
Abstract
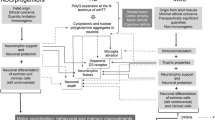
The choroid plexuses (CPs) help maintain the extracellular milieu of the brain by modulating chemical exchange between the cerebrospinal fluid and brain parenchyma, surveying the chemical and immunological status of the brain, detoxifying the brain, secreting a nutritive “cocktail” of polypeptides, and participating in repair processes following trauma. Based on recent pre-clinical studies in animal models, a novel therapeutic approach has been suggested that involves transplanting CP for treating acute and chronic brain diseases. To date most studies have focused on rodent and primate models of Huntington’s disease (HD) with demonstrations that transplants of CP can prevent the behavioral and anatomical consequences of striatal degeneration. Despite the encouraging results that lend support to the possibility of protecting vulnerable neurons in HD, critical basic science issues remain unexamined that limit the translation of the pre-clinical findings into clinical evaluations of CP transplants for HD. Here we briefly outline the logic behind using this novel cell source for transplantation, the pre-clinical data supporting this concept, and most importantly identify several critical, gating issues that remain prior to moving this approach forward in a meaningful clinical manner.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The choroid plexuses (CPs) within the brain ventricles are best known for producing the cerebrospinal fluid (CSF) and forming a physical interface between the peripheral blood and the CSF. It has also become apparent that they play pivotal roles in processes that establish, survey, and maintain the biochemical and cellular status of the central nervous system (CNS) under normal and pathological conditions. As we learn more about the participation of the CP in brain function, it becomes obvious that this small, yet complex organ is subject to subtle anatomical and physiological changes with wide-ranging consequences. There are numerous medical conditions with distinct choroidal pathology ranging from normal aging to Alzheimer’s disease and stroke (Ferrand-Drake 2001; Preston 2001; Serot et al. 2001; Emerich et al. 2004, 2005). Causal relationships between choroidal changes and many medical conditions are poorly understood and some changes in the CP are likely downstream sequelae from the primary clinical condition. But independent of whether the pathology of the CP is directly or indirectly linked to a given disease, the diseased brain from the development to adulthood is subjected to additional stress that over time may exacerbate the primary medical condition or lead to new ones. At the same time, the CP assists in recovery processes by secreting neuroprotective compounds (Chodobski and Szmydynger-Chodobska 2001; Matsumoto et al. 2003; Emerich et al. 2004, 2005) and acting as a site of neurogenesis (Blass-Kampmann et al. 1997; Kitada et al. 2001; Falk and Frisen 2002; Kotani et al. 2002; Li et al. 2002; Chouaf-Lakhdar et al. 2003; Irvin et al. 2004) suggesting both an ongoing and “as needed” role in cellular repair and replacement in the brain. These regenerative functions led to the novel idea that transplanting CP cells into ectopic sites might minimize or prevent neural degeneration and its functional consequences. Here we provide a very brief overview of the evidence that CP epithelial cells possess secretory capabilities that make them attractive candidates for transplantation. We then present the data supporting the notion of transplanting CP cells to repair the damaged CNS with an emphasis on Huntington’s disease (HD). Finally, we outline future studies that, if successful, could allow for clinical examination of CP grafts.
The Choroid Plexus Epithelial Cell as a Transplantable Secretory Cell
The basic underpinning of the hypothesis that CP cells can be used as neuroprotective transplantable cells hinges on their innate capacity to secrete potentially therapeutic compounds. The CP contains a broad array of receptors, serves as a major source of biologically active compounds, and is in an excellent location for distributing the molecules that it secretes both locally and globally to the brain. These capabilities allow the CP to monitor and respond to the biochemistry of the brain by manipulating and maintaining baseline levels of the extracellular milieu throughout the CNS (Chodobski and Szmydynger-Chodobska 2001; Stopa et al. 2001; Matsumoto et al. 2003; Emerich et al. 2004, 2005). The molecules secreted by the CP are able to gain proximal and distal access to the brain parenchyma through volume transmission, convective distribution, and intraparenchymal diffusion/receptor-mediated retrograde transport to neurons whose endings are located near the ependyma or the pia-astroglial membrane (Ferguson et al. 1991; Agnati et al. 1995; Mufson et al. 1999). In this broad view the CP plays an integral role in autocrine/paracrine and distal/endocrine-like effects on target cells in the brain by producing CSF and polypeptides and distributing them from the ventricles to cells within the brain and spinal cord.
