
Abstract
An Upper Devonian to (?)Lower Mississippian section at Hushoot Shiveetiin gol in the Baruunhuurai Terrane of the Central Asian Orogenic Belt (CAOB) was investigated. A generally well-preserved and diverse ostracod fauna was collected from the Late Devonian (Famennian) Samnuuruul Formation in western Mongolia. It is the first rich assemblage described from this area. The ostracod fauna consists of 19 genera and 25 species. Two of the species are proposed new species, and 13 species are described in open nomenclature. The Mongolian ostracod fauna is similar to coeval faunas in western China and Laurussia, but also contains new, endemic species. Two new taxa are described: Beyrichiopsis hushootensis and Ampuloides beckeri. Ostracod findings are characterized by the Eifelian Mega-Assemblage (I-III) representing a nearshore, variable palaeoenvironment which is in accordance with facies analysis provided by other studies.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Reports on Palaeozoic faunas from Mongolia are sparse. Previous papers have described conodonts, corals and echinoderms (e.g. Bolshakova et al. 2003; Webster and Ariunchimeg 2004; Ariunchimeg et al. 2014; Suttner et al. 2019), but Palaeozoic ostracods from Mongolia are previously unreported. Ostracods can provide critical data on palaeoenvironmental and palaeogeographical reconstructions. Many ostracod species and genera from the Devonian exhibit characteristic assemblages ranging from shallow water to deep marine environments (e.g. Bandel and Becker 1975; Wang 1988; Becker and Bless 1990; Becker and Blumenstengel 1995; Becker 2000; Groos-Uffenorde et al. 2000; Casier 2004; Becker et al. 2004; Crasquin and Horne 2018).
In the framework of the International Geoscience Programme (IGCP) 596 and its successor, the Western Mongolia Working Group, our research group conducted fieldwork in western Mongolia in 2012, 2014 and 2018 (Kido et al. 2013; Ariunchimeg et al. 2014; Suttner et al. 2019). Herein, we describe a diverse, generally well-preserved ostracod fauna from the Hushoot Shiveetiin gol section of the Samnuuruul Formation (Famennian, Late Devonian), Baruunhuurai Terrane and Olonbulag Subterrane (Fig. 1). The fauna provides insight on palaeoenvironmental conditions of the section as well as on taxonomic affinities with other Famennian ostracod communities in the CAOB and elsewhere, particularly with Late Devonian ostracod communities of western Junggar, NW China (Song et al. 2017).

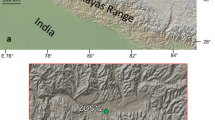
Geological map of the Hushoot Shiveetiin gol section, eastern part (A) and western part (B), based on new mapping data during field work in 2018
Geological setting
The Central Asian Orogenic Belt (CAOB) is the world’s largest Palaeozoic accretionary belt. Mongolia lies in the center of the CAOB and is composed of up to 44 different terrains, including cratonic, metamorphic, passive margin, island arc, accretionary complex and ophiolitic terranes, ranging in age from Proterozoic through the Phanerozoic (Badarch et al. 2002; Xiao et al. 2010; Metcalfe 2011; Choulet et al. 2012; Yang et al. 2013; Li et al. 2017). Studies by Windley et al. (2007), Donskaya et al. (2013) and Yang et al. (2015) suggest that subduction of the Palaeo-Asian Ocean continued during the Middle and Late Palaeozoic before final closure of this ocean and its Turkestan and Junggar branches during the Late Palaeozoic to the Mesozoic (Safonova et al. 2017). Palaeozoic rocks of western Mongolia described in this paper are part of the complex tectonic framework of the CAOB (for a more detailed discussion on plate tectonics, we refer to Safonova et al. 2017, cum lit.).
Deposits of the Baruunhuurai Terrane have a stratigraphic range from Devonian to Carboniferous and are located in the southwestern part of Mongolia close to the border to China. The investigated section is located in the Olonbulag Subterrane (Fig. 1) and ranges from the Devonian (Famennian) to Carboniferous (?early Mississippian). The exposed section belongs to the Samnuuruul Formation which has a thickness of 250–450 m. A detailed facies and stratigraphic analysis of the Hushoot Shiveetiin gol section is published by Ariuntogos et al. (2020, this issue). The ostracods reported herein were collected from both sections (A and B sections) and have a stratigraphical range based on conodonts from the crepida Biozone to the rugosa trachytera Biozone. The zonation scheme used herein follows Hartenfels (2011) and Spalletta et al. (2017).
The Samnuuruul Formation is dominated by siliciclastic rocks with numerous volcaniclastic deposits. Limestones are limited, occur as thin-bedded layers, and are rich in fossils, such as corals, conodonts, crinoids, trilobites, bryozoans and ostracods. The section is divided into an eastern (A) and a western (B) part which was correlated by a diagnostic thick-bedded pyroclastic marker horizon (Fig. 2). Correlation is also proven by conodont data (Ariuntogos et al. 2020, this issue). Sedimentary analysis indicates variable depositional facies settings ranging from shallow-intertidal to open marine (for detailed sedimentology and facies analysis see Ariuntogos et al. 2020, this issue).

Conodont biozones and ostracod levels of Famennian deposits at the Hushoot Shiveetiin gol section
Materials and methods
In this study, 12 samples contain ostracod specimens as single valves and carapaces. The limestone and marl samples between 1 and 4 kg were collected from the eastern (A) and western (B) Hushoot Shiveetiin gol section (see Suttner et al. 2019, Ariuntogos et al. 2020, this issue; Fig. 2). These samples were dissolved in formic and/or acetic acids and sieved into 63 μm, 125 μm, 250 μm and 500 μm fractions. The fractions were dried in an oven at about 40 °C and the most promising conodont-sized fractions were separated by liquid sodium polytungstate. After conodont separation, the light fractions were picked for ostracods and other microfossils. The ostracods were mounted on aluminium stubs, sputtered with gold using a Quorum (Q150R ES) and imaged using a Quanta 650 Field Emission scanning electron microscope in the Central Research Laboratory, Çukurova University, Turkey. The studied ostracods are deposited in the collections of the Mongolian University of Science and Technology, under the collection numbers MUST-RCSP-Ost.00001-98.
Ostracod data
Ostracods were identified in samples from the eastern (A) and western (B) Hushoot Shiveetiin gol section ranging from the crepida into the rugosa trachytera biozones in the Famennian (Ariuntogos et al. 2020, this issue). A systematic list of identified ostracod taxa (19 genera and 25 species) were given Appendix 1 and their stratigraphical distributions in the studied area is provided in Fig. 3. The ostracod fauna of the Hushoot Shiveetiin gol section includes species belonging to the orders Palaeocopida, Platycopida and Podocopida (Figs. 3, 4, 5, 6, 7, and 8). In this study the Palaeozoic ostracod classification by Becker (2002) and Liebau (2005) was used. Ten species out of a total number of 25 species belong to previously known taxa. Two new species are proposed, and because of the materials are insufficient for formal description and naming of a new taxon, thirteen species are left in open nomenclature.

