
Abstract
After a long period of inactivity, recent excavations at the late Miocene Maragheh Formation unexpectedly resulted in the discovery of the first fossil hominoid and second Mesopithecus remains from this area. The discovery motivated a new international initiative to conduct research in these rich fossil sites. These studies focused on the fossil hominoid and its locality, aiming to reveal more about the context of this fossil discovery. Detailed stratigraphy, sedimentology and magnetostratigraphy studies were conducted. New samples from volcaniclastic key horizons (pumice beds) in Dareh Gorg, where the hominoid fossil site is located, were dated by radiometric methods. The radiometric age determinations provide a firm tie-point for the geochronology. The polarity pattern in the palaeomagnetically investigated section corroborates the K-Ar results. The preliminary magnetostratigraphic results suggest that the hominoid locality can be correlated to the normal polarity chron C4n.2n (8.108–7.695 Ma), C4n.1n (7.642–7.528 Ma) or C3Br.1n (7.285–7.251 Ma), placing it at intervals corresponding to the mammal units MN11 or possibly early MN12. The study of fossil hominoid indicates broad affinities with a number of contemporaneous taxa from the Balkan-Iranian palaeoprovince, as well as Siwaliks and southeast Asia. A preliminary analysis of the accompanying (in situ) fauna at the hominoid site indicates the highest similarity of this level to Turolian hominoid- and Mesopithecus-bearing localities in Turkey, Greece and Bulgaria. However, some environmental differences are observed among these localities, based on their faunal structure and taxon properties, as well as in the different masticatory adaptations of their hominoids.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
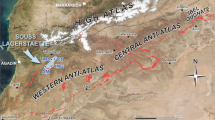
Maragheh is located in the province of East Azarbaijan, northwest Iran. As the capital of the Ilkhanid dynasty (Genghis Khan’s successors, thirteenth century), the city has several historic monuments including the ruins of a world famous observatory. Beyond this glorious past, this region is renowned for preserving some of the richest late Miocene mammal fossil localities in western Asia, collectively known as the Pikermian chronofauna (Bernor 1986; Eronen et al. 2009; Mirzaie Ataabadi et al. 2013). The fossil localities of Maragheh are mainly located between the present villages of Mordagh, Karajabad and Shalilvand (Fig. 1), known in older texts as Mirduq, Kherjabad, Kerjaveh and Sholl’avand, Chollevand (e.g. Kamei et al. 1977; Bernor 1986). However, additional fossil sites occur in a large area between 37° 20′–37° 25′ N latitude and 46° 16′–46° 37′ E longitude, even within the present extension of Maragheh City (Fig. 1).

Location map of Maragheh and its fossil localities. Top left corner: A, fossil localities in Kopran (oldest); B, main fossiliferous areas including the protected zone and hominoid locality in Dareh Gorg (mostly middle Maragheh); C, eastern most and youngest sites in Ilkhchi. The road map shows the exact location of hominoid site between Mordagh and Karajabad. Maps redrawn and modified after Sakai et al. 2016, this issue (top right), Erdbrink et al. 1976 (top left) and Kamei et al. 1977 (bottom)
Fossil bones of Maragheh have probably been known to local villagers for a long time. They were considered the remains of pre-flood animals or those of legendary and monstrous creatures. The Russian explorer Khanikoff is usually credited with first finding the Maragheh fossils in 1840, initially studied in the latter half of the nineteenth century. The main early fossil excavations in Maragheh were carried out by Austrian and French delegations in late nineteenth and early twentieth centuries (Mirzaie Ataabadi and Fortelius 2016, this issue). While the most important studies were conducted during the 1970s by three scientific groups: a combined Dutch-German group led by Erdbrink and assisted by University of Tehran staff (Erdbrink et al. 1976), a joint Kyoto University-Geological Survey of Iran group led by Kamei (Kamei et al. 1977) and the Lake Rezaiyeh Expedition (LRE) led by Professor Campbell and supported by Iranian National Museum of Natural History (MMTT), Tehran, and the LSB Leakey Foundation, Berkeley, California (Campbell et al. 1980).
The late 1970s revolution in Iran stopped the scientific excavations in Maragheh for nearly three decades. At the start of twentieth century, interest of Iranian scientists in renewal of activities in Maragheh fossiliferous areas, plus some national plans in the country to establish a palaeontological research centre motivated Iran’s Department of Environment (DOE) and Iranian National Museum of Natural History (MMTT; Muze Melli Tarikh Tabeie in Persian) to initiate and sponsor new excavations in Maragheh. Thus a new excavation project started in Maragheh in July 2004, under the supervision and leadership of Dr Zeinolabedin Pourabrishami (Emeritus Professor, University of Tabriz).
During this project, initially two sites (sites I and II) were excavated at Middle levels of Maragheh Formation (Bernor 1986) in Dareh Gorg (Gurt Dareseh) area near Mordagh in July and August 2004 (Fig. 2a, b). One of these sites (site II) revealed a very rich fossiliferous layer (Fig. 2a, c). Hence, the team decided to expand the excavations and extend it towards north and west. These complementary excavations were carried out during November and December 2004. The northern extension of this site (site II) proved to be quite successful and more fossils were revealed (Fig. 2d). In late November 2004, quite unexpectedly remains of fossil primates were also discovered in this site (Pourabrishami 2005).

Field photos from excavation and other activities in hominoid locality, Dareh Gorg. a Workers and Maragheh DOE staff in 2004 during the main excavation at site II (section A, Fig. 5); b a 2004 view from the excavation in site II. c Inside the field museum (2015), the main excavation site II (section A, Fig. 5); d the northern extension of site II, where the hominoid and primate fossil have been discovered (sections B, C, Fig. 5); e INSPE team at field station, next to the field museum and hominoid site, Dareh Gorg, 2008 (from right to left: Hideo Nakaya, Yutaka Kunimatsu, Tetsuya Sakai, Mikael Fortelius, Philippe Richir, Yoshihiro Sawada, DOE staff, DOE staff, Sevket Sen, Zhang Zhaoqun and Zhou Wei); f general view of the field museum and station in Dareh Gorg, Mordagh, Maragheh (2009); g Mahito Watabe (centre) and students (Kari Lintulaakso and Pierre Mauries) checking the strata (2009); h Palaeomag team sampling near the field museum in 2015 (from right: Anu Kaakinen, Johanna Salminen and Mohammad Paknia). Photos a, b courtesy of MMTT, c–h M. Mirzaie Ataabadi
Therefore, although some previous excavations (e.g. Japanese and American), which aimed at finding fossil hominoids in Maragheh, were unsuccessful, this late activity by an Iranian team unexpectedly resulted in the discovery of the first fossil hominoid from Maragheh Formation. The hominoid material so far recovered from site II is restricted to the partial maxilla with two teeth (MMTT3453) described in detail elsewhere in this issue (Suwa et al. 2016, this issue). If additional parts of this fossil were present, they have unfortunately been lost.
Upon the request of MMTT, one of us (MMA) visited these excavations and the hominoid site in June 2005 for scientific evaluation. At that time, the first section of excavation (original site II known here as part A in Fig. 5) was temporarily covered but the northern extension or the hominoid site (part B in Fig. 5) was semi-open, apparently due to more recent excavations. The work was done hastily and many fossils were damaged and smaller tiny fossils were apparently neglected, which caused a possible bias in the fossil site towards larger bones. Since most of the work was done, MMA proposed preservation of the fossils by using proper material and methods and suggested a detailed study of the locality because of the importance of the fossil hominoid discovery. During this first visit, the excavation site supervisor (Mr. Pashaei, a local archaeologist) discussed the discovery circumstances of fossil hominoid and showed the approximate place of its occurrence in the northern extension site II (Fig. 2d and part B in Fig. 5).
Meanwhile, the large primate specimen was identified by the team leader (Z. Pourabrishami) as a primate and was taken to Tabriz University for conservation, while other fossils were preserved in situ following the decision of DOE/MMTT to turn the site into the first fossil field museum in Iran. The first stage of a permanent cover over the fossil site was built in 2005 and completed later (Fig. 2f). Also, 10 km2 of the Maragheh fossiliferous area (including the hominoid site and its surroundings) was nominated as a national protected zone by DOE.
Because of the importance of the hominoid discovery in Maragheh, MMA tried to persuade officials in the DOE/MMTT to facilitate the study of the site and area by an international team. This fortunately happened later in 2006, thanks to Dr Delavar Najafi Hajipour, the DOE deputy for natural environment. Therefore, MMTT-University of Helsinki-Japanese initiative known as the International Sahand Palaeoenvironment Expedition (INSPE) was initiated (Mirzaie Ataabadi et al. 2013 and papers in this issue). In September 2007, the first visit of the INSPE team to Maragheh was undertaken. During the visit, the field museum (Site II/hominoid site) was also inspected and preliminary studies undertaken by some of us. The INSPE initiative later secured two field seasons, in May 2008 and again in May 2009, during which both small and large excavations were carried out in the Dareh Gorg area. Although these new excavations did not produce new hominoid remains, detailed study of stratigraphy, sedimentology and geochronology was carried out by an international team of experts in the vicinity of Dareh Gorg and around the hominoid fossil site (Fig. 2e, g). In late May, 2009 the INSPE team could finally access the hominoid specimen in the MMTT office in Tehran and one of us (MF) investigated and documented the specimen and made casts of the teeth for further studies. The limited access to the specimen was due to circumstances that developed around this specimen in Maragheh. At one point the specimen was mistakenly attributed to Homo, which gave rise to various controversies surrounding the specimen. Therefore, the DOE decided to keep the specimen in Tehran.
The unrests following the presidential elections in summer 2009 in Iran temporarily interrupted the research of the INSPE team. In June 2015, the palaeomagnetism team finally visited Maragheh hominoid site and conducted its first field season there, sampling the stratigraphic interval in which the hominoid site occurs (Fig. 2h). Here, a synopsis of the recent studies by INSPE team in Maragheh is presented and more details of the hominoid site and its fossils are disclosed. A preliminary investigation of the palaeoecology based on the fossils present in the hominoid-bearing level of Middle Maragheh interval is also presented.
Material and methods
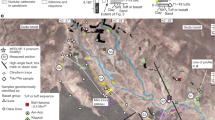
The fossil material studied here in a preliminary manner is kept in situ in the DOE field museum in Dareh Gorg, Mordagh area, Maragheh (Fig. 2c–f). The only exceptions are the primate fossils, which are in MMTT office in Pardisan Park, Tehran (Fig. 3). The fossils were studied on site and identified. Measurements were taken when possible and photographs of some specimens were taken for more detailed identification (Fig. 4). Each site was divided into square metres blocks and photographs of each block were taken from a controlled level. Fossil distribution sketch maps were later drawn based on these photographs and then placed together to make the final map (Fig. 5). The final sketch was then checked on site to correct the possible errors.

Fossil primates from site II, Middle Maragheh interval, Dareh Gorg, Maragheh. a, b Hominoid specimen (MMTT3453); c, d Mesopithecus specimen (MMTT3553). Photos courtesy of MMTT, scales in centimetre

Fossils as preserved in the field museum. a–d Some fossil clusters at site II/hominoid locality, black scale bar 20 cm. e–h Individual fossils from the site including Rhinocerotidae, Equidae, Hyaenidae, Proboscidea, Giraffidae and Bovidae, black scale bar 10 cm. (photos a–d, h M. Mirzaie Ataaabdi 2015 and e–g M. Fortelius 2009)

The distribution of fossils in the field museum as seen in A–C sections/clusters. The fossil hominoid has been discovered in B (asterisk). The distance between A and B (M-O) is covered and possibly bones may occur beneath. Each mesh is 1 m2
Data for comparisons of crown height percentages, faunal resemblance and community structure of the Maragheh hominoid level with other sites were obtained from the NOW database (Fortelius 2016). We used faunal resemblance indices (FRI) to investigate the taxonomic expression of potential palaeoecological/palaeoenvironmental similarities between Maragheh hominoid level and other late Miocene (Vallesian/Turolian) hominoid-bearing localities in the eastern Mediterranean region (Greco-Iranian or Balkan-Iranian palaeoprovince). Faunal lists from Sivapithecus-bearing localities of the Indian subcontinent (Barry et al. 2002) were also included. This survey follows investigations on palaeobiogeographic analysis by Fortelius et al. (1996), Bernor et al. (2001), and more recently Mirzaie Ataabadi et al. (2013). We undertake genus-level faunal resemblance index (GFRI) studies using both the Simpson (1943) and the Dice (Sokal and Sneath 1963) indices. Dice FRI is calculated as 2C/(A + B), where C is the number of shared taxa between two faunas, and A and B are the total number of taxa present in fauna 1 and fauna 2. Simpson’s FRI is calculated as C/smaller of (A or B).
Geology
The Azarbaijan region in northwest Iran is geologically distinguished by the concentration of Cenozoic volcanic activity that continued up to the Miocene and even the Plio/-Pleistocene. Maragheh is part of this region and its main volcanic edifice is the Sahand Volcano. Mt. Sahand is a large volcanic complex which covers an area of about 10,000 km2 and, despite its circular outline, is not a single volcano. A series of distinct volcanic cones are arranged along an east-west trend collectively forming this enormous volcanic massif (Mirzaie Ataabadi et al. 2013; Sawada et al. 2016, this issue). After Paleogene volcanic activities, the late Paleogene-early Neogene sedimentation began with oxidised terrestrial deposits (Lower Red Fm.), a marine unit (Qom Fm.) in the middle and a regressive cycle of terrestrial deposits (Upper Red Fm.) on top. These sequences can be observed in areas east and south east of Maragheh. The Sahand volcanic massif activities in the Miocene are associated with Alpine tectonic phases and continued up to the Quaternary. Due to this eruptive activity, different kinds of volcanic and volcaniclastic sediments have been deposited in the Maragheh area (Pourabrishami 2005).
The late Miocene deposits of the Maragheh Basin consist of a thick sequence of volcaniclastic continental strata with a basal pyroclastic unit, known as the Basal Tuff (Campbell et al. 1980). The Basal Tuff represents a single air-fall unit with local thickness of over 80 m. This unit is a uniform, unbedded and structureless deposit of white, devitrified ash that represents a tremendous pyroclastic event with considerable outcrops south and northeast of the central fossiliferous area. This unit is useful for long-range intra-basin correlations (Campbell et al. 1980; Bernor et al. 1980; Bernor 1986). Kamei et al. (1977) designated the entire 500–600-m-thick Late Miocene sequence in Maragheh Basin as the Maragheh Formation. They differentiated two units: a lower fossiliferous member (160 m) and an upper non-fossiliferous part forming the upland hills of Mt. Sahand. Campbell et al. (1980) restricted the Maragheh Formation to the lower 300 m volcaniclastic and fossiliferous series. The fossil-bearing sequence of Maragheh Basin is confined to the lower 150 m of a 300-m-thick Maragheh Formation. This formation seems to rest with a low-angle regional unconformity on the Basal Tuff Formation.
Stratigraphy and sedimentology
Kamei et al. (1977) identified intercalated tuff and pumice beds around Dareh Gorg and uphill on the Sahand and named them successively as the Basal Tuff, the Mordaq Tuff, the Lower Pumice, the White Fine Tuff, the Upper Pumice, the “Scoria” Bed, the Pumice Falls, the Sargizeh Tuff and the Korde Deh Ash Flow. Campbell et al. (1980), Bernor et al. (1980) and Bernor (1986) used different names for key beds from Kamei et al. (1977) except for the Basal Tuff, Mordaq Tuff (Fig. 6) and Korde Deh Ash Flow. Kamei et al. (1977)’s Lower Pumice and Upper Pumice Beds were referred to as Gurt Dareseh Pumice and “Loose Chippings Marker”, respectively (Campbell et al. 1980; Bernor et al. 1980; Bernor 1986). Bernor (1986) stated that the continuity of each key bed is limited and these key beds cannot be used for the extensive correlation within Maragheh Formation, except for the Basal Tuff. However, within smaller areas, such as Dareh Gorg, most of the key beds have lateral extensions and can be used for correlation (Fig. 6).

Tentative correlation of three stratigraphic columns from Dareh Gorg section by A, Japanese (Kamei et al. 1977); B, American (Bernor et al. 1980); and C, INSPE (Mirzaie Ataabadi et al. 2013) teams (redrawn and modified after Kamei et al. 1977 and Mirzaie Ataabadi et al. 2013). The correlation is based on several key beds and horizons from above-mentioned works including, from base to top, Basal Tuff, Mordagh Tuff, Lower Pumice (LP), White Fine Tuff (WFT and WT), Upper Pumice and Loose Chippings. LP in section C is probably different from LP by Japanese in section A. Correlation of LP and Upper Pumice from section A to other sections is not certain and hence is marked by dashed line and question mark. Also note that MP (section C) is believed to be reworked from LP, hence showing similar ages (Sakai et al. 2016 and Sawada et al. 2016, this issue). Numbers in the left side of columns A-B refer to fossil localities by Japanese and American teams. Numbers in parenthesis next to keybeds indicate absolute dates. In section B, dates are Ar/Ar (Swisher 1996) and 8.2 next to site II refers to approximate interpolated age (Ma) for MMTT sites 1a, 5, 36, 8, 42 (after Mirzaie Ataabadi et al. 2013) which could corresponds to the age of hominoid level based on interpolation according to Swisher 1996 dates. In section C, the lowermost date belongs to Mordagh Tuff and the uppermost refers to Upper Pumice (Sawada et al. 2016, this issue). Site II (C) refers to hominoid locality (asterisk)
Since our studies stand on the geological works carried out by Kamei et al. (1977), names of the key beds as defined in their work are used here. However, our Lower Pumice was likely not recorded by them due to its possible local occurrence and less extensive continuation. Also, a thick pumice-rich bed referred here as the Middle Pumice might be the Lower Pumice of Kamei et al. (1977). The white coloured tuffaceous interval discovered ca. 10 m above the Middle Pumice is referred to here as “White Tuff” (Fig. 6).
Campbell et al. (1980) proposed three litho/biostratigraphic intervals for the Maragheh Fm.: the Lower, Middle and Upper Maragheh. The boundaries of these intervals were refined by Bernor (1986). These units are defined based on their distance from the “Loose Chippings Marker (LC)”. The interval ranging from −150 to −52 m from “Loose Chippings” is defined as Lower Maragheh, from −52 to −20 m from the LC the Middle Maragheh, and from −20 to +7 m from the LC as Upper Maragheh (Bernor 1986).
The hominoid-bearing site of the Maragheh is located in one of the richest fossiliferous areas of this Formatiom (Middle Maragheh interval) in Dareh Gorg. The position of the fossil site is N 37° 21′ 35.8″ and 46° 24′ 25.5″ E. This area has been one of the main locations excavated by all groups who conducted research in Maragheh. However, only the Japanese and American groups documented the exact position of their sites in their stratigraphic columns (Fig. 6).
A tentative correlation of the hominoid fossil level in our stratigraphic column (Fig. 6C) to those of the American (Mordagh-Karajabad) section (Bernor et al. 1980; Mirzaie Ataabadi et al. 2013) and the Japanese section in Dareh Gorg (Kamei et al. 1977) shows that the hominoid level was not excavated by these groups, even though presence of fossils at that level was documented (Fig. 6A, B). This tentative correlation is made possible by tracing several key marker beds in the area, including the Lower and Upper Pumices, the White (Fine) Tuff and the Loose Chippings. The American (MMTT) sites 1a and 5 in Dareh Gorg are thus interpreted to be probably above the hominoid level. While the sites 8, 42, 36 in Aliabad, adjacent to Dareh Gorg, probably close to the hominoid level, have produced few fossils and faunal list for them do not appear to be available (Bernor 1978). Accordingly, the Japanese excavation levels seem to be located about 10 m (site I, locality 6 in Fig. 6A) to 25 m (sites II, III, IV, locality 8 in Fig. 6A) above the hominoid level. It seems that only the Japanese localities 2 and 3 (Fig. 6A), which were not been excavated (Kamei et al. 1977), might have been at the same level with Maragheh hominoid site. Therefore, unfortunately the excellent material collected by these groups from Dareh Gorg does not directly help our understanding of the faunal composition of the hominoid level of the Maragheh For-mation.
The lithology of the hominoid site, which can be observed best from a vertical wall behind and along the excavation site (field museum now), is dominated by silty and sandy deposits with little intrusion of gravels. The fossiliferous layer itself is dominated by clay and tuffaceous clay and in part by silts and fine sands. It is apparently bounded by two layers of palaeosols. The hominoid fossil site is located ca. 8 m below a thick pumice bed known as the Middle Pumice (Fig. 6C). Facies analysis shows a dominance of floodplain depositional environments (cf. Sakai et al. 2016, this issue), characterised by massive silty sand and sandy silt beds, commonly exhibiting palaeosol formation and poorly sorted texture, with a few intercalated laminated silts. A notable feature is the thick pumice bed (“the Middle Pumice”), which consists of stratified sand and pebble beds with occasional cobble-rich horizons in the lower part while the upper part mainly exhibits well sorted laminated silts and fine sand. This unit has been interpreted as representing hyperconcentrated flow deposits, likely to have been accumulated from a single flow (Sakai et al. 2016, this issue).
Chronology and palaeomagnetism
Kamei et al. (1977) described different aspects of the Maragheh Formation, along with K-Ar and zircon fission track ages from Maragheh volcanites and tephras (Fig. 6A). Bernor et al. (1980) provided zircon fission track, and plagioclase and hornblende K-Ar ages from the Maragheh Formation and its basal tuff. Swisher (1996) subsequently reported laser total fusion 40Ar-39Ar plagioclase ages from the basal tuff and tuff beds of the Maragheh Formation (Fig. 6B).
Based on these studies, the Maragheh fauna has a chronologic range of nearly 9 Ma to less than 7.4 Ma but the bulk of fossil material is from the middle and upper parts of the fossiliferous section (“Middle and Upper Maragheh”). For the “Middle Maragheh”, where several groups have collected fossils, the current oldest interpolated age is 8.165 Ma, while the youngest localities (“Upper Maragheh”) date to 7.68 and 7.4 Ma (Mirzaie Ataabadi et al. 2013).
Sawada et al. (2016, this issue) report new hornblende and plagioclase K-Ar ages for refinement of the geochronology in Maragheh Formation. Their K-Ar ages from the hornblende and plagioclase separates are as follows: (1) 8.14 ± 0.27 Ma (mean value), as represented by the Mordagh Tuff bed, (2) 7.54 ± 0.22 Ma from the Lower Pumice beds A, (3) 6.95 ± 0.28 Ma from the Lower Pumice beds B, (4) 7.87 ± 0.29 Ma from the Middle Pumice bed and (5) 6.96 ± 0.31 Ma from the Upper Pumice bed (Fig. 6C). The K-Ar ages are consistent with the stratigraphic division, except for those from the Middle Pumice bed. The age of the Middle Pumice bed is older than that of the Lower Pumice beds. It is inferred that these pumice clasts were derived (reworked) from the Lower Pumice beds, as supported by the fact that the Middle Pumice bed represents flood deposits. Furthermore, the chemical compositions of glass, hornblende and biotite from the Middle pumice bed are very similar to those from the Lower Pumice bed. This suggests that all these pumices were of the same magmatic origin (Sawada et al. 2016, this issue).
Two previous palaeomagnetic campaigns have been also carried out for the Maragheh Formation by Erdbrink et al. (1976) and by Kamei et al. (1977). Erdbrink et al. (1976) sampled different tuffaceous and other volcanic layers at ten levels along a 79.5-m composite section in Maragheh. Seven horizons provided reliable results showing normal polarities for the lowermost samples and reversed polarities for the overlying levels. Kamei et al. (1977) continued palaeomagnetic studies by sampling nine levels at Dareh Gorg section from coarse biotite tuff below the Ignimbritic tuff (Mordagh tuff) and spanning up to the Korde deh ash flow, far above the Upper Pumice. According to Kamei et al. (1977), reasonable results were obtained from four sites: a reversed polarity from the coarse Biotite tuff below the Ignimbritic tuff and normal polarities for the Ignimbritic tuff, the White fine tuff, and for the Korde deh ash flow. Nevertheless, with these sparse palaeomagnetic data, it was not possible to draw any conclusions about the age of the section.
Recently, we started a high-resolution palaeomagnetic study in Maragheh sequence (Salminen et al. 2016, this issue). One hundred fifteen levels along an approximately 27-m-thick interval were sampled in the middle Maragheh Formation at Dareh Gorg during the first field season (2015). Characteristic remanent magnetization directions obtained by alternating field demagnetization define a coherent magnetostratigraphy that is supported by thermal demagnetization on a set of sister specimens Three polarity intervals were recognised; the reversed polarity magnetozone at the middle part of the section at around 15–21 m, bounded by normal polarities above and below.
Our results provide a reliable preliminary magnetostratigraphy for the Middle Maragheh interval in Dareh Gorg section, where the hominoid fossil occurs. The extracted polarity pattern does not allow a unique correlation to the geological time scale, but based on the paleontological constraints and recent K-Ar ages, three correlations to the geomagnetic polarity time scale seem likely.
In light of recent K-Ar ages (Sawada et al. 2016, this issue), we prefer correlating the reversed polarity zone (Fig. 7) to C4n.1r, C3Br.3r or C3Br.1r. This suggests placing the recently discovered hominoid locality to the upper part of the normal polarity chron C4n.2 (8.108–7.695 Ma), C4n.1n (7.642–7.528 Ma) or C3Br.1n (7.285–7.251 Ma). On the other hand, using the interpolated ages of MMTT localities in Dareh Gorg, which approximately correspond to the hominoid level (Mirzaie Ataabadi et al. 2013), an age of ca. 8.2 Ma (>8.17 Ma below localities MMTT 1A and 5) can be inferred for this site. These interpolations are based on single crystal Ar-Ar dates (Swisher 1996) that have very narrow error bars compared to K-Ar ages. Thus, the Maragheh hominoid level is likely equivalent to the European mammal unit MN11 or possibly early MN 12. Palaeomagnetism study of the whole Middle Maragheh succession will hopefully resolve this minor discrepancy.

Lithological column and interpreted magnetostratigraphic polarity for Middle Maragheh interval close to the hominoid site. In the polarity column, black/white denotes normal/reversed polarity. Grey shaded zones represent undefined or transitional polarity. Correlation of the magnetostratigraphy to the Astronomically Tuned Neogene Time Scale (ATNTS) of the geological time scale 2012 (Gradsttein et al. 2012) is also indicated and three possible options and their ages are shown
Taphonomy and palaeoenvironment
About 160 large fossil bones in the Maragheh hominoid fossil site in Dareh Gorg are kept in situ in the field museum (Figs. 2c, d, f and 5). According to Campbell et al. (1980), Maragheh fossils do not show preferred orientation, and completely articulated fossils are rare. Nevertheless, the preservation of each fossil is good. Bernor (1986) stated that fossils were floated in massive beds of overbank origin. Mirzaie Ataabadi et al. (2013) pointed to the presence of a single, almost complete articulated small carnivore from MMTT locality 13 reported by Bernor 1978. Kamei et al. (1977) found that some fossils do show a preferred orientation and are concentrated to form a pile of fossils, leading them to infer the effects of stream. They also described a peculiar occurrence of fossils obliquely oriented against bedding plain. Similar conditions to those described by Kamei et al. (1977) are seen in the Maragheh hominoid site, with fossils showing preferred orientation and occurring in densely concentrated piles (Fig. 5). The floated occurrence within massive beds and oblique occurrence are similar to the fabric of gravels in debris flow deposits (e.g. Postma et al. 1983; Naruse and Masuda 2006). The debris flow could contribute to good fossil preservation as the upper part of the flow forms a rigid plug in which sediment and water mixture form a non-Newtonian laminar flow, suppressing the interaction among clasts and minimising abrasion of the fossil surface during transport (Nemec and Steel 1984; Mazza and Ventra 2011). This hypothesis must be tested by the future taphonomic and sedimentological analysis, hopefully leading to a more concrete understanding of the fossil preservation process during the deposition of the Maragheh Formation (Sakai et al. 2016, this issue).
The Maragheh fossil assemblage is usually considered as part of the Eurasian savanna fauna of the late Miocene. Campbell et al. (1980) discussed the environment at the time of Maragheh Formation based on the faunal assemblage and suggested a closer habitat than previously thought, a savanna dominated by woodland with wooded grassland nearby. Strömberg et al. (2007) indicated the presence of grassland based on analysis of phytoliths and indicated high temperature relative to other “Pikermian” localities. Preliminary results of mesowear analysis by Bernor et al. (2014) indicated a mosaic habitat with woodland and some open savanna environment. Yamada et al. (2016, this issue) also provide mesowear data for sympatric bovid and equid species from a single quarry in Middle Maragheh (above the hominoid level) that support the mosaic vegetation proposed by previous studies. More humid climate suitable for woodland at this time is supported by O-isotope analysis carried out in the northern Iran, which showed a decrease in aridity during 9.6–7.6 Ma (Ballato et al. 2010).
Although the studied interval is short, the application of facies analysis indicates that deposition in fluvial channels was controlled by ephemeral flash streams, indicating a seasonal climate. Distinct roots in the palaeosols suggest the presence of trees but the more mature palaeosols are only infrequently interbedded in the succession and clear evidence of trees has not been discovered from other intervals. From the sedimentological point of view, the distribution of tree cover may not have been extensive around the study site.
Palaeontology and palaeoecology
Mirzaie Ataabadi et al. (2013) provided an updated account of the mammalian species reported from Maragheh Fm. This faunal list is further updated (Mirzaie Ataabadi and Fortelius 2016, this issue) according to recent research in the systematic studies of Maragheh fossils (e.g. Solounias and Danowitz 2016, this issue). Bernor (1978, 1986) integrated all known stratigraphic records of fossil mammals to develop the first Maragheh mammalian biostratigraphy. He originally subdivided the Maragheh Formation into three units based on the stage-of-evolution of the Hipparion s.s. lineage: “Lower Maragheh” defined by the first occurrence of Hipparion gettyi at Kopran; “Middle Maragheh” by the first occurrence of Hipparion prostylum; and “Upper Maragheh” by the first occurrence of Hipparion campbelli. However, Bernor et al. (2016), this issue show that early Turolian Middle Maragheh horizon is better defined by occurrence of aff. Hippotherium brachypus and Cremohipparion aff. moldavicum. Indeed, Hipparion prostylum, originally defined based on skull morphology alone, may not occur at Maragheh. Solounias and Danowitz (2016), this issue have also revised the giraffid material of Maragheh and have increased the giraffid diversity significantly. Based on their analyses, the following genera including a new species should be included/updated in the Maragheh faunal list: Alcicephalus neumayri, Samotherium boissieri, Samotherium major and Honanotherium bernori n. sp.
Primate fossils
Before the discovery of the new primates from MMTT site II in Dareh Gorg, the only record of these animals was the cercopithecid colobine monkey (Mesopithecus pentelici) collected by Mecquenem (1924–25). The provenance of this specimen is not precisely known but as discussed by Bernor (1986), the best estimate for its occurrence is Middle Maragheh. The discovery of the new specimens from site II makes this estimate reliable.
The new primate fossils from Maragheh include the first hominoid catalogued as MMTT 3453 (Fig. 3a, b) and new Mesopithecus material (MMTT 3553, Fig. 3c, d). They are housed at the DOE/MMTT head quarters in Pardisan Park, Tehran, Iran. The new Mesopithecus specimen is a right mandibular fragment with the lower fourth premolar and first molar (p4 and m1). Compared to previous specimen (MNHN (Muséum National d’Histoire Naturelle, Paris)-MAR 3967), which is a complete lower jaw, this is not a significant discovery. However, due to its known provenance and co-occurrence with a large hominoid, it is remarkable. There is only the Azmaka locality in Bulgaria that yields possible contemporaneous occurrence of Mesopithecus and a hominoid (Spassov et al. 2012).
The Maragheh hominoid specimen is a left maxillary posterior alveolar fragment containing the upper second and third molars (M2 and M3) with well-preserved crowns and greater wear in the M2 than the M3 (Fig. 3a, b). Both M2 and M3 crowns are large-sized and are broader (buccolingually) than long. They are broadly comparable in size to those of Ouranopithecus macedoniensis and Sivapithecus parvada, and are larger than in Ankarapithecus meteai. One characteristic of MMTT 3453 is the M3 > M2 size relationship, especially in buccolingual dimension. This condition is not matched in known O. macedoniensis and A. meteai individuals, and it is more expressed in the single specimen of Ouranopithecus turkae.
O. turkae is probably temporally the closest to the Maragheh hominoid (Fig. 8). Despite its temporal and geographical proximity to the Iranian hominoid, O. turkae differs from the Maragheh hominoid considerably. Although the two share a M3 > M2 size relationship, but this condition is much more exaggerated in O. turkae, which also shows a suite of other derived features probably associated with enhanced postcanine mastication (Güleç et al. 2007). These features are not seen in the Maragheh hominoid. The general crown size and shape similarities of Maragheh hominoid with O. macedoniensis are greater than with O. turkae. However, O. macedoniensis also appears to express postcanine enhancement somewhat more so than MMTT 3453. The tendency for a more horizontal occlusal wear in O. macedoniensis suggests a somewhat stronger emphasis of postcanine crushing function in that species. To the contrary, the M3 size dominance seen in MMTT 3453 is not matched in O. macedoniensis.

Chronology and stratigraphic position of the hominoid/Mesopithecus-bearing localities within the eastern Mediterranean region (modified after Koufos et al. 2016). Maragheh here refers to hominoid level of Middle Maragheh interval. The estimated ages of the localities are from Koufos et al. (2016), Kaya et al. (2016), Spassov et al. (2012), and Kappelman et al. (2003b). ELMA, European Land Mammal Age
If the Maragheh hominoid was phylogenetically related to O. macedoniensis, the two might have diverged earlier in the Vallesian, prior to the known time range of O. macedoniensis (Fig. 8). Such a scenario may imply a parallel acquisition of large postcanine size at the eastern and western ends of the emerging Pikermian palaeobiome. In such a scenario, some presumably derived postcanine features characteristic of O. macedoniensis would have been secondarily modified in MMTT 3453. Given the subtle morphological features under consideration, such a transition is not impossible. However, the broadly contemporary presence of O. turkae that seemingly enhanced the O. macedoniensis-like masticatory adaptation makes such an opposite transition somewhat unlikely.
Considering MMTT 3453 as a phylogenetic descendent of the geographically more proximate mid Vallesian A. meteai may also necessitate a secondary shift towards a dietary adaptation with somewhat less emphasis in crushing, this despite an increase in postcanine size. Again, given the subtle features under consideration and small known sample sizes of A. meteai (n = 2), this hypothesis maybe less complicated in terms of morphological transitions. However, it would entail a postcanine size increase to have taken place within the Ankarapithecus clade or in the A. meteai lineage. Geographically, this would have occurred within, or adjacent to the developing Pikermian palaeobiome. Finally, the morphological similarities with the Sivapithecus species caution that the Iranian hominoid may represent an instance of western range expansion of S. parvada or an earlier offshoot of the Sivapithecus clade (see Suwa et al. 2016, this issue, for further discussion).
Hominoid site faunal structure
As mentioned above, about 160 large fossil bones are present in situ at hominoid fossil site in Dareh Gorg, Maragheh (Figs. 2c, d and 5). Most of the identifiable bones (Number of Identifiable Specimens, NISP) belong to bovids, equids, giraffids and proboscideans, while rhinos, carnivores and primates are less common (Tables 1 and 2, Figs. 4 and 9a). About 40 % of the bones are unidentified due to lack of diagnostic features, poor preservation and breakage or sediment cover. Some of them are probably identifiable with more detailed investigation.

Number of identifiable specimens (NISP), minimum number of individuals (MNI) and community structure (ComS) at the hominoid locality in Maragheh (a–c). d–o Panels indicate the bovid body mass spectra based on three categories at different hominoid/Mesopithecus-bearing localities within the eastern Mediterranean region including Middle Maragheh interval and Maragheh hominoid level. n, number of bovid species in each locality
In terms of number of individuals present at the site (Minimum Number of Individuals, MNI), bovids are by far the most abundant, represented by at least 24 individuals (based on horn cores). Giraffids and equids are represented by at least three individuals each. Primates are represented by two individuals, and proboscideans, rhinos and carnivores have at least one individual each (Table 2, Fig. 9b). Although a general bias towards large bones may exist in the site, and identification of the bones, especially the postcranials, should be regarded as preliminary here, one cannot neglect the sheer abundance of bovids in the site (Fig. 9c). Generally it is the hipparionine horses that are the most abundant (by NISP), both in many Maragheh sites and in other faunas of Iran (Mirzaie Ataabadi et al. 2011a, b). Thus, the observed predominance of bovids in the site likely reflects some special aspects of the palaeoecology and palaeoenvironment.
Kostopoulos and Bernor (2011) showed that (Middle) Maragheh, as well as Pikermi, is dominated by small-sized bovid taxa, under 50 kg body weight (Fig. 9o here), while in Samos, another classic Pikermian site, the majority of bovids are medium-sized (50–150 kg body weight). In their view, the differences in the bovid body size spectra indicate that environmental conditions were similar in Maragheh and Pikermi compared to Samos (Mirzaie Ataabadi et al. 2013). Presence of a cercopithecid monkey Mesopithecus in Maragheh and Pikermi and its absence from Samos might also be related to such environmental differences (Kostopolous and Bernor 2011).
