
Abstract
During the IPY-CAML TAN0802 survey with RV Tangaroa, marine communities on the shelf and slope of the Ross Sea, as well as around the remote Scott Island and seamounts east of the Balleny Islands, were sampled using various sampling gears (epibenthic sled, rough-bottom fish trawl, beam trawl and multicorer). Amongst the numerous benthic samples obtained, an important collection of hydroids was present. Thirty-five species, including Sertularella pseudovervoorti sp. nov., have been recorded. Anthoathecata are represented only by Bouillonia denhartogi. The collection is essentially dominated by Leptothecata, with 34 species, belonging to the families Lafoeidae, Haleciidae, Kirchenpaueriidae, Schizotrichidae, Sertularellidae, Staurothecidae, Symplectoscyphidae, Tiarannidae and Zygophylacidae. Symplectoscyphidae is by far the most diversified family with 12 species (34%), followed by Kirchenpaueriidae and Staurothecidae with 5 each (14%) and Haleciidae with 4 species (11%). At the generic level, Symplectoscyphus with seven species, Antarctoscyphus, Oswaldella and Staurotheca with five species and Halecium with four are the most speciose genera. Symplectoscyphus frondosus is clearly the species with the highest occurrence. The depth range of 15 species is increased, considerably in most cases. Several species so far considered to be inhabitants of the Antarctic shelf clearly extend their distribution to the slope. Twenty species were found on the shelf and slope of the Ross Sea, with Stegopoma plicatile representing the new single record for the area, raising the number of known species to 78. Ten species (including the new species) come from the virtually unknown area of Scott Island, 8 constituting new records, raising the number of species known in this isolated area to 11. Sixty-eight percent of the species are Antarctic endemic and 94% are restricted to Antarctic or Antarctic/sub-Antarctic waters.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The recognition of International Polar Year (IPY) from March 2007 to March 2009 led to a large international collaborative research in both Antarctica and the Arctic. Thus, the TAN0802 trip by the R/V Tangaroa, which took placed between January and March 2008, was funded for a New Zealand Census of Antarctic Marine Life (CAML) project to support biodiversity studies in the Ross Sea region (Antarctica). For this IPY-CAML project, three areas of study (the Ross Sea, waters around Scott Island and seamounts east of the Balleny Islands) were considered, each of which was stratified by depth.
Benthic hydroids from the Ross Sea have recently been reviewed by Peña Cantero (2017), after studying the collection gathered during the New Zealand BioRoss 2004 survey with R.V. Tangaroa. The collection consisted of 61 species, of which 22 were new records in the area, so that the number of valid known species of benthic hydroids of the Ross Sea was raised to 77, making it one of the most diverse, and relatively well-known, Antarctic areas for benthic hydrozoans. In contrast, hydroids from the area around Scott Island are virtually unknown. Only three species (Stegolaria irregularis Totton, 1930, Staurotheca dichotoma Allman, 1888 and Schizotricha anderssoni Jäderholm, 1904), all from the same station, have been reported off this remote island located approximately 500 km northeast of Cape Adare (Naumov and Stepanjants 1962). As for the Balleny Islands, 25 species of benthic hydroids have been reported (Peña Cantero 2009).
I present here the results of the study of the collection of benthic hydroids of the Ross Sea and the area of Scott Island (no hydroids came from the seamounts east of the Balleny Islands) collected during the New Zealand TAN0802 voyage with R/V Tangaroa in order to complement the study on the hydroids of the Ross Sea by Peña Cantero (2017) and increase knowledge about the virtually unknown fauna of benthic hydroids that inhabit the area of Scott Island. The collection was particularly interesting both by the samples collected in the remote Scott Island area and by the bathymetric interval considered during the survey, extending from the shelf to the slope and, therefore, sampling depths little explored for Antarctic benthic hydroids, since most knowledge about them comes from shelf.
Material and methods
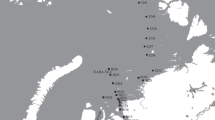
The collection studied here was gathered during the New Zealand IPY-CAML TAN0802 survey with RV Tangaroa, which took place from January to March 2008. Samples come from the shelf and slope of the Ross Sea and from bottoms around Scott Island (cf. Fig. 1 for the location of the sampling sites in which hydroids were found), and were collected with three different sampling gears (epibenthic sled, rough-bottom fish trawl and beam trawl). Thirty-one samples containing hydroids were collected at depths between 283 and 2283 m (cf. Table 1). Hydrozoans were fixed in 98% ethanol, and later transferred to 70% ethanol.

Area of study and location of sampling sites (see Table 1 for more details)
The material studied is deposited in the National Institute of Water and Atmospheric Research Invertebrate Collection at Wellington (NIWA), New Zealand.
Species of Leptothecata Cornelius, 1992 are mainly presented following a recent phylogenetic proposal (Maronna et al. 2016).
Benthic hydroids of the Ross Sea have recently been reviewed by Peña Cantero (2017), and species of Antarctoscyphus Peña Cantero, García Carrascosa and Vervoort, 1997 by Peña Cantero et al. (2017). I refer to these papers as links to previous references concerning the species; synonymy is only included for those species not considered in those studies. Descriptions are only provided for a species new to science described in this study and for Antarctoscyphus fragilis Peña Cantero, Svoboda & Vervoort, 1999, a particularly interesting species from a biogeographic point of view; the remaining species are well known and there are excellent descriptions in the literature, of which I provide references when dealing with each species. On the other hand, to facilitate the species identification, photographs of most of the species are included, as well as measurements for those species in which size is important for their identification; measurements are not given for species morphologically easy to identify, either because there is only one species in the genus [e.g. Abietinella operculata (Jäderholm, 1903)] or because they can be recognised directly from the photographs provided. Diagnostic characters are included only when they are necessary to confirm species identity. In the section on ecology and distribution, I focused on the data obtained in the present study, but I also provide information about the general bathymetric and geographic distribution of the species. For the biogeographic discussion, I follow the distribution patterns considered by Peña Cantero and García Carrascosa (1999).
Results
Taxonomic account
Superorder Anthoathecata Cornelius, 1992
Tubulariidae Allman, 1864
Bouillonia denhartogi Svoboda, Stepanjants and Ljubenkov, 2006
(Fig. 2)

Bouillonia denhartogi Svoboda, Stepanjants and Ljubenkov, 2006: polyp with gonophores. Scale bar: 5 mm
Bouillonia denhartogi Svoboda et al. 2006: 200–204, Figs. 2b–5, 9–12, 14–16, 17b–19, 20d, 24–28; Peña Cantero, 2012: 854, Fig. 2a, b.
Material examined: Stn 211, one polyp, 16 mm high, with blastostyles, on leg of King crab (NIWA 38226).
Remarks: For an extensive description of this species, I refer to the original description by Svoboda et al. (2006).
Ecology and distribution: Deep-water species, known from depths between 330 and 3450 m (Svoboda et al. 2006); present material collected from 855 to 879 m, epibiotic on a crab leg, and with gonophores in March.
Circum-Antarctic distribution (Peña Cantero 2012). Here reported from a seamount off Scott Island.
Superorder Leptothecata Cornelius, 1992
Order Lafoeida Bouillon, 1984
Family Lafoeidae Hincks, 1868
Filellum antarcticum (Hartlaub, 1904)
Material examined: Stn 144, numerous hydrothecae, with coppiniae, on Oswaldella stepanjantsae Peña Cantero, Svoboda and Vervoort, 1997 and Symplectoscyphus exochus Blanco, 1982 (NIWA 120291).
Measurements (in μm): Hydrothecae: diameter at aperture 100. Gonothecae: height 400, maximum diameter 110, diameter at aperture 60.
Remarks: For an extensive description of this species, also including type material and a SEM study of the coppinia, see Peña Cantero et al. (2004).
Ecology and distribution: Hitherto considered a shelf species (Peña Cantero et al. 1998), recorded from depths between 14 (Millard 1964, 1975) and 540 m (Peña Cantero 2012). Present material collected at depths from 1431 to 1658 m, considerably extending its lower bathymetric limit to bathyal depths, on O. stepanjantsae and S. exochus, and with coppiniae in February.
Mainly known from Antarctic waters, but also reported off South Africa (cf. Peña Cantero et al. 2004). Present material comes from the slope of the Ross Sea.
Order Laodiceida Maronna, Miranda, Peña Cantero, Barbeitos and Marques, 2016
Family Tiarannidae Russell, 1940
Stegopoma plicatile (M. Sars, 1863)
Stegopoma plicatile—Peña Cantero and Ramil, 2006: 952 (synonymy); Peña Cantero, 2012: 855–856; Soto Àngel and Peña Cantero, 2015: 986, Fig. 2i.
Material examined: Stn 144, one stem, 12 mm high, on O. stepanjantsae, and two stem fragments, up to 16 mm high, basibiont of Symplectoscyphus curvatus (Jäderholm, 1917) (NIWA 120292); Stn 206, two stems, 30 and 20 mm high, in very bad condition (NIWA 38169); Stn 210, several stems, up to 40 mm high, on gorgonian (NIWA 38190); Stn 237, one stem, 80 mm high (NIWA 38488); Stn 251, a few stems, up to 40 mm high, on axis of gorgonian, in very bad condition (NIWA 38660); Stn 256, several stems, up to 60 mm high, on coral, in bad condition (NIWA 38698).