Stratigraphical distribution of ostracods according to conodont biozones in the Hushoot Shiveetiin gol section

Ostracods from the Late Devonian Hushoot Shiveetiin gol section (Baruunhuurai Terrane, Mongolia). a–d Hollinella (Keslingella) lionica Becker and Bless 1971. a right valve, MUST-RCSP-Ost.00042), b left valve lateral views (MUST-RCSP-Ost.00043), c right valve (MUST-RCSP-Ost.00075), d right valve, internal view (MUST-RCSP-Ost.00076). e–h Hollinella (Keslingella) aff. praecursor Pokorny, 1950. e right valve (MUST-RCSP-Ost.00006), f left valve lateral views (MUST-RCSP-Ost.00008), g right valve (MUST-RCSP-Ost.00007), h left valve lateral views (MUST-RCSP-Ost.00074). i Aechmina sp., dorsal view (MUST-RCSP-Ost.00092). j Parabolbina sp., right valve, external view (MUST-RCSP-Ost.00084). k–n Pribylites junggarensis Song and Crasquin, 2017 in Song et al. 2017, k right valve lateral view (MUST-RCSP-Ost.00047), l left lateral view of carapace (MUST-RCSP-Ost.00050), m right valve lateral views (MUST-RCSP-Ost.00060), n right lateral view of carapace (MUST-RCSP-Ost.00073). o Coryellina sp., right lateral view of carapace (MUST-RCSP-Ost.00086)

Ostracods from the Late Devonian Hushoot Shiveetiin gol section (Baruunhuurai Terrane, Mongolia). a Amphissites centrocostatus Blumenstengel, 1969, left valve lateral view (MUST-RCSP-Ost.00007). b–e Amphissites irinae Glebovskaja and Zaspelova in Egorov, 1953, b left valve lateral view (MUST-RCSP-Ost.000), c left valve lateral view (MUST-RCSP-Ost.00071), d left valve lateral view (MUST-RCSP-Ost.00013), e left valve lateral view (MUST-RCSP-Ost.00019). f Polytylites sp., right valve lateral view (MUST-RCSP-Ost.00012). g-i Nezamyslia sp., g right valve lateral view (MUST-RCSP-Ost.00076); h. left valve lateral view (MUST-RCSP-Ost.00006), i left valve lateral view (MUST-RCSP-Ost.00008). j-k Berdanella sp., j right valves lateral view (MUST-RCSP-Ost.00059), k right valves lateral view (MUST-RCSP-Ost.00045). l–n Palaeocopida? indet. 1. l left valve lateral view (MUST-RCSP-Ost.00042), m right valve lateral view (MUST-RCSP-Ost.00043), n left valve lateral view (MUST-RCSP-Ost.00075). o Palaecopida? indet. 2. right lateral view of male carapace (MUST-RCSP-Ost.00062)

Ostracods from the Late Devonian Hushoot Shiveetiin gol section (Baruunhuurai Terrane, Mongolia). a–f Palaecopida? indet. 2. a right lateral view of male carapace (MUST-RCSP-Ost.00063), b left lateral view of male carapace (MUST-RCSP-Ost.00064), c left lateral view of female carapace (MUST-RCSP-Ost.00065), d right lateral view of female carapace (MUST-RCSP-Ost.00085), e left lateral view of female carapace (MUST-RCSP-Ost.00087), f left lateral view of male carapace (MUST-RCSP-Ost.00090). g-h Kloedenellidae indet, HS-C-2, g Left valve lateral view (MUST-RCSP-Ost.00030), h right valve lateral view (MUST-RCSP-Ost.00083). i Hypotetragona tremula Becker, 1971, (MUST-RCSP-Ost.00054), right valve lateral view. j-o Beyrichiopsis hushootensis sp. nov., j dorsal view of carapace (MUST-RCSP-Ost.00079), k left valve internal view (MUST-RCSP-Ost.00077), l right valve internal view (MUST-RCSP-Ost.00078), m right valve lateral view of Paratype A (MUSlT-RCSP-Ost.00038), n left valve lateral view (MUST-RCSP-Ost.00037), o right valve lateral view of Holotype (MUST-RCSP-Ost.00041)

Ostracods from the Late Devonian Hushoot Shiveetiin gol section (Baruunhuurai Terrane, Mongolia). a Beyrichiopsis hushootensis sp. nov., a left valve lateral view of Paratype B (MUST-RCSP-Ost.00040). b Bairdia hypsela Rome 1971, right lateral view of carapace (MUST-RCSP-Ost.00025). c Bohlenatia sp., right valve lateral view (MUST-RCSP-Ost.00068). d Pustulobairdia sp., right valve lateral view (MUST-RCSP-Ost.00023). e Baschkirina sp., right lateral view of carapace (MUST-RCSP-Ost.00061). f–g Bairdiocypris sp., f right lateral view of carapace (MUST-RCSP-Ost.00001). g right lateral view of carapace (MUST-RCSP-Ost.00002). h–j Microcheilinella bulongourensis Song and Crasquin, 2017, h left lateral view of carapace (MUST-RCSP-Ost.00093), i left lateral view of carapace (MUST-RCSP-Ost.00094), j left lateral view of carapace (MUST-RCSP-Ost.00070). k–o Ampuloides beckeri sp. nov., k left valve lateral view of Paratype A (MUST-RCSP-Ost.00069), l left valve lateral view of Holotype (MUST-RCSP-Ost.00031), m left valve lateral view of Paratype B (MUST-RCSP-Ost.00035), n left valve lateral view of Paratype C (MUST-RCSP-Ost.00036), o left valve internal view (MUST-RCSP-Ost.00095)

Ostracods from the Late Devonian Hushoot Shiveetiin gol section (Baruunhuurai Terrane, Mongolia). a–c Cribroconcha primaris Kotschetkova, 1980, a left valve lateral view (MUST-RCSP-Ost.00021), b right valve lateral view (MUST-RCSP-Ost.00072), c left valve lateral view (MUST-RCSP-Ost.00080). d–f Cribroconcha honggulelengensis Song and Crasquin, 2017 in Song et al. 2017, d left valve lateral view (MUST-RCSP-Ost.00033), e left valve lateral views (MUST-RCSP-Ost.00081), f. Left lateral view of carapace (MUST-RCSP-Ost.00089)
Systematic Palaeontology
The new species and open nomenclature are given in this part.
Class Ostracoda Latreille, 1802
Order Palaeocopida Henningsmoen, 1953
Superfamily Aechminacea Bouček, 1936.
Family Aechminidae Bouček, 1936.
Aechmina sp. (Fig. 4/i)
Material: Two carapaces, partly damaged
Occurrence. It was found in gracilis gracilis Biozone of Hushoot Shiveetiin gol section (Unit IV).
Superfamily Hollinoidea (=Hollinacea) Swartz 1936
Family Ctenoloculinidae Jaanusson and Martinsson, 1956
Parabolbina sp. (Fig. 4/j)
Material: Three valves, partly damaged.
Remarks: Because of the limited number of carapaces and valves, Parabolbina sp. is described in open nomenclature.
Occurrence: It was found in gracilis gracilis Biozone of Hushoot Shiveetiin gol section (Unit IV).
Superfamily Primitiopsoidea (=Primitiopsacea) Swartz 1936
Family Pribylitidae Pokorný, 1958
Coryellina sp. (Fig. 4/o)
Material: One valve.
Remarks: Coryellina sp. is poorly preserved and does not allow a precise identification.
Superfamily Kirkbyoidea (=Kirkbyacea Ulrich and Bassler 1906)
Family Amphissitidae Knight, 1928
Polytylites sp. (Fig. 5/f)
Material: Seven valves, partly damage.
Remarks: Some morphological features on valves (a rounded lobe occurs in the median part, surface reticulate) are observed. But, they are not detailed enough for formal description and naming of a new species.
Occurrence: Polytylites sp. was found in the rhomboidea Biozone of Hushoot Shiveetiin gol section, sample HS-C-1b (Unit IV).
