
Abstract
The Suoidea from Gračanica in the Bugojno Basin (Bosnia and Herzegovina) are assigned to Choeromorus lemuroides (previously Taucanamo sansaniense) (Taucanaminae, Palaeochoeridae), Bunolistriodon latidens (Listriodontinae, Suidae), and Conohyus simorrensis (Tetraconodontinae, Suidae). These belong to three anagenetic lineages, known from Western Europe to Anatolia. The fossils from Gračanica and these lineages are described in detail. Biostratigraphic correlations in an area extending from Western Europe to Anatolia are proposed on the basis of these lineages. These correlations are consistent with known independent age estimates of the large mammal localities in the different areas. Because the Suoidea from Gračanica belong to groups which have suffered from recent taxonomic inflation, their systematics has been discussed for a correct classification.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The interplay between tectonics and sea level greatly changed European Miocene geography, and the reduction of the Paratethys to what are now the Black and Caspian Seas is part of this process (Rögl and Steinginger 1983; Rögl 1998; Popov et al. 2004). Superposed climatic change led to complex biogeographic changes, which are still far from understood. Europe was not biogeographically homogenous and even in Western Europe, there were many differences in the timing of faunal events (Van der Made et al. 2006; Kälin and Kempf 2009; Van der Meulen et al. 2011, 2012). Today, the limit between the Palearctic and Saharo-Arabian biogeographic realms passes just south of Anatolia (Holt et al. 2013), but there is evidence that from the Mio- till the Pleistocene a long-standing biogeographic limit between Anatolia + Greece and western + central Europe existed (Van der Made and Mateos 2010). This could reflect a different position of the limit between the two biogeographic realms in the Neogene, though alternatively, it may reflect that a wide transition zone with limits of individual taxa at different geographic positions.
The Middle Miocene fossil locality of Gračanica in Bosnia and Herzegovina is thus biogeographically in a very interesting position. Its fauna is a welcome addition to the relatively poor Middle Miocene fossil record of the area between west + central Europe and Greece + Anatolia. It should not be strange, that there are difficulties of biostratigraphic correlation between those areas. Fossil localities in the intermediate area are of interest in that they provide information on biogeography and correlation. Fahlbusch (1991, p. 168) recognised the difficulties arising from the complex biogeographic history of Europe and the need of unequivocal boundary definitions of the biozones. Since dispersal events may be diachronic, he considered the use of stages-of-evolution in continuously evolving lineages as the best method for the objective definition of time boundaries. The fossil Suoidea from Gračanica belong to three lineages, which are known from the Iberian Peninsula to Anatolia. Without pretending that these lineages should be used to define the boundaries of biozones, they have been used previously in correlation between these areas (Van der Made 2003, 2005). At present, there is much more material available to describe the evolution of these lineages which could support or contradict these correlations.
All three suoid lineages where subject to publications, which greatly complicated their taxonomy (e.g. Pickford 2011, 2014, 2016a, 2016b, 2017; Pickford and Laurent 2014). In general, there is much splitting, but also, the rules of the International Code of Zoological Nomenclature are not followed. This obscures evolution and biostratigraphy and affects the classification of the Suoidea from Gračanica at the species, genus, subfamily, and family level. Therefore, it is necessary to discuss taxonomy of the taxa from Gračanica in detail.
The aims of this paper are (1) to describe the fossil Suoidea from Gračanica; (2) to classify them, which implies discussions to clarify taxonomy; (3) to describe their evolutionary lineages quantitatively; and (4) to use these lineages in correlation in the area extending from Western Europe to Anatolia. The major taxonomical problems will be discussed in the Suppl.-Information.
Material and methods
Tooth nomenclature follows Van der Made (1996a) and Prieto et al. (2017). Upper teeth are indicated with superscript (e.g. M3, P2), lower teeth with subscript (M3, P2), and if no separation between upper and lower is made, neither sub-nor superscipt is used (M3, P2). The measurements are taken as indicated by Van der Made (1996a) and are given in mm (unless indicated otherwise). The measurements are indicated in Tables 1, 2, 3, 4, 5 and in the figures with acronyms, which are explained in the Suppl.-Tab. 1.
The material presented here is housed in the Natural History Museum of Vienna, Austria (NHMW). The specimens from Gračanica are compared with specimens from other localities (Fig. 1; Suppl.-Fig. 1; Suppl.-Tab. 2). In Suppl.-Tab. 2, descriptions of those materials are also listed, with the classification used in those publications. In this table, acronyms are used to indicate in which institutes the material was studied, or where it is kept presently. These acronyms are explained in Suppl.-Tab. 3. If no acronym of a collection is given, the data used come from these publications. The stratigraphy of many of these localities is discussed in the Suppl.-Information and the ages are summarised in Suppl.-Tab. 4, where also the relevant literature on stratigraphy or age is given.

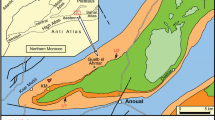
Geographic position of the localities with Choeromorus and Schizoporcus (top), the different species of the B. adelli-latidens-meidamon lineage (middle) and Conohyus and Retroporcus (bottom)
Systematics
The material from Gračanica is described in this section, while information and discussions on taxonomy are presented in the Suppl.-Information. The classification of the taxa mentioned in the text is given in Suppl.-Fig. 2, where it is compared to the classification of other authors.
Palaeochoeridae Matthew, 1924
Selected synonymy
-
1899–1900 Palaeochoeren—Stehlin: pp. 29–522 (intentionally not used as a formal name).
-
1899–1900 Palaeochoeriden—Stehlin: pp. 182, 225, 269, 320, 328, 331, 410–411, 432, 433, 439–440, 453, 481–482 (intentionally not used as a formal name).
-
1909 Palaeochoeridés—Stehlin: p. 505 (not used as a formal name).
-
1924 Palaeochoerinae—Matthew: p. 176.
-
1996b Palaeochoeridae Matthew 1924-Van der Made: p. 302.
-
2011 Yunnanochoerini nov.—Pickford: p. 571.
-
2017 Siderochoeridae nov.—Pickford: p. 43.
-
2017 Schizoporcidae van der Made 2010—Pickford: p. 34.
-
2017 Doliochoeridae Simpson 1945—Pickford: p. 24.
Taucanaminae van der Made, 1997a
Selected synonymy
-
1997a Taucanamini new tribe—Van der Made: p. 131.
-
2010 Taucanaminae Van der Made, 1997—Van der Made: pp. 46, 47, 114.
-
2017 Choeromorinae nov.—Pickford: p. 55.
Choeromorus Gervais, 1850
Selected synonymy
-
1850 Choeromorus Lartet—Gervais: p. 7. Type species: Choeromorus mammilatus Gervais, 1850.
-
1851 Choerotherium Lart.—Lartet: p. 33. Type species: Choerotherium sansaniense Lartet 1851.
-
1945 Taucanamo, new name—Simpson: p. 146. Type species: Choerotherium sansaniense Lartet 1851.
-
2017Siderochoerus nov.—Pickford: p. 43. Type species: Siderochoerus minimus Pickford 2017.
Choeromorus lemuroides (Blainville, 1847)
Selected synonymy
-
1847 S. lemuroides—Blainville: p. 231, pl. 9.
-
1850 Choeromorus mamillatus—Gervais: p. 7, Pl. 33 fig. 4.
-
1850 Choeromorus simplex—Gervais: p. 7, Pl. 33 fig. 5.
-
1851 Choerotherium Sansaniense-Lartet: p. 33.
-
1949 Taucanamo (= Choerotherium) sansaniensis (Lart.)—Thenius: 163 (only the material from Sansan).
-
1956Taucanamo sansaniense (Lartet)—Thenius: p. 366 (only the material from Sansan).
-
1956Taucanamo pygmaeum (Depéret)—Thenius: fig. 31 (the material from Göriach, not figs. 29–30).
-
2012 Choeromorus mammilatus Gervais, 1850—Pickford: pp. 251–265, figs. 1–23.
-
2017Choeromorus lemuroides (Blainville, 1847)—Pickford: pp. 57–73, figs. 42–57.
Holotype: MNHN Sa 4593, symphysis with right I2 and roots or alveoles of the remaining incisors, canines and right and left P1–3, figured by De Blainville (1847, Genus Sus plate 9) and Pickford (2017, fig. 42).
Type locality: Sansan, France
Age of the type locality: Middle Miocene, Middle Aragonian, early Astaracian, MN6
Material
NHMW 2013/0008/0001a—left maxilla with P3 to M3, of the M3 only the anterior lobe is preserved.
NHMW 2013/0008/0001b—right M2
NHMW 2013/0008/0001c—left P3
NHMW 2013/0008/0001d—left P4
NHMW 2013/0008/0001e—left M1
Description and comparison
Of the M3 (Fig. 2), only the first lobe is preserved. It is similar to that of the M2.

Choeromorus lemuroides from Gračanica: NHMW 2013/0008/0001a-left maxilla with P3-M3: a buccal, b occlusal, and c lingual views
The M2 (Figs. 2 and 3) has the roots close together and still fused at a distance of the crown. Fused lingual roots are typical of the Palaeochoeridae (“Old World peccaries”) and unlike in the Suidae. All roots are fused until some distance below the crown (taurodonty), what also occurs in the Palaeochoeridae. The crown is relatively low. The protopreconule (protoconule) is fused to the protocone as in the Palaeochoeridae and not to the cingulum as in most Suidae. The protoprecrista and paraprecrista meet anterior of the proto- and paracone, but posterior of the cingulum. This is a step towards lophodonty. This is like in the later species of Choeromorus and in Pecarichoerus, while Schizoporcus and Yunnanochoerus have better developed lophs. No tetrapreconule (central cusp) is developed and there is only a tetraprecrista, which probably was low. Palaeochoeridae tend to have a small or no tetrapreconule, while most Suidae have a well-developed tetrapreconule. While the tetraprecrista is directed forwards, there is no metaprecrista and no loph is formed by the fusion of these two crests. A longitudinal valley keeps these cusps separate. Such sublophodont morphologies are common in the Taucanaminae. The tetraprecrista meets a protoendocrista. This is unlike in Schizoporcus muenzen-bergensis, where the latter crest is not developed. In the sample from Sansan, it is variable. In Choeromorus inonuensis, it is better developed, but also not universal. There is no cusp in the middle of the posterior cingulum (tetrapostconule) and the tetrapostcrista ends as a narrow crest. This is like in many Palaeochoeridae and unlike in the Suidae. The M2 is narrow as Ch. lemuroides, Ch. inonuensis, and Choeromorus grandaevus, while those of Choeromorus primus and S. muenzenbergensis are wider (Fig. 3). The size (Table 1) is comparable to those of the first two species.

Choeromorus lemuroides from Gračanica: 1NHMW 2013/0008/0001b—right M2: a anterior, b posterior, c buccal, d occlusal, and e lingual views. Bivariate diagrams comparing the widths of the anterior (DTa) or posterior lobes (DTp) and length (DAP) of the teeth from Gračanica with: Ch. primus from Petersbuch 2 and Els Casots; Ch. lemuroides from Sansan; Ch. inonuensis from Inönü I and Paşalar; Ch. grandaevus from Steinheim, La Grive and Anwill; S. muenzenbergensis from Münzenberg, Sandelzhausen and Neudorf Spalte. The lines indicate average proportions
The M1 (Fig. 2) is much worn and little is preserved of the crown. As far as can be seen, the lingual roots are fused as in the Palaeochoeridae and unlike in the Suidae, where they are separate. They are convergent, as in the Taucanamini and Doliochoerini, and not divergent as in the Palaeochoerini. The proportions are as in the M2. While the M2 is in the upper range of the Ch. lemuroides sample, the M1 is in the lower range (Fig. 3). A large M2 and large M3, compared to the M1, are related features in Suoidea. In Choeromorus, this is a progressive feature.
The P4 (Fig. 2) has the para- and metacone placed very close together. They are not separated by a deep or clear furrow on the lingual, nor on the buccal side. Seen from the side, they form a pointed structure, rising far above the cingulum. There is no paraprecrista and no parapreconule (“sagittal cusp”). The protopre- and protopostcristas are short and there is no protoendocrista. All these features are primitive in the Suoidea and common in the Palaeochoeridae. The posterolingual cingulum forms a wide plateau. The tooth is longer than wide (Table 1; Fig. 3), which occurs in some of the species of Choeromorus, unlike what is common in the Suidae, Palaeochoerinae, and Schizoporcus. The size is in the lower ranges of Ch. lemuroides.
The P3 (Fig. 2) has a large paracone with pre- and postcristas. The protocone is a low cusp, arising from the posterolingual cingulum. The tooth is relatively elongate as in Ch. lemuroides and unlike in Ch. primus or Ch. grandaevus or S. muenzenbergensis. The size is in the range of Ch. lemuroides (Fig. 3).
The M1 is much worn and nothing of the crown morphology is preserved. The tooth is elongate. The value indicated in Fig. 4 for the length (DAP) is a minimum value because of wear; the tooth was longer. The tooth is in the ranges of Ch. lemuroides and is smaller than the M1 of Ch. inonuensis (Fig. 4).