Studies of the CP in development and aging also support its potential value in transplantation studies. The CPs develop early during embryogenesis where they help control the extra-cellular environment (Dzieglielewska et al. 2001). Within the immature brain, the blood–CSF barrier is functional and limits passage of substances from the CSF to the brain parenchyma. But while the CP is functional, its permeability is tailored for the embryonic brain allowing low molecular weight compounds to enter the brain more easily than in adulthood. Among the developmentally regulated compounds secreted by the developing CP are morphogens, mitogens, and trophic factors that guide and pattern both the general and specific growth of the CNS (Lammer et al. 1985; Bondy et al. 1990; Ved and Pieringer 1993; Yamamoto et al. 1996; Ba-Charvet et al. 1999; Hu 1999; Herbert et al. 2002). For instance, the embryonic CP contains high levels of IGF-II hypothesized to diffuse to and bind to IGF the receptors on the floor plate cells and activate their role in guiding spinal axon growth (Bondy et al. 1990). As pointed out by Strazielle and Ghersi-Egea (2000), the radial migration of cerebral cortical neurons from the ventricular and sub-ventricular zone to the cortical plate is governed by gradients of soluble factors, such as Slit proteins that are secreted by the CP (Ba-Charvet et al. 1999; Hu 1999). Hu (1999) demonstrated, in vitro, that a soluble factor related to Slit2 is secreted by the CP to aid in establishing a gradient of a repulsive cue guiding cortical neurons away from the ventricular surface. Along these same lines, Yamamoto et al. (1996) showed that the CP modulates neurite outgrowth in the developing cerebellum. Using co-cultures of explanted cerebellum and CP from fetal and infant rat, it was confirmed that CP secretes a soluble neurite-growth factor that correlates with the major milestones of cerebellar morphogenesis. The importance of the CSF distribution of soluble factors during development is also highlighted by the eloquent work of Miyan and colleagues (Mshayekhi et al. 2002; Miyan et al. 2003; Owen-Lynch et al. 2003) who showed that hydrocephalus in rats significantly impairs cortical development leading to the suggestion that the circulating CSF and its associated factors are vital for development along the entire length of the neural tube.
Most of our knowledge about the morphology and function of human CP during development and aging comes from control tissues in studies investigating changes in Alzheimer’s disease (AD). In humans, the length of CP epithelial cells decreases by about 10–11% during life (Serot et al. 2000). The aged epithelial cell cytoplasm becomes rich with Biondi Ring tangles and lipofuchsin deposits (Wen et al. 1999) and the nuclei appear irregular and flattened as the basement membrane thickens (Serot et al. 2000). The stroma also thickens and contains collagen fibers, hyaline bodies, calcifications and psammoma bodies, and the infiltrating arteries become thicker and fragmented (Shuangshoti and Netsky 1970; Serot et al. 2003). These changes are not species specific as similar changes occur in the aged mouse, rat, and human choroid epithelial cells (Sturrock 1998; Serot et al. 2001).
The functions of the CP are energy dependent and the aging CP cannot maintain its normal energy output. The synthesis of lactate dehydrogenase and succinate-dehydrogenase, critical for anaerobic respiration and oxidative phosphorylation, declines in aging rats by 9 and 26%, respectively (Ferrante and Amenta 1987). There are age-dependent increases in the number of cytochrome C oxidase deficient epithelial cells, altering the respiratory mitochondrial chain, and decreasing cellular production of ATP (Miklossy et al. 1998). Reductions in Na+K+–ATPase and the Na+K+–2Cl− co-transporter also occur (Cottrell et al. 2001). These anatomical and enzymatic deteriorations could underlie the diminution of CSF secretion that is decreased by up to 45% in aged animals (Preston 2001). Due to the decreased secretion and the simultaneously increased CSF volume caused by brain atrophy, CSF turnover is longer in elderly rats (7.9 h) than in young rats (2.2 h). In man, CSF production diminishes from 0.41 ml/min at 28 years to 0.19 ml/min at 77 years. Coupled with age-related cerebral atrophy, the turnover of CSF decreases to less than twice daily in elderly subjects versus three to four times daily in young adults. Elevated CSF/plasma ratios have been reported for several proteins during aging (Preston 2001). The cumulative effects of these changes on brain function have not been directly tested but it seems reasonable that such dramatic alterations would lead toward inadequate distribution of nutritive substances, additional cellular stress, and reduced clearance of toxic compounds all of which could play a part in age-related cognitive and motor decline or the development of specific neurological disorders.