Remarks: For a description of this species, I refer to Schuchert (2001).
Ecology and distribution: Deep-water species collected at depths from 385 (Vanhöffen 1910) to 1294 m (Peña Cantero 2012) in the Antarctic region; present material from 602 to 1658 m, extending its lower bathymetric limit in this region, epibiotic on coral, axis of gorgonian and O. stepanjantsae, and basibiont of S. curvatus.
Species with a worldwide distribution, known in Antarctic waters from both East and West Antarctica (cf. Peña Cantero and Ramil 2006). Here reported from the slope of the Ross Sea and from seamounts off Scott Island.
Order Macrocolonia Leclère, Schuchert, Cruaud, Couloux, Manuel, 2009
Incertae Sedis
Billardia subrufa (Jäderholm, 1904)
Material examined: Stn 17, a few stems, up to 90 mm high, with gonothecae, on coral (NIWA 35449).
Remarks: For a description of this species, I refer to Peña Cantero and García Carrascosa (1995).
Ecology and distribution: Eurybathic species (Peña Cantero et al. 2004), found at depths from 25 (Stepanjants 1972) to 1030 m (Peña Cantero et al. 2004); present material at 321 m, epibiotic on coral and with gonothecae in February.
Antarctic-Patagonian distribution (Peña Cantero et al. 2004). Present material from the continental shelf of the Ross Sea.
Staurothecidae Maronna, Miranda, Peña Cantero, Barbeitos and Marques, 2016
Staurotheca antarctica Hartlaub, 1904
(Fig. 3a)

Staurotheca antarctica Hartlaub, 1904: a hydrotheca. Staurotheca dichotoma Allman, 1888: b, c hydrothecae. Staurotheca frigida Peña Cantero, Svoboda and Vervoort, 1997: d, e hydrothecae. Staurotheca nonscripta Peña Cantero, Svoboda and Vervoort, 1997: f hydrotheca. Staurotheca pachyclada (Jäderholm, 1904): g, h hydrothecae. Scale bar: 200 μm (a–d, f–h), 100 μm (e)
Staurotheca antarctica—Peña Cantero and Vervoort, 2003: 2670–2673, Fig. 3 (synonymy); Peña Cantero, 2012: 858–859; 2013: 131; 2014a: 1722, Fig. 5a.
Material examined: Stn 46, a mass of stems, 10 mm in diameter, with female gonothecae (NIWA 36033); Stn 117, a colony fragment, 60 mm long (NIWA 120293).
Measurements (in μm): Hydrothecae: abcauline wall 450, free part of adcauline wall 130–250, adnate part of adcauline wall 450, adcauline wall 580–700, diameter at aperture 160.
Remarks: For an extensive description of this species, including the type series, I refer to Peña Cantero et al. (1997a).
Ecology and distribution: Hitherto considered a shelf species (Peña Cantero and Vervoort 2003), it had been reported from depths between 55 (Peña Cantero and Vervoort 2003) and 708 m (Peña Cantero 2014a); present material collected from 475 to 866, extending its lower bathymetric limit to bathyal depths and with gonothecae in February.
Circum-Antarctic distribution (Peña Cantero and Vervoort 2003). Present material from the continental shelf and slope of the Ross Sea.
Staurotheca dichotoma Allman, 1888
(Fig. 3b, c)
Material examined: Stn 22, a mass of stems, 70 mm high and 70 mm wide, with female gonothecae (NIWA 35630); Stn 94, two masses of stems, 50 and 60 mm in diameter, with female gonothecae (NIWA 36815); Stn 180, a colony, 30 mm high, on stone (NIWA 37901); Stn 256, three masses of stems, up to 60 mm high and 60 mm wide, stolons attached to pebbles, gravel, stones and fragments of shells and corals (NIWA 38677 and 38678); Stn 258, two masses of stems, up to 80 mm high and 100 mm wide, with female gonothecae, stolons attached to gravel (NIWA 38737).
Measurements (in μm): Hydrothecae: free part of adcauline wall 20–200, adnate part of adcauline wall 620–770, adcauline wall 740–870, diameter at aperture 280–310.
Remarks: For a description of this species, I refer to Peña Cantero et al. (1997a) and Peña Cantero and Vervoort (2003).
Ecology and distribution: Hitherto found at depths from 82 (Totton 1930) to 799 m (Peña Cantero et al. 1997a); present material from 447 to 1235 m, distinctly increasing its lower bathymetric limit, epilithic on stones, pebbles and gravel, on fragments of shells, and epibiotic on corals and with gonothecae in February and March.
Antarctic–Kerguelen distribution (Peña Cantero and Vervoort 2003). I report it on the continental shelf of the Ross Sea and on seamounts off Scott Island.
Staurotheca frigida Peña Cantero, Svoboda and Vervoort, 1997a
(Fig. 3d, e)
Staurotheca frigida—Peña Cantero and Vervoort, 2003: 2683–2686, Fig. 8 (synonymy); Peña Cantero, 2008: 459; Soto Àngel and Peña Cantero, 2015: 998, Fig. 8f–g.
Material examined: Stn 17, two masses of stems, 50 and 35 mm long, stolons attached to bryozoans (NIWA 120294).
Measurements (in μm): Hydrothecae: free part of adcauline wall 80–90, adnate part of adcauline wall 330–390, adcauline wall 420–470, abcauline wall 370, diameter at aperture 140–180.
Remarks: For a description of this species, I refer to the original one by Peña Cantero et al. (1997a) or the description by Peña Cantero and Vervoort (2003).
Ecology and distribution: Shelf species (Peña Cantero et al. 1997a), found at depths from 57 (Peña Cantero and Vervoort 2003) to 647 m (Soto Àngel and Peña Cantero 2015); here reported at a depth of 321 m, with stolons attached to bryozoans.
Antarctic-Kerguelen distribution (Peña Cantero and Vervoort 2003). Present material from the shelf of the Ross Sea.
Staurotheca nonscripta Peña Cantero, Svoboda and Vervoort, 1997a
(Fig. 3f)
Material examined: Stn 84, a mass of stems, 40 mm in diameter (NIWA 36727).
Measurements (in μm): Hydrothecae: free part of adcauline wall 340, adnate part of adcauline wall 520, adcauline wall 860, abcauline wall 560, diameter at aperture 210.
Remarks: For an extensive description of this species, see the original description by Peña Cantero et al. (1997a) or that by Peña Cantero and Vervoort (2003).
Ecology and distribution: Shelf species (Peña Cantero and Vervoort 2003), found at depths from 15 (Peña Cantero and Vervoort 2003) to 728 m (Peña Cantero 2014a); present material at 360 m.
Antarctic–Patagonian distribution (cf. Peña Cantero and Vervoort 2003). Present material collected on the shelf of the Ross Sea.
Staurotheca pachyclada (Jäderholm, 1904)

Staurotheca pachyclada (Jäderholm, 1904): a female gonotheca; b male gonotheca. Scale bar: 250 μm
Material examined: Stn 144, several stems and fragments, up to 160 mm high, with male and female gonothecae (NIWA 37471); Stn 161, one stem, 120 mm high (NIWA 37679); Stn 167, three stems, 145, 220 and 205 mm high, with male gonothecae (NIWA 37766).
Measurements (in μm): Female gonotheca (Fig. 4a): height 2000 (with neck), maximum diameter 1500, length of neck 360, diameter at aperture 450. Male gonotheca (Fig. 4b): height 2000, maximum diameter 900, diameter at aperture 200.
Remarks: For a description of this species, including a re-description of the holotype, I refer to Peña Cantero and Vervoort (2003).
This species typically has unbranched stems. In the present material, there are some colonies that have a few secondary stems, but these were formed clearly after the original stem broke. Stems usually give rise to straight unbranched pinnae (few of them have one or two secondary pinnae), arranged alternately in one plane. Some stems have disc-shaped hydrorhizae, clearly indicating attachment on hard substrate.
Ecology and distribution: Eurybathic species (Peña Cantero and Vervoort 2003), found at depths from 42 (Stepanjants 1979) to 1405 m (Peña Cantero and Vervoort 2003); present material at depths between 535 and 1990 m, considerably increasing its lower bathymetric limit; gonothecae in February.
Circum-Antarctic distribution (cf. Peña Cantero and Vervoort 2003). Here reported from the slope of the Ross Sea.
Symplectoscyphidae Maronna, Miranda, Peña Cantero, Barbeitos and Marques, 2016
Antarctoscyphus asymmetricus Peña Cantero, García Carrascosa and Vervoort, 1997
(Fig. 5a)

Antarctoscyphus asymmetricus Peña Cantero, García Carrascosa and Vervoort, 1997: a internode with hydrotheca. Antarctoscyphus elongatus (Jäderholm, 1904): b internode with hydrotheca. Antarctoscyphus fragilis Peña Cantero, Svoboda and Vervoort, 1999: c-d hydrothecae. Antarctoscyphus grandis (Blanco, 1977): e hydrotheca. Antarctoscyphus spiralis (Hickson and Gravely, 1907): f hydrotheca showing internal cusps (arrow). Scale bar: 200 μm
Material examined: Stn 17, two stems, up to 105 mm high (NIWA 35452).
