
Abstract
Hydrocarbons contamination is a worldwide threat to soil ecosystem. The present study was designed to evaluate the remediation potential of bacterial consortium, viz. Bacillus cereus (Acc KR232400), Bacillus altitudinis (Acc KF859970), Commomonas (Delftia sp.) (Acc KF859971), and Stenotrophomonas maltophilia (Acc KF859973), with alfalfa and maize plants in combination with fertilizer to remediate the oily sludge. Oily sludge was mixed with soil in 30:70 ratio, and ammonium nitrate and diammonium phosphate (DAP) were added at a rate of 70 μg/g and 7 μg/g with the consortium (106 cells/ml). Different hydrocarbons were extracted by Soxhlet extraction and examined by using gas chromatography and flame ionization detector (GC-FID). Consortium degraded more than 63% total petroleum hydrocarbons, and significant degradation (80%) of n-alkanes (nC13 to nC29) was witnessed with consortium + fertilizer + alfalfa in 30% oily sludge at 3 days. Degradation of (80%) of n-alkanes (nC13 to nC29) in oily sludge was also noted. In this study, the consortium along with Alfalfa and fertilizer found most efficient option for remediation of hydrocarbons.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
In the last few years, a reasonable amount of oily sludge has been drilled out and stored in open pits within the surroundings of different oil field areas. Moreover, these oil-based industries are also spilling out oily waste to the environment which poses constant threat to the agro-environmental ecosystems. Oily sludge is made up of a wide range of cancer-causing organic or inorganic compounds which are toxic to the microorganisms and plants and also acts as a source of groundwater contamination. These toxic hydrocarbons may enter into the food chain of aquatic and marine life, and hence interrupts their response to chemoreception (Siddiqui et al. 2001). Furthermore, some other disturbing factors may also be toxic but these cancer-causing compounds in oily sludge could lead to remove many of the plant species and marine organisms (Bhattacharya et al. 2003).
Bioremediation is considered one of the finest options to remediate soils which are contaminated with oily sludge in all over the world. Bioremediation involves the use of various genera of microbes as a consortium capable of degrading complex mixture of organic compounds (Dhote et al. 2017; Ite and Ibok 2019; Joo et al. 2008; Reshma et al. 2014). Contradictory evidences are available in the literature that either consortium alone or in combination with fertilizer can enhance the degradation process. Some authors reported that consortium was not as effective alone as with fertilizer when the contamination level is 100% (Alexander 1999). This is probably due to deficiency of essential nutrients in oily sludge required for microbial growth. Microbes in association with plants may be a better option. Plant roots penetrate downward in the soil, thereby increasing the aeration and nutrients availability required for microbial proliferation (Gaskin et al. 2008). It has been reported previously that more than 1000-fold increase in microbial counts were noted in vegetated soil than non-vegetated soils (Siciliano et al. 2003; Das and Chandran 2010; Silva-Castro et al. 2012). The process involves strong interaction between plant root exudates and microbes (Daniel 2004; Nie et al. 2011; Weronika et al. 2013). Rhizoremediation primarily depends on the selection of appropriate plant species that can grow well under oily sludge–contaminated environment (Fang et al. 2001; Escalante-Espinosa et al. 2005; Farinazleen et al. 2004; Huang et al. 2004, Huang et al. 2005; Euliss et al. 2008). Leguminous plants such as alfalfa grew well in soils contaminated with oily sludge, and increased degradation of hydrocarbons was observed in leguminous as compared with non-leguminous plants. This is most probably because of the presence of nodules on the root surface that provide enough surface area and fixed nitrogenous compounds necessary for microbial growth. Schwab et al. (2007) and Kirk et al. (2005) reported that under hydrocarbon stress environment, rhizobacteria can enhance the level of degradation of hydrocarbons.
Plant releases root exudates such as sugars, amino acids, and organic acids for microbes to grow under oily sludge–contaminated rhizosphere (Böltner et al. 2008; Gerhardt et al. 2009). Plant root zone is composed of several classes of microbes which are able to degrade numerous groups of hydrocarbon such as alkanes, cycloalkanes, or PAHs in association with root exudates of tolerant plant which help to remediate oily sludge–contaminated soils (Kuiper et al. 2004, Tang et al. 2010 and 2012; Glick and Stearns 2011).
Little information is available on the role of alfalfa with consortium for the degradation of various groups of hydrocarbon in oily sludge–contaminated soils (Chaudhry et al. 2005). Like alfalfa, maize is also known (Ali et al. 2010) as tolerant to oily sludge contamination most likely because of its greater biomass and well-established root system.
Bacteria faces lag phases in oily sludge which differ depending upon the nature and amount of hydrocarbons present in soil contaminated with oily sludge (Geetha et al. 2013; Palanisamy et al. 2014; Yousefi et al. 2009). Cerqueira et al. 2011 isolated Stenotrophomonas acidaminiphila, Bacillus cibi, and Bacillus megaterium, from petroleum oily sludge and Pseudomonas aeruginosa and Bacillus cereus from oily sludge of Brazil capable of degradation of hydrocarbons. Liu et al. (2010) isolated Bacillus altitudinous from oily sludge–contaminated site and demonstrated that application of such bacteria as inoculum to benzene contaminates soils and significantly enhances the degradation of benzene.
Hydrocarbon contamination is a worldwide threat to the soil ecosystem. The present study was designed to evaluate the remediation potential of bacterial consortium, viz. Bacillus cereus (Acc KR232400), Bacillus altitudinis (Acc KF859970), Commomonas (Delftia sp) (Acc KF859971), and Stenotrophomonas maltophilia (Acc KF859973) with alfalfa and maize plants in combination with fertilizer to remediate oily sludge. Each strain was screened in the laboratory for its capability to degrade total petroleum hydrocarbons (TPHs) (Bano et al. 2015, Shahzad et al. 2015).
Materials and method
Study area
The study area is situated in the south of the Himalaya and Karakorum Mountains named as Kohat-Potwar Plateau (Table 1). It is confined by the Salt Range and Trans-Indus Range in the south and Kalachitta Range in the north (Asif and Fazeelat 2012). Potwar Plateau is an oily prone area. The first oil well was drilled in 1866 near an oil seep at Kundal, Pakistan (Khan et al. 1986a, 1986b). For this study, oily sludge from three different oil fields in this area was collected which varies in their reservoir depth, geological structure and age, API gravity, extent of degradation, and crude oil composition.
The Kohat-Potwar Plateau is a hydrocarbon prone area. According to the previous literature, several source rocks range in age from the Proterozoic-Lower Cambrian to Paleocene and Eocene has been identified until now as a potential rock for hydrocarbons (Quadri and Shuaib 1986; Khan et al. 1986a, 1986b). The Salt Range Formation mainly carbonates is considered to be the oldest rocks shown potential for hydrocarbons whereas the Sardhai and the Chhidru Formation sandy in nature and the Patala Formation usually shales are the most recent source rocks within the Kohat-Potwar Plateau (Jaswal et al. 1997; Ahmad et al. 1996). The potential reservoirs of the Potwar Plateau are Cambrian Khewra and Jutana Formations, Permian Tobra Formation, Jurassic Datta Formation, Cretaceous Chichali Formation, Paleocene Hangu, Locakhart and Patala Formations, and Eocene Sakaser and Chorgali Formations.
Sampling of oily sludge
The oily sludge was collected at a depth of 3.65 cm (because bedrock started after this) from the waste pit of the Missa Kasswal oil field measuring (100 × 100 2) wide and 30.48 m long. The sample texture was clay loam with pH 4.5 and 20% organic matter. The soil pH was determined by Grossman and Reinsch (2020). The soil texture was determined by using the method of Gee and Or (2002). The soil organic matter was determined by using the method of Nelson and Sommers (1983).
Determination of viable bacterial population
The viable bacterial population was determined using the pour plate method as described by Song and Bartha (1990). Suspension of subsamples of sludge (1 g) taken in triplicate was made by adding 9 ml of sterile water to 1 g of sludge sample from each treatment, stirred on a magnetic stirrer. The suspension was centrifuged at 3000 rpm for 10 min. Supernatant was removed, and decimal dilutions were prepared. Aliquots (80 μl) of each dilution () were spread over autoclaved nutrient agar plates (peptone 0.5%, Beef extract/yeast extract 0.3%, agar 1.5%, NaCl 0.5%, pH 7.4) and were incubated at 32 ± 2 °C for 24 h. The bacterial colonies were counted on dilution plates and calculated by the given formula
Consortium preparation
The bacterial strains used in consortium, viz. Bacillus cereus, Bacillus altitudinis, Commomonas, and Stenotrophomonas maltophilia, were isolated from different oil fields of the study area and further selected on the basis of their degradation potential (Bano et al. 2015). The inoculum was prepared as described by Shahzad et al. (2015). The bacterial strains used in consortium were isolated from oil field of the Potwar Plateau in Pakistan and identified by 16S rRNA gene sequencing. For the preparation of consortium, the nutrient broth was purchased from OXOID-UK and autoclaved at 121 °C for 45 min. The isolated strains were inoculated in nutrient broth and incubated in shaker incubator (EXCELLA E24 Germany) at 150 rpm for 48–72 h. Thereafter, the culture was centrifuged at 3000 rpm for 10 min. The pellet was re-suspended in autoclaved distilled water, and the optical density (OD) was adjusted to 1 at 660 nm with a UV-VIS spectrophotometer. The consortium was prepared by mixing pure cultures of four bacterial strains in equal volume of individual strain having an OD of 1 at 660 nm and bacterial density (106 cells/ml). This inoculum was used as working pure culture of the isolated strain. The consortium was prepared by adding pure cultures of four bacterial strains in equal volume.
Soil sample preparation
Samples of oily sludge were placed over three large aluminum trays separately. The oily sludge was hand-mixed with soil thoroughly to ensure homogeneous mixing in the ratio of 30:70 of oily sludge to soil. The application of bacterial consortium was made with fertilizer and without fertilizers, in combination with plant and without plants. Pots measuring 9 × 9 cm2 were filled with 1 kg (1000 g) of 30% sludge. The consortium was added in the pots and was applied in 1:1 ratio (v/v) with fertilizer solution. For fertilizer treatment, aqueous solution of NH4NO3 and (NH4)3PO4 having N to P ratio of 10:1 was pipetted over the oily sludge at the rate of 70 μg/ml and 7 μg/ml, respectively. The seeds of alfalfa and maize were also inoculated with consortium prior to sowing. The inoculation was conducted by soaking the seeds at room temperature for 3–4 h in inoculum suspension prepared earlier. The pots with plants were kept at room temperature having 14-h photoperiod and relative humidity (RH) varied from 75 to 80%.
About 10 g of oily sludge from each treatment was removed at 3, 7, 14, 21, 28, and 60 days of incubation, and the degradation of hydrocarbons was determined by gas chromatography and flame ionization detector (GC-FID) at different intervals. The treatments made are given in Table 2.
Crude oil extraction and fractioning
The different fractions of hydrocarbon were extracted by using Soxhlet method as described by Reeve (1994). The Soxhlet extraction was carried out for each control and treated oily sludge samples in triplicate. The temperature was raised slowly from 20 to 40 °C over a period of 24 h. Solvents used were n-hexane (35 ml) and dichloromethane (100 ml).
Ten-gram triplicates of the control/treated soil samples were removed from the bulk soil samples and were placed in the known weight of cellulose thimble. A cellulose thimble (25-mm neck size and 100 mm in length) with 10 g of soil was placed in the glass Soxhlet extractor and was fixed over the round-bottom flasks which were filled with solvents as mentioned above. A round-bottom flask was placed over the heater and allowed to heat overnight slowly. Thereafter, the oil extract was removed from the round-bottom flask and was placed in a 250-ml glass bottle. Cellulose thimble was removed and was allowed to dry in the laboratory. Once dried, the weight of cellulose thimble was recorded. The difference in the weight before and after the Soxhlet extraction was recorded to determine the amount of total hydrocarbons extracted from the control and treated soils. Thereafter, the Soxhlet extracted solution was placed over the known weight of a roto-evaporator flask and was allowed to evaporate. Once all the solvent was removed, the dried solution was obtained. The weight of the flask was recorded again. This difference in the weight of solution before and after evaporation will gave us the information about the quantity of crude oil extracted from 10 g of soil samples contaminated with oily sludge or treated.
Gas chromatography
The GC-FID analysis was made by using a QC 2010 Shimadzu GC-FID (Japan). The GC-FID was performed as described by Shahzad et al. (2015). Gas chromatography was performed using QC 2010 GC-FID of Shimadzu. This GC was equipped with capillary column of length 30 m and internal diameter of 0.25 mm and 5% phenyl 95% methyl polysiloxane stationary phase of coating of 0.25 μm of thickness (DB-5 MS, J&W Scientific). A sample of 1 μl was removed from bulk liquid sample of (5 ml) and was placed in 2 μl of GC vial. It was diluted with dichloromethane up to the 2 μl mark of the vial and was covered with perforated cap. The vial was placed over the auto sampler, and the GC-FID method was developed to run the sample. The injector and detector (FID) temperature was kept at 280 °C in a pulsed splitless mode. The GC oven was programmed from 75 to 320 °C at 10 °C/min with initial and final hold times of 28.83 min, respectively. Helium was used as a carrier gas, and the flow was maintained constantly at the rate of 1.1 ml/min.
Statistical analysis
The research trial was conducted by using completely randomized design (CRD), and data was analyzed using the Statistix 8.1. The results of total petroleum hydrocarbon were compared by means using least significant difference (LSD) and standard error for all other fractions.
Results
Effect of bacterial consortium on the degradation of total hydrocarbon in oily sludge
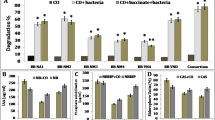
The degradation potential of consortium in oily sludge–contaminated soil is shown in the Fig. 1. The constructed bacterial consortium improved the degradation of total hydrocarbons which was assisted by alfalfa and maize with fertilizer addition. The GC-FID analysis of the oily sludge showed that total hydrocarbons were 47.3 mg/g when extracted with dichloromethane immediately. Negligible degradation of hydrocarbons was noted in all treatments (T1 to T12) at day 0 of incubation. Extraction of day 0 was made immediately after adding consortium, fertilizer, alfalfa, or maize to oily sludge–contaminated soils (T1 to T12). The oily sludge with consortium addition (T2) accelerated the rate of degradation of total hydrocarbons as compared with untreated oily sludge (T1) at day 3. More than 63% of hydrocarbons were degraded with consortium (T2) than untreated oily sludge at day 3. Adding fertilizer (T3) also increased the degradation of total hydrocarbons than untreated oily sludge (T1) at day 3 of incubation; however, the degradation was slower than T2. Combining consortium (T2) and fertilizer (T3), the degradation of TPHs was not improved over a short period of time. Planting alfalfa (T5) also accelerated the degradation of total hydrocarbons than untreated oily sludge (T1), and about 40% of total petroleum hydrocarbons (TPH) were degraded at day 3. Nonsignificant differences in the degradation of total hydrocarbons were observed between T4 and T5. The most effective degradation of TPHs was recorded when consortium, alfalfa, and fertilizer were used in combination (T8). About 80% of (TPHs) were degraded at day 3.

Total hydrocarbon degradation. Alphabets on the bar represent compared means by using LSD values (complete randomize design), and bars having similar letters do not differ significantly; otherwise, they differ at p < 0.05
Planting maize (T9) showed slow degradation of (TPH) in oily sludge than planting alfalfa (T4) at day 14. No change in the rate of degradation of hydrocarbons was observed between maize and alfalfa until day 14. Similarly, consortium application with maize (T10) was not as effective as was with alfalfa (T6). The same trend was observed when fertilizer was added to maize in oily sludge (T11). The bacterial consortium was effective alone but further improvement with maize and fertilizer was not observed even after a long period of time.
Degradation of n-alkanes
Table 3 represents the effect of bacterial consortium on the degradation of all the fractions of hydrocarbons at different incubation intervals. The degradation of different fractions was observed with the application of consortium, fertilizer, alfalfa, and maize. The n-alkanes nC13-nC16 and nC30-nC33 were present in small concentrations whereas nC19-nC29 was present at high concentration (30.2 mg per g of oily sludge). Addition of bacterial consortium in oily sludge (T2) decreased the concentration of nC19-nC29 from 30.2 to 9.2 mg/g at day 3. Similarly, the degradation of nC13-nC16, and nC30-nC33 was also observed at day 3. Similar trend was observed in the degradation of nC17-nC18. However, no further hydrocarbon degradation was observed with alfalfa and fertilizer. The factions of nC30-nC33 were not detected at day 3 in oily sludge with consortium + alfalfa + fertilizer (T8). Maize cannot boost the degradation rate of any carbon chain as compared with a short period of time. Maize delayed the degradation of nC19-nC29, the most abundant carbon chain than the alfalfa.
Degradation of isoprenoids
Table 4 shows the degradation of isoprenoids (non-farnesane, pristane, and phytane) at different intervals. Isoprenoids in oily sludge were present in low concentration such as non-farnesane; pristane and phytane were present at concentrations 0.74 ± 0.02 and 0.5 ± 0.07 mg per kg in oily sludge–contaminated soil at day 3. The concentration of non-farnesane was 0.19 mg per kg at day 21 which was not detected at day 28. Consortium with fertilizer (T4) accelerated the rate of degradation of non-farnesane at shorter time and was not detected after day 7. Pristane and phytane were degraded slowly and about 0.20 mg per kg was present at day 60. The rate of degradation of non- farnesane was rapid in oily sludge with consortium, alfalfa, and maize (T8 and T10) at day 3. Thereafter, the rate of degradation was slow and negligible amount of such compound was present at the end of incubation period. Pristane and phytane were slowly degraded, and around 50% of these compounds were degraded in oily sludge with consortium alone (T2) and with fertilizer (T4) at shorter time.
Bacterial population
Around 106 bacterial cells were inoculated with consortium. Table 5 indicates that the bacterial population in T1 was lower at initial day which increased at days 3 and 7 which again decreased at day 14 until day 28. However, with application in oily sludge (T2), the bacterial population was increased at day 3. Fertilizer addition (T3) did not accelerate the bacterial population during incubation period. However, consortium with fertilizer (T4) increased the bacterial population at day 7. Planting alfalfa (T5) also increased the bacterial population at days 3 to 21. Alfalfa with fertilizer (T6) increased the bacterial population at day 3 whereas it was further increased at day 7. Fertilizer with alfalfa (T7) increased bacterial population at day 7. Consortium, fertilizer, and alfalfa (T8) increased the bacterial population at day 7 which was decreased at day 21 and remained more or the same until day 28. When maize was grown over oily sludge (T9) or in combination with or without consortium or fertilizer (T10, T11, and T12), the bacterial population at day 7 to 28 increased. Maize increased the bacterial population 100-fold from day 3 to day 21 of incubation period.
Discussion
Microbial consortium is used to remediate oily sludge–contaminated soils. This study shows that microbial consortium prepared from Bacillus cereus (Acc KR232400), Bacillus altitudinis (Acc KF859970), Commomonas (Delftia sp.) (Acc KF859971), and Stenotrophomonas maltophilia (Acc KF859973) accelerated the rate of degradation of total petroleum hydrocarbons (TPHs) over a short period of time which indicated that consortium was the best option to remediate oily sludge and these findings are in agreement with the results of Mishra et al. (2001) who demonstrated that consortium prepared from Acinetobacter baumannii, Burkholderia cepacia, and Pseudomonas was capable of degrading TPHs from 69.7 to 5.53 g/kg in 360 days of incubation. The mode of degradation of microbes in oily sludge–contaminated soils is because of β-oxidation. Microbes utilize petroleum hydrocarbons as a carbon or energy source and convert these hydrocarbons into carbon dioxide, fatty acids, and water via β-oxidation (Alvarez 2003; Genard et al. 2013). Dong et al. (2015) reported that Gammaproteobacteria with abundance of alkB genes facilitate TPH degradation. Microbes also require nutrients to further speed up the process of degradation of TPHs in oily sludge. However, the required concentration or ratio of nitrogen and phosphorus is quite variable. Soil deficient in nitrogen and phosphorus may require more N to P than soils sufficient in these nutrients. The oily sludge–contaminated soils of this study were deficient in N to P; therefore, a ratio of 10:1 was enough to degrade TPHs over a short period of time. This is in consistent with the findings of Vieira et al. (2009) who reported that nutrient ratio (C:N:P = 100:20:2.7) and inoculum concentration (1.32 g/l) accelerate the degradation of TPHs and around 71.8% of such compounds were degraded at day 3 of incubation. In this study, a ratio of C to N to P of 30:10:1 was added to the soil with an idea that this ratio was enough to degrade TPHs with microbial consortium. However, the results of this study show that fertilizer alone did not accelerate the degradation of TPHs, but when added with consortium, it enhanced the rate of degradation of TPHs over a short period of time. Plants in combination with microbial consortium are considered an effective medium to further accelerate the rate of degradation of TPHs. The result of this study shows that the level of degradation of hydrocarbons was improved in the presence of fertilizer and alfalfa as compared with consortium alone in a short period of time. The data revealed that consortium can boost up the frequency of degradation of TPHs over a short period of time (Table 2). However, the level of degradation of hydrocarbons was improved in the presence of fertilizer and alfalfa as compared with a short period of time.
The rate of degradation of TPHs was further improved in the presence of an alfalfa plant and fertilizer as compared with the short period of time in the present investigation. These findings are similar with the research outcomes of Kumar et al. (2013). They demonstrated that addition of bacterial consortium to hydrocarbon-contaminated soil was increased when consortium was added under wheat (Triticumaestivum) and mustard (Brassica juncea), respectively at 15 days of incubation. They concluded that consortium was effective under wheat or mustard rhizosphere contaminated with hydrocarbons. Ali et al. (2010) used plants such as maize (Zea mays L.) and tall fescue (Festuca arundinacea) with nutrients to remediate oily sludge–contaminated soils and reported that around 96.3% of (TPHs) were degraded from oily sludge–contaminated soils at day 120 of incubation period.
The results of the present study demonstrate that enhanced hydrocarbon degradation present in oily sludge treated with consortium cannot be directly linked with presence of increased bacterial counts in general. The enhanced TPH degradation in alfalfa with consortium may be attributed to maximum availability of carbon sources required for bacterial growth; furthermore, the alfalfa being a nodulating and N-fixing plant provides fixed N as N source to assist the process of bioremediation (Kirk et al. 2005; Phillips et al. 2009). Maize with consortium delayed the degradation rate of total hydrocarbons and was not as effective as alfalfa to remediate oily sludge–contaminated soils over a short period of time. However, maize increased the bacterial counts by 100-fold at different intervals, and this increase was not linked with increase in degradation of TPHs. Possibly, maize needs a longer lag period and it requires more water and nutrient to support its own growth. The lack of any significant increase or even observed decrease in the bacterial population could be attributed to the fact that the toxicity of hydrocarbons depressed the activity and the proliferation of rhizosphere microorganisms (Merkl et al. 2004, 2005; Muthuswamy et al. 2008). Different varieties of plants produce root exudates of different composition which influence growth of microbial communities; plants resistant to toxic hydrocarbons may have formed certain root exudates which directed the stimulation of specific microbes (including hydrocarbon-utilizing microorganisms); these microflora play vital role in the reduction of toxicity linked with these hydrocarbons (Germida et al. 2002). The greater CFU of microbes from maize rhizosphere grown in 30% sludge is possibly due to its greater biomass and specific composition of root exudates which selectively increase the bacterial population not active in degradation of hydrocarbon as opposed to that in alfalfa, a nodulating leguminous plant. Differences in the degradation abilities were not related with variations in the major microbial population structure or with considerable changes in general microbial abundance but determined by the functional changes in the microbial communities. Even different plants of the same species (Lesuffleur et al. 2007) and even in same variety (Cieśliński et al. 1998; Micallef et al. 2009) can have markedly diverse types of exudation forms in the same growing conditions. The variation in exudation within intra-species has been shown to have beneficial effects on the composition of microbial communities present in rhizosphere (Micallef et al. 2009) and on bacterial gene expression (Mark et al. 2005).
The rate of degradation of TPHs was further improved in the presence of alfalfa plant and fertilizer as compared with consortium alone or in combination with fertilizer but without alfalfa over a short period of time in the present investigation. These findings are similar with the research outcomes of Kumar et al. (2013). The present results are in accordance with the reports of Bento et al. (2005). They demonstrated that bacterial consortium (Bacillus cereus, Bacillus sphaericus, Bacillus fusiformis, Bacillus pumilus, Acinetobacter junii, and Pseudomonas sp.) added to diesel-contaminated soils increased the degradation of nC12-nC23 and nC23-nC40 over 7 days of incubation period. Furthermore, it was demonstrated that the bacterial consortium is capable of degrading light and heavy carbon chain compounds from diesel-contaminated soils.
Further degradation of nC19-nC29 was facilitated in the presence of fertilizer and alfalfa over a short period of time. The results of this study are similar with the results of Plohl et al. (2002) who demonstrated that when unidentified bacterial strain AL-12 from the culture of cyanobacteria was inoculated to motor oil–contaminated soils, the rate of degradation of n-C15 to n-C40 was 98% at day 50 of incubation. However, around 70% n-alkanes n-C15-n-C22, up to 45% of n-C22-n-C30, and up to 20% of n-C30-n-C40 were biodegraded at 7 days of incubation in motor oil–contaminated soils with cyanobacteria unidentified strains than oil-contaminated soils without bacterial strains. Pristane and phytane were found to be the most persistent hydrocarbons and remained in the soil until the end of the incubation period. However, consortium was able to degrade pristane and phytane more than 50% within 3 days of incubation.
Conclusion
The study indicated that TPH degradation was enhanced by constructed consortium which was further accelerated in the presence of alfalfa and fertilizer at room temperature. Persistent hydrocarbons, pristane and phytane, can also be degraded significantly by the consortium. However, maize also helped in degradation but the rate of degradation of TPHs was slower as compared with alfalfa in the presence of consortium and fertilizer. It is anticipated that maize bacteria association may require longer lag period to accelerate the process of degradation of hydrocarbon from oily sludge. The alfalfa with fertilizer and consortium is a suitable combination and recommended for rapid degradation. The degradation can be completed at room temperature with minimal requirement for N and P. The method presented here is economical and environment-friendly.
References
Ahmad S, Alam Z, Khan AR (1996) Petroleum exploration and production activities in Pakistan: Pakistan Petroleum Information Service, 72 p
Ali DZ, Gholamreza NB, Nasser M (2010) Phytoremediation of total petroleum hydrocarbons (TPHs) using plant species in Iran. Turk J Agric 34:429–438
Alexander M (1999) Biodegradation and bioremediation. Gulf Professional Publishing, Houston
Alvarez HM (2003) Relationship between β-oxidation pathway and the hydrocarbon-degrading profile in actinomycetes bacteria. Int Biodeterior Biodegradation 52(1):35–42
Asif M, Fazeelat T (2012) Petroleum geochemistry of the Potwar Basin, Pakistan: II–oil classification based on heterocyclic and polycyclic aromatic hydrocarbons. Appl Geochem 27:1655–1665
Bano A, Shahzad A, Siddiqui S (2015) Rhizodegradation of hydrocarbon from oily sludge. J Bioremediation Biodegrad 6:2
Bhattacharya D, Priyangshu M, Sarma S, Krishnan SM, Banwari L (2003) Evaluation of genetic diversity among Pseudomonas citronellolis strains isolated from oily sludge-contaminated sites. Appl Environ Microbiol 69:1435–1441
Bento FM, Camargo FA, Okeke BC, Frankenberger WT (2005) Comparative bioremediation of soils contaminated with diesel oil by natural attenuation, biostimulation and bioaugmentation. Bioresour Technol 96:1049–1055
Böltner D, Godoy P, Muñoz-Rojas J, Duque E, Moreno-Morillas S, Sánchez L, Ramos JL (2008) Rhizoremediation of lindane by root-colonizing Sphingomonas. Microb Biotechnol 1:87–93
Cerqueira VS, Hollenbach EB, Maboni F, Vainstein MH, Camargo FA, Maria do Carmo RP, Bento FM (2011) Biodegradation potential of oily sludge by pure and mixed bacterial cultures. Bioresour Technol 102:11003–11010
Chaudhry Q, Blom-Zandstra M, Gupta SK, Joner E (2005) Utilising the synergy between plants and rhizosphere microorganisms to enhance breakdown of organic pollutants in the environment (15 pp). Environ Sci Pollut Res 12:34–48
Cieśliński G, Van Rees K, Szmigielska A, Krishnamurti G, Huang P (1998) Low-molecular-weight organic acids in rhizosphere soils of durum wheat and their effect on cadmium bioaccumulation. Plant Soil 203:109–117
Daniel JG (2004) Infection and invasion of roots by symbiotic, nitrogen-fixing rhizobia during nodulation of temperate legumes. Microbiol Mol Biol R 68:280–300
Das N, Chandran P (2010) Microbial degradation of petroleum hydrocarbon contaminants: an overview. Biotechnol Res Int 2011:1–13
Dhote M., Kumar A., Jajoo, A., Juwarkar A (2017). Assessment of hydrocarbon degradation potentials in a plant-microbe interaction system with oil sludge contamination: a sustainable solution. Int J Phytoremediation, 19(12), 1085–1092
Dong Y, Lang Z, Kong X, Lu D, Liu Z (2015) Kinetic and multidimensional profiling of accelerated degradation of oil sludge by biostimulation. Environ Sci Process Impacts 17(4):763–774
Escalante-Espinosa E, Gallegos-Martínez M, Favela-Torres E, Gutiérrez-Rojas M (2005) Improvement of the hydrocarbon phytoremediation rate by Cyperus laxus Lam. inoculated with a microbial consortium in a model system. Chemosphere 59:405–413
Euliss K, C-h H, Schwab A, Rock S, Banks MK (2008) Greenhouse and field assessment of phytoremediation for petroleum contaminants in a riparian zone. Bioresour Technol 99:1961–1971
Fang C, Radosevich M, Fuhrmann JJ (2001) Atrazine and phenanthrene degradation in grass rhizosphere soil. Soil Biol Biochem 33:671–678
Farinazleen MG, Raja NZ, Rahman A, AbuBakar S, Mahiran B (2004) Biodegradation of hydrocarbons in soil by microbial consortium. Int J Bioremediat Biodegr 54:61–67
Gaskin S, Soole K, Bentham R (2008) Screening of Australian native grasses for rhizoremediation of aliphatic hydrocarbon-contaminated soil. Int J Phytoremediation 10:378–389
Gee GW, Or D (2002) 2.4 Particle-size analysis. Methods Soil Anal Part 4 Phys Methods 1(5):255–293
Geetha SJ, Joshi SJ, Kathrotiya S (2013) Isolation and characterization of hydrocarbon degrading bacteria isolate from oil contaminated sites. APCBEE Procedia 5:237–241
Genard B, Miner P, Nicolas JL, Moraga D, Boudry P, Pernet F, Tremblay R (2013) Integrative study of physiological changes associated with bacterial infection in Pacific oyster larvae. PLoS One 8(5):e64534
Gerhardt KE, Huang X-D, Glick BR, Greenberg BM (2009) Phytoremediation and rhizoremediation of organic soil contaminants: potential and challenges. Plant Sci 176:20–30
Germida J, Frick C, Farrell R, Violante A, Huang P, Bollag J, Gianfreda L (2002) Phytoremediation of oil-contaminated soils. Ecological significance of the interactions among clay minerals, organic matter and soil biota 3rd Symposium on Soil Mineral-Organic Matter-Microorganism Interactions and Ecosystem Health, Naples-Capri, Italy, 22-26 May 2000. Elsevier Science BV, pp 169-186
Glick BR, Stearns JC (2011) Making phytoremediation work better: maximizing a plant’s growth potential in the midst of adversity. Int J Phytoremediation 13:4–16
Grossman RE, Reinsch TG (2020) The solid phase. In: Dane JH, Topp GC (eds) Methods of soil analysis, part 4, physical methods, Book Series No. 5. Soils Science Society of America, Madison, pp 201–225
Huang XD, El-Alawi Y, Penrose DM, Glick BR, Greenberg BM (2004) A multi-process phytoremediation system for removal of polycyclic aromatic hydrocarbons from contaminated soils. Environ Pollut 130:465–476
Huang X-D, El-Alawi Y, Gurska J, Glick BR, Greenberg BM (2005) A multi-process phytoremediation system for decontamination of persistent total petroleum hydrocarbons (TPHs) from soils. Microchem J 81:139–147
Ite AE, Ibok UJ (2019) Role of plants and microbes in bioremediation of petroleum hydrocarbons contaminated soils. Int J 7(1):1–19
Jaswal TM, Lillie RJ, Lawrence RD (1997) Structure and evolution of the northern Potwar deformed zone, Pakistan. Am Assoc Pet Geol Bull 81(2):308–328
Joo H-S, Ndegwa PM, Shoda M, Phae C-G (2008) Bioremediation of oil-contaminated soil using Candida catenulata and food waste. Environ Pollut 156:891–896
Khan M, Ahmed R, Raza HA, Kemal A (1986a) Geology of petroleum in Kohat-Potwar depression, Pakistan. AAPG Bull 70:396–414
Khan MA, AhmedR RHA, Kemal A (1986b) Geology of petroleum in Kohat-Potwar depression, Pakistan. Am Assoc Pet Geol Bull 70(4):396–414
Kirk JL, Klironomos JN, Lee H, Trevors JT (2005) The effects of perennial ryegrass and alfalfa on microbial abundance and diversity in petroleum contaminated soil. Environ Pollut 133:455–465
Kuiper I, Lagendijk EL, Bloemberg GV, Lugtenberg BJ (2004) Rhizoremediation: a beneficial plant-microbe interaction. Mol Plant-Microbe Interact 17:6–15
Kumar R, Bharagava RN, Kumar M, Singh SK, Govind K (2013) Enhanced biodegradation of mobile oil hydrocarbons by biosurfactant producing bacterial consortium in wheat and mustard rhizosphere. J Pet Environ Biotechnol 4:2
Lesuffleur F, Paynel F, Bataillé M-P, Le Deunff E, Cliquet J-B (2007) Root amino acid exudation: measurement of high efflux rates of glycine and serine from six different plant species. Plant Soil 294:235–246
Liu WX, Luo YM, Teng Y, Li ZG, Ma LQ (2010) Bioremediation of oily sludge-contaminated soil by stimulating indigenous microbes. Environ Geochem Hlth 32:23–29
Mark GL, Dow JM, Kiely PD, Higgins H, Haynes J, Baysse C, Abbas A, Foley T, Franks A, Morrissey J (2005) Transcriptome profiling of bacterial responses to root exudates identifies genes involved in microbe-plant interactions. Proc Natl Acad Sci U S A 102:17454–17459
Meissner CR, Master M, Rashid MA, Hussain M (1974) Stratigraphy of Kohat Quadrangle, