
Abstract
Plant defensins are small antimicrobial proteins (AMP) that participate in the immune defense of plants through their antibacterial, antiviral and antifungal activities. PgD1 is a defensin from Picea glauca (Canadian Pine) and has antifungal activity against plant pathogens. This activity positions it as an alternative biotechnological agent to pesticides commonly used against these plant fungi diseases. The present study aimed to recombinantly produce PgD1 in Escherichia coli to characterize its in vitro antifungal potential against different phytopathogens. To achieve this, the coding gene was amplified and cloned into pET30a( +). Recombinant plasmid was subsequently introduced into E. coli for the soluble expression of defensin PgD1. To evaluate the antifungal activity of the expressed protein, the growth inhibition test was used in solid and liquid media for approximately 7 days against significant plant pathogens, that cause significant crop damage including: Botrytis cinerea, Colletotrichum gloeosporioides, Colletotrichum musae, Colletotrichum graminicola and Fusarium oxysporum. Additionally, stability assessments included temperature variation experiments and inhibition tests using dithiothreitol (DTT). The results showed that there was significant inhibition of the fungal species tested when in the presence of PgD1. Furthermore, defensin proved to be resistant to temperature variations and demonstrated that part of its stability is due to its primary structure rich in cysteine residues through the denaturation test with dithiothreitol (DTT) where the antifungal activity of PgD1 defensin was inhibited. These data indicate that recombinant PgD1 could be utilized as a plant protection technology in agriculture.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Plant defensins are a class of small cationic proteins typically composed of 45–54 amino acids. They are characterized by a high cysteine content and adopt a structural conformation featuring both mixed beta-sheet and alpha-helix motifs [1,2,3]. Functionally, these proteins serve as antimicrobial peptides integral to the plant's immune defense system due to their small size and antimicrobial activity [1, 4, 5], which is exerted through various mechanisms of action [2].
Studies have categorized plant defensins into 11 distinct groups based on nucleotide sequence similarity, specific targets, and mechanism of action [6, 7]. For instance, group 1 includes defensins such as MtDef4 and RsAFP1, which inhibit alpha-amylases, thereby interfering with the digestive processes of herbivorous insects and pathogens [8]. Group 2 encloses defensins highly expressed in response to plant damage, where the action mechanism is not fully understood [9]. PgD1 from Picea glauca belongs to Group 3 and is a well-studied defensin known for its potent antifungal properties, disrupting the fungal cell membrane, and leading to cell death. Other defensins in this group, such as RsAFP2, have also been shown to bind to specific fungal membrane components, causing membrane permeabilization and subsequent fungal cell death [10, 11]. Group 4 defensins inhibit calcium channels, which are essential for various cellular processes in pathogens. For example, Defensin-like (DEFL) peptides from Arabidopsis thaliana, such as AtPDF2.3 and AtPDF2.6, exhibit calcium channel-blocking activity, thereby disrupting pathogen cellular homeostasis [12]. Interestingly, defensins AX1 and AX2, found in Group 5, specifically target fungi but not bacteria and act by disrupting cell wall integrity and inducing oxidative stress within the fungal cells [5].
The mechanism of action for defensins in Group 6 remains largely unknown [13], while.
group 7 contains defensins such as NaD1 from Nicotiana alata, which function through the production of reactive oxygen species (ROS). NaD1 binds to fungal cell walls and triggers an oxidative burst, leading to fungal cell death [14]. Brassica campestris defensins are classified into Group 8. These defensins have shown broad-spectrum antimicrobial activity, although specific examples and mechanisms are still being elucidated [15]. Group 9 includes defensins like RsAFP2, known to interact with sphingolipids in fungal membranes. This interaction disrupts membrane structure and function, leading to fungal cell death [16]. Defensins PsD1 and MsDef1 in Group 10 interact with pathogen genetic material, specifically binding to DNA and RNA. This interaction inhibits nucleic acid synthesis and disrupts essential cellular processes in the pathogens [17]. Lastly, Group 11 comprises defensins such as Cassia fistula defensin, which inhibit trypsin, a protease essential for the digestive processes of herbivorous insects and pathogens. This inhibition impairs the pathogen's ability to degrade proteins for nutrition, thereby reducing its virulence [18].
Due to their non-toxicity to human, plant, and animal cells [1, 9, 19,20,21,22,23], plant defensins represent a promising biotechnological alternative to chemical control methods [2, 24] as they exhibit activity against important crop pathogens, including Botrytis cinerea, Alternaria brassicae (Berk.), Fusarium culmorum (W.G. Smith) Sacc., Verticillium dahliae (Kleb.), Rhizoctonia solani (Kühn), and Magnaporthe oryzae (B. Couch) [24]. Therefore, they can serve as potential substitutes for chemical control or be utilized in combination with chemicals to mitigate environmental damage [25].