Family Nezamysliidae Żbikowska 1983
Nezamyslia sp. (Fig. 5/g-i)
Material: Three valves, all partly damage.
Remarks: The material is poorly-preserved, thus this species cannot be determined exactly and attributed to a known or new species.
Occurrence: It is observed in the rhomboidea Biozone.
Family Kirkbyellidae Sohn, 196
Berdanella sp. (Fig. 5/j-k)
Material: 4 valves
Description: Outline subquadrate in lateral view, dorsal border straight. Median sulcus (S2) distinct. A lateral ridge with a small posteromedian lobe is on ventral half of the valve. Surface fine reticulate.
Remarks: In outline and surface fine reticulate, the specimens are close to Kirkbyella (Berdanella) unicornis (Coryell and Malkin), 1936, but differ by a lateral ridge with a small posteromedian lobe.
Dimension: L = 1.023–1.433 mm, H = 0.600–0.783 mm.
Occurrence: Berdanella sp. was found in rugosa trachytera Biozone of the Hushoot Shiveetiin gol section, sample HS-C-W1 (Unit VII).
Palaeocopina? indet. 1 (Fig. 5/l-n)
Material: Thirteen valves.
Dimensions: L = 0.600–0.690 mm, H = 0.400–0.480 mm.
Description: Carapace small, nonsulcate, amplete, straigth and long dorsal margin, cardinal angles well defined, obtuse, adventral structure developed and running almost parallel to the free margins, adductorial pit is not clear. The rows of reticulations are observed. The perpendicular ridges are dominated and subparallel to the free margin.
Remarks: The form described herein correlates with the lateral outline and ornamentation of carapace to Reticestus sp. Olempska, 1979 and Reticestus? n.sp. Zagora 1968, except anterodorsal and posterodorsal part and general shape.
Occurrence: The rugosa trachytera Biozone of the Hushoot Shiveetiin gol section, HS-C-W1 (Unit VII).
Palaeocopina? indet. 2 (Fig. 5/o, Fig. 6/a-f).
Material: 33 corroded carapaces.
Dimensions: L = 0.538–0.728 mm, H = 0.400–0.480 mm.
Description: The carapaces and valves are poorly preserved. There is a denticulate dorsal border. Indistinct shallow sulcus (S2). Sexual dimorphism presumably present. The carapace outline of male and female is different. Anterior and posterior margins of females are rounded. Male carapace is preplete (Fig. 5/o, 6/a,b,f). Female carapace is amplete (Fig. 6/c–e).
Occurrence: It occurs from the rhomboidea to the gracilis gracilis zones of the Hushoot Shiveetiin gol section (Unit IV).
Order Platycopida Sars 1866
Superfamily Kloedenelloidea (=Kloedenellacea Ulrich and Bassler 1908)
Kloedenellidae indet. (Fig. 6/g-h)
Material: Two valve
Dimension: (one specimen measured). L = 1.025 mm, H = 0.575 mm.
Occurrence: HS-C-2 includes undetermined kloedenellid ostracod in the rhomboidea Biozone of the eastern section (A) Hushoot Shiveetiin gol section.
Family Beyrichiopsidae Henningsmoen, 1953
Genus Beyrichiopsis Jones and Kirkby 1886
Type-species: Beyrichiopsis fimbriata Jones and Kirkby 1886
Beyrichiopsis hushootensis sp. nov. (Fig. 6/j-o, Fig. 7/a)
Holotype: Right valve, (Fig. 6/o, MUST-RCSP-Ost.00041)
Paratype: Right valve of paratype A (Fig. 6/m, MUST-RCSP-Ost.00038), left valve of paratype B (Fig. 7/a, MUST-RCSP-Ost.00040).
Derivation of name: The name is derived from Hushoot Shiveetiin gol section, Baruunkhuurai Terrane, Mongolia.
Type locality and horizon: The rugosa trachytera Biozone of the Hushoot Shiveetiin gol section, sample HS-C-W1 (Unit VII).
Material: Fifty-six valves.
Diagnosis: Subquadrate and subovate, comma-shaped crests, S2 short.
Dimensions: Holotype: L = 0.980 mm, H = 0.600 mm; Paratypes: L = 0.940–1.020 mm, H = 0.560–0.600; other specimens: L = 0.785–1.080 mm, H = 0.446–0.600 mm.
Description: Outline subquadrate-elongate in lateral view, right valve larger than left valve; hinge line straight; cardinal angle obtuse; median node (L2) prominent; adductorial sulcus (S2) prominent as narrow and short; conspicuous marginal frill; comma-shaped crest on valve surface; marginal ridge present; surface smooth.
Discussion: Beyrichiopsis hushootensis sp. nov. may be compared with Beyrichiopsis glyptopleuroides Green 1963 in Becker et al. 1974. The new species differs in having a comma-shaped lateral crest and smooth ornamentation.
Order Podocopida Sars 1866
Superfamily Bairdioidea (=Bairdiacea Sars 1887)
Family Bairdiidae Sars, 1888
Bohlenatia sp. (Fig. 7/c)
Material: One valve
Dimension: L = 0.600 mm, H = 0.188 mm
Occurrence: It was found in the gracilis gracilis Biozone of the section Hushoot Shiveetiin gol section.
Pustulobairdia sp. (Fig. 7/d)
Material: One valve
Dimension: L = 0.600 mm, H = 0.300 mm.
Occurrence: It was observed in the crepida Biozone of Hushoot Shiveetiin gol section.
Superfamily Bairdiocypridoidea (=Bairdiocypridacea Shaver 1961)
Family Bairdiocyprididae Shaver 1961
Baschkirina sp. (Fig. 7/e)
Material: One carapace
Dimension: L = 0.616, H = 0.350 mm
Occurrence: It was found in the rugosa trachytera Biozone of the Hushoot Shiveetiin gol section.
Bairdiocypris sp. (Fig. 7/f-g)
Material: Five carapaces
Dimension: L = 0.925–0.616 mm, H = 0.550–0.350 mm
Occurrence: It occurs from the crepida to gracilis gracilis biozones of Hushoot Shiveetiin gol section.
Superfamily Healdioidea (=Healdiacea Harlton 1933)
Family Pachydomellidae Berdan and Sohn, 1961
Genus Ampuloides Polenova 1952
Type-species: Ampuloides verrucosa Polenova 1952
Ampuloides beckeri sp. nov. (Fig. 7/k-o)
Holotype: Left valve (Fig. 7/l, MUST-RCSP-Ost.00031)
Paratype: Left valve of Paratype A (Fig./k, MUST-RCSP-Ost.00069), left valve lateral Paratype B (Fig. 7/m, MUST-RCSP-Ost.00035), left valve of Paratype C (Fig. 7/n, MUST-RCSP-Ost.00036)
Derivation of name: Dedicated to German ostracodologist Prof. Dr. Gerhard Becker.
Type locality and horizon: The rhomboidea Biozone of the Hushoot Shiveetiin gol section, sample HS-C-2 (Unit IV).
Material: Twenty-one valves.
Diagnosis: Carapace very swollen, inflated in dorsal view, a narrow groove, cardinal extremity, carapace surface warty (small tubercles).
Dimensions: Holotype: L = 0.690 mm, H = 0.430 mm; Paratypes: L = 0.527–0.700 mm, H = 0.310–0.377.