Harnessing the Choroid Plexus for Transplantation Therapy
The profound age- and disease-related changes in CP have not only clear implications for understanding disease processes, but also raise the largely unexplored possibility that replacing damaged CP or transplanting healthy CP into local areas of cell degeneration can be therapeutic. The diminished function of the aged CP may be much like any number of diseases characterized by secretory cell dysfunction. In principle, transplanting or replacing a failing organ (such as CP) or specific cell type is a logical and direct means of restoring lost function. There is also the potential of harnessing the polypeptide synthesis of the CP through focused local delivery after transplantation into an ectopic brain region. The role of the CP in growth factor and nutrient production makes it a novel potential source of stable, dose-controlled polypeptide delivery (Emerich et al. 2004, 2005). Although the notion of using transplanting CP cells to the damaged brain regions is a new idea, the studies conducted to date support the concept and warrant further investigation.
CP isolated and maintained in vitro exerts potent neuroprotective effects (Borlongan et al. 2004a, b; Emerich et al. 2007). Conditioned media from cultured CP protected embryonic cortical neurons against serum deprivation-induced cell death. This effect was dose dependent and nearly complete when neurons were cultured with 10–30% conditioned media. These data dovetail nicely with a study where mouse CP epithelial cells were cultured with dorsal root ganglion neurons (Chakrabotty et al. 2000). After 4–5 h of co-culture, the neurons developed elongated neuronal processes with elaborate branching patterns. Similar effects have been reported by Kimura et al. (2004). CP epithelial cells that were co-cultured with hippocampal neurons robustly enhanced neurite extension within 24 h. Neurite length, the number of primary neurites, and the number of branches were all increased by co-culture with CP cells. Interestingly, this effect appeared to be primarily due to contact with the choroidal epithelial cells as these tropic effects were notably lessened when only conditioned media were used. Because the neurons extended long branched neurites even when the choroidal cells were killed by ethanol fixation, it was suggested that surface plasma membrane-bound components of the epithelial cells strongly contributed to the enhancement of neurite outgrowth. The ability of CP cells to provide a scaffold for the extension of neurites is consistent with its known production of extracellular matrices including laminin and fibronectin (Peraldi-Roux et al. 1990; Zhao 1990). The trophic and tropic effects of CP establish potentially excellent circumstances for the protection and repair of damaged CNS architecture.
The delivery of neurotrophic factors through CP has also been demonstrated in vivo. One of the first demonstrations of the potential of CP grafts was in a rodent model of spinal cord trauma. Ide et al. (2001) grafted syngeneic fragments of CP into the dorsal funiculus (C2 level) of a damaged adult rat spinal cord. At various times post-transplant, subsets of animals were evaluated histologically to confirm cell survival and determine any regenerative effect on the damaged spinal cord. Electron microscopy and fluorescence histochemistry showed that epithelial cells of the grafted CP survived well and induced regeneration of the damaged axons of the spinal cord. Injections of horseradish peroxidase into the sciatic nerve labeled regenerating fibers extending from the fasciculus gracilis into the graft within 7 days post-transplant. This effect was evident for at least 10 months. Some axons elongated rostrally into the dorsal funiculus and long-duration evoked potentials were recorded 5 mm rostral to the lesion 8–10 months after grafting. These findings indicate that CP epithelial cells have the ability to facilitate axonal growth, suggesting that they are capable of exerting both trophic and tropic effects in vivo.