One such promising defensin is PgD1 from Picea glauca (Moench) Voss., which consists of a 50 amino acid protein and operates through the mechanism of membrane permeabilization. Studies have demonstrated its efficacy against fungal pathogens such as Cylindrocladium floridanum (Sobers & C.P. Seym), Fusarium oxysporum (Schlecht.), and Nectria galligena (Bress.) [26, 27]. PgD1 has structural similarity with other pine defensins like RsAFP1, which are antifungal defensins found in the seeds of radish and have shown great potential in inhibiting fungal growth. Some authors highlighted the effectiveness of the RsAFP1 defensin against the species Botrytis cinerea, Botryosphaeria dothidea, Colletotrichum acutatum, C. gloeosporioides, C. fragariae, Monilinia vaccinii-corymbosi, Phytophthora fragariae, Xanthomonas fragariae, Fusarium oxysporum, Sclerotinia sclerotiorum and Candida albicans [28,29,30,31].
Plant defensins are commonly isolated from different plant parts, including roots, seeds, leaves, flowers, and fruits, across a wide range of species [7, 32]. However, the isolation of defensins from plant tissues may not be efficient, as it is subject to seasonal variations and cultivar flowering patterns, resulting in reduced productivity [33]. In our study, we employed recombinant expression of PgD1 defensin in E. coli followed by its purification. This method, as demonstrated by recent research has proven to be efficient and yield high quantities of the desired product [34].
In light of the growing need for sustainable agricultural practices, this study aims to contribute to crop protection by elucidating the expression, purification, and potent antifungal activity of recombinantly expressed PgD1 defensin, offering insights into its potential as a sustainable solution for plant disease management.
Material and Methods
Cloning and Bioinformatic Analysis
The gene encoding the P. glauca full-length PgD1 protein (83 amino acids) was deposited in the Gen Bank under the accession code AAR84643. The full-length protein of 83 amino acids has a predicted molecular mass of 8895 Da. However, the protein is processed at the N-terminus to remove a 33-amino acid signal peptide, which likely targets this protein to the cell secretory pathway. Therefore, the deduced mature protein is 50 amino acids long [35].
In the present work we amplified and cloned the mature protein coding gene, which corresponds to the last 50 amino acids of the PgD1 protein, which was kindly provided by Dr. Armand Segúin from Natural Resources Canada and the Canadian Forest Service. Synthetic oligonucleotide primers were used to amplify the DNA fragment containing the PgD1 protein coding sequence using standard PCR conditions.
The upstream primer (5'AAGGCC ATG GGTCGAACCTGCAAA ACCCCAAGC 3') included an Nco I restriction site (underlined) by adding two extra amino acids (Met and Gly) at the N-terminal end of PgD1 while the downstream primer (5'GGTGCTCGAGATCAAGGGCAGGGCTTGGAGACGTA3') included an Xho I restriction site (underlined).
Following purification using the PureLink Quick Gel Extraction Kit (Invitrogen®), the amplified fragments were adenylated using Taq DNA polymerase and ligated into pGEM-T-Easy® cloning vector using T4 DNA ligase. The ligation product was transformed into electrocompetent E. coli DH10B cells. Recombinant clones were grown in a liquid LB medium with antibiotic selection, and plasmid DNA was purified using the PureLink Quick Plasmid Miniprep Kit (Invitrogen®).
The purified fragment was cloned into pET30a( +) expression vector between the Nco I and Xho I restriction sites, therefore the full-length expressed protein includes a hexa-histidine tag followed by an S tag™ at the N-terminal end of the protein, producing a full-length protein of 94 amino acids with a predicted molecular weight of 13.7 kDa. The S-tag is a short peptide sequence used for protein purification and detection in biotechnology, and it is a trademark of Novagen, now part of Merck KGaA.To confirm the identity of the cloned gene and ensure the absence of mutations, sequencing of the cloned gene was performed using the Sanger method, and the nucleotide sequence was compared to the PgD1 sequence present in the Gen Bank database using the BLAST tool (http://blast.ncbi.nlm.nih.gov/Blast.cgi). Furthermore, the sequence was submitted to Phyre and Pymol software to predict the three-dimensional structure.
Expression and Purification of PgD1 Defensin
The recombinant plasmid pET30a( +): PgD1, was electroporated into E. coli Rosetta (DE3) cells and the transformed cells were inoculated into LB liquid medium containing kanamycin and chloramphenicol as selection markers and incubated at 37ºC until an OD600 nm of 0.6 was achieved. Induction was 0.05 mM IPTG and the culture was grown at 17 °C for 24 h. For purification, cells were suspended in Tris–HCl pH 8.0; 300 mM NaCl and 1.5 mM imidazole (Buffer A), containing 0.1 mM PMSF, lysozyme (20 mg/mL) and DNAse I (5 μg/mL), and sonication was performed. Cellular debris was removed by centrifugation and the supernatant was applied to a Ni–NTA Agarose column pre-equilibrated with buffer A. The resin was washed with buffer B (50 mM Tris–HCl pH 8.0; 300 mM NaCl; 40 mM imidazole) and proteins were eluted in buffer C (500 mM imidazole; 50 mM Tris–HCl pH 8.0).
The active fractions were dialyzed against buffer A, lyophilized, and stored at 4ºC. The last step of purification consisted of dialysis to remove excess imidazole with 50 mM Tris HCl buffer pH 7.0 at 4ºC. After dialysis, the protein concentration was measured using the Thermo Scientific NanoDrop® 2000 spectrophotometer. Protein purification was confirmed by SDS-Page and Western Blotting. The proteins were transferred to a nitrocellulose membrane and treated with a blocking solution, anti-His Tag primary antibody, and anti-mouse secondary antibodies anchored to the alkaline phosphatase enzyme, for detection of the target protein.
