptake from triacylglycerols in circulating lipoproteins as well as in de novo lipogenesis
As far as the lipolytic pathway is concerned, no changes were observed in gene expression of adipose tissue triglyceride lipase. By contrast, gene expression of hormone-sensitive lipase was significantly reduced by the polyphenol treatment. Although it is well known that the main mechanism by which lipases are activated is phosphorylation, the increase in hormone-sensitive lipase expression suggests a potential increase in lipolysis, which is in good accordance with the enhanced lipolytic response to epinephrine induced by resveratrol observed by Szkudelska et al. [36].
In this cohort of rats, we also observed a significant increase in SIRT-1 and PGC-1α gene expressions, in interscapular brown adipose tissue from rats treated with resveratrol [4]. PGC-1α is a potent inducer of mitochondrial biogenesis, an important part of the thermogenic program. Increased PGC-1α expression was accompanied by a great increase in mitochondrial transcription factor A (TFAM) gene expression, suggesting that resveratrol enhanced mitochondriogenesis. This fact was confirmed by the increase in the mRNA of mitochondrial protein cytochrome C oxidase subunit-2 (COXII), a mitochondrion-encoded protein which is a critical component of the oxidative phosphorylation pathway.
We also analysed the protein expression of UCP1, the first uncoupling protein discovered, which is exclusively expressed in brown adipose tissue [4]. This effect was likely to be related to the increase in PGC-1α. These results are in line with those reported by Lagouge et al. [23] in mice brown adipose tissue using a very high dose of resveratrol (400 mg/kg/day) and a longer experimental period than that used in the present study (15 weeks).
Moreover, rats treated with resveratrol showed significantly greater protein expression of UCP3 and gene expression of TFAM and COXII in skeletal muscle [4]. These results show that resveratrol increased UCP protein expression in two important thermogenic tissues. This could contribute to increased whole-body energy dissipation and consequently to increased energy expenditure, thus reducing energetic efficiency, and it could represent another mechanism of action underlying the body-fat-lowering effects of resveratrol.
In order to gain more insight into the mechanisms underlying the anti-obesity effect of resveratrol in rats, we also analysed the effects of resveratrol on the clock genes. The circadian timing system is composed of several endogenous clocks and is mainly present in the suprachiasmatic nucleus of the hypothalamus, but circadian clocks are also present in peripheral tissues as the adipose tissue. Adipose tissue clock gene desynchronization has been related to the development of certain diseases such as obesity. In this context, it has been proposed that unhealthy lifestyle choices, such as high-fat feeding, produce progressive derangements in temporal communication among different food intake signals by modifying the strength, duration and frequency of circadian rhythms.
We observed that resveratrol treatment led to the downregulation of reverse erythroblastosis virus α (Rev-Erbα). This gene, which is a part of the clock machinery, participates in the regulation of the expression of SREBP-1c, the transcription factor which in turn regulates de novo lipogenesis, as well as in its translocation to the nucleus [24], that is in its maturation and activation. According to these data, it can be proposed that the reduction in lipogenic activities induced by resveratrol could take place via Rev-Erbα [28].
In addition to the model of rats fed an obesogenic diet, we also used a model of genetic obesity in our studies. We administered resveratrol at a dose of 15 mg/kg/day to obese Zucker fa/fa rats for 6 weeks with a standard diet [17]. In this animal model, resveratrol reduced both subcutaneous and internal adipose tissue weights, but the reduction only reached statistical significance in internal depots, mostly in epididymal adipose tissue. As previously mentioned in this review, when we analysed the effects of this dose of resveratrol also for 6 weeks in rats fed an obesogenic diet, the polyphenol did not induce reductions in adipose tissues. Thus, it seems that an obese Zucker rat is a more sensitive model.
When the activity of enzymes involved in triacylglycerol metabolism was assessed in epididymal adipose tissue, we observed that, in general terms, the activity of enzymes involved in de novo lipogenesis was not modified by resveratrol treatment. By contrast, the activity of LPL was significantly reduced. Finally, gene expression of hormone-sensitive lipase was increased, and gene expression of fatty acid translocase/scavenger receptor cluster of differentiation 36 (cd36), which is a transporter of fatty acids, was decreased. These results suggest that resveratrol reduces the uptake of fatty acids for triacylglycerol formation and increases the capacity for triacylglycerol mobilization.