A similar bovid body mass spectrum is also present in the Maragheh hominoid level (Fig. 9n). Occurrence of Mesopithecus at this site together with the hominoid fossil might reflect special palaeoecological conditions suitable for primates within the Pikermian palaeobiome. To explore this hypothesis, we compared the distribution of bovids in three body mass categories (less than 50 kg, 50–150 kg, and more than 150 kg) in primate-bearing Pikermian localities. These localities, which are also used here for other analyses and comparisons, include the following: Greek MN10 localities Xirochori, Ravin de la Pluie and Nikiti 1 which contain Ouranopithecus macedoniensis (Koufos 2006; Koufos and de Bonis 2006); Turkish MN 10 locality Sinap 12 preserving hominoid Ankarapithecus meteai (Kappelman et al. 2003a) and MN11 Çorakyerler site containing O. turkae (Güleç et al. 2007; Kaya et al. 2016); MN10 Georgian locality Udabno preserving the dryopithecine Udabnopithecus garedziensis (Gabunia et al. 2001) and MN12 Bulgarian locality Azmaka which has recently reported to include a hominoid (Spassov et al. 2012). We also included another Greek hominoid (Graecopithecus freybergi) site (Pyrgos Vassilissis) even though it has a longer time span (late Miocene) and short faunal list (Koufos and de Bonis 2005; Fortelius 2016). The Greek MN11 localities Nikiti 2 and Ravin des Zouaves are also included here due to presence of Mesopithecus in them and their role as evidence for the so-called ape-monkey replacement event in Greece (Koufos et al. 2016). We do not include the Sivapithecus and Indopithecus localities from Siwaliks and south/southeast Asia as their closed habitats make them a priori different from the open-adapted faunas of the Pikermian palaeobiome. Nevertheless, we have used some of these localities in our faunal similarity and community structure analyses to show this contrast (Figs. 10a–c, and 11a, b).

Community structure based on percentage of species present in each family in different hominoid/Mesopithecus-bearing localities within the eastern Mediterranean region and south/southeast Asia, as well as Middle Maragheh interval and Maragheh hominoid level

Pair-wise comparisons, based on genus-level faunal resemblance indices (GFRI), of Maragheh hominoid level (a) and Middle Maragheh interval (b) with different hominoid/Mesopithecus-bearing localities within the eastern Mediterranean region and south/southeast Asia. c Crown height diagrams for Maragheh, Middle Maragheh and Maragheh hominoid level as well as hominoid/Mesopithecus-bearing localities within the eastern Mediterranean region
Bovid body mass patterns
Distribution of the three bovid body mass categories in hominoid and some Mesopithecus-bearing localities in Vallesian/Turolian of eastern Mediterranean region is shown in Fig. 9. Similar bovid body mass spectra to Middle Maragheh and Maragheh hominoid level are seen in MN11/MN12 localities of Greece (Nikiti 2, Fig. 9i and Ravin des Zouaves, Fig. 9g), Turkey (Çorakyerler, Fig. 9l) and Bulgaria (Azmaka, Fig. 9j). Among these localities, Çorakyerler and Azmaka are hominoid-bearing and Azmaka also includes Mesopithecus.
The MN10 hominoid localities do not appear particularly similar in this comparison. One likely reason is the small number of bovid taxa in these faunas (two to four species), too few to show meaningful spectra. Among these localities, Sinap 12 (Fig. 9k) and Xirochori 1 (Fig. 9f) are dominated by medium- to large-sized bovids. This is in contrast with MN11/12 sites, which are dominated by small-sized bovids. Ravin de la Pluie (Fig. 9e) and Udabno I (Fig. 9m) have equal numbers of small- and medium/large-sized bovids while Pyrgos Vassilissis (late Miocene, Fig. 9d) and Nikiti 1 (Fig. 9e) are dominated by small bovids. Thus, no distinct pattern is observed among the hominoid-bearing localities of the Balkan-Iranian palaeoprovince regarding the distribution of bovid body mass categories.
However, the pattern seen in Maragheh hominoid level is also well expressed in other MN11/12 localities, indicating that this pattern could be related to the development of the Pikermian chronofauna, which peaks in the Turolian (Eronen et al. 2009). Other Pikermian sites, like Samos, have bovid body mass patterns dominated by large- and medium-sized bovids. Absence of primates in these localities might indicate some palaeoecological setting related to distribution and life of primates (Kostopoulos and Bernor 2011). Similar patterns observed in late Vallesian sites Ravin de la Pluie and Sinap 12 indicate their proto-Pikermian character.
Community structure
The community structure of Maragheh hominoid level also reflects the dominance of bovids, which represent 46 % of genera present (Figs. 9c and 10o). Compared to the 35 % observed for the Middle Maragheh interval as a whole, the dominance of bovids is moderately elevated (Fig. 10l) and may, along with other evidence, support an environmental difference.
Comparison of community structure among hominoid-bearing localities of the Balkan-Iranian palaeoprovince (Fig. 10) shows that the Turolian (MN11/12) localities Çorakyerler (Fig. 10n), Nikiti 2 (Fig. 10i), Ravin des Zouaves (Fig. 10j), Azmaka (Fig. 10k) and Middle Maragheh (Fig. 10l) are dominated by bovids, equids, giraffids and rhinocerotids. Only at Azmaka is a less pronounced predominance of these taxa observed, due to decrease in the percentage of bovids and increase of proboscideans. These localities also share a more even distribution of families. This can be distinguished simply in the pie charts, except for Azmaka. The similar community structure of these Turolian localities indicates less variability than in Vallesian localities described above.
More similarity in community structure of Middle Maragheh with Çorakyerler and Azmaka might be in more environmental similarity of these localities. They have a common cervid (Pliocervus) which is absent in Nikiti 2. This locality is probably drier and has Mesopithecus, even though no big environmental change has been seen in MN10-11 transition in Nikiti and its faunas. Drier conditions in Maragheh are also indicated by predominance of mesodont and hypsodont species (Fig. 11c) supporting the view that the Pikermian primates were capable of existing in regions characterised by overall dry conditions (Kaya et al. 2016).
The community structure in Udabno I (Fig. 10d) however shows a difference from other Pikermian faunas. Bovids are less dominant and there are more close-adapted taxa such as tragulids, cervids, suids and dryopithecin primates. The mean hypsodonty value of Udabno suggests more humid environmental conditions than for the Ouranopithecus localities (Kaya et al. 2016 and Fig. 11c here). A more humid and forested environment favourable for the highly arboreal Dryopithecus (Merceron et al. 2007) is plausible for this site.
Similarly, the greater abundance of close-adapted taxa in localities in south/southeast Asia (Fig. 10a–c) indicates more forested environmental conditions in these localities compared to the open-adapted Pikermian chronofauna (Mirzaie Ataabadi 2010). Studies show closed forested habitats for hominoids in Siwaliks and southeast Asia (Scott et al. 1999; Nelson 2007).
In general, the pattern of community structure seen in the hominoid-bearing localities of the Balkan-Iranian palaeo-province conforms to the patterns observed in the bovid body mass spectra. They suggest more similar distribution of bovid mass categories and mammalian families in the Turolian, while more varied bovid mass categories and less diverse community structure is present in the Vallesian. Overall, this pattern is a reflection of the rise and culmination of the Pikermian chronofauna during the Turolian. The proto-Pikermian characteristic of MN 10 localities suggests that open-country, seasonal conditions likely started to develop in MN10 in Greece and Anatolia (Sinap 12). No significant differences are seen between hominoid-bearing localities in Vallesian or Turolian, nor between hominoid sites and Mesopithecus sites.
Faunal similarity
The Greek localities Ravin de la Pluie and Xirochori 1 (MN 9-10) compared to Maragheh hominoid level show the highest similarity with Dice and Simpson GFRI of 40–50 %. Nikiti 1 has similarity of 35–40 % while Udabno is 28–30 % and Sinap (12) 28–40 %. The Siwaliks at this time show little similarity (10–20 %), mainly due to the co-occurrence of wide-ranging genera like Gazella and Hipparion. During early Turolian (MN11), the similarity of Greek locality Nikiti 2 and Ravin des Zouaves with Maragheh hominoid level is higher than in the Vallesian (45–70 %). The similarity with the hominoid-bearing MN11 locality Çorakyerler in Turkey is 36–60 %.
The only MN12 locality containing hominoids other than Maragheh is Azmaka in Bulgaria, which shows moderate to high levels of similarity with Maragheh hominoid level (35–70 %). Beside the similar age of this locality with Maragheh hominoid level, they are also the only Late Miocene localities in Balkan-Iranian palaeoprovince with co-occurrence of a hominoid with Mesopithecus.
Figure 11a, b illustrates the plot of GFRI in pair-wise comparisons between Middle Maragheh and Maragheh hominoid level and 12 other localities under consideration. Among the Vallesian localities, compared to Middle Maragheh, the Late Miocene Siwaliks containing the hominoid Sivapithecus show the lowest GFRIs. Among the late Vallesian (MN 10) Greek localities, the highest similarity with Middle Maragheh is present in Nikiti 1 with 32–50 % GFRIs. Sinap loc.12 (hominid zone) is similar to Nikiti 1 in showing GFRI of 32–45 % to Middle Maragheh. The Udabno locality in Georgia is close to these two localities by demonstrating 28–50 % similarity. The similarity of other Greek localities (Ravin de la Pluie and Xirochori 1) is about 20 %, less than the other Greek sites compared.
In the early Turolian (MN11), the highest similarity with Middle Maragheh is exhibited by the Turkish locality of Çorakyerler and Ravin des Zouaves in Greece, with similarities of more than 50 % for both indices. There are 14–15 genera in common between these localities. Nikiti 2 in Greece (MN11) also shows high similarity with Middle Maragheh (42–60 %). There are 10 common genera between these localities. The only MN12 hominoid-bearing locality (Azmaka, Bulgaria) also shows high similarity of more than 40 % with Middle Maragheh. They share 13 common genera with each other.
Similar to other analyses conducted here, the FRI analyses show that the similarity of Middle Maragheh in general and Maragheh hominoid level are more to Turolian localities (MN11-12) like Çorakyerler, Nikiti 2 and Azmaka. For Middle Maragheh, both Dice and Simpson indices indicate this high similarity while for hominoid level Simpson index demonstrates the high similarities. This is indeed due to shorter faunal list of Maragheh hominoid level.
Discussion
The assumed replacement of Vallesian hominoids by the cercopithecoid Mesopithecus in the eastern Mediterranean has been seen as an event similar to the earlier demise of hominoids in mainland Europe. Evidence for such a replacement has been seen in northern Greece where localities Ravin de la Pluie/Ravin des Zouves and Nikiti 1/Nikiti 2 are located (Koufos et al. 2016). The spread of more homogenous environments and increasingly seasonal climate in the late Miocene (Fortelius et al. 2002; Eronen and Rook 2004) were thought to be the reasons behind this assumed replacement (Merceron et al. 2010).
However, no major climatic event and vegetation change has been detected in Vallesian/Turolian transition in northern Greece to explain such patterns of species turnover and replacements. Also, recent discovery of large apes in typical Turolian localities show that this proposed ape extinction is not so abrupt in eastern Mediterranean region (Güleç et al. 2007; Spassov et al. 2012; Merceron et al. 2013). Indeed, the Vallesian/Turolian boundary in northern Greece shows climate conditions similar to the present-day Mediterranean region, with high seasonality and pure C3 vegetation (Merceron et al. 2013; Koufos et al. 2016). Bovid palaeodiet and isotope analyses in this area also indicate open landscape with abundant grass in a rich herbaceous layer. There is even evidence for mild cold seasons, unsuitable for present-day apes but not intolerable for monkeys. Therefore, reconstructed climate and environment are quite similar for Ouranopithecus and Mesopithecus, suggesting that they could have occurred in ecological sympatry.
Such ecological overlap is supported by recent research. The dental morphology of the eastern Mediterranean hominoid genus Ouranopithecus (mostly Vallesian) suggests ingestion of hard and abrasive dietary components. Its large body size implies terrestrial foraging for at least a portion of its niche space (Bernor 2007). Thus, Ouranopithecus may have occupied a range of fairly dry habitats, from woodlands to more open regions with a number of trees, bushes and thick grass (Merceron et al. 2005; Spassov et al. 2012; de Bonis and Koufos 2014). Reconstructing the palaeoenvironment in Ouranopithecus-bearing locality Ravin de la Pluie shows open landscape with wealthy herbaceous layer and low tree cover (Merceron et al. 2007).
The living habitat of the cercopithecid Mesopithecus (mostly Turolian) also appears to have ranged from terrestrial to arboreal (Youlatos and Koufos 2010) and its palaeodiet shows inclusion of hard/brittle objects like seeds (Merceron et al. 2009). The palaeoenvironment of this genus was open and low covered with sparse shrub/bushland and grassy cover (Youlatos 2003). Mesowear study of herbivores from Mesopithecus-bearing localities in Bulgaria suggests similar slightly wooded homogenous landscape with developed grassy herbaceous layer (Clavel et al. 2012).
Nevertheless, the climate did become warmer and drier across the Vallesian/Turolian transition and afterwards (Rey et al. 2013). Palaeobotanical data show decline of thermophilous, water-dependent taxa and increase in mesothermic, seasonal adapted taxa (Jiménez-Moreno et al. 2007). The Vallesian/Turolian transition as evidenced in Nikiti fauna shows a slight increase in dryness in MN11 (Nikiti 2) compared to the hominoid-bearing (MN 10) Nikiti 1 (Koufos et al. 2016). Our crown height analyses (Fig. 11c) also indicates such a subtle change.
Therefore, it seems that there was no abrupt change to affect hominoids in the eastern Mediterranean region. Our analyses of community structure and bovid body mass distribution show that the difference between Vallesian and Turolian faunas was not sharp. Koufos et al. (2016) showed that important structural reorganisation of the mammalian fauna happened near this boundary in northern Greece. This was expressed by doubling of meso-herbivore diversity and the proportion of intermediate feeders. The different patterns seen in community structure between the early and late Vallesian can thus be explained. This reorganisation very well demonstrates the development of Pikermian chronofauna and thus these Vallesian sites can be regarded as proto-Pikermian in nature, as was previously shown for the Anatolian Sinap 12 locality (Mirzaie Ataabadi et al. 2013).
This reorganisation has been thought to include replacement of large hominoids with Mesopithecus in northern Greece, but recent discoveries (Güleç et al. 2007; Spassov et al. 2012) and the current study (Suwa et al. 2016, this issue) cast doubt on this. Not only did large hominoids survive into the Turolian but they also co-existed with Mesopithecus in at least two localities (Middle Maragheh and Azmaka). Comparison of Turolian hominoid sites with Maragheh show more similarity of this site with Çorakyerler than Azmaka (Fig. 11a, b). This is probably due to more closed-adapted taxa present in the later site (Fig. 10k). The Azmaka environment is presented as a forest savanna/open woodland mosaic with seasonal precipitation and bushland/woodland vegetation (Spassov et al. 2012). The crown height distribution of brachydont, mesodont and hypsodont taxa in this locality (Fig. 11c) also indicates this difference with presence of more brachydont taxa (60 %) in Azmaka. Similar conditions occur in Pikermi, where 60 % brachydont taxa are present (Mirzaie Ataabadi et al. 2013). Maragheh in contrast show dominance of mesodont and hypsodont taxa. Çorakyerler is intermediate with more brachydont taxa than in Maragheh but less than in Azmaka. According to the mean hypsodonty proxy (Fortelius et al. 2014; Kaya et al. 2016), central Anatolia was a more arid region throughout the late Miocene than was eastern Europe (Fig. 11c here).
This indicates that environmental conditions in Greece and Bulgaria were more or slightly more humid than in Maragheh and Anatolia in the late Miocene (MN12). Previous studies (Kostopoulos 2009; Kostopoulos and Bernor 2011; Mirzaie Ataabadi et al. 2013) showed that conditions towards the centre of the sub-Paratethyan province may have been different from those at the eastern and western edges of this area. Thus, localities like Çorakyerler and its fauna might reflect a period of increased provincial endemism in central Anatolia during the late Vallesian to early Turolian (Geraads and Güleç 1999). Enhanced postcanine mastication seen in the hominoid O. turkae from Çorakyerler (Güleç et al. 2007), a trait probably missing in the Maragheh hominoid (Suwa et al. 2016, this issue) and less pronounced in O. macedoniensis from northern Greece, might reflect such differences in local setting.
While the anatomy may suggest a more mesic character for the Maragheh hominoid than for its counterpart at Çorakyerler, one should avoid pressing the evidence. Today, chimpanzees are well known to have a wide range of habitat preferences (equatorial forest to woodland savanna). Like them, these late Miocene hominoids might also have exhibited plasticity in their habitat preferences (Merceron et al. 2007). Combined with the rarity at Maragheh of both Mesopithecus and the hominoid, it is possible that small patches of relatively mesic woodlands were partially sampled at some of the localities. Such evaluations may be tested by future more in-depth studies after further fossil cleaning and excavations proceed.
Campbell et al. (1980) made much of the apparent absence of hominoid primates at Maragheh and in “Pontian” faunas generally. The discovery of hominoid remains at Maragheh nicely rounds off a trend of re-interpretation of both the nature of the Pikermian palaeobiome and the ecology of late Miocene hominoids. Contrary to the earlier views and somewhat counter intuitively, it now appears that the open biomes of the Balkan-Iranian palaeoprovince in fact included refuges for hominoids long after their forest-living relatives had disappeared from mainland Europe at the end of the Vallesian (Casanovas-Vilar et al. 2011). This view entails the re-assessment of late Miocene hominoid ecology to have possibly contained terrestrial, open-adapted forms as well as forest-adapted ones. It also suggests that the extinction of these ecologically differentiated hominoids had different causes in different biomes and that the expectation of synchrononous extinction of hominoids across geographic regions is unwarranted.
Conclusions
Discovery of a fossil hominoid from a classical Pikermian locality has increased the stratigraphic and geographic range of these primates in the late Miocene of western Eurasia. Survival of large apes at the Vallesian/Turolian boundary is now evident from three localities in eastern Mediterranean region. The co-occurrence of Mesopithecus with a large ape in Middle Maragheh is significant because it confirms the optional sympatry known previously only from a single locality (Azmaka in Bulgaria). The Turolian record of hominoids from the Balkan-Iranian palaeoprovince and the study of their accompanying faunas indicate a general high faunal and structural similarity, possibly overlain by a palaeoenvironmental gradient from a more humid west to a drier east. Presence of hominoids and Mesopithecus in localities at the eastern and western ends of this province with apparent faunal differences suggests that such co-occurrence was due to locally rather than regionally favourable conditions. Alternatively, late Miocene hominoids may have been ecologically more flexible than has been appreciated.
References
Ballato, P., Mulch, A., Landgraf, A., Strecker, M. R., Dalconi, M. C., Friedrich, A., & Tabatabaei, S. H. (2010). Middle to late Miocene Middle Eastern climate from stable oxygen and carbon isotope data, southern Alborz mountains, N Iran. Earth and Planetary Science Letters, 300, 125–138.
Barry, J. C., Morgan, M. E., Flynn, L. J., Pilbeam, D., Behrensmeyer, A. K., Raza, S. M., Khan, I. A., Badgley, C., Hicks, J., & Kelley, J. (2002). Faunal and environmental change in the late Miocene Siwaliks of northern Pakistan. Paleobiology, 28, 1–71.
Bernor, R. L. (1978). The mammalian systematics, biostratigraphy and biochronology of Maragheh and its importance for understanding late Miocene hominoid zoogeography and evolution. Los Angeles: University of California Dissertation.
Bernor, R. L. (1986). Mammalian biostratigraphy, geochronology and zoogeographic relationships of the late Miocene Maragheh fauna, Iran. Journal of Vertebrate Paleontology, 6(1), 76–95.
Bernor, R. L. (2007). New apes fill the gap. Proceedings of the National Academy of Sciences of the United States of America, 104(50), 19661–19662.
Bernor, R. L., Woodburne, M. O., & Van Couvering, J. A. (1980). A contribution to the chronology of some Old World faunas based on hipparionine horses. Geobios, 13(5), 25–59.
Bernor, R. L., Fortelius, M., & Rook, L. (2001). Evolutionary biogeography and paleoecology of the Oreopithecus bambolii Faunal Zone (late Miocene, Tusco-Sardinian Province). Bolletino della Società Paleontologica Italiana, 40(2), 139–148.
Bernor, R. L., Semprebon, G. M., & Damuth, J. (2014). Maragheh ungulate mesowear: interpreting paleodiet and paleoecology from a diverse fauna with restricted sample sizes. Annales Zoologici Fennici, 51, 201–208.
Bernor, R., Mirzai Ataabadi M, Meshida, K., & Wolf, D. (2016). The Maragheh hipparions, late Miocene of Azarbaijan, Iran. In M. Mirzaie Ataabadi and M. Fortelius (Eds.) The late Miocene Maragheh mammal fauna; results of recent multidisciplinary research. Palaeobiodiversity and Palaeoenvironments, 96(3). doi: 10.1007/s12549-016-0235-2.
Campbell, B. G., Amini, M. H., Bernor, R. L., Dickinson, W., Drake, R., Morris, R., Van Couvering, J. A., & Van Couvering, J. A. H. (1980). Maragheh: a classical late Miocene vertebrate locality in northwestern Iran. Nature, 287, 837–841.
Casanovas-Vilar, I., Alba, D. M., Garces, M., Robles, J. M., & Moya-Sola, S. (2011). Updated chronology for the Miocene hominoid radiation in Western Eurasia. Proceedings of the National Academy of Sciences of the United States of America, 108, 5554–5559.
Clavel, J., Merceron, G., Hristova, L., Spassov, N., Kovachev, D., & Escarguel, G. (2012). On Mesopithecus habitat: insights from late Miocene fossil vertebrate localities of Bulgaria. Journal of Human Evolution, 63, 162–179.
de Bonis, L., & Koufos, G. (2014). First discovery of postcranial bones of Ouranopithecus macedoniensis (Primates, Hominoidea) from the late Miocene of Macedonia (Greece). Journal of Human Evolution, 74, 21–36.
Erdbrink, D. P. B., Priem, H. N. A., Hebeda, E. H., Cup, C., Dankers, P. O., & Cloetingh, S. A. P. L. (1976). The bone bearing beds near Maragheh in NW Iran. Proceedings Koninklijke Nederlandse Akademie van Wetenschappen B, 79(2), 85–113.
Eronen, J. T., & Rook, L. (2004). The Mio-Pliocene European primate fossil record: dynamics and habitat tracking. Journal of Human Evolution, 47, 323–341.
Eronen, J. T., Mirzaie Ataabadi, M., Micheels, A., Karme, A., Bernor, R. L., & Fortelius, M. (2009). Distribution history and climatic controls of the Late Miocene Pikermian chronofauna. Proceedings of the National Academy of Sciences of the United States of America, 106, 11867–11871.
Fortelius, M., coordinator. (2016). New and Old World Database of Fossil Mammals (NOW). University of Helsinki. http://www.helsinki.fi/science/now/.
Fortelius, M., Werdelin, L., Andrews, P., Bernor, R. L., Gentry, A., Humphrey, L., Mittmann, H. W., & Viranta, S. (1996). Provinciality, diversity, turnover, and paleoecology in land mammal faunas of the later Miocene of Western Eurasia. In R. L. Bernor, V. Fahlbusch, & H. W. Mittmann (Eds.), The evolution of western Eurasian Neogene mammal faunas (pp. 414–448). New York: Columbia University Press.
Fortelius, M., Eronen, J. T., Jernvall, J., Liu, L., Pushkina, D., & Rinne, J. (2002). Fossil mammals resolve regional patterns of Eurasian climate change during 20 million years. Evolutionary Ecology Research, 4, 1005–1016.
Fortelius, M., Eronen, J. T., Kaya, F., Tanq, H., & Raia, P. (2014). Evolution of Neogene mammals in Eurasia: environmental forcing and biotic interactions. Annual Review of Earth and Planetary Sciences, 42, 579–604.
Gabunia, L., Gabashvili, E., Vekua, A., & Lordkipanidze, D. (2001). The late Miocene hominoid from Georgia. In L. de Bonis, G. D. Koufos, & P. Andrews (Eds.), Hominoid evolution and climatic change in Europe volume 2: phylogeny of the Neogene hominoid primates of Eurasia (pp. 316–325). Cambridge: Cambridge University Press.
Geraads, D., & Güleç, E. (1999). On some spiral-horned antelopes (Mammalia: Artiodactyla: Bovidae) from the late Miocene of Turkey, with remarks on their distribution. Paläontologische Zeitschrift, 73, 403–409.
Gradsttein, F. M., Ogg, J. G., Schmiz, M. D., & Ogg, D. B. (2012). The geological time scale (Vol. 1). Boston: Elsevier.
Güleç, E., Sevim, A., Pehlevan, C., & Kaya, F. (2007). A new great ape from the late Miocene of Turkey. Anthropological Science, 115, 153–158.
Jiménez-Moreno, G., Popescu, S.-M., Ivanov, D., & Suc, J.-P. (2007). Neogene flora vegetation and climate dynamics in southeastern Europe and the northeastern Mediterraenan. In M. Williams, A. M. Haywood, F. J. Gregory, & D. N. Schmidt (Eds.), Deep-time perspectives on climate change: marrying the signal from computer models and biological proxies (pp. 503–516). London: The Micropalaeontological Society.
Kamei, T., Ikeda, J., Ishida, H., Ishida, S., Onishi, I., Partoazar, H., Sasajima, S., & Nishimura, S. (1977). A general report of the geological and paleontological survey in Maragheh area, north-west Iran, 1973. Memoirs of the Faculty of Science, Kyoto Universoty Series Geol. and Mineral., 43, 131–164.
Kappelman, J., Richmond, B. G., Seiffert, E. R., Maga, A. M., & Ryan, T. M. (2003a). Hominoidea (Primates). In M. Fortelius, J. Kappelman, S. Sen, & R. Bernor (Eds.), Geology and paleontology of the Miocene Sinap Formation, Turkey (pp. 42–66). New York: Columbia University Press.
Kappelman, J., Duncan, A., Feseha, J., Lunkka, J. P., et al. (2003b). Chronology. In M. Fortelius, J. Kappelman, S. Sen, & R. Bernor (Eds.), Geology and paleontology of the Miocene Sinap Formation, Turkey (pp. 90–124). New York: Columbia University Press.
Kaya, F., Kaymakçi, N., Bibi, F., Eronen, J. T., Pehlevan, C., Erkman, A. C., Langereis, C. G., & Fortelius, M. (2016). Magnetostratigraphy and paleoecology of the hominid-bearing locality Çorakyerler, Tuǧlu Formation (Çankiri Basin, Central Anatolia). Journal of Vertebrate Paleontology, 36, e1071710. pp.12.
Kostopoulos, D. S. (2009). The Pikermian Event: temporal and spatial resolution of the Turolian large mammal fauna in SE Europe. Palaeogeography, Palaeoclimatology, Palaeoecology, 274(1–2), 82–95.
Kostopoulos, D. S., & Bernor, R. L. (2011). The Maragheh bovids (Mammalia, Artiodactyla): systematic revision and biostratigraphic-zoogeographic interpretation. Geodiversitas, 33, 649–708.
Koufos, G. D. (2006). The Neogene mammal localities of Greece: faunas, chronology and biostratigraphy. Hellenic Journal of Geosciences, 41, 183–214.
Koufos, G. D., & de Bonis, L. (2005). The Late Miocene hominoids Ouranopithecus and Graecopithecus. Implications about their relationships and taxonomy. Annales de Paléontologie, 91, 227–240.
Koufos, G. D., & de Bonis, L. (2006). New material of Ouranopithecus macedoniensis from late Miocene of Macedonia (Greece) and study of its dental attrition. Geobios, 39, 223–243.
Koufos, G. D., Kostopoulos, D. S., Vlachou, T. D., & Konidaris, G. E. (2016). Synthesis. In G. D. Koufos & D. S. Kostopoulos (Eds.), Palaeontology of the upper Miocene vertebrate localities of Nikiti (Chalkidiki Peninsula, Macedonia, Greece) (pp. 147–154). Lyon: Geobios. p. 49.
Mazza, P. P. A., & Ventra, D. (2011). Pleistocene debris-flow deposition of the hippopotamus-bearing Collecurti bone bed (Macerata, Central Italy): taphonomic and paleoenvironmental analysis. Palaeogeography, Palaeoclimatology, Palaeoecology, 310, 296–314.
Mecquenem, R. (1924–25). Contribution l’étude des fossiles de Maragha. Annales de Pa1éonto1ogie, 13/14 (1924–25), 135–160
Merceron, G., Blondel, C., de Bonis, L., Koufos, G. D., & Viriot, L. (2005). A new dental microwear analysis: application to extant primates and Ouranopithecus macedoniensis (late Miocene of Greece). Palaios, 20, 551–561.
Merceron, G., Schulz, E., Kordos, L., & Kaiser, T. M. (2007). Paleoenvironment of Dryopithecus brancoi at Rudabanya, Hungary: evidence from dental meso- and microwear analyses of large herbivorous mammals. Journal of Human Evolution, 53, 331–349.
Merceron, G., Scott, J., Scott, R. S., Geraads, D., Spassov, N., & Ungar, P. S. (2009). Folivory or fruit/seed predation for Mesopithecus, an earliest colobine from the late Miocene of Eurasia. Journal of Human Evolution, 57, 732–738.