In Vitro Antifungal Activity Assay
The evaluation of the antifungal potential of PgD1 defensin was carried out using two techniques: the growth inhibition test in solid medium in a Petri dish, and in liquid medium in a 96-well plate.
For the first technique, fungal growth was monitored by inoculating fungi (B. cinerea, C. gloeosporioides, C. musae, C. graminicola, and F. oxysporum) on Petri dishes and inserting 0.5 cm paper discs containing 2 mM of PgD1 defensin into plates with Potato Dextrose Agar (PDA). The plates were stored at 28 °C for 6 days. After this period, growth was measured.
For the second technique, fungal growth was monitored in microtiter plates containing 200 µL of liquid Potato Dextrose medium, inoculated with 2000 spores of each fungus (B. cinerea, C. gloeosporioides, C. musae, C. graminicola, and F. oxysporum), and supplemented with varying concentrations of PgD1 defensin (0.5 µM, 0.75 µM, 1 µM, 1.5 µM, 2 µM, and 4 µM). The plates were stored at 28 °C, and absorbance was measured at 595 nm for 6 days. Percentages of growth inhibition were assessed by analyzing the area under the curve (AUC) of the growth curves for each replicate (three).
All experiments were repeated at least twice. The means were subjected to statistical analysis using the Student's t-test.
Temperature Variation Resistance Test
For the thermal stability assessment of PgD1, fungal growth was monitored in microtiter plates containing 200 µL of liquid Potato Dextrose medium, inoculated with 2000 spores of C. gloeosporioides and supplemented with 2 μM of PgD1 defensin previously treated at different temperatures. Briefly, the protein was incubated at 25 °C, 50 °C, 75 °C, or 100 °C for 30 min before being added to the growth media. The plates were stored at 28 °C for 3 days, and absorbance was measured at 595 nm. Growth inhibition percentages were determined by analyzing the area under the curve (AUC) of the growth curves for each replicate (three).
Denaturation Assay Using Dithiothreitol (DTT)
For the denaturation assay, fungal growth was monitored in microtiter plates containing 200 µL of liquid Potato Dextrose medium, inoculated with 2000 spores of C. gloeosporioides, supplemented with 2 μM of PgD1 defensin previously treated with 0.1 mM or 1 mM DTT. Defensin PgD1 was treated with DTT for 1 h and added to the wells containing C. gloeosporioides spores in liquid media, and incubated at 28 °C over three days and compared to a control reaction without DTT.
Results
Cloning and Bioinformatic Analysis
A DNA fragment encompassing the PgD1 sequence was successfully amplified via PCR, resulting in a specific product of 154 bp. After amplification, the DNA fragment was ligated into the pGEM vector. The ligated construct was then transformed into E. coli DH10B cells. Positive clones harboring the desired insert were identified through colony PCR screening, and one colony was propagated in LB medium and subjected to plasmid extraction using the PureLink Quick Plasmid Miniprep kit (Invitrogen®). The extracted plasmid, designated pGEM::Def3, was further characterized by PCR and subcloned into the pET30a( +) vector using the Nco I and Xho I restriction sites, resulting in the construct pET30a( +)::Def. This construct was transformed into electrocompetent E. coli DH10B cells. Finally, the sequence of the recombinant vector pET30a( +)::Def2 was confirmed by sequencing, with the sequence data provided by the sequencing service displayed in Fig. 1.

Sequencing of pET30::Def2
The obtained sequence was analyzed using the BLAST tool (http://blast.ncbi.nlm.nih.gov/Blast.cgi) to observe the similarity between the pET30::Def2 clone sequence and the PgD1 defensin sequence present in the database under GenBank accession number AAR84643. The sequence obtained showed 100% identity with the PgD1 gene of P. glauca, as shown in Fig. 2.

Search results using BLAST comparing the sequence of interest (query) against a database (subject). The sequence of interest from the pET30( +)::Def2 clone amplified by PCR.
The defensin PgD1 amino acid sequence was submitted to the Phyre and Pymol software programs to the 3D structure which enabled analysis of specific regions of the proteins as shown in Fig. 3.

3D structures of PgD1 P. glauca defensin generated according to the bioinformatics program Phyre 2 and analyzed in Pymol 3.0 highlighted positive charges in purple
Hydrogen bonds and repulsive interactions contribute to the stability of protein secondary structures like alpha helices and beta sheets, while disulfide bonds (Fig. 4A) provide additional stability to the tertiary structure, crucial for maintaining the protein's integrity amidst varying environmental conditions. In antimicrobial proteins, positive charges (Fig. 3), often attributed to amino acid residues like lysine or arginine, facilitate electrostatic interactions with negatively charged components on microbial cell surfaces. This interaction aids in membrane disruption and penetration into microbial cells, where the antimicrobial protein can exert its activity, targeting vital intracellular components and processes, ultimately leading to microbial cell death.