Choeromorus lemuroides from Gračanica: 1 NHMW 2013/0008/0001c—left P3: a buccal, b occlusal, and c lingual views; 2 NHMW 2013/0008/0001d—left P4: a buccal, b occlusal, and c lingual views. Bivariate diagrams comparing the width of the anterior (DTa) or posterior lobe (DTp) and length (DAP) of these premolars and the M1 with C. minimus from Petersbuch 62; Ch. primus from Petersbuch 2 and 7, Artenay, Montréal-du-Gers, Bézian and Els Casots; Ch. lemuroides from Petersbuch 31 and 68, Göriach, Thannhausen and Sansan and; Ch. inonuensis from Inönü I, Paşalar, Petersbuch 39 and 108 and Prebreza; Ch. grandaevus from Steinheim, Przeworno and La Grive; S. muenzenbergensis from Münzenberg, Neudorf Spalte, Sandelzhausen and Petersbuch 54. The lines indicate average proportions
The P4 (Fig. 4 (2)) has only one main cusp. This is the primitive state for the Suoidea. In the Palaeochoeridae, it occurs in the Taucanamini and in the Suidae in the Tetraconodontinae and is occasionally known from some Hyotheriinae. Schizoporcus has a P4 with a well-developed metaconid. There is a well-developed protoprecrista and where this reaches the wide anterior cingulum, a protopreconulid is formed. There is a protopost- and protoendocristid. The latter is directed posterolingually. There is a very low and well-worn hypoconid. The low hypoconid correlates to the paracone and metacone on the P4 being very close together and high, a primitive feature in the Suoidea. Primitive Suidae, like Hyotherium have already a larger hypoconid, and as a result, the para- and metacone on the P4 form a lower and less pointed structure. Next to the hypoconid, there is another small cusplet on the talonid. The tooth is very elongate for a P4 and in this respect, it is similar to Ch. lemuroides and Ch. inonuensis and it is also in their metrical ranges (Fig. 3).
The P3 (Fig. 4 (1)) has a shape that is similar to that of the P4, but it is simpler. In particular, the hypoconid is very small and not well differentiated from the protopostcristid. In side view, the tooth is triangular and this cusp does not even show up. This is a primitive morphology. Compared to the premolars of suids like Hyotheriinae, Listriodontinae, and Suinae, the P4 is relatively high and pointed. Metrically, it is within the ranges of Ch. lemuroides.
Specific and generic classification of the palaeochoerid from Gračanica
The fused lingual roots and shape of the protopreconule of the upper molars are similarities to the Palaeochoeridae. The molars with crests that suggest lophodonty are a similarity to several Taucanaminae, while the later species of Schizo-porcus and Yunnanochoerus have better developed anterior lophs in the upper molars. The P4 with a single main cusp is a difference with Schizoporcus. The elongate molars and premolars are a similarity to the later species of the genus Choeromorus (= Taucanamo). The size is relatively large for this genus and best fits Ch. lemuroides (= T. sansaniense) (Fig. 5). The material is assigned here to the latter species. The classification by Van der Made (2010) is used to arrive to this identification. However, subsequently, a more complex classification of these Suoidea has been proposed (Pickford 2011, 2012, 2016b, 2017), which would change the species, genus, tribe, subfamily, and family name of the suoid from Gračanica. Those proposals are discussed in the Suppl.-Information.

The size of the M3 and the size of the M3 relative to the M1 in Choeromorus and S. muenzenbergensis. The localities are ordered from old at the bottom to young at the top. Provenance of data as in Suppl.-Tab. 2
Suidae Gray, 1821
Listriodontinae Gervais, 1859
Bunolistriodon Arambourg, 1963
Selected synonymy
-
1933 Bunolistriodon—Arambourg: p. 137 (name not available, because no type species was indicated).
-
1963 Bunolistriodon Arambourg 1933—Arambourg: pp. 903–904, with type species Bunolistriodon lockharti.
-
1995 Eurolistriodon nov.—Pickford and Moyà Solà: p. 344 (name not available, because no type species was indicated).
-
2006 Eurolistriodon gen. nov.—Orliac: pp. 968–969, with type species Eurolistriodon adelli Pickford and Moyà-Solà 1995.
Bunolistriodon latidens (Biedermann, 1873)
Selected synonymy
-
1873 Sus latidens m.—Biedermann: p. 11, pl. 7
-
1899–1900 Listriodon latidens Biedermann—Stehlin: pp. 13, 85, 173, 285, 327, 426, 462, pl.1, fig. 15, pl. 3, figs. 31–33
-
1990a Bunolistriodon latidens (Biedermann 1873)—Van der Made: pp. 86, 93
-
1995 Eurolistriodon latidens—Pickford and Moyà Solà
Holotype: NSSW no. 99—mandible with left and right P2-M3, Cm, I1 and left I2, figured by Biedermann (1873, pl. 1, fig. 15), Stehlin (1899–1900, pl. 1 fig. 15, pl. 3, fig. 31–33) and Van der Made (1996a, pl. 18, figs. 2-3).
Type locality: Veltheim, Switzerland.
Age of the type locality: Middle Miocene, MN5
Material
NHMW 2013/0012/0001—right maxilla with M2–3; same individual as 0002
NHMW 2013/0012/0002—left M3 and lingual half of left M2; same individual as 0001
NHMW 2013/0012/0003—left maxilla with P4 and roots of a completely worn M1
NHMW 2013/0012/0004—right I1–3 and left Cm
NHMW 2013/0012/0005—right premaxilla with root of the I1 and an alveolus of the I2/3 and part of the maxilla with flattened Cm
NHMW 2013/0012/0006—right maxilla with P3–4 and M3 and with roots/alveoles of M1–2
NHMW 2013/0012/0007—left maxilla with P4-M3; probably same individual as 0008
NHMW 2013/0012/0008—left mandible with M1–3 and right mandible with P2-M3, pressed together
NHMW 2013/0012/0009—left I1
NHMW 2013/0012/0010—left P4
NHMW 2013/0012/0011—fragment of left M3; lingual wall of second and third lobes
NHMW 2013/0012/0012—second phalanx III or IV, right of the axis of the foot
NHMW 2013/0012/0013—right astragalus, proximo-medial fragment
Description and comparison
The M3 (Fig. 6 (6)) has the protoendocristid and metaendocristid directed to each other and connecting behind the centers of the proto- and metaconid. The protoconid and metaconid are massive cusps; between them and anterior to the endocristids, there is a clear anteroposterior furrow. The result is close to a loph, as occurs in Bunolistriodon and not a complete loph, as in Listriodon. There is a hypopreconulid in the middle of the transverse valley, as in many Suidae, including Bunolistriodon, while in Listriodon, there is just a hypoprecristid. The hypoconid and entoconid remain well separated by a clear antero-posterior directed valley. With a primitive loph in the anterior lobe and no loph in the posterior lobe, the tooth is sub-lophodont. Nearly all Suidae are bunodont, without any lophs, while within the Listrio-dontinae sub-lophodonty and lophodonty are found. Al sublophodont Palaeochoeridae are much smaller. The third lobe consists of a large pentaconid and a pentapreconulid close to the axis of the tooth.

Bunolistriodon latidens from Gračanica: 1 NHMW 2013/0012/0007—left maxilla with P4-M3: a occlusal view, b buccal view of the P4; 2 NHMW 2013/0012/0006—right maxilla: a occlusal view of the P3, b buccal view of the P4, c occlusal view of the P4; 3 NHMW 2013/0012/0010—left P4: a anterior, b buccal, c occlusal, d lingual, and e posterior views; 4 NHMW 2013/0012/0003—left P4: a buccal andb occlusal views; 5 NHMW 2013/0012/0002—left M3: occlusal view; 6 NHMW 2013/0012/0008—left M3: a buccal, b occlusal, and c lingual views
The M2 and M1 have a similar structure, but lack a third lobe. These molars are relatively small. Though there is a large overlap in size between the larger B. lockharti and the smaller Bunolistriodon adelli and B. latidens, the size of these molars is more similar to that of the latter species.
The M3 (Fig. 6 (1, 5)) has a protoprecrista that is directed to the anterior side of the paracone. When it reaches that cusp, it is very low, but the two connect, forming a primitive loph. No protopreconule is formed on the cingulum. The tetraprecrista is directed to the centre of the tooth, where it ends, without connecting to the metacone, nor giving rise to a well-individualized tetrapreconule in the middle of the transverse valley. This structure is called sub-lophodont, as occurs in Bunolistriodon. The third lobe consists of a pentacone, situated lingually of the axis of the tooth.
The M2 and M1 (Fig. 6 (1)) have morphologies similar to that of the M3, save for lacking a third lobe.
The P4 (Fig. 6 (3)) has a well-developed metaconid placed lingually of the protoconid, forming a transverse loph. There is a flat transverse facet extending over the posterior side of this loph (Fig. 6 (3e)). There is a furrow on the anterior side between these two cusps (Fig. 6 (3a)), as in Bunolistriodon, while in Listriodon, there is a wide depression but no furrow. The hypoconid is placed on the buccal side of the talonid.
The P3 and P2 do not have any indication of a metaconid and the talonids do not have a clear hypoconid. Their morphologies are simple as in Bunolistriodon.
The P4 (Fig. 6 (1, 2, 4)) has a large protoprecrista directed anterobuccally, ending near the axis of the tooth and without forming a loph with the paracone, even though in a worn tooth, the dentine pattern suggests such a structure (Fig. 6 (4b)). The metacone may be more (Fig. 6 (1, 2)) or less individualized (Fig. 6 (4)).
The P3 (Fig. 6 (2)) consists of a large paracone and a small protocone, which is not connected to the cingulum. The latter morphology is common in the Listriodontinae. The protocone and cingulum do not extend much lingually and the tooth is not so wide as in the later Listriodon. There is no cusp or cusplet formed within the protopostcrista. Especially in the later Listriodon primus, such a cusp is formed. Both morphologies are as in Bunolistriodon.
The Cm (Fig. 7) is high and probably it was hypselodont. It has a triangular section with the lingual and labial sides covered with enamel, but the posterior side not. The earliest Suoidea have such triangular teeth with the lingual sides much wider than the labial sides. These teeth are said to be “scrofic,” because they are as in Sus scrofa. In several lineages, the width of the labial side increased, giving rise to “verrucosic” canines. When the labial side became wider, the posterior side became relatively narrower. The different canine types can be well recognised in bivariate diagrams, or using ratios of the widths of the different sides (Fig. 7). Bunolistriodon lockharti and B. adelli tend to have canines with relatively wide posterior (Po) and narrow labial sides (La), while in B. latidens, the posterior side is relatively narrower and in Bunolistriodon meidamon, it is much narrower (Fig. 7). In both, the labial side is relatively wider. As seen in an index comparing the widths of the labial and lingual sides (Li), there is a tendency from B. adelli to B. latidens and B. meidamon to increase the width of the labial side (Fig. 7). The tooth from Gračanica fits best B. latidens.