Merceron, G., Kaiser, T. M., Kostopoulos, D. S., & Schulz, E. (2010). Ruminant diet and the Miocene extinction of European great apes. Proceedings of the Royal Society B, 277, 3105–3112.
Merceron, G., Kostopoulos, D. S., de Bonis, L., Fourel, F., Koufos, G. D., Le’cuyer, C., & Martineau, M. (2013). Stable isotope ecology of Miocene bovids from northern Greece and the ape/monkey turnover in the Balkans. Journal of Human Evolution, 65, 185–198.
Mirzaie Ataabadi, M. (2010). The Miocene of Western Asia; fossil mammals at the crossroads of faunal provinces and climate regimes. Helsinki: Helsinki University Print.
Mirzaie Ataabadi, M. & Fortelius, M. (2016) Introduction to the special issue “The late Miocene Maragheh mammal fauna; results of recent multidisciplinary research”. Palaeobiodiversity and Palaeoenvironments, 96(3). doi:10.1007/s12549-016-0242-3.
Mirzaie Ataabadi, M., Zaree, G., & Orak, Z. (2011a). Large mammals from the new Late Miocene fossil localities in Varzeghan area, northwest Iran. Vertebrata PalAsiatica, 49(3), 311–321.
Mirzaie Ataabadi, M., Mohammadalizadeh, J., Zhang, Z., Watabe, M., Kaakinen, A., & Fortelius, M. (2011b). Late Miocene Large mammals from Ivand (northwestern Iran). Geodiversitas, 33(4), 709–728.
Mirzaie Ataabadi, M., Bernor, R. L., Kostopoulos, D. S., Wolf, D., Orak, Z., Zare, G., Nakaya, H., Watabe, M., & Fortelius, M. (2013). Recent advances in paleobiological research of the Late Miocene Maragheh fauna, northwest Iran. In X. Wang, L. J. Flynn, & M. Fortelius (Eds.), Fossil mammals of Asia: Neogene biostratigraphy and chronology (pp. 546–565). New York: Columbia University Press.
Naruse, H., & Masuda, F. (2006). Visualization of the internal structure of the massive division in experimental sediment-gravity-flow deposits by mapping of grain fabric. Journal of Sedimentary Research, 76, 845–865.
Nelson, S. N. (2007). Isotopic reconstruction of habitat change surrounding the extinction of Sivapithecus, a Miocene hominoid, in the Siwalik Group of Pakistan. Palaeogeography, Palaeoclimatology, Palaeoecology, 243, 204–222.
Nemec, W., & Steel, R. J. (1984). Alluvial and coastal conglomerates: their significant features and some comments on gravelly mass flow deposits. In E. H. Koster & R. J. Steel (Eds.), Sedimentology of gravels and conglomerates (pp. 1–31). Toronto: Canadian Society of Petroleum Geologists Memoir 10.
Postma, G., Roep, T. B., & Ruegg, G. H. J. (1983). Sandy-gravelly mass flow deposits in an ice-margin lake (Saalian, Leuvenumsche Beek Valley, Veluwe, the Netherlands), with emphasis on plug flow deposits. Sedimentary Geology, 34, 59–82.
Pourabrishami, Z. (2005). Study, discovery, excavation and reconstruction of fossil vertebrates from Maragheh and Varzeghan. Department of Environment (DOE/MMTT): Tehran.
Rey, K., Amiot, R., Le’ cuyer, C., Koufos, G. D., Martineau, F., Fourel, F., Kostopoulos, D. S., & Merceron, G. (2013). Late Miocene climatic and environmental variations in northern Greece inferred from stable isotope compositions (O, C) of equid teeth apatite. Palaeogeography, Palaeoclimatology, Palaeoecology, 388, 48–57.
Sakai, T., Zaree, G.R., Sawada, Y., Mirzaie Ataabadi, M. & Fortelius, M. (2016) Depositional environment reconstruction of the Maragheh Formation, East Azarbaijan, Northwestern Iran. In M. Mirzaie Ataabadi and M. Fortelius (Eds.) The late Miocene Maragheh mammal fauna; results of recent multidisciplinary research. Palaeobiodiversity and Palaeoenvironments, 96(3). doi:10.1007/s12549-016-0238-z.
Salminen, J., Paknia, M., Kaakinen, A., Mirzaie Ataabadi, M., Zaree, G., Orak, Z. & Fortelius, M. (2016). Preliminary magnetostratigraphic results from the late Miocene Maragheh Formation, NW Iran. In M. Mirzaie Ataabadi and M. Fortelius (Eds.) The late Miocene Maragheh mammal fauna; results of recent multidisciplinary research. Palaeobiodiversity and Palaeoenvironments, 96(3). doi:10.1007/s12549-016-0239-y.
Sawada,Y., Zaree, G.R., Sakai, T., Itaya, T. Yagi, K., Imaizumi, M., Mirzaie Ataabadi, M., & Fortelius, M. (2016) K–Ar ages and petrology of the late Miocene pumices from the Maragheh Formation, northwest Iran. In M. Mirzaie Ataabadi and M. Fortelius (Eds.) The late Miocene Maragheh mammal fauna; results of recent multidisciplinary research. Palaeobiodiversity and Palaeo-environments, 96(3). doi:10.1007/s12549-016-0232-5.
Scott, R. S., Kappelman, J., & Kelley, J. (1999). The paleoenvironment of Sivapithecus parvada. Journal of Human Evolution, 36, 245–274.
Simpson, G. G. (1943). Mammals and the nature of continents. American Journal of Science, 241, 1–31.
Sokal, R. R., & Sneath, P. H. A. (1963). Principles of numerical taxonomy. San Francisco: Freeman.
Solounias, N., & Danowitz, M. (2016) The Giraffidae of Maragheh and the identification of a new species of Honanotherium. In M. Mirzaie Ataabadi and M. Fortelius (Eds.) The late Miocene Maragheh mammal fauna; results of recent multidisciplinary research. Palaeobiodiversity and Palaeoenvironments, 96(3). doi:10.1007/s12549-016-0230-7.
Spassov, N., Geraads, D., Hristova, L., Markov, G. N., Merceron, T., Stoyanov, K., & Dimitrova, A. (2012). A hominid tooth from Bulgaria: the last pre-human hominid of continental Europe. Journal of Human Evolution, 62, 138–145.
Strömberg, C. A. E., Werdelin, L., Friis, E. M., & Saraç, G. (2007). The spread of grass-dominated habitats in Turkey and surrounding areas during the Cenozoic: phytolith evidence. Palaeogeography, Palaeoclimatology, Palaeoecology, 250, 18–49.
Suwa, G., Kunimatsu, Y., Mirzai Ataabadi, M., Orak, Z., Sasaki, T. and Fortelius, M. (2016) The first hominoid from the Maragheh Formation, Iran. In M. Mirzaie Ataabadi and M. Fortelius (Eds.) The late Miocene Maragheh mammal fauna; results of recent multidisciplinary research. Palaeobiodiversity and Palaeo-environments, 96(3). doi: 10.1007/s12549-016-0234-3.
Swisher, C. C., III. (1996). New 40Ar/39Ar dates and their contribution toward a revised chronology for the Late Miocene of Europe and west Asia. In R. L. Bernor, V. Fahlbusch, & H. W. Mittmann (Eds.), The evolution of western Eurasian Neogene mammal faunas (pp. 64–77). New York: Columbia University Press.
Yamada, E., Hasumi, E., Miyazato, N., Akahoshi, M., Watabe, M., & Nakaya, H. (2016) Mesowear analyses of sympatric ungulates from the late Miocene Maragheh, Iran. In M. Mirzaie Ataabadi and M. Fortelius (Eds.) The late Miocene Maragheh mammal fauna; results of recent multidisciplinary research. Palaeobiodiversity and Palaeoenvironments, 96(3) doi: 10.1007/s12549-016-0237-0.
Youlatos, D. (2003). Calcaneal features of the Greek Miocene primate Mesopithecus pentelicus (Cercopithecoidea: Colobinae). Geobios, 36, 229–239.
Youlatos, D., & Koufos, G. D. (2010). Locomotor evolution of Mesopithecus (Primates, Colobinae) from Greece: evidence from selected astragalar characters. Primates, 51, 23–35.
Acknowledgements
The Department of Environment (environment protection organisation) of the Government of Iran permitted and facilitated this study. We would like to express our gratitude to the former and present heads of the “natural environment division” of this organisation as well as those of the “office of natural history museum, biodiversity and genetic resources” for their support. We also appreciate assistance by the Maragheh governor, mayor, city council, and the DOE office, and the members of the INSPE (International Sahand Palaeoenvironmental Expedition) team and participants of the field workshops held in Maragheh during 2007–2009. We specially thank Philippe Richir (MNHN, Paris), Zhou Wei (IVPP, Beijing), Pierre Mauries and Kari Lintulaakso (University of Helsinki) for their field assistance. MMA is grateful to Dimitris Kostopolous for providing bovid body mass data. We thank Raymond Bernor and Nikolai Spassov for their reviews and valuable comments. MMA, MF, AK and field work in Maragheh were partially supported by the Academy of Finland, RHOI project (NSF-HOMINID- BCS-0321893), Sasakawa Foundation, Nordenskiöld Samfundet and DOE/MMTT.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interests
The authors declare that no conflict of interests exists.
Additional information
This article is a contribution to the special issue "The late Miocene Maragheh mammal fauna; results of recent multidisciplinary research."
Rights and permissions
About this article

Cite this article
Ataabadi, M.M., Kaakinen, A., Kunimatsu, Y. et al. The late Miocene hominoid-bearing site in the Maragheh Formation, Northwest Iran. Palaeobio Palaeoenv 96, 349–371 (2016). https://doi.org/10.1007/s12549-016-0241-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-016-0241-4