Remarks: For an extensive description of this species, I refer to the original description by Peña Cantero et al. (1997); for additional information, including a SEM study, see Peña Cantero et al. (2017).
Ecology and distribution: Shelf species, found at depths between 18 (Peña Cantero et al. 2017) and 429 m (Peña Cantero 2008); present material collected at 321 m.
Circum-Antarctic distribution (Peña Cantero et al. 2017). Present material comes from the shelf of the Ross Sea.
Antarctoscyphus elongatus (Jäderholm, 1904)
(Fig. 5b)
Material examined: Stn 245, one stem, 70 mm high (NIWA 38580); Stn 249, one stem, 45 mm high (NIWA 38625).
Remarks: For a description of this species, I refer to Peña Cantero et al. (1999); for additional information, including a SEM study, see Peña Cantero et al. (2017).
Ecology and distribution: Eurybathic species (Peña Cantero et al. 2017), collected at depths from 10 (Naumov and Stepanjants 1972) to 1958 m (Peña Cantero 2012); present material from 300 to 760 m.
Antarctic-Kerguelen distribution (Peña Cantero 2012). Here reported from a seamount off Scott Island.
Antarctoscyphus fragilis Peña Cantero, Svoboda and Vervoort, 1999
(Fig. 5c, d)
Material examined: Stn 251, two stems, up to 105 mm high, on stone (NIWA 38659).
Description: Stems basally polysiphonic, up to 105 mm high. Stem divided into long internodes by slightly marked perisarc constrictions. Internodes in marked zigzag; each with distal apophysis giving rise to paired branches and a hydrotheca at axil in between. Paired branches divided into long hydrothecate internodes and sparsely branched. Usually each paired branch giving rise to a single secondary branch at its third internode; only on three occasions a secondary branch giving rise to a third-order branch (twice at fourth internode and one at 7th).
Hydrothecae arranged alternately in two planes with a tendency to unilateral disposition, widely separated, non-overlapping. Hydrotheca completely free, directed outwards and almost cylindrical (some widen distally). Adcauline wall mostly straight, but convex basally; abcauline wall mostly straight, but concave basally. Hydrothecal aperture with three cusps, one adcauline and two abcauline.
Measurements (in μm): Distance between cauline apophyses: 3000–6000. Hydrothecae: height 850–930, diameter at aperture 220–280.
Remarks: Present material agrees with previous descriptions (Peña Cantero et al. 1999, 2017) of the species in almost every detail. The only difference relates to the branching of paired branches, which occurs at the third internode here, whereas it takes place at the second in previous reports (Peña Cantero et al. 1999, 2017).
Ecology and distribution: Deep water species, found at depths between 460 (Peña Cantero et al. 1999) and 1920 m (Peña Cantero et al. 2017); present material from 1450 to 1541 m, on stone.
Originally described on the east coast of the Weddell Sea (Peña Cantero et al. 1999), it has recently been reported off the Pacific entrance of the Magellan Strait and, therefore, considered to have a West Antarctic-Patagonian distribution (Peña Cantero et al. 2017). However, the present record, which constitutes the third for the species, comes from a seamount off Scott Island, which allows its distribution to be defined as Antarctic-Patagonian.
Antarctoscyphus grandis (Blanco, 1977a)
(Fig. 5e)
Material examined: Stn 17, two stems, 300 and 120 mm high, on pebbles (NIWA 35556); Stn 117, three stems, up to 230 mm high, with gonothecae (NIWA 37193); Stn 161, one stem, 270 mm high, on animal debris (NIWA 120295).
Remarks: For an extensive description of this species, including the re-description of the holotype, I refer to Peña Cantero et al. (1999); for additional information, including a SEM study, see Peña Cantero et al. (2017).
Ecology and distribution: Eurybathic species (Peña Cantero et al. 2017), found at depths between 15 (Naumov and Stepanjants 1972) and 1042 m (Peña Cantero et al. 2017); present material from 321 to 536 m, on pebbles and animal debris, and with gonothecae in February.
Circum-Antarctic distribution (Peña Cantero et al. 1997). I report it here from the shelf and slope of the Ross Sea.
Antarctoscyphus spiralis (Hickson and Gravely, 1907)
(Fig. 5f)
Material examined: Stn 17, one stem, 60 mm high (NIWA 35462); Stn 46, a basally broken stem, 50 mm high (NIWA 36037); Stn 147, one distal stem fragment, 15 mm long (NIWA 61729).
Remarks: For a description of this species, I refer to Totton (1930); for additional information, including a SEM study, see Peña Cantero et al. (2017).
Ecology and distribution: Eurybathic species (Peña Cantero et al. 2017), found at depths from 6 m (Naumov and Stepanjants 1972) to 2143 m (Peña Cantero et al. 2017); present material from 321 to 1610 m.
Circum-Antarctic distribution (Stepanjants 1979). Here reported from the shelf and slope of the Ross Sea.
Symplectoscyphus anae Peña Cantero, Sovoboda and Vervoort, 2002
(Fig. 6a)

Symplectoscyphus anae Peña Cantero, Svoboda and Vervoort, 2002: a fragment of stem with hydrothecae. Symplectoscyphus curvatus (Jäderholm, 1917): b hydrotheca. Symplectoscyphus exochus Blanco, 1982: c fragment of stem with hydrothecae; d incomplete gonotheca. Symplectoscyphus frondosus Peña Cantero, 2010: e two branch fragments with hydrothecae; f male gonotheca. Symplectoscyphus liouvillei (Billard, 1914): g internode with hydrotheca. Symplectoscyphus nesioticus Blanco, 1977: h fragment of stem with hydrotheca. Scale bar: 200 μm
Material examined: Stn 17, a few stems, up to 6 mm high, on O. stepanjantsae (NIWA 120296); Stn 144, several stems, up to 15 mm high, with gonothecae, on O. stepanjantsae (NIWA 120297).
Measurements (in μm): Hydrothecae: length of abcauline wall 350–460, length of free adcauline wall 320–390, length of adnate adcauline wall 150–240, length of adcauline wall 480–580, diameter at aperture 140–180, diameter at diaphragm 120, maximum diameter 200. Gonothecae: height 1000, maximum diameter 650, length of neck 70, diameter at aperture 160. Cnidome: larger microbasic mastigophores, range 13.0–14.0 × 3.5–4.0.
Remarks: For an extensive description of this species, I refer to the original description by Peña Cantero et al. (2002).
Ecology and distribution: Shelf species (Peña Cantero 2010), collected at depths from 20 (Naumov and Stepanjants 1972) to 640 m (Peña Cantero et al. 2002); present material from 321 to 1658 m, considerably increasing its lower bathymetric limit to the slope, epibiotic on O. stepanjantsae, and with gonothecae in February.
Circum-Antarctic distribution (Peña Cantero et al. 2002). Here reported for the shelf and slope of the Ross Sea.
Symplectoscyphus curvatus (Jäderholm, 1917)
(Fig. 6b)
Material examined: Stn 144, three stems, up to 25 mm high, on O. stepanjantsae and S. plicatile (NIWA 120298); Stn 157, several stems, up to 40 mm high, with one gonotheca, on axis of dead gorgonian (NIWA 37621).
Measurements (in μm): Hydrothecae: length of abcauline wall 850–880, length of free adcauline wall 660–680, length of adnate adcauline wall 360–450, length of adcauline wall 1020–1130, diameter at aperture 350–380, maximum diameter 350.
Remarks: For a description of this species, I refer to Peña Cantero et al. (2002).
Ecology and distribution: Eurybathic species (Peña Cantero 2010), collected at depths from 49 (Peña Cantero 2008) to 2043 m (Peña Cantero 2012); present material between 814 and 1658 m, on axis of dead gorgonian, O. stepanjantsae and S. plicatile, and with gonothecae in February.
Circum-Antarctic distribution (Stepanjants 1979). Here reported for the slope of the Ross Sea.
Symplectoscyphus exochus Blanco, 1982
(Fig. 6c, d)
Material examined: Stn 17, three masses of stems, up to 30 mm in diameter, on O. stepanjantsae (NIWA 35454); Stn 31, a mass of stems, 30 mm in diameter, with immature gonothecae (NIWA 35882); Stn 144, two masses of stems, up to 30 mm in diameter, with gonothecae, on O. stepanjantsae, basibiont of F. antarcticum (NIWA 37463); Stn 157, a mass of stems, 15 mm in diameter (NIWA 37632).
Measurements (in μm): Hydrothecae: length of abcauline wall 270–310, length of free adcauline wall 170–180, length of adnate adcauline wall 220–280, length of adcauline wall 400–450, diameter at aperture 130–150, maximum diameter 180.
Remarks: For a description of this species, I refer to Peña Cantero et al. (2002).
Ecology and distribution: Eurybathic species (Peña Cantero 2017), collected at depths from 15 (Vervoort 1972a) to 1958 m (Peña Cantero 2012); present material from 283 to 1658 m, on O. stepanjantsae, basibiont of F. antarcticum and with gonothecae in February.
Circum-Antarctic distribution (Peña Cantero 2014a). Present material from the shelf and slope of the Ross Sea.
Symplectoscyphus frondosus Peña Cantero, 2010
(Fig. 6e, f)
Material examined: Stn 17, several stems, up to 130 mm high, with male and immature female gonothecae, on stone (NIWA 35537); Stn 117, one stem, 160 mm high, on bryozoans and sponges (NIWA 120299); Stn 121, one stem, 100 mm high (NIWA 37247); Stn 133, a basally broken stem, 50 mm high (NIWA 37385); Stn 144, seven stems, up to 120 mm high (NIWA 37474); Stn 151, one stem, 75 mm high (NIWA 37560); Stn 157, three stems, up to 85 mm high (NIWA 37620); Stn 161, three stems, up to 70 mm high (NIWA 37680); Stn 167, two stems, 130 and 115 mm high, on coral (NIWA 37761); Stn 171, one stem, 68 mm high (NIWA 37831).
Measurements (in μm): Hydrothecae: length of abcauline wall 390–400, length of free adcauline wall 250–320, length of adnate adcauline wall 190–280, length of adcauline wall 440–600, diameter at aperture 130–170, maximum diameter 200. Male gonothecae: height 1800. Female gonothecae (not completely developed): height up to 1660, maximum diameter 500. Cnidome: larger microbasic mastigophores, range 9.5–11.0 × 3.0–3.5.
Remarks: For an extensive description of this species, I refer to the original description by Peña Cantero (2010).
Ecology and distribution: Hitherto considered a shelf species, it had been found at depths between 400 and 564 m (Peña Cantero 2017); present material, however, collected from 321 to 2283, considerably extending its lower bathymetric limit to the slope. Epilithic on stones and epibiotic on coral, bryozoans and sponges and with gonothecae in February.
Endemic to the eastern Ross Sea (Peña Cantero 2017). Present material supports this pattern, as it comes from the shelf and slope of the Ross Sea.
Symplectoscyphus liouvillei (Billard, 1914)
(Fig. 6g)
Material examined: Stn 17, one stem, 180 mm high, with immature gonothecae, stolons attached to debris (NIWA 35458); Stn 161, one basally broken stem, at least 170 mm high, with immature female gonothecae (NIWA 120300).
Measurements (in μm): Hydrothecae: length of abcauline wall 425, length of free adcauline wall 240–300, length of adnate adcauline wall 290–300, length of adcauline wall 530–600, diameter at aperture 180, maximum diameter 200, diameter at diaphragm 140. Female gonothecae (not completely developed): height at least 1750, maximum diameter 600.
Remarks: For an extensive description of this species, I refer to Peña Cantero et al. (2002).
Ecology and distribution: Shelf species (Peña Cantero 2010), found at depths from 65 (Peña Cantero 2008) to 480 m (Peña Cantero 2017); present material between 321 and 536 m, with stolons attached to debris, and with gonothecae in February.
Antarctic-Patagonian distribution (Peña Cantero 2017). Here reported from the shelf and slope of the Ross Sea.
Symplectoscyphus naumovi Blanco, 1969
Material examined: Stn 61, one stem fragment in bad condition, with gonothecae (NIWA 36256).
Measurements (in μm): Hydrothecae: length of abcauline wall 450, diameter at aperture 200. Gonothecae: height (neck included) 1600, maximum diameter 620, length of neck 120, diameter at aperture 240.
Remarks: For a description of this species, see Peña Cantero et al. (2002).
Ecology and distribution: Eurybathic species (Peña Cantero 2010), found from the tidal level (Blanco 1984) to a depth of 1379 m (Peña Cantero and Ramil 2006); present material at 520–522 m, with gonothecae in February.
Circum-Antarctic species (Peña Cantero and Ramil 2006). Here reported from the shelf of western Ross Sea.
Symplectoscyphus nesioticus Blanco, 1977b
(Fig. 6h)
Material examined: Stn 249, several stems, with gonothecae, on Schizotricha trinematotheca Peña Cantero and Vervoort, 2005 (NIWA 120301).
Measurements (in μm): Hydrothecae: length of abcauline wall 375, length of free adcauline wall 350, length of adnate adcauline wall 120, length of adcauline wall 470, diameter at aperture 200.
Remarks: For an extensive description of this species, I refer to Peña Cantero et al. (2002).
Ecology and distribution: Shelf species (Peña Cantero 2017), found at depths between 56 (Peña Cantero 2006) and 701 m (Peña Cantero 2014a); present material from 300 to 340 m, on S. trinematotheca; gonothecae in March.
Circum-Antarctic distribution (Peña Cantero 2014a). Present material from a seamount off Scott Island.
Family Haleciidae Hincks, 1868
Halecium banzare Watson, 2008
Material examined: Stn 17, one stem, 100 mm high, with stolons attached to shell fragments and gravel, and two basally broken stems, up to 120 mm high, with gonothecae (NIWA 120302); Stn 121, one stem, 100 mm high, on pebbles, stolons also on debris (NIWA 37258).
Measurements (in μm): Hydrothecae: height 50, diameter at aperture 150. Gonothecae: height 850–930, maximum diameter 600–860. Cnidome: microbasic euryteles 7 × 3–3.5, microbasic mastigophores 6.5–7 × 2.
Remarks: For a description of this species, I refer to the original description by Watson (2008) and that by Peña Cantero (2014b), also including the re-description of type material.
Ecology and distribution: Hitherto considered a shelf species (Peña Cantero 2017), found at depths between 197 (Peña Cantero 2017) and 603 m (Watson 2008); present material from 321 to 950 m, distinctly extending its lower bathymetric limit to the upper slope, on pebbles and with hydrorhizal stolons attached to shell fragments and gravel, and with gonothecae in February.
Circum-Antarctic distribution (Peña Cantero 2014b). Present material collected on the shelf and slope of the Ross Sea.
Halecium interpolatum Ritchie, 1907
Material examined: Stn 17, several stems, up to 25 mm high, with gonothecae, on O. stepanjantsae (NIWA 120303).
Remarks: For a description of this species, also including the re-description of type material, see Peña Cantero (2014b).
Ecology and distribution: Shelf species (Peña Cantero 2017), found at depths from three (Stepanjants 1979) to 536 m (Peña Cantero 2017); present material at 321 m, epibiotic on O. stepanjantsae, and with gonothecae in February.
Circum-Antarctic distribution (Peña Cantero 2014b). Present material collected on the shelf of the Ross Sea.
Halecium jaederholmi Vervoort, 1972b
Material examined: Stn 224, several polysiphonic stems, up to 40 mm high, on gorgonian (NIWA 38397); Stn 243, a few stems, up to 25 mm high, on axis of gorgonian (NIWA 38552).
Measurements (in μm): Hydrothecae: height 20–25, diameter at aperture 160–170. Cnidome: microbasic euryteles, 10 × 5.
Remarks: For an extensive description of this species, based on the holotype, see Peña Cantero (2014b).
The stems are polysiphonic and there are intermediate internodes following the apophyses that give rise to branches; usually one or two, but up to three have been observed. The first hydrothecate internode of the branch is similar to the rest. Hydrothecae are more or less in one plane. The adcauline hydrothecal wall is adnate to the internode and longer than the abcauline one.
Ecology and distribution: Hitherto considered a shelf species (Peña Cantero 2017), collected at depths between 24 (Vervoort 1972b) and 565 m (Peña Cantero 2017); present material found between 420 and 950 m, extending its bathymetric distribution to the upper slope, epibiotic on gorgonian.
Pan-Antarctic distribution (cf. Peña Cantero 2014b). Present material collected on seamounts off Scott Island.
Halecium pseudoincertus Peña Cantero, 2014b
(Fig. 7)

Halecium pseudoincertus Peña Cantero, 2014: a female gonotheca (frontal view); bidem (lateral view). Scale bar: 250 μm
Material examined: Stn 161, one stem, 220 mm high, with gonothecae (NIWA 120304).
Measurements (in μm): Female gonotheca: height 1220–1430, maximum diameter 440–500 (frontal view), 300 (lateral view). Nematocysts: isorhizas, 10–11 × 3–3.5.
Remarks: For a description of this species, see the original description by Peña Cantero (2014b). Gonothecae of H. pseudoincertus were unknown. Mature female gonothecae found in the material studied. They are fusiform in lateral view and triangular in frontal view (Fig. 7), similar to those of Halecium incertus Naumov and Stepanjants, 1962, although distinctly thinner (cf. Peña Cantero 2014a, b, p. 260). As in H. incertus, the gonotheca has an acrocyst, apparently with only one embryo (the poor state of the material prevents confirmation).
Ecology and distribution: Shelf and slope species (Peña Cantero 2017), found at depths between 96 (Peña Cantero 2009) and 1019 m (Peña Cantero 2008); present material at 535–536 m, and with gonothecae in February.
Circum-Antarctic distribution (Peña Cantero 2014b). Present material from the shelf of the Ross Sea.
Family Zygophylacidae Quelch, 1885
Abietinella operculata (Jäderholm, 1903)
Material examined: Stn 144, two stems, up to 60 mm high (NIWA 120305).
Remarks: For an extensive description of this species, including the re-description of the holotype, see Marques et al. (2005).
Ecology and distribution: Shelf and slope species (Peña Cantero 2017), known from depths between 63 (Peña Cantero and García Carrascosa 1993) and 1500 m (Stepanjants 1979); present material collected from 1431 to 1658 m, extending its lower bathymetric limit.
Antarctic–Patagonian distribution (Stepanjants 1979). Present material from the slope of the Ross Sea.
Sertularellidae Maronna, Miranda, Peña Cantero, Barbeitos and Marques, 2016
Sertularella pseudovervoorti sp. nov.

Sertularella pseudovervoorti sp. nov.: a fragment of stem showing zigzag arrangement of internodes, hydrothecal disposition and branches; b hydrotheca and branch origin; c hydrotheca. Scale bar: 500 μm (a), 250 μm (b, c)

Sertularella pseudovervoorti sp. nov.: a Holotype; b hydrotheca and branch; c hydrotheca. Scale bar: 3 mm (a), 200 μm (b, c)
Material examined: Stn 237, a few stems, up to 15 mm high, on axis of dead gorgonian [Holotype, a 13-mm-high stem with two lateral branches (NIWA 38496); remaining material paratype (NIWA 120192)].
Description: Monosiphonic stems up to 15 mm in height. Stems unbranched, or with a single short branch originating below first hydrotheca; only one stem with two branches, on the same side of stem, from successive hydrothecae (Figs. 8a and 9a). Stem divided into internodes (first internode much longer) arranged in a marked zigzag (Figs. 8a and 9a). Each with a hydrotheca at end of internode (Figs. 8 and 9). Branches originating below hydrotheca (Fig. 8a, b). Hydrotheca cylindrical, almost completely free of internode (Figs. 8 and 9). Adcauline wall slightly convex. Abcauline wall straight or slightly convex basally and somewhat concave distally. Adcauline wall slightly wavy, with three or four undulations. Rim of hydrothecal aperture with four small cusps separated by shallow embayments.
Measurements (in μm): Internodes: length 1300–2100 (first internode 3600–6800), diameter at hydrothecal base 280–360. Hydrothecae: length of abcauline wall 660–750, free part of adcauline wall 630–680, adnate part of adcauline wall 200, adcauline wall 830–880, diameter at aperture 320–350, maximum diameter 370–390.
Remarks: Even when the hydrothecae have the appearance of being entirely free, there is a portion of the adcauline wall that is actually adnate, although it is almost in the same plane with the true base.
The hydrotheca of S. pseudovervoorti sp. nov. resembles that of S. vervoorti El Beshbeeshy, 2011, but is clearly distinguishable because it is approximately half its size (e.g. abcauline length 1345–1624 μm and diameter at aperture 510–614 μm in S. vervoorti, according to El Beshbeeshy 2011). In addition, the hydrotheca is roughly cylindrical in S. pseudovervoorti sp. nov., whereas in S. vervoorti the diameter of the hydrotheca distinctly diminishes basally, reaching its minimum when it becomes free. They also differ by the wavy adcauline wall of S. pseudovervoorti sp. nov. The structure of the colony is also different because S. pseudovervoorti sp. nov. has erect stems divided into internodes arranged in marked zigzag, whereas in S. vervoorti the stem consists of a series of curved internodes in linear arrangement. Finally, while in S. vervoorti hydrothecae are alternately arranged in two planes that form a very acute angle, with a clear tendency to unilateral arrangement, they are arranged alternately in one plane in S. pseudovervoorti sp. nov.
Ecology and distribution: Sertularella pseudovervoorti sp. nov. was collected at a depth of 1520–1560 m off Scott Island, on the axis of a dead gorgonian.
Etymology: The specific name pseudovervoorti refers to the fact that the shape of the hydrotheca in this species is evocative of that of S. vervoorti.
Schizotrichidae Peña Cantero, Sentandreu and Latorre, 2010
Schizotricha nana Peña Cantero, Svoboda and Vervoort, 1996
(Fig. 10a)

Schizotricha nana Peña Cantero, Svoboda and Vervoort, 1996: a a fragment of branch showing internodes with hydrothecae and nematothecae. Schizotricha trinematotheca Peña Cantero and Vervoort, 2005: b-c hydrocladial internodes showing hydrothecae and nematothecae. Schizotricha turqueti Billard, 1906: d a fragment of branch showing internodes with hydrothecae and nematothecae. Scale bar: 200 μm
Material examined: Stn 144, two stems, 88 and 100 mm high, with gonothecae (NIWA 120191).
Remarks: For an extensive description of this species, I refer to the original description by Peña Cantero et al. (1996).
Ecology and distribution: Eurybathic species (Peña Cantero 2017), known from depths between 43 (Stepanjants 1972) and 1890 m (Peña Cantero and Vervoort 2005); present material collected from 1431 to 1658 m, with gonothecae in February.
Circum-Antarctic distribution (Peña Cantero et al. 1996). Reported here for the slope of the Ross Sea.
Schizotricha trinematotheca Peña Cantero and Vervoort, 2005
(Fig. 10b, c)
Schizotricha trinematotheca Peña Cantero and Vervoort, 2005: 809–812, Fig. 2; Peña Cantero, 2009: 1747, Fig. 2 g–h.
Schizotricha anderssoni: Naumov and Stepanjants, 1962: 101, Fig. 20.
Schizotricha unifurcata unifurcata: Stepanjants, 1979: 115 (in part).
Material examined: Stn 243, nine sparsely branched stems (three are unbranched), up to 200 mm high, with gonothecae (NIWA 38553); Stn 245, two much-branched stems, up to 240 mm high, with male and female gonothecae (NIWA 38565); Stn 249, many stems, at least 320 mm high, basibiont of S. nesioticus (NIWA 38621).
Remarks: For an extensive description of this species, I refer to the original description by Peña Cantero and Vervoort (2005); for additional information, see also Peña Cantero (2009).
Schizotricha trinematotheca is similar to Schizotricha anderssoni Jäderholm, 1904 in the high number of nematothecae in the hydrocladial internodes, which can be misleading. However, a careful study of both species allows differentiation.
Schizotricha trinematotheca differs from S. anderssoni by the constant presence of an intermediate ahydrothecate internode after cauline apophyses (its absence being an oddity). Typically, there is also an intermediate internode following hydrocladial apophyses. Hydrocladia are usually unbranched, although the bifurcated condition is also quite frequent and, in this case, the hydrocladial apophyses, which are quite short and provided with one nematotheca, are typically followed by an intermediate ahydrothecate internode provided with up to four nematothecae. In S. anderssoni, however, there is no intermediate ahydrothecate internode after the cauline apophyses (its presence is a rarity). In addition, in S. anderssoni, the hydrocladia are typically bifurcated and the hydrocladial apophyses are not followed by an intermediate internode.
They also differ in the branching pattern. While in S. trinematotheca, stems are usually unbranched or slightly branched (yet there are also quite branched stems); stems are much branched in S. anderssoni. As indicated in Peña Cantero and Vervoort (2005p. 801), ‘Schizotricha anderssoni has much branched stems; indeed, there is continuous branching of the stem. Moreover, this species usually lacks branched hydrocladia. When branched, hydrocladia are either merely bifurcated or they become lower-order stems. Schizotricha anderssoni usually lacks intermediate ahydrothecate internodes in the hydrocladia, except in those becoming lower-order stems’. Even when in S. trinematotheca, it is possible to find quite branched stems, the branching is different: stems of lower order do not originate from a further development of the hydrocladia, as happens in S. anderssoni, but as new stems that seemingly originate from accessory tubes of the polysiphonic part of the oldest stem. This is demonstrated by the elimination of all the accessory tubes of the lower-order stem, which exposes its basal part and shows that it is composed of a series of ahydrothecate internodes preceding the first hydrothecate internode that is also provided with an apophysis. In addition, the diameter of the main tube of the lower-order stems is smaller at the origin, gradually increasing distally to reach the normal diameter of the internodes when the first hydrothecate internode is formed.
Finally, in S. trinematotheca, the heteromerous division of the hydrocladia into internodes is frequent, with the common presence of double internodes (there are even triplets). In addition, hydrocladia are distinctly longer in S. trinematotheca (with up to 29 hydrothecae).
As indicated above, both species share a high number of nematothecae. However, this number is often higher in S. trinematotheca. Schizotricha anderssoni usually has three infrathecal nematothecae at different levels (four are also found), whereas S. trinematotheca usually has four in more or less two pairs (six are also frequent).
The material assigned to S. anderssoni by Naumov and Stepanjants (1962), and later considered to belong to S. unifurcata unifurcata by Stepanjants (1979), probably belongs to S. trinematotheca. It is characterised by the high number of nematothecae, as happens in both S. anderssoni and S. trinematotheca, but ‘in the proximal part of the hydrocladia there are internodes which are not provided with hydrothecae but bear a large number of nematothecae’ (Naumov and Stepanjants 1962, p. 101). As noted above, the presence of ahydrothecate internodes is characteristic of S. trinematotheca. There are also biogeographic reasons to support this idea. Schizotricha anderssoni has always been reported from the South Georgia area (Jäderholm 1904; Peña Cantero and Vervoort 2005), the only record outside that area is that by Naumov and Stepanjants (1962) from the remote area of Scott Island (the material studied here also comes from this area). If the material studied by Naumov and Stepanjants (1962) actually belonged to S. trinematotheca, it would have important biogeographic implications, since S. anderssoni would therefore be an endemism of the South Georgia area.