Previous to our study, Rivera et al. [35] also analysed the effects of resveratrol in lean and obese Zucker rats. They fed rats with a standard diet and treated them with 10 mg/kg/day of polyphenol (orally provided by gavage) for 8 weeks. Resveratrol decreased abdominal fat content in obese rats but not in their lean littermates. This effect was not due to a reduction in food intake. These authors suggested that the body-fat-lowering effect induced by resveratrol in obese rats was mediated, at least in part, by the reduction in de novo lipogenesis because ACX was phosphorylated, that is inhibited, in adipose tissue. ACX phosphorylation was due to the activation of AMPK.
Nagao et al. [30] carried out a study in Otsuka Long-Evans Tokushima fatty (OLETF) rats. This animal model develops a syndrome with multiple metabolic and hormonal disorders that shares many features with human obesity. OLETF rats have hyperphagia due to a lack of receptors for cholecystokinin, and they become obese even by consuming a normal diet. They fed animals with a standard diet supplemented with 0.5% of resveratrol, which, according to the food intake and body weight values provided, represented approximately 200 mg/kg/day. After 4 weeks of treatment, they observed that resveratrol reduced omental and perirenal adipose tissue weights. In order to examine the effect of resveratrol on the oxidation of nutrients, respiratory gas analysis was performed. Carbohydrate oxidation and protein oxidation were significantly lowered in the resveratrol-treated rats after the first week of feeding. Moreover, resveratrol enhanced fat oxidation and suppressed carbohydrate and protein oxidation after 3 weeks of feeding. These results suggest that resveratrol induced an anti-obesity effect through the alteration of metabolic profiles of nutrients. Furthermore, when they analysed gene expression of FAS and PPARs and transcription factors involved in the regulation of lipolysis and adipocyte differentiation, no changes were observed between experimental groups. By contrast, mRNA level of carnitine palmitoyl transferase, a key enzyme in fatty acid β-oxidation, was significantly increased in the perirenal white adipose tissue from rats treated with the polyphenol. The authors proposed that this effect might contribute to the partial enhancement of fat oxidation induced by resveratrol.
All the studies described up to this point in this review show the effects of resveratrol during periods of fat accumulation due to dietary patterns or to genetic alterations. In order to broaden the focus, we carried out a study devoted to determining whether resveratrol was able to increase body fat reduction induced by energy restriction [2]. For this purpose, male Wistar rats were fed a high-fat high-sucrose diet (40% energy as fat) for 6 weeks to obtain a diet-induced obesity model. Then, they were submitted to a mild energy restriction (25%) without or with resveratrol supplementation (30 mg/kg body weight/day) for 2 weeks. Final body weight and white adipose tissue weights were reduced in both restricted groups, but no differences were found among them. FAS, ACX and LPL activities and expressions were also similar in both groups. These results suggested a lack of any adjuvant effect of resveratrol on energy restriction for obesity treatment purposes. The lack of effect of resveratrol supplementation observed in the work mentioned could be due to two main reasons. On the one hand, the effects caused by energy restriction were strong enough to mask the potential positive effects ascribed to resveratrol, which has been clearly observed under other feeding conditions. On the other hand, our experimental procedure might not have been long enough to allow resveratrol to bring about its effects.
Studies in other animal models
In addition to rodents, a study has been reported in mouse lemurs (Microcebus murinus, Cheirogaleidae), a seasonal non-primate model [15]. As in rodents, resveratrol was also effective in this model.
Behavioural and physiological seasonal changes in mouse lemurs are dependent on the photoperiod and are reproduced in captivity using an artificial photoperiodic regimen. In the present study, animals were studied during the weeks following the shift from long to short day lengths, corresponding to the pre-wintering fattening period. The animals were fed ad libitum during 4 weeks with fresh fruit (bananas and apples) and a mixture of cereals, milk and eggs supplemented with the amount of resveratrol needed to provide a dose of 200 mg/kg/day. Nevertheless, due to the fact that the animals tended to eat less food than provided, the real doses during the four treatment weeks ranged from 192 to 168 mg/kg/day.