Bunolistriodon latidens from Gračanica, NHMW 2013/0012/0004: 1 left Cm: buccal view. Bivariate diagrams comparing the width of the lingual (Li), buccal (Bu), and posterior sides (Po) of the Cm from Gračanica with those of the B. adelli-latidens-meidamon lineage and B. lockharti from: Pellécahus, La Artesilla, La Romieu, Chevilly (type locality of B. lockharti), Buñol, Córcoles, Langenau 1, Tavers and Montréal du Gers (type locality of B. tenezarensis; MHNT). The lines indicate average proportions. Diagrams showing the changes in time in La and index 100 La/Li through time. The localities are in approximate order from old at the bottom to young at the top. Veltheim is the type locality of B. latidens, Paşalar is the type locality of B. meidamon and Çandır is the type locality of B. meidamon ultimus
The Cm has tree enamel bands, as is normal in the Suidae: the pre-, post-, and endosyncline. The tooth has little wear and the tip, where the three synclines meet, is preserved. The tooth is flattened by diagenetic compression. It was directed outwards.
The I1 is damaged and the labial side of the crown is gone; what is left of the relevant morphology is similar to that in the I2.
The I2 (Fig. 8 (1)) has a transverse or mesiodistal diameter (DMD) that is much longer than the labiolingual diameter (DLL). This occurs only in Listriodontinae, Celebochoerus, Kolpochoerus, and Hylochoerus. The pre- and postanticlinid are very low, as only occurs in the Listriodontinae. There is no endocristid. This crest is weak or absent in the Listriodontinae; Listriodon retains a weak crest, but in Bunolistriodon, it tends to be weaker or absent. The DMD and DLL fit B. latidens, while the incisors of B. lockharti and B. adelli are narrower and those of B. meidamon wider (Fig. 8).

Bunolistriodon latidens from Gračanica, 1NHMW 2013/0012/0004: 1 right I2: a apical, b labial, and c lingual views; 2 right I3: labial view. Bivariate diagram comparing the mesodistal diameter (DMD) and labiolingual diameter (DLL) of the I2 from Gračanica with: B. lockharti from: La Romieu, Baigneaux-en-Beauce, Gerlenhofen, Langenau 1, Engelswies, Pontlevoy, Tavers and Montréal du Gers, B. adelli, B. latidens and B. meidamon. The lines indicate average proportions. Diagrams showing the changes in time in DMD and index 100 DMD/DLL through time. The localities are in approximate order from old at the bottom to young at the top. Montréal du Gers is the type locality of B. tenezarensis, Veltheim is the type locality of B. latidens, Paşalar is the type locality of B. meidamon and Çandır is the type locality of B. meidamon ultimus. The provenance of the data is indicated in Suppl.-Tab. 2
The I3 (Fig. 8 (2)) is damaged, but what is left shows that the crown was very wide, as in B. latidens and B. meidamon.
The I1 (Fig. 9 (1)) has a low and very elongate crown, with a marked lingual cingulum. The mesial facet is perpendicular to the long axis of the crown. This means that the two incisors formed a transverse crest, as in the Listriodontinae, and not a V-shape as in most Suoidea (see Van der Made 1996a, fig. 25; Van der Made 1997b, fig. 7). It has two clear, equally deep and long, groves on the labial side, separating the main cusp in the middle, from the pre- and postconules. This is as in Bunolistriodon, while Listriodon has only one large grove, which in very elongate specimens, is flanked by two minor groves. Bunolistriodon lockharti and B. adelli have incisors with a DMD that is about twice as long as the DLL, but B. latidens and B. meidamon have relatively much longer DMD (Fig. 9). The tooth from Gračanica fits B. latidens in size and proportions (Fig.10).

Bunolistriodon latidens from Gračanica. 1 NHMW 2013/0012/0009—left I1: a apical, b lingual, c mesial, d lingual, and e distal views. Bivariate diagram comparing the mesodistal diameter (DMD) and labiolingual diameter (DLL) of the tooth from Gračanica with: B. lockharti from: La Romieu, Baigneaux-en-Beauce, Córcoles, Gerlenhofen, Langenau 1, Ravensburg, Pontlevoy, Tavers, Montréal du Gers and Can Canals, and with B. adelli, B. latidens, and B. meidamon. The lines indicate average proportions. Montréal-du-Gers is the type locality of B. tenezarensis, Can Canals is the type locality of B. giganteus, Els Casots is type locality of B. adelli, La Retama is type locality of B. retamaensis, Veltheim is the type locality of B. latidens, Paşalar is the type locality of B. meidamon, and Çandır is the type locality of B. meidamon ultimus. In the lower graphs, the changes in DMD and the index 100 DMD/DLL are shown. The localities are ordered from old (bottom) to young (top). Provenance of data as in Suppl.-Tab. 2

Diagram comparing the 100 DMD/DLL indices of the I1, I2, and I1 as a percentage of the mean of the same tooth in the sample of Paşalar. The localities are in approximate order from old at the bottom to young at the top and the provenance of data is as indicated in Suppl.-Tab. 2
The premaxilla and anterior maxilla are crushed. The maxilla contains the Cm and the premaxilla the alveolus of the I1. The I2–3 are not present and no alveolus of the I2 can be seen, while it is not clear whether a depression is the alveolus of the I3. It is not clear whether these alveoles cannot be identified because the bone is crushed or whether the individual lost these teeth during life and the alveoles closed. In any case, the snout is very wide as is normal in the Listriodontinae.
Some foot bones are preserved but add little additional information.
Classification of the listriodont from Gračanica
The low crowned and wide incisors from Gračanica are indicative of the Listriodontinae. The two equally long and deep labial furrows on the I1 and the sublophodont molars indicate the genus Bunolistriodon (=Eurolistriodon). This is supported by the sublophodont molars, while Listriodon has fully lophodont molars. Here, the classification of Bunolistriodon of Van der Made (1996a, 1997b) is used. Other opinions on the classification of these Suidae are discussed in the Suppl.-Information.
The clearest differences between the different species of Bunolistriodon are seen in the proportions and sizes of the incisors and canines (Van der Made 1996a, figs. 35, 49), differentiating four species. Increased samples of the European and Anatolian I1, I2, and Cm of Bunolistriodon still show four clusters for each of these teeth (Fig. 7, 8, and 9). The clusters of the oblique crosses include a specimen from Chevilly (red circle), type locality of B. lockharti, supporting that these clusters belong to that species. The clusters of the black triangles consist mainly of fossils of the type locality of B. meidamon. The clusters of blue rhombs include material from type locality Veltheim of B. latidens. Bunolistriodon adelli from type locality Els Casots (red dot) clusters with the small black squares (Fig. 9), which in part represents material previously assigned to that species (Van der Made 1996a), supporting this assignation. These graphs support four distinct west Eurasian species: B. lockharti (= B. giganteus = B. tenezarensis), B. adelli (= B. retamaensis = ? B. michali), B. latidens, and B. meidamon.
The incisors from Gračanica (red inverted triangles in Figs. 8 and 9) are far away from those of B. lockharti and B. adelli, as well as from B. tenezarensis/giganteus and B. retamaensis (whether these species are valid or not). They are also far away from B. meidamon, while they are closest to the clusters of B. latidens (Figs. 8 and 9). This is also the case with the canines (Fig. 7). All other morphology described above is compatible with assigning the listriodont fossils from Gračanica to B. latidens.
Tetraconodontinae Lydekker, 1876
Conohyus Pilgrim, 1925
Conohyus simorrensis (Lartet, 1851)
Selected synonymy
-
?1850 Listriodon Lartetii—Gervais: p. 50, plate 20 fig. 1.
-
1851 Sus Simorrensis—Lartet: p. 33 (partially).
-
?1851 Sus? doati—Lartet: p. 33.
-
1859 Sus abnormis, Kaup—Kaup: pp. 7–9, pl. 2, fig. 1.
-
?1882 Sus Valentini—Filhol: pp. 123–124.
-
1972 ?Hyotherium sommeringi MEY. matritensis n. sp.—Golpe-Posse: p. 122
-
1972 Hyotherium sommeringi MEY. matritensis nova. sp.—Golpe-Posse: pp. 155–156.
-
1972 Hyotherium sommerringi matritense GOLPE, 1971—Golpe-Posse: p. 197, pl. 6, fig. 6b.
-
1972 Conohyus melendezi n. sp.—Golpe-Posse: pp. 149, 157.
-
1972 Conohyus melendezi n. sp. GOLPE, 1971—Golpe-Posse: pl. 2, fig. 2b.
-
1986 Conohyus ebroensis nov ap.—Azanza Asensio: pp. 95–105, pl. 2, fig. 1, figs. 2-3?
-
1989 Conohyus simorrensis goeriachensis nov. subsp.—Van der Made: pp. 27–28, pl. 1.
-
?2014 Parachleuastochoerus valentini (Filhol, 1882)—Pickford: pp. 181–185 (partially).
-
2014 Conohyus simorrensis (Lartet 1851)—Pickford and Laurent: pp. 6–9, fig. 2.
-
2014 Retroporcus matritensis nov. comb. (Golpe-Posse, 1972)—Pickford and Laurent: pp. 15–24, figs. 14–26.
-
1999 Conohyus giganteus (Falconer and Cautley, 1847)—Van der Made: 211–212 (European material).
Lectotype: MHNT PAL2011.0.84.1 right mandible with P2-M2 and PAL 2011.0.84.2 left mandible with P4-M3 of the same individual, figured by Pickford and Laurent (2014), fig. 2).
Type locality: Villefranche d’Astarac, Gers, France.
Age of the type locality: Middle Miocene (Ginsburg 1971).
Diagnosis:Conohyus with relatively large premolars and relatively small M3. M1 length about 14–21 mm, M3 length about 24–35 mm, M3 length as a percentage of M1 length about 140–200. (Adapted from Van der Made 1999).
Material:NHMW 2014/0081/0001 right and left M1–3 and left P3 of the same individual.
Description and comparison
The P3 (Fig. 11a–e) has a very simple structure. It has a single main cusp with a pointed tip. From this anterior and posterior crests, protoprecristid and protopostcristid descend towards the base of the crown. There is no clearly developed posterior cusp (hypoconid or protopostconulid). There is a bulge on the posterior part of the buccal side. This simple structure is typical of the Tetraconodontinae. Other Suidae tend to have a main cusp that is not so pointed, but that has a wider and blunter tip, especially if seen from the side. They also tend to have a cusp on the talonid (hypoconid) or a cusplet in the protopostcristid (protopostconulid). The tooth is very large (Table 3), and it is much bigger than the P3 of the different species of Parachleuastochoerus (save for a single aberrant specimen from La Grive) and of Retroporcus sindiensis, while it clusters with the P3 of Conohyus (Fig. 11). Its length must have been a little over 183% of its width.

Conohyus simorrensis from Gračanica: 1 NHMW 2014/0081/0001—left P3: a anterior, b buccal, c posterior, d occlusal, and e lingual views. Bivariate diagram comparing the width of the posterior lobe (DTp) and length (DAP) of the P3 of C. simorrensis from Puente de Vallecas, Göriach, Au, Villefranche d’Astarac, Alhambra, Paşalar, Carpetana, Elgg, Klein Hadersdorf, Kleineisenbach, Tutzing and Pitten; R. sindiensis from Mala Miliva, Somosaguas and from the Indian Subcontinent; P. steinheimensis from: Manchones I, La Grive oc, Steinheim, Can Almiral, Hostalets, Castell de Barberà, and Wissberg; P. steinheimensis olujici from Lučane; P. huenermanni from Rudabánya and Can Ponsic I; P. crusafonti from Can Llobateres. Provenance of data as indicated in Suppl.-Tab. 2
The M3 (Fig. 12 (1)) has a structure that is similar to that of the anterior molars, save for that it has a third lobe. There is one main cusp in the third lobe, the pentaconid, and it is situated a little buccally of the axis of the tooth. It is preceded by a pentapreconulid. The tooth is not very elongate and its length is 158% of its width and 153% the length of the M1. Both M3 are unworn and their crowns are low with a hypsodonty index of 56 to 68. Compared to other Conohyus samples, these M3 are among the shortest (Fig. 12).

Conohyus simorrensis from Gračanica, NHMW 2014/0081/0001: 1 left M3: a buccal, b occlusal, c lingual, and d anterior views. Change in the anterior width (DTa) and length (DAP) of the M3 of Conohyus and R. sindiensis through time—localities from old (bottom) to young (top) and R. sindiensis from the Indian Subcontinent and of different ages at the bottom. Provenance of data as in Suppl.-Tab. 2
The M1 (Fig. 13 (2)) and M2 (Fig. 13 (1)) have the same morphology, but differ in size (Table 3). They are bunodont and they had four roots. The M2 are worn, but not enough to expose the dentine and when unworn, the crowns must have been slightly higher than the values in Table 3. With a hypsodonty index (100 Ha/DTa) of 75, which must have been only a little higher, the crowns are markedly lower than wide. They are also wide (length about 122–127% of the width in the M2 and 131–132% in the M1). The length of the M2 is about 115% the length of the M1. The latter value may be much more in a late tetraconodont and in most Suinae, this is near 140%. These three proportions are typical in the primitive or early Suidae. Compared to other samples of Conohyus, these teeth are among the smallest (Fig. 13).

Conohyus simorrensis from Gračanica, NHMW 2014/0081/0001: 1 left M2: a buccal, b occlusal, c lingual, d anterior, and e posterior views; 2 left M1: occlusal view. Change in the posterior width (DTp) and length (DAP) of the M1–2 of Retroporcus and Conohyus through time—localities from old to young and R. sindiensis of different ages at the bottom. Provenance of data as in Suppl.-Tab. 2
Classification of the tetraconodont from Gračanica
The teeth from Gračanica described here belong probably to the same individual. The P3 with a high main cusp and a low talonid and a size, which is large in absolute terms (Fig. 11), but also relative to the molars, suggests that the species belongs to the Tetraconodontinae.
The suid from Gračanica does not belong to Parachleuastochoerus (= Versoporcus; Suppl.-Information) because its P3 is much larger and relatively wider (Fig. 11). In Conohyus, the P3 is large compared to the molars whereas in Parachleustochoerus, it is much smaller (Fortelius et al. 1996: fig. 28.7; Van der Made 1999: fig. 3). The P3 from Gračanica is large, relative to the molars like in Conohyus. The size of the molars from Gračanica (in particular of the M1) is comparable to the earlier stages of C. simorrensis (Figs. 11, 12, and 13, Supplementary Fig. 9). The P3 from Gračanica is longer and relatively narrower than those from Mala Miliva and Somosaguas and of R. sindiensis from Pakistan (Fig. 11). These fossils from Gračanica are assigned here to C. simorrensis. This implies that Gračanica is younger than Somosaguas (zone E, 14.05–13.75 Ma), Sansan (just over 14.163 Ma), Mala Miliva, Petrovac and Bâlâ, which have R. sindiensis (see Suppl.-Information).
The Gračanica Suoidea and biostratigraphy
Figure 14 is a further developed version of previously published figures (Van der Made 2003 fig. 9, 2005, fig. 1). It includes the three suoid lineages described here in detail, as well as some others, which are of interest here. The lineage of Hyotherium is included in Fig. 14. This lineage was described in detail by Van der Made (2010), recognising two subspecies of H. sommerringi. Recent literature on this lineage is discussed in the Suppl.-Information. The central part of Fig. 14 gives the evolutionary lineages and solid squares indicate the presence of the different chrono-species or chrono-subspecies in the localities. Open squares indicate possible presence. The localities are ordered in the same way as in Figs. 5, 7, 8, 9, 10, 12, and 13. The localities with more than one lineage indicate the positions of the grades of evolution of different species relative to each other and the relative positions of localities. Though there is still some possibility to move localities up and down in this scheme, this is limited, and the more lineages are included, the more rigid the scheme becomes. The scheme is based on biometry.

The temporal distribution of the species of Choeromorus, Bunolistriodon, Conohyus and various other selected suoid lineages. To the left the localities from the Iberian Peninsula, the local biozonation (Z to G3) and its correlation to the MN units and ATNTS2012 mainly after Daams et al. (1999b), Hernández-Ballarín and Peláez-Campomanes (2017). To the right localities from the rest of Europe and of Anatolia, MN units and Central Paratethys stages modified after Hilgen et al. (2012). Lines indicate estimated ages of localities according to various authors (see text). In some case, there are alternative ages (dashed lines) or age ranges. Solid squares indicate presence of a species in a locality and open squares indicate uncertainty (normally indicated with: cf., aff., sp. or?)
On the left of Fig. 14 are the Astronomically Tuned Neogene Time Scale (ATNTS2012) after Hilgen et al. (2012), the Spanish biozones and their ages and the proposed correlations the MN units as well as the Spanish and Portuguese localities (largely after: Daams et al. 1999a, 1999b; Hernández-Ballarín and Peláez-Campomanes 2017; Mein 2000). As indicated above, for most of the large mammal localities, only the time range of the biozone is available. This results in a long time interval for sites from a long zone like Dc, but a short time interval for sites in a short zone like F. On the right, Fig. 14 gives localities from other parts of Europe, when available with their published ages or age ranges based on palaeomagnetics, radiometric dates, etc., as well as the Central Paratethys stages, MN units, and the ATNTS2012. It should be noted that MN units have different estimated ages in Spain and Germany and Switzerland (Van der Meulen et al. 2011). Most of the localities on the right side do not have reliable and precise age constraints. The ages or age ranges of many localities are discussed in the Suppl.-Information and the localities and their published or inferred ages are listed in Suppl.-Tab. 4.
The central part of Fig. 14 provides a scheme in which localities have ages relative to many of the other localities (older/younger than). The addition of chronological information of many localities does not only provide absolute ages (or age ranges) to the scheme, but also more rigidity. This still does not provide precise ages for many localities. It is not possible to indicate graphically the possible age ranges for each site in the figure, but regarding the figure well, the age constraints of the sites can be seen. There is much more material and much better age control than when these lineages were first proposed Van der Made (1989, 1996a, 1997a, 1997b, 2003, 2005, 2010).
What strikes in Fig. 14 is that the Iberian large mammal localities are not evenly distributed in time, or at least the localities with the large mammals studied here. There are biozones without a single locality with these taxa. On the right side of the figure, this is not so apparent, but there are similar hiatuses for Germany, Austria, and possibly France as well. Different basins might have hiatuses at different times. It was noted before that the European fossil record in general is not evenly distributed (Van der Made 2011: fig. 1).
The replacement of Hyotherium by Conohyus/Retroporcus in Central and Western Europe
Conohyus simorrensis replaced Hyotherium sommerringi in the Central Paratethys (Mottl 1970; Rabeder and Steininger 1975). The appearance of Conohyus was taken as indicative of MN6 (e.g. Mein, 1975a, Mein, 1977). Later, on a European scale, there appeared to be an overlap (Van der Made 1990b; De Bruijn et al. 1992), but with improved dating, the extinction of Hyotherium in Europe appeared to be earlier (Van der Made 2010). The question, whether such a replacement existed or not, and when it may have happened, has relevance for the age of Gračanica where Conohyus is present.
At present, it appears that the latest Hyotherium record is in Rümikon (14.163–14.609 Ma) and Thannhausen (a little older than 14.55 Ma). Given the fact that Rümikon and Thannhausen have inferred ages close to or younger than 14.55 Ma, occurrences of C. simorrensis, which are younger than 14.55 Ma, would fit the model of Conohyus replacing Hyotherium. However, the new species Conohyus olojici was named and some fossils have been reclassified as Retroporcus sindiensis.
Bernor et al. (2004) described the species C. olujici from Lučane (15.0 Ma). It is tempting to believe that this represents the first Conohyus in Europe, but Van der Made et al. (2014) classified it as Parachleuastochoerus steinheimensis olujici and Pickford (2016b) as Parachleuastochoerus huenermanni.
Fossils previously included in C. simorrensis were placed in various species in the genus Retroporcus (Pickford and Laurent 2014) and are here classified as a single species R. sindiensis, which is present in Bâlâ, Petrovac, Mala Miliva, Sansan, Somosaguas, and Montjeo de la Vega (Suppl.-Tab. 2). The latter two localities are in zone E (14.05–13.75 Ma). Sansan is in the top of C5ADn (close to 14.163 Ma; see discussion in the Suppl.-Information). The specimens from Bâlâ and Petrovac seem to be isolated finds. Mala Miliva is in sediments, which interfinger with the marine Badenian, suggesting it is older than 13.8 Ma (Suppl.-Information). The faunal list is short but includes two other species of Suoidea. The specific identity of the palaeochoerid from this site has been a problem. It is probably Choeromorus lemuroides (see Suppl.-Information). The other species is Bunolistriodon latidens. The temporal ranges of these species are discussed below and are compatible with Mala Miliva being younger than Rümikon, but still within chron C5ADn (14.609–14.163 Ma).
With such an age for Mala Miliva, the model of Conohyus/Retroporcus replacing Hyotherium is possible. The dispersal of R. sindiensis into western Eurasia would then be anterior to 14.163 Ma (Sansan, top of chron C5ADn), but younger than Rümikon (chron C5ADn, 14.609–14.163 Ma) with the latest Hyotherium. Considering the historic model of a replacement of Hyotherium by Conohyus, which did not recognise Retroporcus as different from Conohyus (Mottl 1970; Rabeder and Steininger 1975), Gračanica with Conohyus should be younger than 14.069 Ma, the oldest possible date of the last Hyotherium.
The replacement of Bunolistriodonlockharti by Listriodon as a criterion for the MN5–6 transition
Gračanica has been placed in MN4–6 on the basis of rodents (Wessels et al. in press, this issue), MN5 on the basis of Rhinocerotidae (Becker and Tissier in press, this issue), MN5–6 on the basis of ruminants (Aiglstorfer and Mayda in press, this issue), beavers (Stefen in press, this issue) and chalicotheres (Coombs and Göhlich in press, this issue), and MN6 on the basis of carnivores (Bastl et al. in press, this issue). These papers do not necessarily contradict each other, but more probably reflect that different definitions of the MN units were used. The MN units were defined using many different criteria (Mein 1975a, 1975b, 1977, 1979, 1990), but it should not be surprising that the use of different criteria may give different results.
The appearance of Listriodon splendens is consistently used to separate MN6 from MN5 (Mein 1975a, 1975b, 1977, 1979, 1990). The replacement of Bunolistriodon by Listriodon is the only faunal event indicated by De Bruijn et al. (1992, tabs 8-12), which separates MN5 from MN6 in both western and central Europe. The event is thus an important one. It should be noted that at the time of these publications, B. lockharti was the only species of that genus recognised in Europe. The presence of a second European species (Van der Made and Alférez 1988) and eventually of the lineage Bunolistriodon adelli-B. latidens-B. meidamon (Van der Made 1996a) was later. Bunolistriodon lockharti was a contemporary of B. adelli and even occurred in the same localities (Córcoles) and later, B. latidens and B. meidamon were contemporaries of L. splendens and occurred in the same localities (Inönü 1, Paşalar, Çandır). By contrast, B. lockharti and L. splendens are not known from the same localities and there is no evidence that were contemporaries. It seems thus that the replacement of B. lockharti by L. splendens is a relevant event for the MN5–6 transition in western Eurasia. Listriodon is known from over 100 localities in Europe and Turkey (Van der Made 1996a), and its stratigraphic distribution can be more reliably known than of many other taxa which are used in biostratigraphy.
A similar replacement occurs broadly at the same time in China, where Bunolistriodon intermedius is replaced by L. splendens. Bunolistriodon intermedius has verrucose Cm, like B. lockharti and could be closely related or identical to that species. The Chinese Listriodon were named L. mongoliensis, L. lishanensis, L. xinanensis, and L. robustus, but there are no convincing features that show them to be different from L. splendens. I have previously listed L. robustus as a synonym of B. intermedius on the basis of published photographs (Van der Made 1996a). Liu Liping (now Stockholm) showed me the originals and indeed, these belong to a primitive Listriodon, not to Bunolistriodon. The appearance of L. splendens and the replacement of a local species of Bunolistriodon could prove to be a good criterion for the correlation of the MN5–6 transition across Eurasia from Spain and Portugal to China.
Castelnau d’Arbieu has B. lockharti and is one level lower in the local stratigraphic sequence than Sansan (about 14.163 Ma) with L. splendens. Other late B. lockharti are from Lisbon Vb (later half of zone Dc,15.2–14.75 Ma) and Ravensburg. The latter locality is in the Upper Fresh Water Molasse (OSM) and is probably latest MN5, but no precise age is known. A molar from Georgensgmünd (NMB) was assigned to Bunolistriodon sp. (Van der Made 1996a), but more material (MNB) belongs clearly to B. lockharti. Berger (2010, 2011) described and figured all presently known material and assigned it to that species. Georgensgmünd is in the OSM. Its age is controversial (before/after the Ries Event), but probably is slightly less than 15 Ma (see Suppl.-Information).
From Sansan (top C5ADn, a little older than 14.163 Ma) and Inönü 1 onwards, there is a dense record of Listriodon in Europe. A fragmentary incisor from Rümikon (14.163–14.609 Ma) (NMB OSM1080) belongs to a listriodont, but its identification is insecure. A cast from Stätzling (BSP) was assigned to Listriodon (Van der Made 1996a). This locality is similar in age to Thannhausen (close to, but older than, 14.55 Ma).
The replacement of B. lockharti by L. splendens seems to have occurred between about 15 and 14.55 Ma. This is in either in zone Dc or Dd and well before zone F, which in Spain has the first record of L. splendens. Puente de Vallecas is probably the only Iberian record of Bunolistriodon in zones Dd and E and was previously assigned to B. lockharti (Morales and Soria 1985). Even though these fossils were later assigned to B. latidens, the first appearance of Listriodon at 13.75 Ma in zone F was not questioned (Van der Made 1996a). Daams et al. (1998, fig. 1) indicated “Bunodont Listriodon” in zones C to E and “Lophodont Listriodon” in zones F to G1 and correlated the E-F transition to that of MN5–6. It seems now that the replacement of B. lockharti by L. splendens is a much older event between about 15 and 14.55 Ma. A correlation of Sansan to just below the top of C5ADn at 14.163 Ma dates Sansan within the time range of zone Dd (14.75–14.05 Ma). The position of Sansan is thus well above the MN5–6 transition, if this is taken at the replacement of B. lockharti by L. splendens. The following section will provide arguments that Gračanica is younger than the appearance of Listriodon and, based on this criterion, should be placed in MN6.
Faunal events and the age of Gračanica
A series of faunal events, both older and younger than Gračanica, are of biostratigraphic interest and help to date the locality (Fig. 14 and Suppl.-Fig. 10).
The first is the evolution of Choeromorus primus to Ch. lemuroides (Suppl.-Information). Since Gračanica has Ch. lemuroides; this event puts a maximum age on the locality. The evolution occurred after Els Casots (zone C, 16.45–15.98 Ma). The earliest Ch. lemuroides might be from Lavardens, in a level between those of La Romieu and Castelnau d’Arbieu. None of these sites have independent age estimates.
The second is the evolution of B. adelli to B. latidens (Suppl.-Information). Bunolistriodon adelli is well known from the Iberian Peninsula, but might be present in Chios, and B. latidens is mainly known from Serbia and Turkey, though there are some records in the Iberian Peninsula. Bunolistriodon adelli is present in various localities of zone C and Dc (15.47–14.75 Ma) and evolved into B. latidens present in Lisbon Vb (Olival da Susana, Quinta do Farinheira, etc.; zone Dc, 15.47–14.75 Ma; Fig. 14). The latter identification is mainly based on one I1 from Olival Susana, but it is wider than any B. adelli I1 (Suppl.-Fig. 8). Given the number of sites with B. adelli in zone Dc, this evolution may have happened closer to 14.75 than to the 15.47 Ma. This further limits the possible age of Gračanica.
The third event is the replacement of Bunolistriodon lockharti by L. splendens, which was discussed in the previous section. In Europe and Anatolia, it seems to have happened between about 15 and 14.55 Ma and a similar event occurred in China. It is of relevance, since it is one of the criterions used to define the transition of MN5 to MN6. Gračanica is younger than this event (see below).
The fourth event is the replacement of Hyotherium by a Retroporcus, which is discussed in a previous section. This event happened within chron C5Adn (14.609–14.163 Ma).
The fifth event is the evolution of Retroporcus to Conohyus (Suppl.-Information). The youngest localities with Retroporcus and age control are Sansan (just over 14.163 Ma) and Montejo de la Vega and Somosaguas (zone E, 14.05–13.75 Ma). One of the oldest localities with Conohyus is Puente de Vallecas (zone E). This suggests that this transition occurred within zone E and between 14.05 and 13.75 Ma. A P4 from Sankt Oswald is generally assigned to C. simorrensis and is longer and more elongate (measurements: Van der Made 1998, Tab. 7) than in R. sindiensis. The site is said to be in sediments that interfinger with marine sediments of the “Lagenidenzone” (Rabeder 1978), which would imply an age older than 13.9 Ma (Mandic et al. 2019, fig. 10). This suggests that the replacement of Retroporcus by Conohyus happened between 14.05 and 13.9 Ma. Since Gračanica has Conohyus, this indicates a maximum age of 14.05 Ma for this site.
The sixth event is the evolution of Ch. lemuroides to Ch. inonuensis (Suppl.-Information). The fifth and sixth events bracket Gračanica, which has already Conohyus, but retains Ch. lemuroides. The event must be younger than Sansan (slightly older than 14.163 Ma), because that locality still has Ch. lemuroides. None of the localities with Ch. inonuensis has independent age control, so the only thing that can be said is that this event must be older than the next faunal event.
The seventh event is the evolution of B. latidens to B. meidamon (Suppl.-Information). The B. latidens I1 from Gračanica is outside the range of the older samples of the species and comparable to Prebreza, a late, possibly the latest, locality with this species, suggesting that Gračanica was relatively close to this faunal event. The latest B. latidens with an independent indication of numerical age is from Carpetana (zone F, 13.75–13.60 Ma). This identification is based on only one incisor, but it is clearly outside the size range of the large sample from Paşalar (Suppl.-Fig. 8). The oldest locality with B. meidamon is Paşalar, but which lacks independent age control.
The eighth event is the replacement of Ch. inonuensis by Ch. grandaevus. This must have happened between Paşalar, without independent age control, and Steinheim with Megacricetodon gregarius, which in the Swiss molasse is restricted to the top of C5ACn (Kälin and Kempf 2009). Therefore, the event must have happened before 13.739Ma. Gračanica should be clearly older than this date.
The ninth event is the evolution of B. m. meidamon to B. m. ultimus. This happened between Paşalar, without independent age control, and Çandır. The latter locality has a palaeomagnetic section with four possible correlations to the global scale (Suppl.-Information). The two most likely are to C5ACn (13.739–14.070 Ma) or to C5ABn (13.363–13.608 Ma). A correlation of Çandır to C5ACn would concentrate the last three faunal events in the top of C5Acn and the base of zone F (13.75–13.739 Ma). It seems more likely that Çandır should be correlated to C5ABn.
Despite the fact that the localities are from an area extending from Lisbon to central Anatolia, and despite the known problems of correlation with MN units, all evolutionary events in these lineages are consistent with the known ages or age ranges of the localities. There are, however, some remarkable implications. For instance, Sansan is not to be correlated to zone F, as previously believed, but more likely to the top of zone Dd. This follows from the age estimates from zone D and Sansan being placed in the top of chron C5Adn, but now, this is also supported by biostratigraphy. The combination of various lineages provides more precise and robust age estimates. For instance, Petersbuch 39 and 108 date probably from between 14.163 (Sansan, where Ch. inonuensis is not yet present) and 13.363 Ma (the youngest possible age of Çandır, where Ch. inonuensis is replaced by Ch. grandaevus). The estimated age ranges for each of the localities, for which no previous age estimate was available, and for each of the faunal events, are testable hypotheses.
Gračanica is bracketed between events 1–5 and 6–9 (after 14.05 and well before 13.739 Ma). The use of multiple events to date Gračanica implies enhanced support for its estimated age.
Improved biochronology and the biogeography and evolution of Bunolistriodon
The anagenetic lineage B. adelli-latidens-meidamon was described and the origin of this lineage was discussed over 20 years ago (Van der Made 1996a), but it was not resolved whether B. lockharti and B. adelli appeared in Europe by dispersal at the same time, at different times, or whether one evolved from the other within Europe. The increased biochronologic resolution, as synthesized in Suppl.-Fig. 11, is relevant for the understanding and dating of the biogeography and evolution of Bunolistriodon.
In the Iberian Peninsula, the first localities with Bunolistriodon are in zone Ca (after 16.45 Ma), but in Germany, this is in Grimmelfingen (17.6–17.2 Ma), Langenau (17.2–16.8 Ma), Gerlenhofen (16.8–16.6 Ma), and Engelswies (16.6 Ma), which are all older. At the first sight, the same happens as with several micro mammal events, which are used in biostratigraphy (Van der Meulen et al. 2011, 2012): the taxa appear later in Spain than in the Molasse Basin. However, there are no large mammal localities in the Iberian Peninsula placed in zone B and those of zone A are much rarer than those of zones C and D. It is possible that this later appearance of Bunolistriodon in the Iberian Peninsula is apparent. This can only be solved when more large mammal localities of these ages are found. Thus, it is not clear whether the later appearance of Bunolistriodon in the Iberian Peninsula is real or an artifact.
While the oldest European Bunolistriodon localities have all B. lockharti, nearly all B. adelli localities are from the Iberian Peninsula. It was cited from two localities in France and Spain: Echzell and Bézian (Van der Made 1996a). Bunolistriodon michali (Paraskevaidis 1940) is a small species, of which the incisors and canines are not known. It was considered to be a possible synonym of B. latidens (Fortelius et al. 1996). More recent age estimates for Chios (Koufos 2006) suggest that it is older than B. latidens and that therefore it might rather be a synonym of B. adelli. The oldest B. adelli from zone C are more similar to B. lockharti than those of zone D. The available data suggest the following scenario (Suppl.-Fig. 11):
-
1.
B. lockharti dispersed into Western Europe (17.6–17.2 Ma).
-
2.
Not later than early zone Ca (16.45 Ma), it spread into the Iberian Peninsula.
-
3.
B. adelli evolved there from B. lockharti (later in zone Ca, before 16.3 Ma). Bunolistriodon adelli inherited its three-lobed I1 from B. lockharti, even though in the oldest samples, the tripartite division is still variable and mainly visible near the tip of the tooth in little worn molars.
-
4.
B. lockharti dispersed again into the Iberian Peninsula (zone Cb, 16.3–15.98) and B. adelli dispersed into Europe reaching Chios (not later than 15.974–15.16 Ma). The two species were sympatric.
-
5.
B. adelli evolved into B. latidens (zone Dc, 15.47–14.75 Ma).
-
6.
B. lockharti went extinct, when L. splendens dispersed into Europe (about 15–14.55 Ma).
-
7.
In the Balkans and Anatolia, B. latidens gave rise to B. meidamon (after the onset of zone F, 13.75 Ma).
-
8.
The lineage went extinct before 13.363 Ma.
Ecology of the Suoidea of Gračanica
With three species, the Suoidea are prominent in the fauna of Gračanica. There are at least two individuals of Bunolistriodon latidens, while the other species are represented each with at least one individual.
The most important feature of the Suoidea is their adaptation to rooting. This is a unique adaptation and provides the Suoidea with access to food sources that are not used by other ungulates. They are omnivores and target animals that live in the soil or hide underground (invertebrates and small vertebrates) as well as nutrient-rich underground plant parts (roots, tubercles, etc.). Depending on the climates in which they live, rooting provides the Suoidea access to food during dry or cold seasons, when other food sources are restricted (e.g. African wart hogs in the dry season; Cumming 1975). Suoidea have a variety of rooting styles, some root to an average depth of 8 cm and others to 25 cm, and adaptations to rooting, which can be seen in the fossils (Ewer 1958; Sicuro and Oliveira 2002). When Suoidea root, they push their snout into the soil and subsequently elevate the snout. Species that root deep have wide occiputs, for the insertion of powerful neck muscles, which are high above the occipital condyles, so that the muscles have a greater momentum. Long narrow snouts are another adaptation. Species with large canines probably do not root very deep, as is the case with the living African wart hog. When the snout is in the soil, the snout disc is moved for searching food items. This disc is moved with muscles, of which the size and power can be judged from the origin on the skull. The incisors are used in digging and in the extraction of food items. Narrow high-crowned incisors are an adaptation to rooting. The lingual crests in the lower incisors are another feature, which probably is related to rooting, providing grip when wrenching roots. In many species, the I3 does not evolve a higher crown, while related to this, the I2 becomes elongated, occluding with the lateral side of the I2 (Van der Made 2010).
All Suidae of which the skulls are known have wider and higher occiputs than the Palaeochoeridae (“Old World peccaries”) and the Tayassuidae (American peccaries). The highest and widest occiputs appear from the Late Miocene onwards. The skull of Conohyus simorrensis is not known, but the skull of its ancestor Retroporcus sindiensis has already a clearly more elevated and wider occiput than in the peccaries, while its snout was narrow (Colbert 1938). Its incisors have high, but not extremely high, crowns and the I2 is moderately elongated. Its I1–2 have clear lingual crests. Conohyus simorrensis was clearly adapted to rooting and presumably targeted deeper food items than the living peccaries. It may have had a broadly similar level of adaptation as Hyotherium, while later Suidae improved on that. Conohyus is far less abundant in the Middle Miocene of Europe, than its relative Parachleuastochoerus. It tends to be represented by few specimens in the localities where it occurs. An exception is Göriach, where it is very common. The site is in coal, suggesting possibly dense vegetation or forest and certainly a humid soil with a thick organic layer. Such a soil would be rich in food items for Conohyus. Also, the fossil locality of Gračanica is in coal and probably the environment had optimal conditions for Conohyus.
All known skulls of the Palaeochoeridae have narrow and low occiputs. There is no indication for powerful snout muscles on the face. Their incisors tend to be low crowned and in Choeromorus, they lack lingual crests (Suppl.-Fig. 5). The I2 is not elongated. This suggests that they are no powerful rooters. However, the Palaeochoeridae have elbows, which allow some rotation, which is rare in the Artiodactyla, and third phalanges, which suggest the existence of powerful nails or claws, different from the typical hoofs of other Suoidea. These and other features, such as a long pisiform, suggest that they may have dug with their feet. Living species of Suoidea are known to dig occasionally with their forefeet (Cumming 1975). The localities where the Palaeochoeridae are most abundant tend to be fissure fillings: Coderet, Tomerdingen, various sites at Petersbuch, and also La Grive. The fissures are in limestone. Landscapes with limestone tend to have soils that are not very deep. Choeromorus evolved towards lophodonty, without fully reaching this state. Lophodonty in Suoidea is taken as an adaptation to folivory. All this suggests that Choeromorus lemuroides may have specialised more on folivory than other Suoidea and that it depended relatively less on rooting.
Bunolistriodon latidens evolved very wide incisors. The Listriodontinae in general show the trend to decrease the crown height and increase the width of the incisors. The lingual crests on the I1–2 tend to become less prominent. They also lack lingual wear facets on these incisors. This has been interpreted as an evolution away from the rooting habit (Leinders 1977; Van der Made 1996a). The Listriodontinae evolved towards lophodonty and the species of the lineage of B. latidens reached a stage, which is called sublophodont. Morphology and micro wear suggests that the listriodonts specialised in folivory and the wide incisors have been interpreted to be a specialisation to bulk feeding, perhaps on herbs (Leinders 1977; Hunter and Fortelius 1994; Van der Made 1996a). If this species abandoned rooting and was a specialised folivore, maybe also frugivore, there must have been abundant edible leaves all year round. This suggests a lesser degree of seasonality than today, or at least a lesser impact of the seasons on the vegetation. Another feature, which suggests that rooting was not important to B. latidens and B. meidamon, is the gigantic canines of the males, which had lengths of over 30 cm (Van der Made 1996a, Pl. 18 fig. 1, Pl. 37 fig. 6): these would not allow deep rooting. While the lower canines have sharp edges and could have been used as weapons, especially the upper canines are large and would be well visible from a distance. The upper canines had a display function and served to impress competing males. Bovidae use horns to fight and to display. The smaller horns occur in species, which are territorial and which live in pairs in closed habitats, while the largest and most ornamented horns occur in species, which live in more open environments and in which a male defends a territory and attempts to retain many females (“harem”) in this territory, while in non territorial species, there are large mixed herds and also the females tend to have large horns (Jarman 1974; Estes 1974). In Suidae, this has a parallel in that most living species are territorial, live in pairs, and the males have relatively small upper canines, while in the African warthog (Phacochoerus), both males and females have large canines, the females live in groups, and one male defends his position near the group of females. Based on the canine as a display structure, the social structure of B. meidamon has been interpreted as males living with large groups of females in environments where these canines are visible from far (Van der Made 2003). Bunolistriodon latidens may have been folivorous, feeding on large quantities of herbs in a landscape with open spaces.