Ecology and distribution: Shelf species, previously found at depths between 70 and 579 m (Peña Cantero 2009); present material from 300 to 760 m, extending its lower bathymetric limit, basibiont of S. nesioticus, and with gonothecae in March.
Previously considered endemic to the Balleny Islands (Peña Cantero 2009), present material comes from a neighbouring seamount off Scott Island.
Schizotricha turqueti Billard, 1906
(Fig. 10d)
Material examined: Stn 17, one stem, 125 mm high, with male? gonothecae (NIWA 120306); Stn 144, two unbranched stems, 75 and 135 mm high, with female gonothecae (NIWA 120307).
Remarks: For an extensive description of this species, I refer to Peña Cantero et al. (1996); for additional information see also Peña Cantero and Vervoort (2005) and Peña Cantero (2017).
Ecology and distribution: Eurybathic species (Peña Cantero and Vervoort 2005), collected from the intertidal level (Billard 1906) to a depth of 1890 m (Peña Cantero and Vervoort 2005); present material from 321 to 1658 m, with gonothecae in February.
Circum-Antarctic distribution (Peña Cantero and Vervoort 2005). Reported here for the shelf and slope of the Ross Sea.
Kirchenpaueriidae Stechow, 1921
Oswaldella bifurca (Hartlaub, 1904)
(Fig. 11a–c)

Oswaldella bifurca (Hartlaub, 1904): a cauline apophysis showing axillary nematophores (arrows); b distal hydrocladial internode with pointed end; c hydrocladial internode showing hydrotheca and mesial inferior nematophore. Oswaldella garciacarrascosai Peña Cantero, Svoboda and Vervoort, 1997: d cauline apophysis showing ‘mamelon’ (arrow); e hydrocladial internode showing hydrotheca and mesial superior and mesial inferior nematophore; f detail of hydrotheca and mesial superior and mesial inferior nematophore. Oswaldella laertesi Peña Cantero, 2007: g detail of mesial inferior nematophore (arrow). Oswaldella rigida Peña Cantero, Svoboda and Vervoort, 1997: h hydrocladial internode showing hydrotheca and mesial inferior nematotheca (arrow). Scale bar: 200 μm (b, e, h), 100 μm (a, c, d, f), 50 μm (g)
Oswaldella bifurca—González Molinero and Peña Cantero, 2015: 403, 433, Figs 1, 16A, 17A, 19A (synonymy).
Material examined: Stn 117, several stems, up to 52 mm high, on shell fragments (NIWA 120308); Stn 144, two stems, 40 and 43 mm high (NIWA 37497).
Remarks: For an extensive description of this species, I refer to Peña Cantero and Vervoort (2004); for a recent SEM study, see González Molinero and Peña Cantero (2015).
Ecology and distribution: Deep-water species (González Molinero and Peña Cantero 2015), found at depths from 342 to 1610 m (Peña Cantero and Vervoort 2004); present material from 475 to 1658 m, on shell fragments.
Circum-Antarctic distribution (Peña Cantero and Vervoort 2004). Here reported from the slope of the Ross Sea.
Oswaldella garciacarrascosai Peña Cantero, Svoboda and Vervoort, 1997b
(Fig. 11d–f)
Oswaldella garciacarrascosai—González Molinero and Peña Cantero, 2015: 409, 435, Figs 6, 16F, 17F, 19F (synonymy).
Material examined: Stn 180, several stems, up to 70 mm high, with gonothecae (NIWA 37896); Stn 182, a few stems, up to 60 mm high, on stone (NIWA 37969).
Remarks: For a lengthy description of this species, see the original description by Peña Cantero et al. (1997b); additionally, for a SEM study, see González Molinero and Peña Cantero (2015).
Our material is perfectly in agreement with previous description of the species. Stems are monosiphonic, unbranched and divided into internodes. Hydrocladia are unbranched. There are two axillary nematophores and one mamelon on cauline apophyses. The hydrotheca is elongate and there is no mesial inferior nematotheca. See González Molinero and Peña Cantero (2015) for a SEM study on part of this material.
Ecology and distribution: Shelf species (González Molinero and Peña Cantero 2015), previously found at a depth of 330–340 m (Peña Cantero et al. 1997b); present material from 410 to 455 m, with gonothecae in February.
Circum-Antarctic distribution (González Molinero and Peña Cantero 2015). Present material collected on a seamount south off Scott Island, representing the second finding of the species.
Oswaldella laertesi Peña Cantero, 2007
(Fig. 11g)
Material examined: Stn 17, two stems, 280 and 100 mm high, plus an stem fragment, 130 mm long, with male gonothecae (NIWA 35536); Stn 117, one stem broken into two parts, 345 mm in total length, with female gonothecae (NIWA 37198); Stn 121, two stems, 140 and 110 mm high (NIWA 37257).
Remarks: For a description of this species, I refer to the original description by Peña Cantero (2007); see González Molinero and Peña Cantero (2015) for a SEM study on part of this material.
Ecology and distribution: Shelf and slope species (Peña Cantero 2017), found at depths between 343 and 736 m (Peña Cantero 2007); present material from 321 to 950 m, increasing its lower bathymetric limit, and with gonothecae in February.
East Antarctic distribution (Peña Cantero 2007), only known from the Ross Sea region. Present material comes from the shelf and slope of the Ross Sea.
Oswaldella rigida Peña Cantero, Svoboda and Vervoort, 1997b
(Fig. 11h)
Material examined: Stn 31, one stem fragment, 20 mm long (NIWA 35841).
Remarks: For a description of this species, see the original description by Peña Cantero et al. (1997b); see also González Molinero and Peña Cantero (2015) for a SEM study of the species.
As characteristic of this species, cauline apophyses are almost perpendicular to the stem and are provided with two axillary nematophores, the mesial inferior nematotheca of the hydrocladial internodes is strongly marked (Fig. 11h) and the hydrocladial internodes have a typical embayment in their basal half (Fig. 11h).
Ecology and distribution: Shelf and slope species (González Molinero and Peña Cantero 2015), known from depths between 80 (Peña Cantero et al. 1997b) and 1157 m (Peña Cantero 2014a); present material collected at 283 m.
Circum-Antarctic distribution (Peña Cantero et al. 1997b). Here reported for the shelf of the Ross Sea.
Oswaldella stepanjantsae Peña Cantero, Svoboda and Vervoort, 1997b
(Fig. 12)

Oswaldella stepanjantsae Peña Cantero, Svoboda and Vervoort, 1997: a cauline apophysis showing axillary nematophores (left arrows) and ‘mamelons’ (right arrows); b detail of axillary nematophore showing collar of perisarc; c frontal view of mesial inferior nematotheca (arrow); d lateral view of mesial inferior nematotheca (arrow). Scale bar: 100 μm (a), 50 μm (b–d)
Material examined: Stn 17, a few stems and fragments, up to 470 mm, basibiont of H. interpolatum, S. anae and S. exochus (NIWA 35451); Stn 117, two stems, 180 and 120 mm high (NIWA 120309); Stn 144, numerous stems, up to 250 mm high, basibiont of F. antarcticum, S. anae, S. curvatus and S. exochus (NIWA 37472); Stn 157, three branched stems, up to 200 mm high (NIWA 37618); Stn 161, three stems, up to 300 mm high (NIWA 37683); Stn 178, one branched stem, 105 mm high (NIWA 37880).
Remarks: For an extensive description of this species, I refer to Peña Cantero and Vervoort (2004); see also González Molinero and Peña Cantero (2015) for a SEM study.
Cauline apophyses are provided with up to six axillary nematophores (Fig. 12a) that emerge from simple perisarc holes and are provided with a very short collar-shaped nematotheca (Fig. 12b), and up to two ‘mamelons’. Hydrocladial internodes are characteristically provided with a tiny, nail-shaped, mesial inferior nematotheca, placed on a sharp step (Fig. 12c, d).
Ecology and distribution: Eurybathic species (Peña Cantero 2017), known from depths between 36 and 1890 m (Peña Cantero and Vervoort 2004); present material collected from 321 to 1658 m, basibiont of F. antarcticum, H. interpolatum, S. anae, S. curvatus and S. exochus.
Circum-Antarctic distribution (Peña Cantero and Vervoort 1998). Present material comes from the shelf and slope of the Ross Sea, and from a seamount south of Scott Island.
General biodiversity
Thirty-five species, one of them new to science (Sertularella pseudovervoorti sp. nov.) were found in the collection of benthic hydroids collected during the IPY-CAML TAN0802 survey on the shelf and slope of the Ross Sea and around the area of Scott Island (Table 2).
Symplectoscyphidae is by far the most diversified family with 12 species (34%), followed by Kirchenpaueriidae and Staurothecidae each with 5 species (14%) and Haleciidae with 4 (11%). Symplectoscyphus with seven species is the most speciose genus, followed by Antarctoscyphus, Oswaldella and Staurotheca with five and Halecium with four. In general, hydroid occurrence is low; 25 species were found in 1 or 2 stations. Symplectoscyphus frondosus is the species with the highest occurrence, being present in ten stations (34%), followed by S. plicatile and O. stepanjantsae, found in six (21%), and S. dichotoma in five (17%).