Alignment, generated to the bioinformatics program MEGA 11 and a structural comparison using the program Pymol 3.0. PgD1 (A) defensin amino acid predicted structure compared to another plant defensin RsAFP1 (B) that exhibit known antifungal activity (C) alignment of the amino acid sequences of A and B identical amino acids are shaded in black
The striking resemblance in sequence (Fig. 4C) and conformation (Fig. 4A and B) between the defensin PgD1 and RAsFP1 suggests a potential similarity in their antifungal properties. The first 33 amino acids of PgD1 sequence (absent in RsAFP1) correspond to the signal peptide that is removed to form the mature protein. The signal peptide is also observed as a helix in PgD1 3D structure (Fig. 4A) but absent in RsAFP1 (Fig. 4B). The shared structural characteristics, particularly in their 3D arrangements, likely represent the functional moiety responsible for their antimicrobial activity.
Expression and Purification of PgD1 Defensin
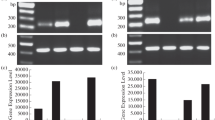
For the expression of the recombinant protein PgD1 (pET30a( +)::Def2), E. coli BL21(DE3) was chosen. This strain contains the lambda DE3 prophage, which includes the gene for T7 RNA polymerase under the control of a lacUV5 promoter. This setup allows for the induction of T7 RNA polymerase expression with IPTG. During the expression, various optimization procedures were tested, including different concentrations of IPTG and various induction temperatures. The optimal condition determined for our process was an induction temperature of 17 °C, which resulted in enhanced expression of the protein in the soluble fraction. To identify the expressed PgD1 defensin, 15% SDS-PAGE was utilized, allowing for the separation of proteins based on their molecular size. As depicted in Fig. 5, an intense band is visible in lane three, corresponding to a protein with a molecular mass of approximately 14 kDa—matching the expected size for the protein. In lane seven, the presence of PgD1 is observed post-purification, confirming the effectiveness of the purification protocols employed. Lanes four, five, and six exhibit the absence of the 14 kDa band. This absence is attributed to the histidine tail of PgD1 defensin binding to the Ni–NTA agarose during the purification process, indicating successful adherence to the chromatographic column as anticipated.

SDS-Page product of the recombinant protein pET30a( +)Def2. Lane 1: Thermo-scientific marker; Lane 2: pET30a( +) without insert; Lane 3: Expression supernatant. Lane 4: first elution step “Flow through”; Lane 5: elution with 40 mM imidazole wash buffer; Lane 6: elution with 80 mM imidazole wash buffer; Lane 7: pET30a( +)Def2 after elution with 300 mM imidazole
The Western Blot technique was used to demonstrate the presence of the protein through its histidine tail. The proteins were transferred to the nitrocellulose membrane and treated with a blocking solution, anti-His Tag primary antibody, and anti-mouse secondary antibodies anchored in the alkaline phosphatase enzyme, for detection of the target protein. Figure 1S shows a band signaling a protein with approximately 14 kDa, which indicates the presence of the PgD1 defensin (Fig. 1S—supplementary material).
In vitro Antifungal Activity Assay
Antifungal activity assays were conducted in microtiter plates against plant pathogenic fungi using varying concentrations of recombinant PgD1, ranging from 0.5 µM to 4.0 µM. The results demonstrated that PgD1 defensin significantly inhibited the growth of C. gloeosporioides at concentrations from 0.5 µM to 4.0 µM (Fig. 6).

Antifungal Activity of Defensin PgD1 Against C. gloeosporioides. This figure illustrates fungal growth curves after 6 days of treatment with varying concentrations of recombinant PgD1 defensin, ranging from 0.5 µM to 4.0 µM. Statistically significant differences in fungal growth inhibition are indicated, with a significance threshold set at p < 0.05. Analyses were performed using GraphPad Prism software, version 10. The asterisk indicates a significant difference from the controls (* = 0.05, ** = p < 0.01, *** = p < 0.001 and **** = p < 0.0001)
The inhibition of the growth of other strains of C. gloeosporioides (isolated from different plants) was also tested. The tests with C. gloeosporioides showed very similar results at concentrations of 2 µM and 4 µM. Consequently, tests on other Colletotrichum fungi were conducted using only the 2 µM concentration. We can see that the PgD1 defensin promoted the inhibition of fungal growth for all strains of C. gloeosporioides (Fig. 7), corroborating the results of the previous experiment (Fig. 6). In the inhibition test with C. graminicola, PgD1 defensin reduced fungal growth but did not completely inhibit it (see Fig. 8).

Antifungal Activity of Defensin PgD1 Against C. gloeosporioides Isolated from Various Plants. This figure displays growth curves of C. gloeosporioides after 6 days of treatment with 2 µM of recombinant PgD1 defensin. Each panel represents the fungus isolated from a different plant: (A) onion, (B) guava, (C) apple, and (D) pepper. Statistically significant differences in fungal growth inhibition are indicated, with a significance threshold set at p < 0.05. Analyses were performed using GraphPad Prism software, version 10. The asterisk indicates a significant difference from the controls (* = 0.05, ** = p < 0.01, *** = p < 0.001 and **** = p < 0.0001)

Antifungal Activity of Defensin PgD1 Against C. musae and C. graminicola. This figure displays growth curves of (A) C. musae and (B) C. graminicola after 6 days treatment with 2 µM of recombinant PgD1 defensin. Statistically significant differences in fungal growth inhibition are indicated, with a significance threshold set at p < 0.05. Analyses were performed using GraphPad Prism software, version 10. The asterisk indicates a significant difference from the controls (* = 0.05, ** = p < 0.01, *** = p < 0.001 and **** = p < 0.0001)
In the test with C. musae total inhibition was observed at a concentration of 2 µM, as shown in Fig. 8.