Description: The lateral outline of the carapace is preplete, valves strongly asymmetrical and inflated, a narrow groove runs parallel to the front edge in the lateral view of valve, the most swollen part of the carapace located in the posterior half, left valve overlaps the right one, cardinal extremity in anterodorsal part of left valve, depressing straight hinge margin, the bow-shaped projection occurs in the ventral margin of the left valve, the surface is covered with small rounded tubercles (warty). The tubercles are only visible in well-preserved specimens.
Discussion. Ampuloides beckeri sp. nov. differs from Ampuloides pumilus Olempska, 1979 (Famennian of the Holy Cross Mountains, Poland) by the absence of a nodular process occuring in the anterodorsal part and shape/outline of the carapace. Also, Ampuloides beckeri sp. nov. differs from Ampuloides kellerwaldensis Casier and Lethiers, 1999 (Schmidt Quarry, Frasnian/Famennian section in the Kellerwald, Germany) by the presence of an anterior furrow and by the smaller development of cardinal extremities.
Palaeoenvironmental analysis
Ostracods can provide critical data for reconstructing Palaeozoic environments, especially in the Devonian. Ostracods can be used to distinguish ecologically three mega-assemblages (ecotype), such as the Eifelian (thick-shelled and sculptured ostracod assemblages), Thuringian (thin-shelled species with smooth and spinose valves) and Myodocopid (=Entomozoacean) mega-assemblages (ecotype) (finger-print ostracods) (e.g. Bandel and Becker 1975; Groos-Uffenorde et al. 2000; Becker 2000; Becker et al. 2004; Casier 2004, 2017; Crasquin and Horne 2018). Also, in-depth overall information and references are given and discussed on palaeoecology of ostracods by Casier (2017). Three mega-assemblages and six assemblages of ostracods numbered 0 to V indicative for lagoonal-, semi-restricted- and marine environments above and below fair weather wave-base or storm wave-base in the Devonian are described by Casier (2017). Wang (1988) proposed five ostracod associations (leperditiid, palaeocopid, smooth-podocopid, spinose-podocopid and entomozoacean) ranging from nearshore to deep basins in Palaeozoic strata of South China.
The Devonian ostracod genera and species from the Hushoot Shiveetiin gol section (Baruunhuurai Terrane) are characterized by palaecopids, platycopids and podocopids (Fig. 9). Calcareous levels (Unit II, IV, V and VII) from the section yielded many ostracod specimens even if they lived in a high physical stress environment characterized by frequent and often voluminous pyroclastic eruptions (Ariuntogos et al. 2020, this issue). However, some genera and species are particularly useful for palaeoenvironmental interpretation. Critical assemblages in this study are characterized by the smooth-shelled and ornamented-shell (reticulate, spinous and node-bearing) ostracods.

Distribution and abundance of the palaecopid, platycopid and podocopid ostracods from the Hushoot Shiveetiin gol section
Hollinoideans lived in marine environments and lived worldwide in nektobentonic, epineritic and pelagic facies realms. They occur together with kloedenellids and eridostracans (Becker 1980, 1982, 2002) and are indicative of more restricted environments. Hypotetragona occurs in marine near-shore environments (Bless 1983). Also, weakly ornamented ostracods such as the Knoxiella-Beyrichiopsis assemblage represent a supratidal environment in the late Famennian in the Dinantian Basin of Belgium (Becker et al. 1974). Primitiopsoidean occur in epineritic as well as in pelagic facies realms (Becker 2002; Adamczak 2005). Kirkbyoidean were marine ostracods ranging from near-shore (e.g. found in shallow-water sections in the Eifel area, Germany) to deep-sea (such as in Thuringia, Germany; see Bandel and Becker 1975; Olempska 1999). The studies on fossil and modern bairdiids indicate that they occur from near-shore to bathyal environments of normal marine salinity (Melnyk and Maddocks 1988). Thin-shelled, spinose bairdiids indicating Thuringian mega-assemblage are absent in the samples from the Hushoot Shiveetiin gol section.
Common short spiny podocopid (Pustulobairdia, Ampuloides) association with kirkbyoid (Amphissites) forms are found in Unit II (sample HS-C-11a, Figs. 2 and 3). The thick-shelled and sculptured palaeocopids (Hollinella, Pribylites, Nesamyslia and Amphissites) and smooth to weak ornamented podocopids (Bairdia, Ampuloides and Cribroconcha) are very common in the upper part of Unit IV (sample number HS-C-2, Figs. 2, 3, 9). Paleocopids are dominant in the other parts of Unit IV. Podocopids are also observed in the upper part of Unit IV (sample number HS-C-W15). Relatively increases in the number of ostracod genera and species point to fully marine conditions which correspond with limestone or slightly silicified limestone. But, the ostracods are represented by a few species and genera in the upper parts of Unit IV (sample number HS-C-W14) and Unit V (sample number HS-C-W12). A very diverse assemblage containing palaecopid + platycopid + podocopid ostracods were observed at the end of Unit VII (sample HS-C-W1, Figs. 2, 3, 9). This assemblage was found in short-lived pure limestone and might be related to a sea-level rise. However, the Hushoot Shiveetin gol section is characterized by a very variable facies setting ranging from lagoonal, shallow-intertidal to open marine. Hydrodynamic conditions are also very variable ranging from high-energy environment to semi-restricted conditions and a number of remarkable sea-level changes (Ariuntogos et al. 2020, this issue).
Overall, the ostracod assemblage of the Hushoot Shiveetiin gol section is considered to be ecologically equivalent to the Eifelian mega-assemblage which include semi-restricted (Ass. I), marine above (Ass. II) or below (Ass. III) fair-weather wave-base or storm wave-base environments. These assemblages correspond to foreshore Ostracod Assemblage-1 (OA1), nearshore Ostracod Assemblage-2 (OA2) and offshore Ostracod Assemblage-23 (OA3) environments of the Hongguleleng Formation (western Junggar, NW China) in an open oceanic island arc (Song et al. 2019). Also, these results can be correlated with the Eifelian mega-assemblage indicative of a nearshore-offshore setting (palaeocopid and smooth-podocopid associations) in the Gelaohe Formation from the Devonian-Carboniferous (D/C) transition, South China (Song and Gong 2019).
The Eifelian mega-assemblage is characterized by variable environments which are confirmed by sedimentological criteria and facies analysis (Ariuntogos et al. 2020, this issue).
Palaeobiogeographical relationships
The Hushoot Shiveetiin gol section (Baruunhuurai Terrane) belongs to an active island arc complex in the CAOB. Ostracod assemblages of this section consist of cosmopolitan, regional and endemic, new species. Some of the identified ostracods are similar to coeval faunas reported from studies in Russia, China, Poland, Belgium, Germany, Spain, Czech and Turkey and references are given below (Fig. 10).

Geographical and stratigraphical distributions of ostracods of the Hushoot Shiveetiin gol section (see references in section 5). □ Eifelian, * Givetian, x Frasnian, o Famennian, ● Tournaisian, ■ Upper Devonian
Hollinella (Keslingella) aff. praecursor occurs in the rhomboidea Biozone of the eastern and western parts of the Hushoot Shiveetiin gol section. This species is known from the Late Devonian in Spain (Iberian Chains, Gozalo 1994), from the Middle Devonian from Red Coral Limestone in the Czech Republic (Late Givetian, Pokorny 1950), the Middle Devonian from the Eifel area, Germany (Ahrdorf Beds, Eifelian, Bless and Jordan 1971), from Late Devonian in Belgium (Frasnian, Dinant Basin, Casier and Olempska 2008a), and from the Late Devonian from Turkey (Famennian in Taurids, E. Şeker Zor, personal communication, October 14, 2019).
Hollinella (Keslingella) lionica occurs in the gracilis gracilis and rugosa trachytera biozones of the western part. It is known from the Late Devonian in France (Frasnian of Ardennes and Boulonnais, Lethiers 1984), the Dinant Basin in Belgium (Becker 1971; Casier and Olempska 2008a, b), and the Late Devonian in Turkey (Famennian in the Taurids; E. Şeker Zor, personal communication, October 14, 2019).
Amphissites irinae occurs in both, the eastern and western sections within crepida and rhomboidea biozones. This species is known from the Late Devonian (Frasnian) of Russia (Egorov 1953; Zaspelova 1959; Evdokimova 2015; Zhuravlev et al. 2006), Poland (Zbikowska 1983), Germany (Harz Mountains, Blumenstengel 1969), and Spain (Famennian of Cantabrian Mountains, Becker 1977).
Amphissites centrocostatus occurs from the rhomboidea to rugosa trachytera biozones of the western section. This species is known from the Late Devonian in Germany (Harz Mountains, Frasnian, Blumenstengel 1969), Spain (Famennian, Central Pyrenees; Sanchez de Posada et al. 2008), and from the Pontids in Turkey (Nazik et al. 2012).
Hypotetragona tremula is known from the Late Devonian (Frasnian) of Dinant Basin, Belgium (Becker 1971; Becker and Bless 1974 in Becker et al. 1974; Casier and Olempska 2008a, b).
Bairdia hypsela occurs from the crepida to the gracilis gracilis biozones of both sections. It is known from Upper Devonian-Carboniferous strata from the Holy Cross Mountains, Poland, from the Dinant Basin in France and Belgium, as well as from Germany (Rhenish Massif and Thuringia), from Turkey (Taurids and Pontids), and from Morocco and northern Algeria (Olempska 1979; Becker et al. 1993; Casier et al. 2005; Nazik et al. 2012).
Three new species (Microcheilinella bulongourensis, Pribylites junggarensis and Cribroconcha honggulelengensis) which were identified in the Late Devonian of western Junggar, NW China by Song et al. (2017) were also found in Hushoot Shiveetiin gol section. Cribroconcha primaris occurs in the rhomboidea Biozone of the eastern section. It is known from Tournaisian from the Southern Urals (Kotschetkova 1980), the Devonian/Carboniferous boundary beds of Kozhim, Polar Urals (Nemirovskaya et al. 1992) and of South Urals (Kulagina 2013).
Furthermore, the new species (Beyrichiopsis hushootensis, Ampuloides beckeri) and species described in open nomenclature (Palaeocopina? indet. 1 and 2, Nezamyslia sp., Polytylites sp., Parabolbina sp., Coryellina sp., Berdanella sp., Pustulobairdia sp., Aechmina sp., Bairdiocypris sp., Baschkirina sp., Bohlenatia sp. and Kloedenellidae indet.) are considered as endemic species. These data may support the hypothesis that the CAOB was a diversity hotspot during the Famennian also for ostracods beside other organisms (see Waters et al. 2020, this issue). As a result, the studied ostracod fauna consist of Euramerican/Laurusian assemblages (Hollinella (Keslingella) lionica, Hollinella (Keslingella) aff. praecursor, Amphissites irinae, Amphissites centrocostatus, Hypotetragona tremula and Bairdia hypsela), China (Microcheilinella bulongourensis, Pribylites junggarensis and Cribroconcha honggulelengensis) and endemics.
Conclusions
In total, 25 ostracod species belonging to 19 genera from the Late Devonian (Famennian) Hushoot Shiveetiin gol section (Mongolia) are documented and figured. They were found in sediments ranging from the crepida to rugosa trachytera biozones. The Mongolian ostracod fauna is similar to coeval faunas known from Laurussia but also from China. Two new taxa are described. Ostracod assemblages are ecologically equivalent to the Eifelian mega-assemblage (I-III) and generally represent nearshore, variable palaeoenvironment. The diverse ostracod Eifelian mega-assemblage expanding from shallow-water refugia supports the idea of the CAOB as a biodiversity hotspot in the aftermath of the F/F extinction.
References
Adamczak, F. J. (2005). [Edited by G. Becker]: Contributions to Palaeozoic Ostracod Classification [POC], No. 37. The Superfamily Primitiopsacea Swartz, 1936. Family Primitiopsidae Swartz, 1936. Mid-Palaeozoic genera. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte, 17–45.
Ariunchimeg, Ya., Bayasgalan, A., Waters, J.A., Kido, E., Suttner, T. J., Sersmaa, G., Undariya, J., Otgonbaatar, D., (2014). IGCP 596 and IGCP 580 field workshop guidebook. 8-18th August, 2014, Ulaanbaatar, Mongolia. Paleontological Center, Mongolian Academy of Sciences, pp. 1–55.
Ariuntogos, M., Königshof, P., Hartenfels, S., Jansen, U., Nazik, A., Carmichael, S.K., Waters, J.A., Gonchigdorj, J., Cronier, C., Ariunchimeg, Ya, Paschall, O., Dombrowski, A. (2020). The Hushoot Shiveetiin gol section (Baruunhuurai Terrane): Sedimentology and facies from a Late Devonian island arc setting. Palaeobiodiversity and Palaeoenvironments. (this issue).
Badarch, G., Cunningham, W. D., & Windley, B. F. (2002). A new terrane subdivision for Mongolia: implications for the Phanerozoic crustal growth of central Asia. Journal of Asian Earth Sciences, 21, 87–104.
Bandel, K., & Becker, G. (1975). Ostracoden aus Paläozoischen pelagischen Kalken der Karnischen Alpen (Silurium bis Unterkarbon). Senckenbergiana Lethaea, 56, 1–84.
Becker, G. (1971). Paleoecology of Middle Devonian ostracods from Eifel region, Germany. Paléoécologie des Ostracodes. Bulletin du Centre Recherches Pau-SNPA, 5, 801–816.
Becker G. (1977). Thuringian ostracods from the Famennian of the Cantabrian Mountains. In: H. Löffler & D. Danielopol (Eds.), Aspects of ecology and zoogeography of recent and fossil Ostracoda (pp. 459–474). Kluwer Academic Publishers.
Becker, G. (1980). Givetische Ostracoden aus der Bohrung Boussu in Belgien. Mémoires pour servir à l'explication des Cartes géologiques et minières de la Belgique, 20, 1–17.
Becker, G. (1982). Ostracoden aus Cephalopoden-führendem Oberdevon im Kantabrischen Gebirge (N-Spanien). 2. Bairdiacea, Cytheracea und Entomozoacea. Palaeontographica (A), 178(4–6), 109–182.
Becker, G. (2000). Progress in mid Palaeozoic palaeoceanographical studies from Ostracoda – from local to global importance (a review). Senckenbergiana Lethaea, 80(2), 555–566.
Becker, G. (2002). Contributions to Palaeozoic ostracod classification [poc], no. 24. Palaeozoic Ostracoda. The standard classification scheme. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 226, 165–228.
Becker, G., & Bless, M. J. M. (1971). Zur Verbreitung der Ostracoden-Familie Hollinellidae Bless and Jordan. Mit Beschreibung neuer Funde aus dem Mittel- und Oberdevon Westeuropas. Senckenbergiana lethaea, 52, 537–567.
Becker, G., & Bless, M. J. M. (1974). Ostracode stratigraphy of the Ardenno-Rhenish Devonian and Dinantian (with special reference to the Eifelian/Givetian boundary in the Eifel region and to the Famennian/Tournaisian faunas in the Dinant Basin). International Symposium on Belgian Micropaleontological Limits from Emsian to Visean, Namur, 1, 1–52.