Discussion
Bouillonia denhartogi is the only Anthoathecata present in a collection that is clearly dominated by Leptothecata (Table 2). The scarcity of anthoathecates is quite surprising since 18 Anthoathecata are known from the Ross Sea (Peña Cantero 2017). This may be related to the relatively poor state of the material studied (anthoathecates are in general badly damaged when obtained with indirect sampling methods), but also to the bathymetric range of the survey, since most samples come from slope bottoms, whereas anthoathecates apparently have a higher occurrence in shallow waters (Peña Cantero 2004). Concerning the sampling methods, it is worth mentioning the relatively high diversity of anthoathecates present in studies based on material obtained by scuba diving. For example, Stepanjants (1972) reported 11 species of anthoathecates, from a total of 32 species of benthic hydroids, at depths between 0 and 50 m in the Davis Sea, and Peña Cantero et al. (2013) found 11 anthoathecates, in addition to 9 leptothecates, from 0 to 40 m in the Ross Sea. These studies clearly demonstrate that when hydroids are collected carefully the diversity of anthoathecates is higher. Scuba diving also allows sampling habitats (e.g. rocky bottoms) usually avoided when indirect sampling methods are used due to the difficulty in sampling them. Studies in which hydroids are carefully collected by scuba diving, but also kept alive in the laboratory, like that by Peña Cantero et al. (2013), are particularly important, because they provide evidence of a much higher species richness, noticing a hidden diversity of inconspicuous and even undescribed species, and allow reconstructing life cycles, which in turn can help to clarify the systematic position of certain species [see for example the case of Phialella belgicae (Hartlaub, 1904) in Peña Cantero et al. (2013)].
Twenty species were found inhabiting the shelf and slope bottoms of the Ross Sea. All are already known from the area (see Peña Cantero 2017), except S. plicatile that was collected on the slope. However, it is worth mentioning that samples from deep waters, particularly those of the slope, have allowed an extension of the known bathymetric range for almost half of the species (Fig. 13). This increase has been particularly noteworthy in F. antarcticum, S. anae and S. frondosus, so far considered shelf species, but which have now been shown to inhabit deep-water bottoms of the slope (S. frondosus reaching to 2300 m).

Bathymetric range of the species studied (depth in metres)
Benthic hydroids from the remote area of Scott Island were virtually unknown; only S. irregularis, S. dichotoma and S. anderssoni being recorded from there (Naumov and Stepanjants 1962). Here, I report ten species, including Sertularella pseudovervoorti sp. nov. Eight of them are new records and, therefore, the number of species known in this poorly explored and isolated Antarctic area is raised to 11.
Despite the limited information about the use of substrate that can be obtained when using indirect sampling methods (cf. Peña Cantero 2008, 2013, 2014a), it is worth mentioning that 19 species were attached to a substrate (Table 2). Although most of them were found on a single substrate (Table 2), this does not imply any specificity (except perhaps for B. denhartogi), as they have been found in other substrates in previous studies. In the collection, species were found growing on both non-living (e.g. stones, gravel, animal debris) or living (e.g. sponges, bryozoans, hydroids) substrates. Staurotheca dichotoma is the species with the highest variety of substrates (Table 2). Little is known about the substrate preferences of most species that form large erect colonies (e.g. S. pachyclada, Schizotricha species or Oswaldella species) because they usually come on board unattached or basally broken, although there are a few exceptions, such as S. frondosus, found on stones, corals, bryozoans and sponges (Table 2). The hydrozoan species studied seem not to be a particularly favoured substrate for other hydroids. Of the 35 species present in the collection, only 4 are basibionts of other hydroids, and 3 of them of a single species (Table 2). Only Oswaldella stepanjantsae harbours a relatively high hydroid diversity (five species). Its suitability as substrate for other hydroids has been demonstrated in previous studies; for example, Peña Cantero (2014a) found 17 different species.
In relation to the geographic distribution, the present study provided some important results. Firstly, the finding of A. fragilis, which represents the third record for the species. Originally described on the east coast of the Weddell Sea (Peña Cantero et al. 1999), it has recently been reported off the Pacific entrance of the Magellan Strait and has been considered to have a West Antarctic-Patagonian distribution (Peña Cantero et al. 2017). However, the present record from the area of Scott Island, on the opposite side of Antarctica, considerably extends its known geographic distribution, which can be defined as Antarctic-Patagonian. Second, the present study supports the idea that S. frondosus and O. laertesi are endemic to the shelf and slope of the eastern Ross Sea. They have never been reported elsewhere on the Antarctic continental shelf and appear to be absent further north, as they were not found in the area of Scott Island, and seem to be also absent from the Balleny Islands (Peña Cantero 2009). Finally, the hydrozoan fauna of Scott Island, although with typical representatives of the Antarctic fauna, has some exclusive species, such as S. pseudovervoorti sp. nov., which is endemic to this area, and S. trinematotheca, which is limited to it and the neighbouring area of the Balleny Islands, making it an interesting area from a biogeographic perspective.
Sixty-eight percent of the species are endemic to Antarctic waters (Table 2). They are endemic to East Antarctica (4 species, 11%) or circum-Antarctic, that is, present in both East and West Antarctica (57%). This level of endemism is similar to that found in other Antarctic areas [e.g. 65% off Queen Mary Coast (Peña Cantero 2014a)], but lower than in others [e.g. 81% off Low Island (Peña Cantero 2013)]. There is also a significant group of species typically found in Antarctic waters that are also present in sub-Antarctic waters. These include species with Antarctic-Patagonian (5 species, 14%), Antarctic-Kerguelen (3 species, 9%) and Pan-Antarctic (1 species, 3%) distribution. Thus, most species are restricted to Antarctic or Antarctic/sub-Antarctic waters (33 species, 94%); only F. antarcticum and S. plicatile are found outside these waters. The latter is a deep-sea species reported worldwide, but the former is commonly found in Antarctic waters, although there is a record from off South Africa.
References
Billard A (1906) Hydroïdes. In: Joubin L (ed) Expédition Antarctique Française (1903–1905) commandée par le Dr Jean Charcot, Masson et Cie, Paris, pp 1–20
Blanco OM (1984) Contribución al conocimiento de hidrozoos antárticos y subantárticos. Contrib Inst Antárt Argent 294:1–53 pls 1–47
El Beshbeeshy M (2011) Thekate Hydroiden von Patagonischen Schelf (Cnidaria, Hydrozoa, Thecata). Edited by Jarms G. Verh Naturwiss Ver Hamburg 46:19–233
González Molinero A, Peña Cantero AL (2015) SEM study of species of Oswaldella Stechow, 1919 (Cnidaria, Hydrozoa, Kirchenpaueriidae), with an annotated checklist of the species of the genus. Zootaxa 4052:401–441
Jäderholm E (1904) Mitteilungen ueber einige von der Schwedischen Antarctic-Expedition 1901–1903 eingesammelte Hydroiden. Arch Zool exp gén 4 Notes et revue 1:1–14
Maronna MM, Miranda TP, Peña Cantero AL, Barbeitos MS, Marques AC (2016) Towards a phylogenetic classification of Leptothecata (Cnidaria, Hydrozoa). Sci Rep 6:18075
Marques AC, Peña Cantero AL, Migotto AE (2005) Redescription and systematic status of the Antarctic genus Abietinella Levinsen, 1913 (Lafoeidae, Hydrozoa, Cnidaria). J Nat Hist 39:1443–1455
Millard NAH (1964) The Hydrozoa of the south and west coasts of South Africa. Part II. The Lafoeidae, Syntheciidae and Sertulariidae. Ann S Afr Mus 48:1–56
Millard NAH (1975) Monograph on the Hydroida of southern Africa. Ann S Afr Mus 68:1–513
Naumov DV, Stepanjants SD (1962) Hydroida (Thecaphora) collected by the Soviet Antarctic Expedition on the M/V Ob in antarctic and subantarctic waters. In: Biol Res Soviet Antarct Exped 1955–1958 1 (in Russian). Issled Fauny Morei 1:68–106
Naumov DV, Stepanjants SD (1972) Marine invertebrates from Adélie land collected by the XIIth and XVth French Antarctic expeditions. 3. Hydroida. Téthys Suppl 4:25–60
Peña Cantero AL (2004) How rich is the deep-sea Antarctic benthic hydroid fauna? Polar Biol 27:767–774