Inhibition was also tested for the fungi B. cinerea and F. oxysporum, using defensin concentrations of 0.5 µM to 4 µM, measuring growth after 7 days. The results show that there was complete inhibition for the fungus B. cinerea, however for the fungus F. oxysporum there was a significative reduction in growth, but there was no total inhibition (Fig. 9).

Antifungal Activity of Defensin PgD1 Against B. cinerea and F. oxysporum. This figure depicts the antifungal effects of varying concentrations of recombinant PgD1 defensin (0.5 µM to 4 µM) on the growth of (A) B. cinerea and (B) F. oxysporum. Differences in fungal growth inhibition were considered statistically significant at a p-value of less than 0.05. Data analysis was conducted using GraphPad Prism software, version 10
The antifungal activity assay, carried out in a Petri dish, against phytopathogenic fungi was carried out with a concentration of 2 µM of recombinant PgD1. The results showed that PgD1 defensin reduces the growth of C. gloeosporioides, B. cinerea and F. oxysporum, however only for the fungus B. cinerea was this reduction statistically significant (Fig. 10). The asterisk indicates a significant difference from the controls (* = 0.05, ** = p < 0.01, *** = p < 0.001 and **** = p < 0.0001).

Antifungal Activity of Defensin PgD1 on Petri Dishes. This figure demonstrates the antifungal effects of 2 µM recombinant PgD1 defensin against fungal growth in Petri dishes for (A) C. gloeosporioides, (B) B. cinerea, and (C) F. oxysporum.. Statistically significant differences in fungal growth inhibition were considered at a p-value of less than 0.05. Analyses were performed using GraphPad Prism software, version 10. The asterisk indicates a significant difference from the controls (* = 0.05, ** = p < 0.01, *** = p < 0.001 and **** = p < 0.0001)
Temperature Variation Resistance Test
The stability test at different temperatures showed that PgD1 defensin against C. gloeosporioides is resistant to temperature variation. As shown in Fig. 11 the antifungal activity remains above 90% even when the defensin is subjected to heating at 50ºC, 75ºC, and 100ºC.

Assessment of the Thermal Resistance of Defensin PgD1. This figure illustrates the effectiveness of PgD1 defensin after heat treatment by comparing the percentage of fungal growth inhibition at various temperatures to a control (25 °C) with no heat treatment and assumed 100% growth. The results show that fungal growth inhibition was 98% at 50 °C, 96% at 75 °C, and 95% at 100 °C. The statistical significance of differences was determined at a p-value less than 0.05. Data analysis was conducted using GraphPad Prism software, version 10
Denaturation Assay Using Dithiothreitol (DTT)
Dithiothreitol (DTT) is a denaturing agent that cleaves the disulfide bonds formed between cysteine residues [36]. The PgD1 defensin denaturation test used 2 µM of the protein treated with DTT in two concentrations 0.1 mM and 1 mM. The results showed that in the presence of DTT in both concentrations, defensin PgD1 was completely inhibited, with the growth of the fungus C. gloeosporioides occurring at the same rate as the control sample (without defensin) (Fig. 12).

Evaluation of Defensin PgD1 treated with DTT. Samples were treated with 0.1 mM and 1 mM DTT concentrations, with and without Defensin PgD1, compared to controls (without Def. and DTT; and with Def without DTT). Differences were considered significant at the level of p < 0.05. The analyses were performed using the GraphPad Prism 10 software. The asterisk indicates a significant difference from the controls (* = 0.05, ** = p < 0.01, *** = p < 0.001 and **** = p < 0.0001)
Discussion
Almost 30 years ago, the first articles were published describing plant defensins as part of the innate immune system of plants [33]. From its discovery, until now, we've learned about the role of these proteins in protecting plants against invading agents like fungi and bacteria. Today, these defensins are explored as antimicrobial proteins (AMPs) against numerous pathogens. In our study, we expressed and purified PgD1, a defensin from P. glauca, and tested the capacity to inhibit fungal growth. This protein is rich in cysteines and has a molecular weight of 13.752 kDa.
PgD1 exhibits similarities in terms of cysteine content and molecular weight with other defensins, such as defensin NaD1 (Nicotiana alata) [37], defensin HsAFP1 (Heuchera sanguinea) [38] and defensin RsAFP1 (Raphanus sativus) [28]. These defensins also demonstrate high cysteine content, which is characteristic of their structure and stability and this conservation suggests that the structural and functional roles of cysteines are critical in maintaining not only the stability but also the antimicrobial activity of these proteins. The presence of disulfide bonds is also necessary for pore formation in membranes, which is fundamental to antimicrobial activity [6, 39]. Studies found that antimicrobial activity is closely associated with the stability of the β-hairpin structures, and removing disulfide bonds significantly diminishes their antimicrobial function [40]. This was confirmed in our study through the DTT denaturation test (Fig. 12), which showed a complete loss of antifungal function after disulfide bond reduction. Other amino acid residues in the primary protein structure are also crucial. In the study on the RsAFP1 defensin, it was observed that mutating specific amino acids leads to a loss of function [41]. Since PgD1 and RsAFP1 defensins share these residues (Fig. 4), part of their antifungal properties can be attributed to these amino acids [39].