Becker, G., & Bless, M. J. M. (1990). Biotope indicative features in Palaeozoic ostracods: a global phenomenon. In R. Whatley & C. Maybury (Eds.), Ostracoda and Global Events. British Micropalaeontological Societv Publication Series (pp. 421–436). London, New York: Chapman & Hall.
Becker, G., & Blumenstengel, H. (1995). Ostracoden vom Thüringer Ökotyp aus der “Postriff-Kappe” des Rübeländer Riffs (Elbingeröder Komplex, Harz; Obere crepida-Zone, Oberdevon). Abhandlungen und Berichte für Naturkunde, 18, 63–101.
Becker, G., Bless, M. J. M., Streel, M., & Thorez, J. (1974). Palynology and ostracode distribution in the Upper Devonian and basal Dinantian of Belgium and their dependence on sedimentary facies. Mededelingen Rijks Geologische Dienst, 25, 9–99.
Becker, G., Clausen, C.-D., & Leuteritz, K. (1993). Verkieselte Ostracoden vom Thüringer Ökotyp aus dem Grenzbereich Devon/Karbon des Steinbruchs Drewer (Rheinisches Schiefergebirge). Courier Forschunginstitut Senckenberg, 160, l–l30.
Becker, G., Lazreq, N., & Weddige, K. (2004). Ostracods of Thuringian provenance from the Devonian of Morocco (Lower Emsian–Middle Givetian; south-western anti-Atlas). Palaeontographica A, 271, 1–109.
Berdan, J., & Sohn, I.G., (1961). Family Pachydomellidae Berdan & Sohn, n.fam. In R.C. Moore (ed.) Treatise on Invertebrate Paleontology (pp. Q.373–Q.374). Part Q, Arthropoda 3, Crustacea, Ostracoda.
Bless, M. J. M. (1983). Late Devonian and Carboniferous ostracode assemblages and their relationship to the depositional environment. Bulletin de la Société Belge de Géologie, 92, 31–52.
Bless, M. J. M., & Jordan, H. (1972). Ostracodes of the family Hollinellidae. Mededelingen Rijks Geologische Dienst Serie C, 3(1), 1–155.
Blumenstengel, H. (1969). Oberdevonische Ostracoden aus der Bohrung Mandelholz 18/56 (Harz, Elbingeroder Komplex). Freiberger Forschungshefte C, 256, 7–36.
Bolshakova, L. N., Bondarenko, O. B., Minjin, C., Ulitina, L. M., & Sharkova, T. T. (2003). Paleontology of Mongolia: corals and stromatoporoids (pp. 199–256). Moskow: Nauka (in Russian).
Casier, J.-G. (2004). The mode of life of Devonian entomozoacean ostracods and the myodocopid mega-assemblage proxy for hypoxic events. Bulletin de l’Institut Royal des Sciences Naturelles de Belgique Sciences de la Terre, 74, 73–80.
Casier, J.-G. (2017). Ecology of Devonian ostracods: application to the Frasnian/Famennian boundary bioevent in the type region (Dinant Synclinorium, Belgium). Palaeobiodiversity and Palaeoenvironments, 97, 553–564.
Casier, J.-G., & Olempska, E. (2008a). Early Frasnian ostracods from the Arche quarry (Dinant Synclinorium, Belgium) and the Palmatolepis punctata isotopic event. Acta Palaeontologica Polonica, 53(4), 635–646.
Casier, J.-G., & Olempska, E. (2008b). Middle Frasnian (Devonian) ostracods from the Frasnes railway section (Dinant Synclinorium, Belgium); taxonomy, biostratigraphy, paleoeeology. In E. Steurbaut, J.W.M. Jagt, & E.A. jagt-Yazykova. (Eds.), Annie V. Dhondt memorial volume. Bulletin de l'Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre, 78, 51–66.
Casier, J.-G., Lebon, A., Mamet, B., & Préat, A. (2005). Ostracods and lithofacies close to the Devonian-Carboniferous boundary in the Chanxhe and Rivage sections, northeastern part of the Dinant Basin, Belgium. Bulletin de l'Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre, 75, 95–126.
Choulet, F., Cluzel, D., Faure, M., Lin, W., Wang, B., Chen, Y., Wu, F. Y., & Ji, W. (2012). New constraints on the pre-Permian continental crust growth of Central Asia (West Junggar, China) by U-Pb and Hf isotopic data from detrital zircon. Terra Nova, 24, 189–198. https://doi.org/10.1111/j.1365-3121.2011.01052.x.
Crasquin, S., & Horne, D. J. (2018). The palaeopsychrosphere in the Devonian. Lethaia, 51, 547–563.
Donskaya, T. V., Gladkochub, D. P., & Mazukabzov, A. M. (2013). Late Paleozoic-Mesozoic subduction-related magmatism at the southern margin of the Siberian continent and the 150 million-year history of the Mongol-Okhotsk Ocean. Journal of Asian Earth Sciences, 62, 79–97. https://doi.org/10.1016/j.jseaes.2012.07.023.
Egorov, V.G. (1953). Ostrakody franskogo yarusa russkoy platformy. 2. Bairdiidae, Hollinidae, Kirkbyidae. Ostracoda from the Frasnian stage of the Russian platform-2: Bairdiidae, Hollinidae, Kirkbyidae Trudy VNIGRI, Moskov Filial, 1-133 (in Russian).
Evdokimova, I.O. (2015). New data on the Frasnian ostracods from the middle Timan region, Russia: taxonomy, biostratigraphy, palaeoecology. In B. Mottequin, J. Delayer, P. Königshof, C. Perestianni & S. Olivie (Eds). Strata, Travaux de Géologie sédimentaire et Paléontologie, Série 1: communications, 16: 48-49.
Gozalo, R. (1994). Geologia y Paleontologỉa (Ostrácodos) del Devónica superior de Tabuenca (NE de la Cadena Ibérica oriental). Memorias del Museo Paleontológico de la Universidad de Zaragoza, 6, 1–291.
Green, R. (1963). Lower Mississippian ostracodes from the Banff formation, Alberta. Research Council of Alberta Bulletin, 11, 1–201.
Groos-Uffenorde, H., Lethiers, F., & Blumenstengel, H. (2000). Ostracodes and Devonian stratigraphy. Courier Forschunginstitut Senckenberg, 220, 99–111.
Harlton, B. H. (1933). Micropaleontology of the Pennsylvanian Johns Valley shale of the Ouachita mountains, Oklahoma, and its relationship to the Mississippian Caney shale. Journal of Paleontology, 7(1), 3–29.
Hartenfels, S. (2011). Die global Annulata-Events und die Dasberg-Krise (Famennium, Oberdevon) in Europa und Nord-Afrika – hochauflösende Conodonten-Stratigraphie, Karbonat-Mikrofazies, Paläoökologie und Paläodiversität. Münsterische Forschungen zur Geologie und Paläontologie, 105, 17–527.
Henningsmoen, G. (1953). Classification of Paleozoic straight-hinged ostracods. Norsk Geologisk Tidsskrift, 31, 185–288.
Jones, T.R., & Kirkby, J.W. (1886). Notes on the Palaeozoic bivalved Entomostraca, no. 22. On some undescribed species of British carboniferous Ostracoda. Annals and Magazine of Natural History, London, series 5, 18, 249-269.