Peña Cantero AL (2006) Benthic hydroids from the south of Livingston Island (south Shetland Islands, Antarctica) collected by the Spanish Antarctic expedition Bentart 94. Deep Sea Res II 53:932–948
Peña Cantero AL (2007) Breaking molds: Oswaldella laertesi, sp. nov., a unique Antarctic species of Oswaldella Stechow, 1919 (Cnidaria: Hydrozoa: Kirchenpaueriidae). Zootaxa 1612:63–68
Peña Cantero AL (2008) Benthic hydroids (Cnidaria: Hydrozoa) from the Spanish Antarctic expedition Bentart 95. Polar Biol 31:451–464
Peña Cantero AL (2009) Benthic hydroids (Cnidaria, Hydrozoa) from the Balleny Islands (Antarctica). Polar Biol 32:1743–1751
Peña Cantero AL (2010) On a new Antarctic species of Symplectoscyphus Marktanner-Turneretscher, 1890 (Cnidaria, Hydrozoa, Sertulariidae), with an annotated checklist of the Antarctic species of the genus. Zootaxa 2494:29–44
Peña Cantero AL (2012) Filling biodiversity gaps: benthic hydroids from the Bellingshausen Sea (Antarctica). Polar Biol 35:851–865
Peña Cantero AL (2013) Benthic hydroids from off Low Island (Southern Ocean, Antarctica). Mar Ecol 34(Suppl 1):123–142
Peña Cantero AL (2014a) Benthic hydroids (Cnidaria, Hydrozoa) from the continental shelf and slope off Queen Mary coast (East Antarctica). Polar Biol 37:1711–1731
Peña Cantero AL (2014b) Revision of the Antarctic species of Halecium Oken, 1815 (Cnidaria, Hydrozoa, Haleciidae). Zootaxa 3790:243–280
Peña Cantero AL (2017) Benthic hydroids (Cnidaria, Hydrozoa) from the Ross Sea (Antarctica) collected by the New Zealand Antarctic expedition BioRoss 2004 with RV Tangaroa. Zootaxa 4293(1):1–65
Peña Cantero AL, García Carrascosa AM (1993) The coppinia of Abietinella operculata (Lafoeidae: Hydrozoa Leptomedusae) and its systematic position. J Nat Hist 27:1003–1011
Peña Cantero AL, García Carrascosa AM (1995) Hidrozoos bentónicos de la campaña Antártida 8611. Publ Espec Inst Esp Oceanogr 19:1–148
Peña Cantero AL, García Carrascosa AM (1999) Biogeographical distribution of the benthic thecate hydroids collected during the Spanish “Antártida 8611” expedition and comparison between Antarctic and Magellan benthic hydroid faunas. Sci Mar 63:209–218
Peña Cantero AL, Ramil F (2006) Benthic hydroids associated with volcanic structures from Bransfield Strait (Antarctica) collected by the Spanish Antarctic expedition GEBRAP96. Deep Sea Res II 53:949–958
Peña Cantero AL, Vervoort W (1998) On two new species of Oswaldella Stechow, 1919 (Cnidaria, Hydrozoa) from Bransfield Strait (Antarctica). Polar Biol 20:33–40
Peña Cantero AL, Vervoort W (2003) Species of Staurotheca Allman, 1888 (Cnidaria: Hydrozoa: Sertulariidae) from US Antarctic expeditions, with the description of three new species. J Nat Hist 37:2653–2722
Peña Cantero AL, Vervoort W (2004) Species of Oswaldella Stechow, 1919 (Cnidaria: Hydrozoa: Kirchenpaueriidae) from US Antarctic expeditions, with the description of three new species. J Nat Hist 38:805–861
Peña Cantero AL, Vervoort W (2005) Species of Schizotricha Allman, 1883 (Cnidaria: Hydrozoa: Halopterididae) from US Antarctic expeditions with the description of two new species. J Nat Hist 39:795–818
Peña Cantero AL, Svoboda A, Vervoort W (1996) Species of Schizotricha Allman, 1883 (Cnidaria, Hydrozoa) from recent antarctic expeditions with R.V. “Polarstern”, with the description of a new species. Zool Meded 70:411–435
Peña Cantero AL, García Carrascosa AM, Vervoort W (1997) On Antarctoscyphus (Cnidaria, Hydrozoa), a new genus of antarctic hydroids and the description of two new species. Polar Biol 18:23–32
Peña Cantero AL, Svoboda A, Vervoort W (1997a) Species of Staurotheca Allman, 1888 (Cnidaria, Hydrozoa) from recent Antarctic expeditions with R.V. ‘Polarstern’, with the description of six new species. J Nat Hist 31:329–381
Peña Cantero AL, Svoboda A, Vervoort W (1997b) Species of Oswaldella Stechow, 1919 (Cnidaria, Hydrozoa) from recent Antarctic expeditions with R.V. “Polarstern”, with the description of eight new species. Zool J Linnean Soc 119:339–388
Peña Cantero AL, García Carrascosa AM, Vervoort W (1998) On the species of Filellum Hincks, 1868 (Cnidaria: Hydrozoa) with the description of a new species. J Nat Hist 32:297–315
Peña Cantero AL, Svoboda A, Vervoort W (1999) Species of Antarctoscyphus Peña Cantero, García Carrascosa and Vervoort, 1997 (Cnidaria, Hydrozoa, Sertulariidae) from recent Antarctic expeditions with R.V. Polarstern, with the description of two new species. J Nat Hist 33:1739–1765
Peña Cantero AL, Svoboda A, Vervoort W (2002) Species of Symplectoscyphus Marktanner-Turneretscher, 1890 (Cnidaria: Hydrozoa, Sertulariidae) from recent Antarctic expeditions with R.V. Polarstern, with the description of four new species. J Nat Hist 36:1509–1568
Peña Cantero AL, Svoboda A, Vervoort W (2004) Antarctic hydroids (Cnidaria: Hydrozoa) of the families Campanulinidae, Lafoeidae and Campanulariidae from recent Antarctic expeditions with R.V. Polarstern, with the description of a new species. J Nat Hist 38:2269–2303
Peña Cantero AL, Boero F, Piraino S (2013) Shallow-water benthic hydroids from Tethys bay (Terra Nova Bay, Ross Sea, Antarctica). Polar Biol 36:731–753
Peña Cantero AL, Roig Ferrer E, Miranda TP (2017) Species of Antarctoscyphus Peña Cantero, García Carrascosa and Vervoort, 1997 (Cnidaria: Hydrozoa: Symplectoscyphidae) collected by US Antarctic expeditions: biogeographic implications. J Nat Hist 51:1437–1477
Schuchert P (2001) Hydroids of Greenland and Iceland (Cnidaria, Hydrozoa). Meddelelser om Grønland, bioscience 53. The Danish Polar Center, Copenhagen
Soto Àngel JJ, Peña Cantero AL (2015) On the benthic hydroids from the Scotia Arc (Southern Ocean): new insights into their biodiversity, ecology and biogeography. Polar Biol 38:983–1007
Stepanjants SD (1972) Hydroidea of the coastal waters of the Davis Sea (collected by the XIth Soviet Antarctic Expedition of 1965–1966 In: Biol res Soviet Antarct Exped 5 (in Russian). Issled Fauny Morei 11:56–79
Stepanjants SD (1979) Hydroids of the antarctic and subantarctic waters. In: Biol res Soviet Antarct Exped 6 (in Russian). Issled Fauny Morei 20:1–200, pls 1–25
Svoboda A, Stepanjants SD, Ljubenkov J (2006) The genus Bouillonia (Cnidaria: Hydrozoa: Anthoathecata). Three species from the northern and southern hemispheres, with a discussion of bipolar distribution of this genus. Zool Meded 80–4:185–206
Totton AK (1930) Coelenterata. Part V Hydroida Nat Hist Rep Br Antarct Terra Nova Exped 1910 5:131–252 pls 1–3
Vanhöffen E (1910) Die Hydroiden der Deutschen Südpolar-Expedition 1901–1903. Dtsch Südpolar Exped 1901–1903(11):269–340
Vervoort W (1972a) Hydroids from submarine cliffs near Arthur Harbour, Palmer Archipelago, Antarctica. Zool Meded 47:337–357
Vervoort W (1972b) Hydroids from the Theta, Vema and Yelcho cruises of the Lamont-Doherty geological observatory. Zool Verh 120:1–247
Watson JE (2008) Hydroids of the BANZARE expeditions, 1929–1931: the family Haleciidae (Hydrozoa, Leptothecata) from the Australian Antarctic territory. Mem Mus Vic 65:165–178
Acknowledgments
I thank Dr. Ashley Rowden, National Institute of Water & Atmospheric Research (NIWA), for giving me the opportunity to study the material. I also thank Sadie Mills and Dr. Kareen Schnabel, from the same institution, for their help with different aspects related to the collection. This research was funded by the New Zealand Government under the New Zealand International Polar Year Census of Antarctic Marine Life Project (Phase 1: So001IPY; Phase 2; IPY2007-01). I gratefully acknowledge project governance provided by the Ministry of Fisheries Science Team and the Ocean Survey 20/20 CAML Advisory Group (Land Information New Zealand, Ministry of Fisheries, Antarctica New Zealand, Ministry of Foreign Affairs and Trade, and National Institute of Water and Atmospheric Research).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The author declares that he has no conflict of interest.
Ethical approval
All applicable international, national, and/or institutional guidelines for the care and use of animals were followed by the author.
Sampling and field studies
All necessary permits for sampling and observational field studies have been obtained by the author from the competent authorities and are mentioned in the acknowledgements, if applicable.
Additional information
Communicated by S. Piraino
This article is registered in ZooBank under urn:lsid:zoobank.org:pub:2642909E-5E32-4A3F-A5E1-C1A528CD1D55
Rights and permissions
About this article

Cite this article
Peña Cantero, Á.L. Benthic hydroids off Scott Island and the shelf and slope of the Ross Sea (Antarctica) collected during the IPY-CAML TAN0802 voyage by R/V Tangaroa. Mar Biodiv 49, 863–885 (2019). https://doi.org/10.1007/s12526-018-0865-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12526-018-0865-x