Similar benefits of CP have been observed in the models of cerebral ischemia. Isolated CP obtained from rodents was tested for its neuroprotective effects in a conventional rodent model of stroke (Borlongan et al. 2004a). Rats received a 1-h middle cerebral artery (MCA) occlusion immediately followed by the transplantation of alginate-encapsulated rat CP on the cortex overlying the striatal region that would be normally infarcted following MCA occlusion. Behavioral testing on days 1–3 following surgery using the elevated body swing test and Bederson neurological examination revealed profound motor and neurological impairments in control animals that were significantly improved in the study group that received alginate-encapsulated CP transplants. Histological analysis revealed that the behavioral improvements were accompanied by a 35–40% decrease in the volume of striatal infarction as assessed by 2,3,5-triphenyltetrazolium chloride staining. Similar functional and structural benefits of encapsulated porcine CP transplants were observed in the same model (Borlongan et al. 2004b).
In a more recent study, neonatal porcine CP was encapsulated within alginate microcapsules and tested for its neuroprotective potential in a rat model of HD (Borlongan et al. 2004c; Borlongan et al. 2008). In these studies, the animals received stereotaxic transplants of either empty capsules or CP-loaded capsules directly into the striatum. Three days later, the same animals received unilateral injections of the excitotoxin quinolinic acid (QA) into the ipsilateral striatum. After surgery, animals were behaviorally tested for function of their forepaws using the placement test. As expected, the QA lesion produced a profile of anatomical and behavioral effects that resemble those seen in the clinical condition. Transplanted animals gained body weight post-surgery more rapidly than controls and also demonstrated significant improvements on the placing test. When given ten trials on the behavioral test, the control rats were only able to make one to two correct motor responses. In stark contrast, the CP-transplanted rats were indistinguishable from normal animals on this task as they made greater than nine out of ten correct responses. Nissl-stained sections demonstrated that CP transplants significantly reduced the volume of the striatal lesion produced by QA by approximately 80%. A significant protection of cholinergic but not diaphorase-positive neurons was also seen. Future studies should more carefully evaluate the extent of anatomical protection produced by CP grafts. As seen in the previously published reports in stroke models, transplants of encapsulated rat CP produced behavioral and histological protection indistinguishable from that obtained with porcine CP (Borlongan et al. 2004c, 2008).
Importantly, the data obtained in rats have been replicated in a non-human primate HD model (Emerich et al. 2006). Prior to transplantation, PCR confirmed that the porcine CP cells used in those studies expressed transthyretin and immunocytochemistry demonstrated ZO-1 and tubulin staining typical of CP epithelial cells. In vitro, CP-conditioned media enhanced the survival and preserved neurite number and length on serum-deprived neurons. Cynomolgus primates were transplanted with CP-loaded capsules into the caudate and putamen followed by QA lesions 1 week later. Control monkeys received empty capsules plus QA. When evaluated 1 month later, detailed histological analysis confirmed that the CP transplants significantly protected striatal neurons as revealed by stereological counts of NeuN-positive neurons (8% loss vs. 43% in controls) and striatum volume (10% decrease vs. 40% in controls). All in all, these data demonstrate that transplanted CP cells can be used to potently protect striatal neurons from excitotoxic damage and that this strategy may ultimately prove relevant for the treatment of HD.
The Road to the Clinic
The few transplant studies conducted to date have been generally encouraging. Still, if CP transplants are to be examined for their neuroprotective ability in HD (or any disease) numerous scientific studies will be required to answer specific unresolved questions. These studies are, of course, not exclusive of the extensive tissue procurement, monitoring and testing, quality control, and manufacturing issues for any cellular product. Below, we provide a starting outline for these future lab-to-clinic efforts.
Evaluation in Relevant and Multiple Animal Models
To date, all of the pre-clinical HD animal studies have been conducted using the QA lesion model (Borlongan et al. 2004c, 2008; Emerich and Thanos 2006; Emerich et al. 2006. Although this model does provide a useful initial assessment of the ability of a given approach to mitigate the loss of striatal neurons, the model does not faithfully recapitulate either the slow degenerative nature or the genetic component of the disease. Future studies should examine the impact of CP grafts in both more chronic chemical lesion models (i.e., 3-NP) as well as a clinically useful genetic mouse model of HD. These studies must include detailed, long-term examinations of motor and cognitive function, and should correlate behavioral changes to any potential attenuation of neuronal loss and cellular protein aggregation. Additional studies (preferably in small and large animal models) should be performed to understand the therapeutic dose–response curve for CP transplants. All of the published studies to date have used similar numbers of cells. Of note, testing a meaningful range of cells may require modification of the currently used encapsulation system. This may also be the case in any mouse studies where the current size of the alginate microcapsules is potentially large enough to preclude non-traumatic intrastriatal implantation. To this end, the need for testing the efficacy and safety of CP transplants in multiple species and multiple animal models of HD, coupled with carefully designed experiments to reveal the therapeutic and tolerable doses, and appropriate delivery approach (i.e., modifying the encapsulation technique), and timing of administration should proceed in order to realize the optimal transplant regimen.
Mechanism of Action
We and others have suggested that the mechanism by which CP transplants exert their beneficial effect is through secreted proteins with trophic activity. While reasonable, this hypothesis is untested. Assuming that CP epithelial cells are a source of numerous potentially therapeutic molecules, it could be a daunting task to determine the precise compound or combination of compounds underlying any observed benefit. To begin the address this issue we propose the following:
-
(a)
Use genomic and proteomic approaches to identify the protein and small molecule secretome of CP cells.
-
(b)
Once potential molecules are identified begin testing them alone and in combination for their neuroprotective effects. This should begin using established in vitro cell culture models. In vivo studies could then be performed in which the activity of selected compounds is blocked (potentially using antibodies, chemical blockade, or transgenic animals).
-
(c)
Demonstrate, in vivo, that the identified compound is elevated in areas where any neuroprotection is produced by CP transplants. The establishment of this information will also be invaluable for studies aimed at determining the extent of diffusion from grafted CP cells, the pharmacokinetic profile of that diffusion, the numbers of implants needed, the duration of bioactivity of the grafted cells, and so on.
We maintain the position, however, that bolus delivery of growth factors may not effectively mimic the therapeutic benefits afforded by CP transplants. CP cells likely secrete yet unknown growth factors, which may not be detected by genomic or proteomic profiling. Furthermore, the regulated secretion of growth factors by CP presents as a unique vehicle for the delivery of these substances to the brain, which will be difficult to replicate with injection of exogenous therapeutic molecules. Accordingly, the studies enumerated above are designed to reveal a mechanism of CP’s therapeutic action so that strategies can be designed to further enhance the secretory profile of CP cells and improve its transplant-mediated functional outcome.
Long-Term Viability and Function of the Grafted Cells
HD is obviously a chronic disease and any effective transplantation approach will likely be required to function on the order of years if not decades. To date, all of the studies in animal models of HD have been short term (approximately 1 month) and no systematic study has been published that quantifies the survival and function of encapsulated CP cells for even these short periods of time. Given that frequent re-implantation into the brain parenchyma is not desirable or practical, this knowledge will be crucial for moving this approach forward and understanding how an effective dose of secreted product can be titrated in the face of an ongoing degenerative process.
Identification of Optimal Donor Species and Age
Grafts of CP cells from neonatal pigs and young adult rats appear to be beneficial in the studies published to date and neonatal porcine CP has been proposed for clinical trials. It is, however, an open question whether neonatal animals are the optimal source or not. It is also unknown how long CP cells would possess a secretory profile that is appropriate. This issue is highlighted by a recent study (Emerich et al. 2007) where aged rat CP (24-month-old donor animals) was compared with young adult rat CP (3–4-month-old donor animals) for its neuroprotective capacity. Results indicated that the aged CP was metabolically less active, secreted significantly less VEGF, and was less potent in vitro and in vivo when tested for its ability to protect striatal neurons against insult. Future studies should simply test CP from different aged animals and species to identify the best potential donor age. This also raises the issue of using a xenograft for clinical trials. While xenogeneic tissue can certainly be brought to the clinic it might be useful to evaluate human cell sources as well, including CP cell lines. Aside from the practical considerations of monitoring any potential viral risks from xenogeneic sources such as pigs it is quite conceivable that lowering the technological requirements for encapsulation to maintain its immunoprotective functions for extended periods of time would make the transition from the lab to the clinic easier and more likely to succeed. An alternative approach is to use autologous human CP in order to circumvent the graft-versus-host disease complications associated with transplantation therapy. The initial harvest from autologous sources will not yield sufficient cells to treat large numbers of patients, and techniques for expanding these cells in culture might be needed to generate an ample supply of these transplantable cells. Clearly, additional studies are warranted to determine the optimal donor species and age, as well as to explore cell culture techniques designed to enhance CP cell proliferation with the aim of producing a clinical product (i.e., homogenous cell population that can be cryopreserved).
Definition and Optimization of the Cellular Composition of the CP Graft
The published CP transplant studies do not use a purified epithelial cell graft. The encapsulated cells, while primarily appearing to be epithelial cells, are clusters of cells that include portions of the undigested fibroblastic stroma. Although these clusters provide significant benefits as tested to date it is an open question whether a purified population of epithelial cells would perform comparably to a mixed, more natural population of CP cells. Ideally, a purified population could be used as this would significantly lower the amount of product characterization needed.
Safety Considerations
Once the above-said questions are answered, standard safety studies will be required to confirm the biocompatibility of the encapsulation material (if used), lack of host systemic response to graft, lack of toxic effect to the brain and peripheral organs and cells, absence of structural changes including cellular sprouting and hyperplasia, and so on.
Conclusions
Intracerebral transplants of CP epithelial cells appear capable of mitigating neural trauma in rodent lesion models of stroke and HD. With regard to HD, significant protection of striatal neurons has been reported in both rats and monkeys lesioned with the excitotoxin QA. Still, numerous scientific questions remain unanswered that, at present, limit the possibility of clinical evaluations. Here we have outlined many of these studies and suggest that successful outcomes would raise what is currently an interesting hypothesis to the level of serious clinical consideration.
References
Agnati LF, Zoli M, Stromberg I, Fuxe K (1995) Intercellular communication in the brain: wiring versus volume transmission. Neuroscience 69:711–726
Ba-Charvet NKT, Brose K, Marillat V (1999) Slit2-mediated chemorepulsion and collapse of developing forebrain axons. Neuron 22:463–473
Blass-Kampmann S, Kindler-Rohrborn A, Deissler H, D’Urso D, Rajewsky MF (1997) In vitro differentiation of neural progenitor cells from prenatal rat brain: common cell surface glycoprotein on three glial cell subsets. J Neurosci Res 48:95–111
Bondy CA, Werner H, Roberts CT Jr, LeRoith D (1990) Cellular pattern of insulin-like growth factor-I (IGF-I) and type I IGF receptor gene expression in early organogenesis: comparison with IGF-II gene expression. Mol Endocrinol 4:1386–1398
Borlongan CV, Elliott RB, Skinner SJM, Geaney M, Vasconcellos A, Emerich DF (2004a) Intraparenchymal grafts of rat choroid plexus protect against cerebral ischemia in adult rats. Neuroreport 15:1543–1547
Borlongan CV, Skinner SJM, Geaney M, Vasconcellos AV, Elliott RB, Emerich DF (2004b) Intracerebral transplants of porcine choroid plexus provide structural and functional neuroprotection in a rodent model of stroke. Stroke 35:2206–2210
Borlongan CV, Geaney M, Vasconcellos A, Elliott R, Skinner SS, Emerich DF (2004c) Neuroprotection of striatal neurons by choroid plexus grafts in QA-lesioned rats. Neuroreport 15:2521–2525
Borlongan CV, Skinner SJM, Geaney M, Emerich DF (2008) Transplants of encapsulated rat choroid plexus exert neuroprotection in a rodent model of Huntingtons disease. Cell Transplantation 16:987–992
Chakrabotty S, Kitada M, Matsumoto N, Taketomi M, Kimura N, Ide C (2000) Choroid plexus ependymal cells enhance neurite outgrowth from dorsal root ganglion neurons in vitro. J Neurocytol 10:707–717
Chodobski A, Szmydynger-Chodobska J (2001) Choroid plexus: target for polypeptides and site of their synthesis. Microsc Res Tech 52:65–82
Chouaf-Lakhdar L, Fevre-Montange M, Brisson C, Strazielle N, Gamra H, Didier-Bazes M (2003) Proliferative activity and nestin expression in periventricular cells of the adult rat brain. Neuroreport 14:633–636
Cottrell DA, Blakely EL, Johnson MA, Ince PG, Borthwick GM, Turnbull DM (2001) Cytochrome oxidase deficient cells accumulate in the hippocampus and choroid plexus with age. Neurobiol Aging 22:265–272
Dzieglielewska KM, Ek J, Habgood MD, Saunders NR (2001) Development of the choroid plexus. Microsc Res Tech 2:5–20
Emerich DF, Thanos CG (2006) In vitro culture duration does not impact the ability of encapsulated choroid plexus transplants to prevent neurological deficits in excitotoxin-lesioned rats. Cell Transplant 15:595–602
Emerich DF, Vasconcellos AV, Elliott RB, Skinner SJM, Borlongan CV (2004) The choroid plexus: function, pathology and therapeutic potential of its transplantation. Expert Opin Biol Ther 4:1191–1201
Emerich DF, Skinner SJM, Borlongan CV, Vasconcellos AV, Thanos CG (2005) The choroid plexus in the rise, fall, and repair of the brain. Bioessays 27:262–274
Emerich DF, Thanos CG, Goddard M, Skinner SJM, Geany MS, Bell WJ, Bintz B, Schnieder P, Chu Y, Rangasamy S, Babu S, Borlongan CV, Boekelheide K, Hall S, Bryant B, Kordower K (2006) Neuroprotective effects of porcine choroid plexus in a monkey model of Huntington’s disease. Neurobiol Dis 23:471–480
Emerich DF, Schneider P, Bintz BE, Hudak J, Thanos CG (2007) Aging reduces the neuroprotective capacity, VEGF secretion, and metabolic activity of choroid plexus epithelial cells. Cell Transplant 16:697–705
Falk A, Frisen J (2002) Amphiregulin is a mitogen for adult neural stem cells. J Neurosci Res 69:757–762
Ferguson IA, Schweitzer JB, Bartlett PF, Johnson EM Jr (1991) Receptor-mediated retrograde transport in CNS neurons after intraventricular administration of NGF and growth factors. J Comp Neurol 313:680–692
Ferrand-Drake M (2001) Cell death in the choroid plexus following transient forebrain global ischemia in the rat. Microsc Res Tech 52:130–136
Ferrante F, Amenta F (1987) Enzyme histochemistry of the choroid plexus in old rats. Mech Ageing Dev 41:65–72
Herbert JM, Mishina Y, McConnell SK (2002) BMP signaling is required locally to pattern the dorsal telencephalic midline. Neuron 35:1029–1041
Hu H (1999) Chemorepulsion of neuronal migration by Slit2 in the developing mammalian forebrain. Neuron 23:703–711
Ide C, Kitada M, Chakrabortty S, Taketomi M, Matsumoto N, Kikukawa S, Mizoguchi A, Kawaguchi S, Endoh K, Suzuki Y (2001) Grafting of choroid plexus ependymal cells promotes the growth of regenerating axons in the dorsal funiculus of rat spinal cord: a preliminary report. Exp Neurol 167:242–251
Irvin DK, Nakano I, Paucer A, Kornblum HI (2004) Patterns of Jagged1, Jagged2, Delta-like 1 and Delta-like 3 expression during late embryonic and postnatal brain development suggest multiple functional roles in progenitors and differentiated cells. J Neurosci Res 75:330–343
Kimura K, Matsumoto N, Kitada M, Mizoguchi A, Ide C (2004) Neurite outgrowth from hippocampal neurons is promoted by choroid plexus ependymal cells in vitro. J Neurocytol 33:465–476
Kitada M, Chakrabortty S, Matsumoto N, Taketomi M, Ide C (2001) Differentiation of choroid plexus ependymal cells into astrocytes after grafting into the pre-lesioned spinal cord in mice. Glia 36:364–374
Kotani M, Osanai T, Tajima Y, Kato H, Imada M, Kaneda H, Kubo H, Sakuraba H (2002) Identification of neuronal cell lineage-specific molecules in the neuronal differentiation of P19 EC cells and mouse central nervous system. J Neurosci Res 67:595–606
Lammer EJ, Chen DT, Hoar RM (1985) Retinoic acid embryopathy. N Engl J Med 313:837–841
Li Y, Chen J, Chopp M (2002) Cell proliferation and differentiation from ependymal, subependymal and choroid plexus cells in response to stroke in rats. J Neurol Sci 193:137–146
Matsumoto N, Kitayama H, Kitada M, Kimura K, Noda M, Ide C (2003) Isolation of a set of genes expressed in the choroid plexus of the mouse using suppression subtractive hybridization. Neuroscience 117:405–415
Miklossy J, Kraftsik R, Pillevuit O, Lepori D, Genton C, Bosman FT (1998) Curly fiber and tangle-like inclusions in the ependyma and choroid plexus-a pathogenetic relationship with the cortical Alzheimer-type changes? J Neuropathol Exp Neurol 57:1202–1212
Miyan JA, Nabiyouni M, Zendah M (2003) Development of the brain: a vital role for cerebrospinal fluid. Can J Physiol Pharmacol 81:317–328
Mshayekhi F, Draper CE, Banister CM, Pourhasem M, Owen-Lynch PJ, Miyan JA (2002) Deficient cortical development in the hydrocephalic Texas (H-Tx) rat: a role for CSF. Brain 125:1859–1874
Mufson EJ, Kroin JS, Sendera TJ, Sobreviela T (1999) Distribution and retrograde transport of trophic factors in the central nervous system: functional implications for the treatment of neurodegenerative diseases. Prog Neurobiol 57:451–484
Owen-lynch PJ, Draper CE, Mashayekhi F, Banister CM, Miyan JA (2003) Defective cell cycle control underlies abnormal cortical development in the hydrocephalic Texas rat. Brain 126:623–631
Peraldi-Roux S, Nguyen-Than B, Nao B, Hirn M, Gabrion J (1990) Choroidal ependymocytes in culture: expression of polarity and function. Int J Dev Neurosci 8:575–588
Preston JE (2001) Ageing choroid plexus-cerebrospinal fluid system. Microsc Res Tech 52:31–37
Serot JM, Béné MC, Foliguet B, Faure GC (2000) Morphological alterations of the choroid plexus in late-onset Alzheimer’s disease. Acta Neuropathol 99:105–108
Serot JM, Béné MC, Foliguet B, Faure GC (2001) Choroid plexus and ageing: a morphometric and ultrastructural study. Eur J Neurosci 14:794–798
Serot JM, Béné MC, Faure GC (2003) Choroid plexus, ageing of the brain, and Alzheimer’s disease. Front Biosci 8:515–521
Shuangshoti S, Netsky MG (1970) Human choroid plexus: morphologic and histochemical alterations with age. Am J Anat 128:73–96
Stopa EG, Berzin TM, Kim S, Song P, Kuo-Leblanc V, Rodriguez-Wolf M, Baird A, Johanson CE (2001) Human choroid plexus growth factors: what are the implications for CSF dynamics in Alzheimer’s disease. Exp Neurol 167:40–47
Strazielle N, Ghersi-Egea JF (2000) Choroid plexus in the central nervous system: biology and physiopathology. J Neuropathol Exp Neurol 59:561–574
Sturrock RR (1998) An ultrastructural study of the choroid plexus of aged mice. Anat Anz 165:379–385
Ved HS, Pieringer RA (1993) Regulation of neuronal differentiation by retinoic acid alone and in cooperation with thyroid hormone or hydrocortisone. Dev Neurosci 15:49–53
Wen GY, Wisniewski HM, Kascsak RJ (1999) Biondi ring tangles in the choroid plexus of Alzheimer’s disease and normal aging brains: a quantitative study. Brain Res 832:40–46
Yamamoto M, McCaffery P, Drager UC (1996) Influence of the choroid plexus on cerebellar development: analysis of retinoic acid synthesis. Brain Res Dev Brain Res 93:182–190
Zhao FC (1990) Four patterns of laminin-immunoreactive structure in developing rat brain. Brain Res Dev Brain Res 55:191–202
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Emerich, D.F., Borlongan, C.V. Potential of Choroid Plexus Epithelial Cell Grafts for Neuroprotection in Huntington’s Disease: What Remains Before Considering Clinical Trials. Neurotox Res 15, 205–211 (2009). https://doi.org/10.1007/s12640-009-9021-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12640-009-9021-5