Kido, E., Suttner, T. J., Waters, J. A., Ariunchimeg, Y., Sersmaa, G., Atwood, J. W., & Webster, G. D. (2013). Devonian deposits of the Baruunkhuurai Terrane, western Mongolia (IGCP 596 Field Workshop). Episodes, 36(4), 242–254. https://doi.org/10.18814/epiiugs/2013/v36i4/001.
Kotschetkova, N.M. (1980). New Ostracodes of the late Tournaisian in the Urals. In: Palaeozoic Stratigraphic and palaeontology in the south of Urals. Akad. Nauk URSS, section bachkirienne, 60–66 (in Russian with English summary).
Kulagina, E. I. (2013). Taxonomic diversity of foraminifers of the Devonian-Carboniferous boundary interval in the south Urals. Bulletin of Geosciences, 88(2), 265–282.
Lethiers, F. (1984). Zonation du Dévonien Supérieur par les ostracodes (Ardenne et Boulonnais). Revue de Micropaleontologie, 27, 30–42.
Li, P., Sun, M., Rosenbaum, G., Jourdan, F., Li, S., & Cai, K. (2017). Late Paleozoic closure of the Ob-Zaisan Ocean along the Irtysh shear zone (NW China): implications for arc amalgamation and oroclinal bending in the central Asian Orogenic Belt. Geological Society of American Bulletin, 129(5–6), 547–569. https://doi.org/10.1130/B31541.31541.
Liebau, A. (2005). A revised classification of the higher taxa of the Ostracoda (Crustacea). Hydrobiologia, 538, 115–137.
Melnyk, D. H., & Maddocks, R. F. (1988). Ostracode biostratigraphy of the Permo-Carboniferous of central and north-central Texas, part I: paleoenvironmental framework. Micropaleontology, 34, 1–20.
Metcalfe, I. (2011). Palaeozoic-Mesozoic history of SE Asia. Geological Society London, Special Publications, 355, 7–35. https://doi.org/10.1144/SP355.2.
Nazik, A., Çapkınoğlu, Ş., & Şeker, E. (2012). Famennian Ostracods from İstanbul zone (Gebze, Kocaeli, NW Turkey) and their Paleogeographical relations. Geologica Carpathica, 63(5), 355–363.
Nemirovskaya, T. I., Chermnykh, V. A., Kononova, L. I., & Pazukhin, V. N. (1992). Conodonts of the Devonian-carboniferous boundary section, Kozhim, Polar Urals, Russia. Annales Societe Geologique Belgique, 115, 629–647.
Olempska, E. (1979). Middle to Upper Devonian Ostracoda from the southern Holy Cross Mountains, Poland. Palaeontologia Polonica, 40, 57–162.
Olempska, E. (1999). Silicified shallow-water ostracodes from the early Carbonifeous of South China. Acta Palaeontologica Polonica, 44(4), 383–436.
Pokorny, V. (1950). The ostracods of the middle Devonian red coral limestones of Celechowice. Sbomik Statniho Geologiceskeho Ustavu Ceskoslovenske Republiky, 17, 513–630.
Polenova, E. N. (1952). Ostracoda from the Upper Givetian of the Russian platform. Trudy VNIGRl, n. s., 60, 65–156.
Rome, R. (1971). Contribution a l’etude des Ostracodes du Tournaisien inferieur de la Belgique. Memoires de l'Institut geologique de l’Universite de Louvain, 27, 1–46.
Safonova, I., Kotlyarov, A., Krivonogov, S., & Xiao, W., (2017). Intra-oceanic arcs of the Paleo-Asian Ocean. Gondwana Research, 50, 167–194. https://doi.org/10.1016/j.gr.2017.04.005.
Sanchez de Posada, L. C., Sanz-Lopez, J., & Gozalo, R. (2008). Ostracod and conodont faunal changes across the Frasnian-Famennian (Devonian) boundary at Els Castells, Spanish central Pyrenees. Revue de Micropaleontologie, 51, 205–219.
Sars, G. O. (1866). Oversigt af marine ostracoder. Norske VidenskapsAkademi, Förhandlingar, 1865, 1–130.
Sars, G. O. (1887). Nye bidrag til kundskaben om middelhavets invertebrafauna: 4. Ostracoda mediterranea (sydeuropaeiske ostracoder). Archiv for Mathematik og Naturvidenskab, 12, 173–324.
Shaver, R.H. (1961). Family Bairdiocyprididae Shaver, n. fam. In R.C. Moore (ed.) Treatise on Invertebrate Paleontology, Part Q, Arthropoda 3. Geological Society of America and University of Kansas Press, Boulder and Kansas, Q.364-Q.368.
Song, J.-J., & Gong, Y.-M. (2019). Ostracods from the Devonian-Carboniferous transition in Dushan of Guizhou, South China. Palaeobiodiversity and Palaeoenvironments, 99, 117–127. https://doi.org/10.1007/s12549-018-0322-7.
Song, J.-J., Crasquin, S., & Gong, Y.-M. (2017). Ostracods of the Late Devonian Frasnian/Famennian transition from Western Junggar, Xinjiang, NW China. Alcheringa: An Australasian Journal of Palaeontology, 41(2), 250–276.
Song, J.-J., Crasquin, S., & Gong, Y-M- (2019). Late Devonian benthic ostracods from western Junggar, NW China: implications for palaeoenvironmental reconstruction. Geological Journal, 54, 91–100. https://doi.org/10.1002/gj.3156.
Spalletta, C., Perri, M. C., Over, D. J., & Corradini, C. (2017). Famennian (Upper Devonian) conodont zonation: revised global standard. Bulletin of Geosciences, 91, 31–57. https://doi.org/10.3140/bull.geosci.1623.
Suttner, T. J., Kido, E., Ariunchimeg, Y., Sersmaa, G., Waters, J. A., Carmichael, S. K., Batchelor, C. J., Ariuntogos, M., Hušková, A., Slavík, L., Valenzuela-Ríos, J. I., Liao, J.-C., & Gatovsky, Y. A. (2019). Conodonts from Late Devonian island arc settings (Baruunkhuurai Terrane, western Mongolia). Palaeogeography, Palaeoclimatology, Palaeoecology (pp. 1–22). Elsevier. https://doi.org/10.1016/j.palaeo.2019.03.001.
Swartz, F. M. (1936). Revision of the Primitiidae and Beyrichidae with new Ostracoda from the Lower Devonian of Pennsylvania. Journal of Paleontology, 10, 541–586.
Ulrich, E.O., & Bassler, R.S., (1906). New American Paleozoic Ostracoda. Notes and descriptions of the Upper Carboniferous genera and species. United States National Museum Paper, 30, 149–164.
Ulrich, E. O., & Bassler, R. S. (1908). New American Paleozoic Ostracoda. Preliminary revision of the Beyrichiidae with descriptions of new genera. Proceedings of the United States National Museum, 35, 277–325.
Wang, S. Q. (1988). Late Paleozoic ostracode associations from South China and their paleoecological significance. Acta Palaeontologica Sinica, 27, 91–102 [in Chinese with English abstract].
Waters, J. A., Waters, J. W., Königshof, P., Carmichael, S. K., & Ariuntogos, M. (2020). Famennian crinoids and blastoids (Echinodermata) from Mongolia. Palaeobiodiversity and Palaeoenvironments. (this issue).
Webster, G. D., & Ariunchimeg, Y. (2004). The northern most Emsian crinoid known, a Devonian fauna from the Chuluun Formation, Shine Jinst area, Southern Mongolia. Geobios, 37, 481–487.