The rate of evolutionary increase in incisor width in the Bunolistriodon adelli-latidens-meidamon lineage was not constant, but increased greatly after 13.75 Ma. This is within a period of major climatic change, called the Mid-Miocene Crisis (Krijgsman et al. 1994). Probably, these morphological changes reflect the intake of larger quantities of less nutritive food and are related to environmental change. At this time, the geographical distribution of the lineage became progressively restricted ending in Anatolia. This may reflect a response to the Mid-Miocene cooling, perhaps causing greater impact of seasonal fluctuations on the vegetation, first at higher latitudes, later at lower latitudes. The presence of B. latidens in Gračanica suggests that this locality shared aspects of its environment (i.e., those to which this listriodont adapted) with Anatolia, but not with western and central Europe. During MN6, the environment in which B. latidens-meidamon lived progressively disappeared from Europe.
Two of the three species of Suoidea from Gračanica seem to have been more specialised in folivory (perhaps also frugivory) rather than in rooting. This suggests the presence during all or most of the year of abundant herbs or other low vegetation, either as an under storey in a forest or in open spaces. The impact of the seasons on the vegetation may have been less than today, which is compatible with a higher inferred temperature, based on insects (Wedmann and Skartveit in press, this issue). The canines of Bunolistriodon suggest open spaces, while Conohyus is more suggestive of closed environments, or at least soils with a thick horizon of organic material, which in turn suggests humid conditions. This is not strange, given the presence of coal and beavers at Gračanica (Stefen in press, this issue). This interpretation is compatible with others of the environment of Gračanica having habitats ranging from swampy forest to drier and more open environments (Harzhauser et al. in press, this issue; Becker and Tissier in press, this issue; Coombs and Göhlich in press, this issue).
Conclusions
The study of the Suoidea from Gračanica led to the following conclusions:
-
The species Choeromorus lemuroides (Taucanaminae, Palaeochoeridae), Bunolistriodon latidens (Listriodontinae, Suidae), and Conohyus simorrensis (Tetraconodontinae, Suidae) have been identified.
-
The Suoidea from Gračanica belong to anagenetic lineages, which are known from Europe and Anatolia. Many new fossils of these lineages have become available since these lineages were first described and dating of these materials has improved. The new data support these lineages.
-
A biostratigraphic scheme is presented based on the evolutionary stages of these three lineages and some other Suoidea. The proposed correlations across Europe and Anatolia appear to be consistent, despite known provincialism in rodents, which are widely used in used for biostratigraphy.
-
The Suoidea suggest an age for Gračanica between 14.05 and 13.739 Ma and probably more towards the middle of this period
-
The ages of many other localities can be estimated more precisely. Among these, of particular interest are the hominoid localities Çandır (13.608–13.363 Ma) and Paşalar (13.75–13.739 Ma) and the locality Mala Miliva (14.609–14.163 Ma).
-
The Suoidea suggest the presence of abundant vegetation, possibly forest, but also of open spaces at Gračanica. Seasonality may have had less impact on the vegetation than today. The soil may have been humid with a thick layer of organic material.
-
A revised phylogeny of the Taucanaminae is presented.
-
The early history of Bunolistriodon in Europe was not clear. The available information suggest that Buno-listriodon lockharti dispersed first into central Europe and later into the Iberian Peninsula, where it may have given rise to Bunolistriodon adelli (= Bunolistriodon michali). The two species became sympatric in an area stretching from Portugal to Greece.
-
The evolution in the Bunolistriodon adelli-latidens-meidamon lineage was not constant and the rate of evolution increased greatly around 13.75–13.739 Ma, which reflects a trend to bulkfeeding and is probably related to climatic change.
-
Retroporcus sindiensis dispersed into Anatolia and Europe, which may have happened between 14.609 and 14.163 Ma. Between 14.05 and 13.75 Ma, it gave rise to Conohyus simorrensis.
-
The Suoidea from Gračanica belong to groups which have suffered taxonomic inflation, taxa have been named on too little evidence and disregarding the rules of the ICZN (1999). The cases, which affect the classification of the Suoidea from Gračanica, are discussed in the Suppl.-Information.
-
Choeromorus lemuroides is not classified as Choeromorinae and Siderochoeridae, nor as Yunnanochoeridae, because of articles 36.1 and 40.1 of the ICZN (1999), the Choeromorinae is a junior synonym of Taucanaminae and Siderochoeridae and Yunnanochoeridae would be synonyms of Taucanamidae, if such a family would be recognised.
-
Choeromorus lemuroides is classified as Palaeochoeridae, because Palaeochoerus does not belong to the Suidae (as has been claimed) and the Palaeochoeridae, Schizoporcidae, Siderochoeridae, and Doliochoeridae share many apomorphies and are here retained in a single family, the Palaeochoeridae.
-
The genus Choeromorus and its likely direct ancestor Siderochoerus minimus share a clear apomorphy and as a result, Siderochoerus is included in Choeromorus. Several other synonymies concerning genera and species are discussed.
-
The species Retroporcus complutensis is a synonym of R. sindiensis.
-
The designation of a lectotype for C. simorrensis by Pickford and Laurent (2014) may not be valid, but changed the definition of the species.
References
Aiglstorfer, M. & Mayda, S. (in press). Ruminantia from the middle Miocene of the Gračanica coalmine (Bugojno Basin, Bosnia-Herzegovina). In U.B. Göhlich, & O. Mandic (Eds.) The drowning swamp of Gračanica (Bosnia-Herzegovina) - a diversity hotspot from the middle Miocene in the Bugojno Basin. Palaeobiodiversity and Palaeoenvironments. https://doi.org/10.1007/s12549-018-0354-z. [this issue]
Arambourg, C. (1933). Mammifères Miocènes du Turkana (Afrique orientale). Annales de Paleontologie, 22, 123–146 2 plates.
Arambourg, C. (1963). Le genre Bunolistriodon Arambourg 1933. Bulletin de la Société géologique de France, Série, 7, 903–911 plate 24.
Azanza Asensio, B. (1986). Estudio geologico y Palaeontológico del Mioceno del sector oeste de la comarca de Borja. Cuadernos de Estudios Borjanos, 17–18, 63–126.
Bastl, K., Nagel, D., Morlo, M., & Göhlich, U.B. (in press). The Carnivora (Mammalia) from the middle Miocene locality of Gračanica (Bugojno Basin, Gornji Vakuf, Bosnia and Herzegovina). In U.B. Göhlich, & O. Mandic (Eds.) The drowning swamp of Gračanica (Bosnia-Herzegovina) - a diversity hotspot from the middle Miocene in the Bugojno Basin. Palaeobiodiversity and Palaeoenvironments. https://doi.org/10.1007/s12549-018-0353-0. [this issue]
Becker, D., & Tissier, J. (in press). Rhinocerotidae from the early middle Miocene locality Gračanica (Bugojno Basin, Bosnia-Herzegovina). In U.B. Göhlich, & O. Mandic (Eds.) The drowning swamp of Gračanica (Bosnia-Herzegovina) - a diversity hotspot from the middle Miocene in the Bugojno Basin. Palaeobiodiversity and Palaeoenvironments. https://doi.org/10.1007/s12549-018-0352-1. [this issue]
Berger, G. (2010). Die miozäne Flora und Fauna (MN5) der historischen Fossil-Lagerstätte Georgensgmünd (M.fr.) (pp. 1–191). Nürnberg: Naturhistorische Gesellschaft.
Berger, G. (2011). Weitere Schnecken- und Säugetierfunde von der untermiozänen (MN5) Fossil-Lagerstätte Georgensgmünd (Mfr.). Natur und Mensch, Jahresemitteilungen, 2010, 91–98.
Bernor, R. L., Bi, S., & Radovcic, J. (2004). A contribution to the evolutionary biology of Conohyus olujici n. sp. (Mammalia, Suidae, Tetraconodontinae) from the early Miocene of Lučane, Croatia. Geodiversitas, 26(3), 509–534.
Biedermann, W.G.A. (1873). Petrefacten aus der Umgegend von Winterthur 4. Reste aus Veltheim (pp. 1–16, 9 plates). Winterthur: J. Westfehling.
Blainville, H.M.D. de (1847). Ostéographie ou description iconographique comparée du squelette et du système dentaire de mammifères récents et fossiles pour servir de base à la zoologie et à la géologie. Vol. 4, sur les hippopotames et les cochons. (pp. 248 pp., 93 pls.) Paris: Bertrand.
Bruijn, H. de, Daams, R., Daxner-Höck, G., Fahlbusch, V., Ginsburg, L., Mein, P., Morales, J., Heizmann, E., Mayhew, D. F., Van der Meulen, A. J., Schmidt-Kittler, N., & Telles Antunes, M. (1992). Report of the RCMNS working group on fossil mammals, Reisensburg 1990. Newsletters on Stratigraphy, 26(2/3), 65–118.
Colbert, E. H. (1938). Fossil mammals from Burma in the American Museum of Natural History. Bulletin of the American Museum of Natural History, 74, 255–436.
Coombs, M. & Göhlich, U.B. (in press). Anisodon (Perissodactyla, Chalicotheriinae) from the middle Miocene locality Gračanica (Bugojno Basin, Gornji Vakuf, Bosnia and Herzegovina). In U.B. Göhlich, & O. Mandic (Eds.) The drowning swamp of Gračanica (Bosnia-Herzegovina) - a diversity hotspot from the middle Miocene in the Bugojno Basin. Palaeobiodiversity and Palaeoenvironments. https://doi.org/10.1007/s12549-018-0357-9. [this issue]
Cumming, D.H.M. (1975). A field study of the ecology & behaviour of warthog. Trustees of the National Museums and Monuments of Rhodesia, Museum memoir, 7. Salisbury.
Daams, R., Alcalá, L., Álvarez Sierra, M. A., Azanza, B., van Dam, J. A., Van der Meulen, A. J., Morales, J., Nieto, M., Peláez-Campomanes, P., & Soria, D. (1998). A stratigraphical framework for Miocene (MN4-MN13) continental sediments of Central Spain. Comptes Rendus de l’Académie des Sciences de la terre et des planètes, 327, 625–631.
Daams, R., Meulen, A. J. Van der, Alvarez Sierra, M. A., Peláez-Campomanes, P., Calvo, J. P., Alonso Zarza, M. A., & Krijgsman, W. (1999a). Stratigraphy and sedimentology of the Aragonian (Early to Middle Miocene) in its type area (North-Central Spain). Newsletters in Stratigraphy, 37(3), 103–139.
Daams, R., Meulen, A. J. Van der, Alvarez Sierra, M. A., Peláez-Campomanes, P., & Krijgsman, W. (1999b). Aragonian stratigraphy reconsidered, and a re-evaluation of the middle Miocene mammal biochronology in Europe. Earth and Planetary Science Letters, 165, 287–294.
Estes, R.D. (1974). Social organizationof the Afrian Bovidae. Calgary Symposium, 166–205.
Ewer, R. F. (1958). Adaptive features in the skulls of African Suidae. Proceedings of the Zoological Society of London, 131, 135–155.
Fahlbusch, V. (1991). The meaning of MN-zonation: considerations for a subdividion of the European continental Tertiary using Mammals. Newsletters on Stratigraphy, 24(3), 159–173.
Falconer, H., & Cautley, T. (1847). Fauna Antiqua Sivalensis. London, 1846–1849.
Filhol, H. (1881). Note relative une nouvelle èspece de Sus fossile trouvée dans Ies argiles à Dinotherium de Valentine (Haule-Garonne). Bulletin de la Societe philomathique de Paris, 7th series, 5(1), 123–124.
Fortelius, M., Made, J. van der, & Bernor, R. L. (1996). Middle and Late Miocene Suoidea of Central Europe and the Eastern Mediterranean: evolution, biogeography and paleoecology. In R. L. Bernor, V. Fahlbusch, & S. Rietschel (Eds.) The evolution of Western Eurasian Neogene Mammal Faunas (pp. 344–377). New York: Columbia University Press.
Gervais, P. (1848–1852). Zoologie et Paléontologie françaises (animaux vertébrés) ounouvelles recherches sur les animaux vivants et fossiles de la France (Vol. 1–3). Paris: Bertrand.
Gervais, P. (1859). Zoologie et Paléontologie françaises (2nd ed.pp. 1–544). Paris: Bertrand.
Ginsburg, L. (1971). Les faunes de Mammifères burdigaliens et vindoboniens des bassins de la Loire et de la Garonne. Mémoires du BRGM, 78(1), 153–167.
Golpe Posse, J. M. (1972). Suiformes del Terciario Español y sus yacimientos. Paleontología y Evolución, 2, 1–197 7 plates.
Gray, J. E. (1821). On the natural arrangement of vertebrose animals. London Medical Repository, 15(1), 296–310.
Harzhauser, M., Mandic, O., Nordsieck, H., & Neubauer, T.A. (in press). A new Helicidae (Gastropoda) from the Middle Miocene of Bosnia and Herzegovina, with a revision of the genus Paradrobacia. In U.B. Göhlich, & O. Mandic (Eds.) The drowning swamp of Gračanica (Bosnia-Herzegovina) - a diversity hotspot from the middle Miocene in the Bugojno Basin. Palaeobiodiversity and Palaeoenvironments. https://doi.org/10.1007/s12549-018-0344-1. [this issue]
Hernández-Ballarín, V., & Peláez-Campomanes, P. (2017). Updated biochronology of the Madrid Basin: species distribution and implications for the Middle Miocene rodent faunas of Spain. Historical Biology. https://doi.org/10.1080/08912963.2016.1276580.
Hilgen, F.J., Lourens, L.J., van Dam, J.A., Beu, A.G., Boyes, A.F., Cooper, R.A., Krijgsman, W., Ogg, J.G., Piller, W.E., & Wilson, D.S. (2012). The neogene period. In Gradstein, F.M., Ogg, J. G., Schmitz, M. D.& Ogg, G.(Eds.) The geologic time scale, Elsevier, (pp. 923–978)
Holt, B. G., Lessard, J. P., Borregaard, M. K., Fritz, S. A., Araújo, M. B., Dimitrov, D., Fabre, P. H., Graham, C. H., Graves, G. R., Jønsson, K. A., Nogués-Bravo, D., Wang, Z., Whittaker, R. J., Fjeldså, J., & Rahbek, C. (2013). An update of Wallace’s zoogeographic regions of the world. Science, 339, 74–78.
Hunter, J. P., & Fortelius, M. (1994). Comparative dental occlusal morphology, facet development, and microwear in two sympatric species Listriodon (Mammalia, Suidae) from the Middle Miocene of western Anatolia (Turkey). Journal of Vertebrate Paleontology, 14(1), 105–126.
ICZN. (1999). International Code on Zoological Nomenclature (4th ed.pp. 1–306). London: International Trust for Zoological Nomenclature.
Jarman, P. J. (1974). The social organisation of antelpe in relation to their ecology. Behaviour, 58, 215–267.
Kälin, D., & Kempf, O. (2009). High-resolution stratigraphy from the continental record of the Middle Miocene Northern Alpine Foreland Basin of Switzerland. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 254(1–2), 177–235.
Kaup, J. J. (1859). Beiträge zur näheren Kenntnis der urweltlichen Säugethiere 4 (pp 1–31). Darmstadt: J. G. Heyer.
Koufos, G. D. (2006). The Neogene mammal localities of Greece: faunas, chronology and biostratigraphy. Hellenic Journal of Geosciences, 41(1), 183–214.
Krijgsman, W., Langereis, C. G., Daams, R., & Meulen, A. J. Van der (1994). Magnetostratigraphic dating of the middle Miocene climate change in the continental deposits of the Aragonian type area in the Calatayud-Teruel basin (Central Spain). Earth and Planetary Science Letters, 128, 513–526.