Resveratrol reduced body mass gain, activated energy expenditure by inducing an increase in resting metabolic rate and a decrease in torpor patterns that play key roles in energy saving in these lemurs and decreased spontaneous food intake. At a molecular level, resveratrol activated SIRT1 and lipid metabolism and increased glucose-dependent insulinotropic polypeptide (GIP), which induces an increase in energy expenditure, motor activity and mobilization of fat stores. All of these changes participated in the limitation of pre-wintering fattening processes.
Studies in human beings
Only a reduced number of studies addressing the potential anti-obesity effect of resveratrol in humans have been reported so far (Table 3). In the near future, more data will be available because at present, there are several ongoing studies. Table 4 summarizes only ongoing studies designed to analyse the effects of resveratrol alone (not included in a polyphenol mixture) including parameters related to body fat or energy balance as outcomes (Table 4).
Yoshino et al. [42] analysed the metabolic effects induced by resveratrol supplementation in Caucasian postmenopausal women showing normal weight or overweight. Resveratrol was administered for 12 weeks at a dose of 75 mg/day. At the end of the total experimental period, no changes were observed in body weight, total fat mass, subcutaneous abdominal fat and intra-abdominal fat between both groups.
Taking into account that in animal models, the beneficial effects of resveratrol seem to be mediated via SIRT1, the authors analysed the gene expression of these mediators, as well as that of PGC-1α in white adipose tissue, but no differences were observed between placebo group and resveratrol-treated group.
Thus, the final conclusion of this study was that, under these experimental conditions, resveratrol supplementation was not effective. The lack of positive effects may be due to the fact that the subjects in this study were not obese. As already stated, some subjects were of normal weight and others slightly overweight. The vast majority of animal studies showing positive effects of resveratrol were performed in obese animals or animals fed an obesogenic diet (Tables 1 and 2).
Timmers et al. [39] analysed the effects of resveratrol administration on whole-body energy expenditure, substrate utilization and lipolysis in adipose tissue among other aspects. For this purpose, healthy obese males were randomly distributed in a placebo group or a resveratrol group treated with 150 mg resveratrol/day for 4 weeks. As far as energy expenditure was concerned, a lower sleeping metabolic rate was observed in the resveratrol-treated group and no differences were found in diet-induced thermogenesis or in a physical activity index. Finally, a higher respiratory quotient was noticeable in the resveratrol-treated group. The authors suggested that resveratrol administration was efficient in inducing an improvement in metabolic flexibility. Nevertheless, these effects were not accompanied by changes in body weight. Unfortunately, in this study, the authors did not provide data concerning adipose tissue or body fat. They later on addressed a new study with the same cohort for analysing abdominal and subcutaneous adipocyte size [22]. They observed that resveratrol supplementation induced a shift toward an increased proportion of small adipocytes. Moreover, a microarray was carried out, which showed increased expression of genes involved in adipogenesis. Although the authors proposed that these effects may underline the previously reported improved insulin sensitivity [39], it is not possible to draw conclusions concerning the net effect on adiposity.
In another study, Poulsen et al. [32] carried out a double-blinded, placebo-controlled parallel-group trial with healthy obese subjects in order to analyse the effect of resveratrol on energy and substrate metabolism as well as on metabolic biomarkers of obesity. In this case, the experimental period length was shorter and the polyphenol dose was greater. The treatment consisted of three tablets of resveratrol daily (500 mg/day) for 4 weeks. Participants were asked not to alter their normal way of life. Resveratrol did not induce significant changes in body mass, lean body mass and total body fat mass. Similarly, resting energy expenditure and lipid oxidation rate remained unchanged even though a higher dose was used. Previously, in this review, when the study reported by Yoshino et al. [42] was mentioned, we proposed that the lack of effect of resveratrol on body fat may have been due to participants not being obese. However, in the study reported by Poulsen et al., where subjects showed BMI ≥30 kg/m2, resveratrol was also ineffective, even at a higher dose. In this case, it is important to emphasize that the treatment period was quite short (4 weeks).
Crandall et al. [14] also analysed the effects of a short period of resveratrol administration on overweight to obese old subjects with impaired glucose tolerance. They administered resveratrol at doses of 1, 1.5 and 2 g/day for 4 weeks. Participants were instructed not to alter their usual dietary and physical activity patterns. In line with the results obtained by Poulsen et al. [32], regardless of the dose used, no changes were observed in body weight or percentage of body fat. Nevertheless, the authors indicated that the small number of subjects participating in their study reduced the ability to detect differences in some study outcomes.
Obese or overweight subjects were also used in the study reported by Chachay et al. [11]. Men whose body mass index was greater than 25 kg/m2 and with a waist circumference >90 cm, with non-alcoholic fatty liver disease, received placebo or 3 g of resveratrol during 8 weeks. No changes were observed in body weight, total abdominal fat or fat distribution. These results are in good accordance with those reported by Poulsen et al. [32] although the treatment period was longer, 8 weeks instead of 4 weeks, and the dose of resveratrol used was higher.
Mendez-del Villar et al. [29] carried out a study devoted to examining the effect of resveratrol administration on metabolic syndrome, insulin sensitivity and insulin secretion. In addition, they measured parameters related to obesity such as total body weight, body mass index and waist circumference. Subjects with a diagnosis of metabolic syndrome were treated with placebo or 500 mg of resveratrol three times per day before meals for 12 weeks. In this case, all the anthropometric parameters measured were significantly reduced, indicating that polyphenol treatment induced a decrease in body fat.
This is the only study showing a clear reduction in body fat induced by resveratrol. Two important aspects to point out are that this experimental period was longer than those used in other studies reporting no effects of this polyphenol, and that participants were obese. Thus, it can be hypothesised that treatment periods of at least 12 weeks are needed for resveratrol to exhibit body fat-lowering effects in obese humans.
Concluding remarks
Although the number of studies carried out in humans is low so far, the reported data show that animals seem to respond more readily to resveratrol treatment than humans. In order to shed light onto this different pattern of response, comparisons between experimental designs should be made. The doses used in the reported studies range from 5 to 400 mg/kg/day in mice, from 6 to 60 mg/kg/day in rats and from 75 to 3000 mg/day in humans. By using the Reagan-Shaw formula to know the equivalence between these doses and those used in human studies [34], it can be deduced that the doses used in human studies are in the same range as those used in animal studies (equivalent to 30–2.220 mg/day). Consequently, the reduced response to resveratrol in humans cannot be attributed to the use of lower doses in humans than in animals.
With regard to the experimental period length, in the reported studies, animals were treated for between 4 weeks and 6 months. In humans, the studies were performed for 4 to 12 weeks. Taking into account that lifespan is clearly shorter in rodents than in humans, in general terms, it can be stated that, in animal studies, treatments were longer than in human studies. This can be one of the reasons contributing to the reduced responses observed in humans.
Another important aspect to be considered is the age of individuals. Thus, while animals used in the reported studies were young, with the exception of the study reported by Baur et al. [7], volunteers participating in human studies are adults. This suggests that, perhaps, resveratrol is more efficient in young individuals. Furthermore, it is important to emphasize that the vast majority of the reported animal studies were performed by administering resveratrol during the fattening period. Thus, in some experimental designs, the polyphenol was administered at the same time as an obesogenic diet. In other cases, resveratrol was provided to young genetically obese rodents during the fattening period. By contrast, in humans, resveratrol was administered to overweight or obese subjects who were not apparently in a fattening period. According to the experimental designs of animal studies, it seems that in animal studies, resveratrol avoided or reduced fattening, thus showing a preventive effect on obesity. By contrast, resveratrol was administered in humans not to prevent but to treat obesity.