The recombinant defensin PgD1 exhibited antifungal activity against five phytopathogenic fungi: C. gloeosporioides, C. musae, C. graminicola, F. oxysporum, and B. cinerea. It has been found that PgD1 also has activity against F. oxysporum [35].
We specifically chose different species within the Colletotrichum genus because each one is a significant pathogen to various important crops. For instance, C. gloeosporioides is a known pathogen of apples, C. musae affects bananas, and C. graminicola is detrimental to maize. By testing the defensin against these different species, we aim to demonstrate its broad-spectrum activity and its potential applicability in diverse agricultural contexts.
At the time of this writing, no studies had examined PgD1 against Colletotrichum and Botrytis species. However, it has been reported that PaDef, another pine defensin, also has antifungal activity against C. gloeosporioides and B. cinerea [42]. Studies have shown that the pine defensins PsDef and PgD5 are effective against B. cinerea and F. oxysporum, respectively [43].
Besides efficient expression, plant defensins are also resistant to degradation caused by temperature changes. This makes them suitable for various biotechnological uses including application in agriculture (antimicrobial activity) and health (anticancer activity, antimalarial activity) [44,45,46]. The PgD1 defensin has shown stability under varying temperatures, which aligns with findings from other studies [47, 48].
In this study, we have demonstrated the potent antifungal activity of a plant defensin against significant phytopathogens. We also developed a robust and efficient recombinant expression system, which produced a thermoresistant protein that can be easily produced and purified on a large scale. [49, 50].
Given its stability, relatively simple production, and excellent potential to inhibit phytopathogenic fungi, PgD1 defensin can be a viable alternative or complement to pesticides, reducing the socio-environmental impacts of chemical pesticides. Using defensins as antifungals represents a promising biotechnological innovation that warrants further research for improving environmental health.
Data Availability
The data that support the findings of this study are available on request from the corresponding author, M.L.B.M.
References
da Silva Gebara R, Taveira GB, de Azevedo Dos Santos L, Calixto SD, Simão TLBV, Lassounskaia E, Muzitano MF, Teixeira-Ferreira A, Perales J, Rodrigues R, de Oliveira Carvalho A, Gomes VM (2020) Identification and characterization of two defensins from capsicum annuum fruits that exhibit antimicrobial activity. Probiotics Antimicrob Proteins 12(3):1253–1265. https://doi.org/10.1007/s12602-020-09647-6
Vriens K, Cammue B, Thevissen K (2014) Antifungal plant defensins: mechanisms of action and production. Molecules 19:12280–12303
Cornet B, Bonmatin J-M, Hetru C, Hoffmann JA, Ptak M, Vovelle F (1995) Refined three-dimensional solution structure of insect defensin A. Structure 3:435–448. https://doi.org/10.1016/S0969-2126(01)00177-0
Lima AM, Azevedo MIG, Souza LM et al (2022) Plant antimicrobial peptides: An overview about classification, toxicity and clinical applications. Int J Biol Macromol 214:10–21. https://doi.org/10.1016/j.ijbiomac.2022.06.043
Lacerda AF, Vasconcelos ÃRAR, Pelegrini PCB, Grossi De Sá MF (2014) Antifungal defensins and their role in plant defense. Front Microbiol 5:116
Thomma BPHJ, Cammue BPA, Thevissen K (2002) Plant defensins. Planta 216(2):193–202
De Coninck B, Cammue BPA, Thevissen K (2013) Modes of antifungal action and in planta functions of plant defensins and defensin-like peptides. Fungal Biol Rev 26(4):109–120
Gao AG, Hakimi SM, Mittanck CA, Wu Y, Woerner BM, Stark DM ... Hiatt WR (2000) Fungal pathogen protection in potato by expression of a plant defensin peptide. Nat Biotechnol 18(12): 1307–1310
Lay FT, Anderson MA (2005) Defensins–components of the innate immune system in plants. Curr Protein Pept Sci 6(1):85–101
Sagaram US, Pandurangi R, Kaur J, Smith TJ, Shah DM (2011) Structure–activity determinants in antifungal plant defensins MtDef4 and MtDef5: Novel insights into functional significance of prosegment. J Biol Chem 286(39):35641–35649
Almeida MS, Cabral KM, Zingali RB, Kurtenbach E (2000) Characterization of two novel defense peptides from pea (Pisum sativum) seeds. Arch Biochem Biophys 378(2):278–286
De Jesus MB, Franco OL (2022) Antifungal Mechanisms of Plant Defensins: An Overview. In: Plant Antimicrobial Peptides. Springer, Cham