Windley, B. F., Alexeiev, D., Xiao, W., Kröner, A., & Badarch, G. (2007). Tectonic models for accretion of the central Asian Orogenic Belt. Journal of the Geological Society, 164, 31–47. https://doi.org/10.1144/0016-76492006-022.
Xiao, W. J., Huang, B. C., Han, C. M., Sun, S., & Li, J. L. (2010). A review of the western part of the Altaids: a key to understanding the architecture of accretionary orogens. Gondwana Research, 18(2–3), 253–273. https://doi.org/10.1016/j.gr.2010.01.007.
Yang, G., Li, Y., Santosh, M., Yang, B., Zhang, B., & Tong, L. (2013). Geochronology and geochemistry of basalts from the Karamay ophiolitic mélange in West Junggar (NW China): Implications for Devonian–Carboniferous intra-oceanic accretionary tectonics of the southern Altaids. Geological Society of America Bulletin, 125(3–4), 401–419. https://doi.org/10.1130/B30650.1.
Yang, G., Li, Y., Tong, L., & Yang, B. (2015). Oceanic Island basalts from the Darbut and Karamay Ophiolitic Mélange in West Junggar (NW China): product of a Middle Devonian Mantle Plume? Acta Geologica Sinica, 89, 105–106. https://doi.org/10.1111/1755-6724.12308_63.
Zagora, K. (1968). Ostracoden aus dem Grenzbereich Unter-Mitteldevon von Ostthüringen. Geologie, 17, 1–91.
Zaspelova, V. S. (1959). Ostracoda and their value for the stratigraphy of the Devonian of the north−western regions of the Russian platform. Trudy Vsesoûznogo neftânogo naučno-issledovatel’skogo geologo-razvedočnogo instituta (VNIGRI), Mikrofauna SSSR 10. Novaâ seriâ, 136, 5–131.
Żbikowska, B. (1983). Middle to Upper Devonian ostracods from northwestern Poland and their stratigraphic significance. Palaeontologia Polonica, 44(3), 108.
Zhuravlev, A. V., Sokiran, E. V., Evdokimova, I. O., Dorofeeva, L. A., Rusetskaya, G. A., & Małkowski, K. (2006). Faunal and facies changes at the Early-Middle Frasnian boundary in the north-western east European platform. Acta Palaeontologica Polonica, 51(4), 747–758.
Acknowledgments
The authors are grateful to Jana Anger (Senckenberg Institute, Frankfurt) for conodont sample preparations and selections of ostracod specimens under the binocular microscope. Also, A. Nazik would like to thank Peter Königshof (Senckenberg Institute, Frankfurt) inviting me for studying ostracod findings from Mongolia. We appreciate the Çukurova Üniversity, Central Laboratory Management and Expert Derya Akalan taking of scanning electron microscope photographs. The PhD student (A.M.) and her supervisor (P.K.) acknowledge funding from both, German Academic Exchange Service (DAAD, Research Grant–Doctoral Programme in Germany, September 01, 2018–September 01, 2021; 57381412) and German Science Foundation (DFG, KO 1622-19/1). This paper is a contribution to IGCP Project 652 (Reading geologic time in Palaeozoic rocks: the need for an integrated stratigraphy [2017–2021]). We are grateful to Junjun Song (China) and Marie-Béatrice Forel (France) for their critical comments and suggestions on the manuscript.
Funding
This study was supported by the German Academic Exchange Service (DAAD, Research Grant–Doctoral Programme in Germany, September 01, 2018–September 01, 2021; 57381412) and German Science Foundation (DFG, KO 1622-19/1).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This is a contribution to a special series on The Central Asian Orogenic Belt (CAOB) during Late Devonian: new insights from southern Mongolia.
Appendix
Appendix
Taxonomic list of identified ostracod taxa from Hushoot Shiveetiin gol section, Mongolia
Class Ostracoda Latreille, 1802
Order Palaeocopida Henningsmoen, 1953
Superfamily Hollinoidea (=Hollinacea) Swartz 1936
Family Hollinellidae Bless and Jordán 1972
Hollinella (Keslingella) lionica Becker and Bless, 1971 (Fig. 4/a-d)
Hollinella (Keslingella) aff. praecursor Pokorny, 1950 (Fig. 4/e-g)
Superfamily Aechminoidea Bouček, 1936
Family Aechminidae Bouček, 1936
Aechmina sp. (Fig. 4/i)
Family Ctenoloculinidae Jaanusson and Martinsson, 1956
Parabolbina sp. (Fig. 4/j)
Superfamily Primitiopsoidea (=Primitiopsacea) Swartz 1936
Family Pribylitidae Pokorný, 1958
Pribylites junggarensis Song and Crasquin, 2017 in Song et al. 2017 (Fig. 4/k-n)
Coryellina sp. (Fig. 4/o)
Superfamily Kirkbyoidea (=Kirkbyacea Ulrich and Bassler 1906)
Family Amphissitidae Knight, 1928
Amphissites centrocostatus Blumenstengel, 1969 (Fig. 5/a)
Amphissites irinae Glebovskaja and Zaspelova in Egorov, 1953 (Fig. 5/b-e)
Polytylites sp. (Fig. 5/f)
Family Nezamysliidae Żbikowska, 1983
Nezamyslia sp. (Fig. 5/g–i)
Family Kirkbyellidae Sohn, 1961
Berdanella sp. (Fig. 5/j–k)
Palaeocopina? indet. 1 (Fig. 5/l–n)
Palaeocopina? indet. 2 (Fig. 5/o, Fig. 6/a–f)
Order Platycopida Sars 1866
Superfamily Kloedenelloidea (=Kloedenellacea Ulrich and Bassler 1908)
Family Kloedenellidae
Kloedenellidae indet. (Fig. 6/g–h)
Family Geisinidae Sohn, 1961
Hypotetragona tremula Becker, 1971 (Fig. 6/i)
Family Beyrichiopsidae Henningsmoen, 1953
Beyrichiopsis hushootensis sp. nov. (Fig. 6/j–o, Fig. 7/a)
Order Podocopida Sars 1866
Superfamily Bairdioidea (=Bairdiacea Sars 1887)
Family Bairdiidae Sars, 1888
Bairdia hypsela Rome 1971 (Fig. 7/b)
Bohlenatia sp. (Fig. 7/c)
Pustulobairdia sp. (Fig. 7/d)
Superfamily Bairdiocypridoidea (=Bairdiocypridacea Shaver, 1961)
Family Bairdiocyprididae Shaver, 1961
Baschkirina sp. (Fig. 7/e)
Bairdiocypris sp. (Fig. 7/f–g)
Family Pachydomellidae Berdan and Sohn 1961
Microcheilinella bulongourensis Song and Crasquin, 2017 (Fig. 7/h–j)
Ampuloides beckeri sp. nov. (Fig. 7/k–o)
Superfamily Healdioidea (=Healdiacea Harlton 1933)
Family Healdiidae
Cribroconcha primaris Kotschetkova, 1980 (Fig. 8/a–c)
Cribroconcha honggulelengensis Song and Crasquin, 2017 (Fig. 8/d–f)
Rights and permissions
About this article

Cite this article
Nazik, A., Königshof, P., Ariuntogos, M. et al. Late Devonian ostracods (Crustacea) from the Hushoot Shiveetiin gol section (Baruunhuurai Terrane, Mongolia) and their palaeoenvironmental implication and palaeobiogeographic relationship. Palaeobio Palaeoenv 101, 689–706 (2021). https://doi.org/10.1007/s12549-020-00446-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-020-00446-z