In addition to differences in experimental designs, metabolic differences between animals and human cannot be discarded. Thus, de novo lipogenesis, which is a metabolic pathway inhibited by resveratrol, is very relevant in adipose tissue from rodents but not in human adipose depots.
Finally, it should be emphasized that further studies in humans are needed in order to better characterize the potential anti-obesity effects of resveratrol. In this scenario, various population groups (gender, age, obesity degree, etc.) should be used.
Change history
05 October 2017
Volume 73 issue 3 was published with an incorrect cover date. Correct is August 2017. The Publisher apologizes for this mistake and all related inconveniences caused by this.
References
Aguirre L, Fernández-Quintela A, Arias N, Portillo MP (2014) Resveratrol: anti-obesity mechanisms of action. Molecules 19:18632–18655
Alberdi G, Macarulla MT, Portillo MP, Rodríguez VM (2014) Resveratrol does not increase body fat loss induced by energy restriction. J Physiol Biochem 70:639–646
Alberdi G, Rodríguez VM, Miranda J, Macarulla MT, Arias N, Andrés-Lacueva C, Portillo MP (2011) Changes in white adipose tissue metabolism induced by resveratrol in rats. Nutr Metab (Lond) 8:29
Alberdi G, Rodríguez VM, Miranda J, Macarulla MT, Churruca I, Portillo MP (2013) Thermogenesis is involved in the body-fat lowering effects of resveratrol in rats. Food Chem 141:1530–1535
Andrade JM, Paraíso AF, de Oliveira MV, Martins AM, Neto JF, Guimarães AL, de Paula AM, Qureshi M, Santos SH (2014) Resveratrol attenuates hepatic steatosis in high-fat fed mice by decreasing lipogenesis and inflammation. Nutrition 30:915–919
Arias N, Macarulla MT, Aguirre L, Milton I, Portillo MP (2016) The combination of resveratrol and quercetin enhances the individual effects of these molecules on triacylglycerol metabolism in white adipose tissue. Eur J Nutr 55:341–348
Baur JA, Pearson KJ, Price NL, Jamieson HA, Lerin C, Kalra A, Prabhu VV, Allard JS, Lopez-Lluch G, Lewis K, Pistell PJ, Poosala S, Becker KG, Boss O, Gwinn D, Wang M, Ramaswamy S, Fishbein KW, Spencer RG, Lakatta EG, Le Couteur D, Shaw RJ, Navas P, Puigserver P, Ingram DK, de Cabo R, Sinclair DA (2006) Resveratrol improves health and survival of mice on a high-calorie diet. Nature 444:337–342
Borriello A, Cucciolla V, Della Ragione F, Galletti P (2010) Dietary polyphenols: focus on resveratrol, a promising agent in the prevention of cardiovascular diseases and control of glucose homeostasis. Nutr Metab Cardiovasc Dis 20:618–625
Burkon A, Somoza V (2008) Quantification of free and protein-bound trans-resveratrol metabolites and identification of trans-resveratrol-C/O-conjugated diglucuronides—two novel resveratrol metabolites in human plasma. Mol Nutr Food Res 52:549–557
Burns J, Yokota T, Ashihara H, Lean ME, Crozier A (2002) Plant foods and herbal sources of resveratrol. J Agric Food Chem 50:3337–3340
Chachay VS, Macdonald GA, Martin JH, Whitehead JP, O’Moore-Sullivan TM, Lee P, Franklin M, Klein K, Taylor PJ, Ferguson M, Coombes JS, Thomas GP, Cowin GJ, Kirkpatrick CM, Prins JB, Hickman IJ (2014) Resveratrol does not benefit patients with nonalcoholic fatty liver disease. Clin Gastroenterol Hepatol 12:2092–2103.e2096
Chang CC, Lin KY, Peng KY, Day YJ, Hung LM (2016) Resveratrol exerts anti-obesity effects in high-fat diet obese mice and displays differential dosage effects on cytotoxicity, differentiation, and lipolysis in 3T3-L1 cells. Endocr J 63:169–178