Parisi K, Shafee TM, Quimbar P (2019) The evolution, function and mechanisms of action for plant defensins. Semin Cell Dev Biol 88:107–118
van der Weerden NL, Bleackley MR, Anderson MA (2013) Properties and mechanisms of action of naturally occurring antifungal peptides. Cell Mol Life Sci 70(19):3545–3570
Stotz HU, Thomson JM, Wang Y, Kuc J (2000) Defense genes in Brassica napus: Inducible chitinase and β-1,3-glucanase mRNAs in leaves detected by PCR amplification. Plant Mol Biol 42(1):103–109
Thevissen K, Kristensen HH, Thomma BP, Cammue BP, François IE (2007) Therapeutic potential of antifungal plant and insect defensins. Drug Discov Today 12(21–22):966–971
Caruso C, Bellincampi D, Camardella L, Muccilli V, Faoro F, Falcone G ... Bertini L (2001) A basic protein from wheat kernel with antifungal activity. Phytochemistry 57(7): 921–927
Carvalho AO, Gomes VM (2009) Plant defensins—prospects for the biological functions and biotechnological properties. Peptides 30(5):1007–1020
Van Der Weerden NL, Anderson MA (2013) Plant defensins: Common fold, multiple functions. Fungal Biol Rev 26(4):121–131. https://doi.org/10.1016/j.fbr.2012.08.004
Parisi K, Doyle SR, Lee E, Lowe RGT, van der Weerden NL, Anderson MA, Bleackley MR (2019) Screening the Saccharomyces cerevisiae nonessential gene deletion library reveals diverse mechanisms of action for antifungal plant defensins. Antimicrob Agents Chemother 63(11). https://doi.org/10.1128/aac.01097-19
Cools TL et al (2017) Antifungal plant defensins: Increased insight into their mode of action as a basis for their use to combat fungal infections. Future Microbiol 12(5):441–454. https://doi.org/10.2217/fmb-2016-0181
Hegedüs N, Marx F (2013) Antifungal proteins: more than antimicrobials? Fungal Biol Rev 26(4):132–145. https://doi.org/10.1016/j.fbr.2012.07.002
Tavares LS et al (2008) Biotechnological potential of antimicrobial peptides from flowers. Peptides 29(10):1842–1851. https://doi.org/10.1016/j.peptides.2008.06.003
Rojas Arias ACR, Espitia HMZ (2010) Plant defensins and their potential use as pest controllers in agriculture. Acta Biol Colomb 15(3):33–46
Pérez NS, Obregón GAE, Avera YH, et al (2022) Transgenic soybean modification event for greater resistance to Asian rust. Anais do ACC. Havana 12(3). http://scielo.sld.cu/scielo.php?script=sci_arttext&pid=S2304-01062022000300007&lng=es&nrm=iso. Acesso em 09 fev. 2024
Picart P, Pirttila AM, Raventos D, Kristensen HH, Sahl HG (2012) Identification of spruce defensin encoding genes: Characterization of PgD5, a conserved spruce defensin with strong antifungal activity. BMC Plant Biol 12:180
Azmi S, Hussain MK (2021) Analysis of structures, functions, and transgenicity of phytopeptides defensin and thionin: a review. Beni-Suef Univ J Basic Appl Sci 10(5). https://doi.org/10.1186/s43088-020-00093-5
Hammerschlag FA (2004) In vitro inhibitory activity of antimicrobial peptides cecropin, α-thionin DB4, and γ-thionin RsAFP1 against several pathogens of strawberry and highbush blueberry. HortScience HortSci 39(5):1053–1055. https://doi.org/10.21273/HORTSCI.39.5.1053
Sadhu S, Jogam P, Gande K et al (2023) Expression of the radish defensin gene (RsAFP2) in chickpea (Cicer arietinum L.) confers resistance to Fusarium wilt disease. Mol Biol Rep 50:11–18. https://doi.org/10.1007/s11033-022-08021-9
Djami-Tchatchou AT, Tetoria M, Godwin J, Codjoe JM, Li H, Shah DM (2023) Small cationic cysteine-rich defensin-derived antifungal peptide controls white mold in soybeans. J Fungi 9:873. https://doi.org/10.3390/jof9090873
Vriens K, Cools TL, Harvey PJ, Craik DJ, Braem A, Vleugels J, De Coninck B, Cammue BP, Thevissen K (2016) The role of defensins in plant defense. Peptides 75:71–79
Hoskin DW, Ramamoorthy A (2008) Studies on anticancer activities of antimicrobial peptides. Biochim Biophys Acta 1778:357–375. https://doi.org/10.1016/j.bbamem.2007.11.008
Broekaert WF, Terras FR, Cammue BP, Osborn RW (1995) Plant defensins: novel antimicrobial peptides as components of the host defense system. Plant Physiol. Aug;108(4):1353-8. https://doi.org/10.1104/pp.108.4.1353
Liu Y, Liu L, Yang C et al (2022) Identificação molecular e atividade antifúngica de uma defensina (PaDef) de Spruce. J Plant Growth Regul 41:494–506. https://doi.org/10.1007/s00344-021-10316-3
Pervieux I, Bourassa M, Laurans F et al (2004) A spruce defensin showing strong antifungal activity and increased transcript accumulation after wounding and jasmonate treatments. Physiol Mol Plant Pathol 64:331–341. https://doi.org/10.1016/j.pmpp.2004.09.008