Lartet, E. (1851). Notice sur la Colline de Sansan, suivi d'une Récapitulation des diverses espèces d'animaux vertébrés fossiles, trouvés soit à Sansan dans d'autres Gisements du terrain teritiare miocène dans le bassin sous-pyrénéen (pp. 1–45, pl. 1). Auch: J.A. Portes.
Leinders, J. J. M. (1977). A functional interpretation of the dental morphology of Listriodon (Suina). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, Series B, 80, 61–69.
Lydekker, R. (1876). Fossil mammalian faunae of Indian and Burma. Records of the Geological Survey of India, 9, 86–106.
Made, J. van der (1989). A Conohyus-lineage (Suidae, Artiodactyla) from the Miocene of Europe. Revista Espanola de Paleontologia, 4, 19–28.
Made, J. van der (1990a). Iberian Suoidea. Paleontologia I Evolució, 23, 83–97.
Made, J. van der (1990b). A range chart for European Suidae and Tayassuidae. Paleontologia I Evolució, 23, 99–104.
Made, J. van der (1996a). Listriodontinae (Suidae, Mammalia), their evolution, systematics and distribution in time and space. Contributions to Tertiary and Quaternary Geology, 33(1–4), 3–254 19 tables on microfiche: 1–54.
Made, J. van der (1996b). Albanohyus, a small Miocene pig. Acta Zoologica Cracoviense, 38(1), 293–303.
Made, J. van der (1997a). Systematics and stratigraphy of the genera Taucanamo and Schizochoerus and a classification of the Palaeochoeridae (Suoidea, Mammalia). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, 100(1–2), 127–139.
Made, J. van der (1997b). On Bunolistriodon (=Eurolistriodon) and kubanochoeres. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, 100(1–2), 141–160.
Made, J. van der (1998). Vertebrates from the open-cast mine Oberdorf (Western Styria Basin, Austria). Mammalia 8: Rhinocerotidae. Annalen des Naturhistorisches Museum in Wien, 99A, 163–168.
Made, J. van der (1999). Biometric trends in the Tetraconodontinae, a subfamily of pigs. Transactions of the Royal Society of Edinburgh: Earth Sciences, 89, 199–225.
Made, J. van der (1999b). Intercontinental relationship Europe-Africa and the Indian Subcontintent. In G. Rössner & K. Heissig (Eds.) The Miocene land mammals of Europe (pp. 457–472). München: Verlag Dr. Friedrich Pfeil.
Made, J. van der (2003). Suoidea (pigs) from the hominoid locality of Çandır in Turkey. Courier Forschungsinstitut Senckenberg, 240, 149–179.
Made, J. van der (2005). Errata and reply to guest editor’s notes. Courier Forschungsinstitut Senckenberg, 254, 473–477.
Made, J. van der (2010). The pigs and “Old World peccaries” (Suidae and Palaeochoeridae, Suoidea, Artiodactyla) from the Miocene of Sandelzhausen (southern Germany): phylogeny and an updated classification of the Hyotheriinae and Palaeochoeridae. Paläontologische Zeitschrift, 84, 43–121.
Made, J. van der (2011). Biogeography and climatic change as a context to human dispersal out of Africa and within Eurasia. Quaternary Science Reviews, 30, 1353–1367.
Made, J. van der, & Alférez, F. (1988). Dos suidos (Listriodontinae) del Mioceno Inferior de Córcoles (Guadalajara, España). In Coloquio homenaje a Rafael Adrover: “Bioeventos y Sucesiones faunisticas en el Terciario continental Iberico”. Sabadell 14–16 dic. 1988. Resumenes. Sabadell.
Made, J. van der, & Mateos, A. (2010). Longstanding biogeographic patterns and the dispersal of early Homo out of Africa and into Europe. Quaternary International, 223–224, 195–200.
Made, J. van der, Morales, J., & Montoya, P. (2006). Late Miocene turnover in the Spanish mammal record in a wider context. Palaeogeography, Palaeoclimatology, Palaeoecology, 238, 228–246.
Made, J. van der, Prieto, J., Aiglstorfer, M., Böhme, M., & Gross, M. (2014). Taxonomic study of the pigs (Suidae, Mammalia) from the late Middle Miocene of Gratkorn (Austria, Styria). In M. Böhme, M. Gross, J. Prieto (Eds.) The Sarmatian vertebrate locality Gratkorn, Styrian Basin. Palaeobiodiversity and Palaeoenvironments, 94(1), 595–617.
Mandic, O., Sant, K., Kallanxhi, M. E., Ćorić, S., Theobalt, D., Grunert, P., de Leeuw, A., & Krijgsman, W. (2019). Integrated bio-magnetostratigraphy of the Badenian reference section Ugljevik in southern Pannonian Basin - implications for the Paratethys history (middle Miocene, Central Europe). Global and Planetary Change, 172, 374–395.
Matthew, W. D. (1924). Third contribution to the Snake Creek fauna. Bulletin of the American Museum of Natural History, 50, 59–210.
Mein, P. (1975a). Résultats du groupe de travail des Vertébrés. IUGS Commission on Stratigraphy, subcommision on Neogene Strat. Region. Comm. Mediterr., 78–81.
Mein, P. (1975b). Proposition de biozonation du Néogène méditerranéen à partir des mammifères. Trabajos sobre Neógeno-Cuaternario, 4, 112.
Mein, P. (1977). Tables 1–3. In M. T. Alberdi, & E. Aguirre, Round table on mastostratigraphy of the W. Mediterranean Neogene. Trabajos Sobre Neogeno Cuaternario, 7.
Mein, P. (1979). Rapport d'activité du groupe de travail vertébrés mise à jour de la biostratigraphie du Neogène basée sur les mammifères. Annales de Géologie des Pays Helléniques, tome hors série, 3, 1367–1372.
Mein, P. (1990). Updating of MN zones. In E. H. Lindsay, V. Fahlbusch, & P. Mein (Eds.) European Neogene Mammal Chronology (pp. 73–90). New York & London: Plenum Press.
Mein, P. (2000). La biochronologie des Mammifères néogènes d’Europe. L’échelle MN, son application à la succession des faunes du Portugal. Ciências da Terra (UNL), 14, 335–342.
Meulen, A. J. Van der, García-Paredes, I., Álvarez-Sierra, M. Á., Hoek Ostende, L. W. van den, Hordijk, K., Oliver, A., López-Guerrero, P., Hernández-Ballarín, V., & Peláez-Campomanes, P. (2011). Biostratigraphy or biochronology? Lessons from the Early and Middle Miocene small Mammal Events in Europe. Geobios, 44, 309–321.
Meulen, A. J. Van der, García-Paredes, I., Álvarez-Sierra, M. Á., Hoek Ostende, L. W. van den, Hordijk, K., Oliver, A., & Peláez Campomanes, P. (2012). Updated Aragonian biostratigraphy: small mammal distribution and its implications for the Miocene European Chronology. Geologica Acta, 10(2), 159–179.
Morales, J., & Soria, D. (1985). Carnivoros y Artiodactylos de la provincia de Madrid. In M. T. Alberdi (Ed.), Geología y paleoecología del Terciario continental de la provincia de Madrid (pp. 81–89). Madrid: CSIC.
Mottl, M. (1970). Die jungtertiären Säugetierfaunen der Steiermark, Südösterreichs. Mitteilungen des Museums für Bergbau, Geologie und Technik am Landesmuseum Joanneum Graz, 31, 1–92.
Paraskevaidis, I. (1940). Eine obermiocäne Fauna von Chios. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, 83, 363–442 plates 11–14.
Pickford, M. (2011). Small suoids from the Miocene of Europe and Asia. Estudios Geológicos, 67(2), 541–578.
Pickford, M. (2012). Les Suoidea (Artiodactyla) de Sansan: systématique, paléoécologie, biogéographie et biochronologie. Mémoires du Muséum National d'Histoire Naturelle, 203, 249–277.
Pickford, M. (2014). Sus Valentini Filhol (1882) from St Gaudens (MN 8–9) France: blighted from the outset but a key to understanding late Middle Miocene Tetraconodontinae (Suidae, Mammalia) of Europe. Mainzer Naturwissenschaftlich Archiv, 51, 67–220.
Pickford, M. (2016a). Revision of European Hyotheriinae (Suidae) and Doliochoeridae (Mammalia). Münchener Geowissenschaftliche Abhandlungen, Series A, 44, 1–270.
Pickford, M. (2016b). Biochronology of European Miocene Tetraconodontinae (Suidae, Artiodactyla, Mammalia) flowing from recent revision of the subfamily. Annalen des Naturhisorischen Museums in Wien, Serie A, 118, 175–244.
Pickford, M. (2017). Revison of “peccary-like” Suoidea (Artiodactyla: Mammalia) from the Neogene of the Old World. Münchener Geowissenschaftliche Abhandlungen, Series A, 46, 1–144.
Pickford, M., & Laurent, Y. (2014). Valorisation of palaeontological collections: nomination of a lectotype for Conohyus simorrensis (Lartet, 1851), Villefranche d’Astarac, France, and description of a new genus of tetraconodont. Estudios Geológicos, 70(1), 1–28.
Pickford, M., & Moyà Solà, S. (1994). Taucanamo (Suoidea, Tayassuidae) from the Middle Miocene (MN04a) of Els Casots, Barcelona, Spain. Comptes Rendus de l’Academie des Sciences, Paris, 319(série II), 1569–1575.
Pickford, M., & Moyà-Solà, S. (1995). Eurolistriodon gen. nov. (Suoidea, Mammalia) from Els Casots, early Middle Miocene, Spain. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, 98, 343–360.
Pilgrim, G.E. (1925). Presidential address to the geological section of the 12th Indian Science Congress (pp. 200–218). Proceeedings of the 12th Indian Science Congress.
Popov, S. V., Rögl, R., Rozanov, A. Y., Steininger, F. F., Shcherba, I. G., & Kovac, M. (2004). Lithologial-Paleogeographic maps of Paratethys, 10 maps Late Eocene to Pliocene. Courier Forschungsinstitut Senckenberg, 250, 1–46 10 maps.
Prieto, J., Antoine, P. O., Made, J. van der, Métais, G., The Phuc, L., Truong Quan, Q., Schneider, S., Ngoc Tran, D., Vasilyan, D., The Viet, L., & Böhme, M. (2017). Biochronological and palaeobiogeographical significance of the earliest Miocene mammal fauna from Northern Vietnam. Palaeobiodiversity and Palaeoenvironments, 98(2), 287–313.
Rabeder, G. (1978). Die Säugetiere des Badenien. In A. Papp, I. Cicha, I. Seneš, & F. F. Steininger (Eds.) Chronostratigraphie und Neostratotypen. Miozän der Zentralen Paratethys. M4 Badenien (Moravien, Wielicien, Kosovien) (pp. 467–481). Bratislava: VEDA.
Rabeder, G., & Steininger, F. (1975). Die direkten biostratigraphischen Korrelationsmöglichkeiten von Säugetierfaunen aus dem Oligo/Miozän der zentralen Paratethys. In Proceedings of the VIth Congres (pp. 177–183). Regional Committee on Mediterranean Neobgene Stratigraphy - Bratislava.
Rögl, F. (1998). Palaeogeographic considerations for Mediterranean and Paratethys seaways (Oligocene to Miocene). Annalen des Naturhistorischen Museums Wien, 99a, 279–310.
Rögl, F., & Steinginger, F. F. (1983). Vom Zerfall der Tethys zu Mediterran und Paratethys. Annalen des Naturhistorischen Museums Wien, 85A, 135–163.
Sicuro, F. L., & Oliveira, L. F. B. (2002). Coexistence of peccaries and feral hogs in the Brazilian Pantanal wetland: an ecomorphological view. Journal of Mammalogy, 83(1), 207–217.
Simpson, G. G. (1945). The principles of classification and a classification of mammals. Bulletin of the American Museum of Natural History, 85, 1–350.
Stefen, C. (in press). The castorids (Mammalia, Castoridae) from the (early) middle Miocene of Gračanica (Bosnia-Herzegovina). In U. B. Göhlich, & O. Mandic (Eds.) The drowning swamp of Gračanica (Bosnia-Herzegovina) - a diversity hotspot from the middle Miocene in the Bugojno Basin. Palaeobiodiversity and Palaeoenvironments. https://doi.org/10.1007/s12549-018-0365-9. [this issue]
Stehlin, H.G. (1899–1900). Ueber die Geschichte des Suiden-gebisses. Abhandlungen der schweizerischen paläontologischen Gesellschaft, first part, 1899, 26, 1–336, second part, 1900, 27, 337–527, 10 plates.
Stehlin, H. G. (1909). Remarques sur les faunules de mammifères des chouches éocènes et oligocènes du Bassin de Paris. Bulletin de la Société Géololgique de France, 19, 488–520.
Thenius, E. (1949). Die Tortone Säugetierfauna von Neudorf an der March (CSR) und ihre Bedeutung für die Helvet-Torton-Grenze. Anzeiger der Österreichische Akadademie der Wissenschaften mathematisch-naturwissenschaftliche Klasse, 7, 160–171.
Thenius, E. (1956). Die Suiden und Tayassuiden de steirischen Tertiärs. Sitzugsberichte der Akademie von Wissenschaften, Wien, 1(165), 337–382.
Wedmann, S. & Skartveit, J. (in press). First record of March flies (Insecta: Diptera: Bibionidae) from the Miocene Gračanica mine (Bugojno, Bosnia-Herzegovina). In U.B. Göhlich, & O. Mandic (Eds.) The drowning swamp of Gračanica (Bosnia-Herzegovina) - a diversity hotspot from the middle Miocene in the Bugojno Basin. Palaeobiodiversity and Palaeoenvironments. https://doi.org/10.1007/s12549-018-00369-w. [this issue]
Wessels, W., Bruijn, H. De, Markovic, Z. & Milivojevic, M. (in press). Small mammals from the opencast lignite mine Gračanica (Bugojno, middle Miocene), Bosnia and Herzegovina. In U.B. Göhlich, & O. Mandic (Eds.) The drowning swamp of Gračanica (Bosnia-Herzegovina) - a diversity hotspot from the middle Miocene in the Bugojno Basin. Palaeobiodiversity and Palaeoenvironments. https://doi.org/10.1007/s12549-018-0366-8. [this issue]
Acknowledgements
I thank U. Göhlich for inviting me to study the Suoidea from Gračanica, the following persons for discussions on themes related with this paper: M. Böhme, U. Göhlich, O. Mandic, P. Pelaez Campomanes, J. Prieto, G. Rössner, D. Vasilyan, W. Wessels, and as reviewers M. Orliac and an anonymous person. The following persons gave me access to fossils used for comparison, or helped me in any other way: J. Agustí, L. Alcalá, F. Alférez, B. Alpagut, M. Arif, B. Azanza, O. Bardot, Basu, P. Brewer, S. Calzada, E. Cioppi, A. Currant, G. Daxner Höck, F. Duranthon, T. Engel, B. Engesser, V. Fahlbusch, M. Fortelius, S. Fraile, J.L. Franzen, J. Galkin, L. Ginsburg, C. de Giuli, U. Göhlich, A. González Alonso, W. Gräf, C. Guérin, E. Güleç, O. Hampe, K. Heissig, E. Heizmann, J. Hooker, M. Hugueney, K.A. Hünermann, S.T. Hussain, K. Kowalski, Y. Laurent, H. Lutz, E. Menéndez, S. van der Mije, J. Morales, R. Niederl, R. O’Leary, S. Pavic, M. Philippe, K. Rauscher, M. Raza, L. Rook, G. Rössner, M. Rummel, G. Saraç, U. Schmid, H.K. Schmutz, P.Y. Sondaar, I. Stefanović, M. Telles Antunes, E. Ünay, L. Via, R. Ziegler.
Funding
This paper benefited from projects Synthesys GB-TAF-4119 and AT-TAF-3663 and from projects CGL2004-04169/BTE, CGL2008-03881 of the Spanish Ministry of Science, and CGL20l5 65387-C3-3-P from the Spanish Ministery of Economy and Competitiveness.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is a contribution to the special issue “The drowning swamp of Gračanica (Bosnia-Herzegovina) - a diversity hotspot from the Middle Miocene in the Bugojno Basin"
Electronic supplementary material
ESM 1
(PDF 3.79 mb)
Rights and permissions
About this article

Cite this article
van der Made, J. The Suoidea from the Middle Miocene of Gračanica (Bugojno Basin, Bosnia and Herzegovina)—evolution, taxononomy, and biostratigraphy. Palaeobio Palaeoenv 100, 321–349 (2020). https://doi.org/10.1007/s12549-020-00420-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-020-00420-9