Cho SJ, Jung UJ, Choi MS (2012) Differential effects of low-dose resveratrol on adiposity and hepatic steatosis in diet-induced obese mice. Br J Nutr 108:2166–2175
Crandall JP, Oram V, Trandafirescu G, Reid M, Kishore P, Hawkins M, Cohen HW, Barzilai N (2012) Pilot study of resveratrol in older adults with impaired glucose tolerance. J Gerontol A Biol Sci Med Sci 67:1307–1312
Dal-Pan A, Blanc S, Aujard F (2010) Resveratrol suppresses body mass gain in a seasonal non-human primate model of obesity. BMC Physiol 10:11
Gu H, Li K, Li X, Yu X, Wang W, Ding L, Liu L (2016) Oral resveratrol prevents osteoarthritis progression in C57BL/6J mice fed a high-fat diet. Nutrients 8:233
Gómez-Zorita S, Fernández-Quintela A, Lasa A, Hijona E, Bujanda L, Portillo MP (2013) Effects of resveratrol on obesity-related inflammation markers in adipose tissue of genetically obese rats. Nutrition 29:1374–1380
James PT, Rigby N, Leach R, Force IOT (2004) The obesity epidemic, metabolic syndrome and future prevention strategies. Eur J Cardiovasc Prev Rehabil 11:3–8
Jeon SM, Lee SA, Choi MS (2014) Antiobesity and vasoprotective effects of resveratrol in apoE-deficient mice. J Med Food 17:310–316
Kim KH, Park Y (2011) Food components with anti-obesity effect. Annu Rev Food Sci Technol 2:237–257
Kim S, Jin Y, Choi Y, Park T (2011) Resveratrol exerts anti-obesity effects via mechanisms involving down-regulation of adipogenic and inflammatory processes in mice. Biochem Pharmacol 81:1343–1351
Konings E, Timmers S, Boekschoten MV, Goossens GH, Jocken JW, Afman LA, Müller M, Schrauwen P, Mariman EC, Blaak EE (2014) The effects of 30 days resveratrol supplementation on adipose tissue morphology and gene expression patterns in obese men. Int J Obes 38:470–473
Lagouge M, Argmann C, Gerhart-Hines Z, Meziane H, Lerin C, Daussin F, Messadeq N, Milne J, Lambert P, Elliott P, Geny B, Laakso M, Puigserver P, Auwerx J (2006) Resveratrol improves mitochondrial function and protects against metabolic disease by activating SIRT1 and PGC-1alpha. Cell 127:1109–1122
Le Martelot G, Claudel T, Gatfield D, Schaad O, Kornmann B, Sasso GL, Moschetta A, Schibler U (2009) REV-ERBalpha participates in circadian SREBP signaling and bile acid homeostasis. PLoS Biol 7:e1000181
Leiherer A, Mündlein A, Drexel H (2013) Phytochemicals and their impact on adipose tissue inflammation and diabetes. Vasc Pharmacol 58:3–20
Macarulla MT, Alberdi G, Gómez S, Tueros I, Bald C, Rodríguez VM, Martínez JA, Portillo MP (2009) Effects of different doses of resveratrol on body fat and serum parameters in rats fed a hypercaloric diet. J Physiol Biochem 65:369–376
Mendes KL, de Pinho L, Andrade JM, Paraíso AF, Lula JF, Macedo SM, Feltenberger JD, Guimarães AL, de Paula AM, Santos SH (2016) Distinct metabolic effects of resveratrol on lipogenesis markers in mice adipose tissue treated with high-polyunsaturated fat and high-protein diets. Life Sci 153:66–73
Miranda J, Portillo MP, Madrid JA, Arias N, Macarulla MT, Garaulet M (2013) Effects of resveratrol on changes induced by high-fat feeding on clock genes in rats. Br J Nutr 110:1421–1428
Méndez-del Villar M, González-Ortiz M, Martínez-Abundis E, Pérez-Rubio KG, Lizárraga-Valdez R (2014) Effect of resveratrol administration on metabolic syndrome, insulin sensitivity, and insulin secretion. Metab Syndr Relat Disord 12:497–501
Nagao K, Jinnouchi T, Kai S, Yanagita T (2013) Effect of dietary resveratrol on the metabolic profile of nutrients in obese OLETF rats. Lipids Health Dis 12:8
Ohara K, Kusano K, Kitao S, Yanai T, Takata R, Kanauchi O (2015) ε-Viniferin, a resveratrol dimer, prevents diet-induced obesity in mice. Biochem Biophys Res Commun 468:877–882
Poulsen MM, Vestergaard PF, Clasen BF, Radko Y, Christensen LP, Stødkilde-Jørgensen H, Møller N, Jessen N, Pedersen SB, Jørgensen JO (2013) High-dose resveratrol supplementation in obese men: an investigator-initiated, randomized, placebo-controlled clinical trial of substrate metabolism, insulin sensitivity, and body composition. Diabetes 62:1186–1195
Qiao Y, Sun J, Xia S, Tang X, Shi Y, Le G (2014) Effects of resveratrol on gut microbiota and fat storage in a mouse model with high-fat-induced obesity. Food Funct 5:1241–1249
Reagan-Shaw S, Nihal M, Ahmad N (2008) Dose translation from animal to human studies revisited. FASEB J 22:659–661
Rivera L, Morón R, Zarzuelo A, Galisteo M (2009) Long-term resveratrol administration reduces metabolic disturbances and lowers blood pressure in obese Zucker rats. Biochem Pharmacol 77:1053–1063
Szkudelska K, Nogowski L, Szkudelski T (2009) Resveratrol, a naturally occurring diphenolic compound, affects lipogenesis, lipolysis and the antilipolytic action of insulin in isolated rat adipocytes. J Steroid Biochem Mol Biol 113:17–24
Szkudelska K, Szkudelski T (2010) Resveratrol, obesity and diabetes. Eur J Pharmacol 635:1–8
Tauriainen E, Luostarinen M, Martonen E, Finckenberg P, Kovalainen M, Huotari A, Herzig KH, Lecklin A, Mervaala E (2011) Distinct effects of calorie restriction and resveratrol on diet-induced obesity and fatty liver formation. J Nutr Metab 2011:525094
Timmers S, Konings E, Bilet L, Houtkooper RH, van de Weijer T, Goossens GH, Hoeks J, van der Krieken S, Ryu D, Kersten S, Moonen-Kornips E, Hesselink MK, Kunz I, Schrauwen-Hinderling VB, Blaak EE, Auwerx J, Schrauwen P (2011) Calorie restriction-like effects of 30 days of resveratrol supplementation on energy metabolism and metabolic profile in obese humans. Cell Metab 14:612–622
Voigt A, Ribot J, Sabater AG, Palou A, Bonet ML, Klaus S (2015) Identification of Mest/Peg1 gene expression as a predictive biomarker of adipose tissue expansion sensitive to dietary anti-obesity interventions. Genes Nutr 10
WHO (2014) Obesity and overweight. In, http://www.who.int/mediacentre/factsheets/fs311/en/
Yoshino J, Conte C, Fontana L, Mittendorfer B, Imai S, Schechtman KB, Gu C, Kunz I, Rossi Fanelli F, Patterson BW, Klein S (2012) Resveratrol supplementation does not improve metabolic function in nonobese women with normal glucose tolerance. Cell Metab 16:658–664
Acknowledgements
This research has been supported by MINECO (AGL-2015-65719-R), Fondo Europeo de Desarrollo Regional (FEDER), University of the Basque Country (ELDUNANOTEK UFI11/32), Instituto de Salud Carlos III (CIBERobn) and Basque Government (IT-572-13).
Author information
Authors and Affiliations
Corresponding author
Additional information
An erratum to this article is available at https://doi.org/10.1007/s13105-017-0593-x.
Rights and permissions
About this article

Cite this article
Fernández-Quintela, A., Carpéné, C., Fernández, M. et al. Anti-obesity effects of resveratrol: comparison between animal models and humans. J Physiol Biochem 73, 417–429 (2016). https://doi.org/10.1007/s13105-016-0544-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13105-016-0544-y