Cui X, Huang Y, Peng Z, Li Z, Cen S (2024) Mammalian antimicrobial peptides: defensins and cathelicidins. In: Tang Y-W, Hindiyeh MY, Liu D, Sails A, Spearman P, Zhang J-R (eds) Mol Medical Microbiol, 3rd edn. Academic Press, pp 551–573. https://doi.org/10.1016/B978-0-12-818619-0.00103-9
Lay FT, Mills GD, Poon IK, Cowieson NP, Kirby N, Baxter AA, Van Der Weerden NL, Dogovski C, Perugini MA, Anderson MA, Kvansakul M, Hulett MD (2012) The molecular basis for the antifungal activity of plant defensins. J Biol Chem 287:19961
Vriens K, Cools TL, Harvey PJ, Craik DJ, Spincemaille P, Cassiman D, Braem A, Vleugels J, Nibbering PH, Drijfhout JW, De Coninck B, Cammue BP, Thevissen K (2015) The antifungal plant defensin PvD1 targets the lipid transfer protein LTP1. PLoS ONE 10:e0132701–e0132701
Shalovylo YI, Yusypovych YM, Hrunyk NI, Roman II, Zaika VK, Krynytskyy HT, Nesmelova IV, Kovaleva VA (2021) Seed-derived defensins from Scots pine: structural and functional features. Planta 254(6):129. https://doi.org/10.1007/s00425-021-03788-w
Wang J, Dou X, Canção J, Outros (2019) Antimicrobial peptides: promising alternatives in the post-antibiotic era. Med Res Rev 39: 831–859. https://doi.org/10.1002/med.21542
Samblanx GW, Goderis IJ, Thevissen K, Raemaekers R, Fant F, Borremans F et al (1997) Mutational analysis of a plant defensin from radish (Raphanus sativus L.) reveals two adjacent sites important for antifungal activity. J Biol Chem 272:1171–1179
Liu Y, Liu L, Yang C et al (2022) Molecular Identification and Antifungal Activity of a Defensin (PaDef) from Spruce. J Plant Growth Regul 41:494–506. https://doi.org/10.1007/s00344-021-10316-3
Kovalera V, Krynytskyy H, Gout I et al (2011) Recombinant expression, affinity purification and functional characterization of Scots pine defensin 1. Appl Microbiol Biotechnol 89:1093–1101. https://doi.org/10.1007/s00253-010-2935-2
de Oliveira Carvalho A, Moreira Gomes V (2011) Plant defensins and defensin-like peptides - biological activities and biotechnological applications. Curr Pharm Design 17(38). Editoras Bentham Science. https://doi.org/10.2174/138161211798999447
Ermakova EA, Faizullin DA, Idiyatullin BZ, Khairutdinov BI, Mukhamedova LN, Tarasova NB, Toporkova YY, Osipova EV, Kovaleva V, Gogolev YV, Zuev YF, Nesmelova IV (2016) Structure of Scots pine defensin 1 by spectroscopic methods and computational modeling. Int J Biol Macromol 84:142–152. https://doi.org/10.1016/j.ijbiomac.2015.12.011
Ishaq N, Bilal M, Iqbal HMN (2019) Medicinal potentialities of plant defensins: A review with applied perspectives. Medicines 6:29. https://doi.org/10.3390/medicines6010029
Kovalera V, Bukhteeva I, Kit OY et al (2020) Plant Defensins from a structural perspective. Int J Mol Sci. https://doi.org/10.3390/ijms21155307
Wu J, Zhou X, Chen Q et al (2022) Defensins as a promising class of tick antimicrobial peptides: a scoping review. Infect Dis Poverty. https://doi.org/10.1186/s40249-022-00996-8
Deo S, Turton KL, Kainth T et al (2022) Strategies for improving antimicrobial peptide production. Biotechnol Adv 59:107968. https://doi.org/10.1016/j.biotechadv.2022.107968
Parisi K, McKenna JA, Lowe R, Harris KS, Shafee T, Guarino R, Lee E, van der Weerden NL, Bleackley MR, Anderson MA (2024) Hyperpolarisation of mitochondrial membranes is a critical component of the antifungal mechanism of the plant defensin, Ppdef1. J Fungos 10:54. https://doi.org/10.3390/jof10010054
Acknowledgements
Authors acknowledge Dr. Armand Segúin from Natural Resources Canada and the Canadian Forest Service for providing PgD1 gene.
Funding
Partial financial support was received from Grant PAP2023011000003 FAPESC No 48/2022.
Author information
Authors and Affiliations
Contributions
E. M.R. P.T cloned, expressed, and purified defensing.
L. C. C. wrote the manuscript and assisted with antifungal experiments.
G.F. D. S Wrote the manuscript, prepared the figures, and conceptualized the project.
M. J. G. isolated fungal species, wrote the manuscript and assisted with antifungal experiments.
R. C. isolated fungal species and revised the manuscript.
L. R. isolated fungal species and revised the manuscript.
M.L.B. M. Wrote the manuscript, prepared the figures and conceptualize the project.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Alaide Cristina de Bem Matos and Elisa Maria Rodriguez Pazinatto Telli contributed equally.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
de Bem Matos, A.C., Telli, E.M.R.P., Camillo, L.C. et al. Plant Defensin PgD1 a Biotechnological Alternative Against Plant Pathogens. Probiotics & Antimicro. Prot. (2024). https://doi.org/10.1007/s12602-024-10333-0
Accepted:
Published:
DOI: https://doi.org/10.1007/s12602-024-10333-0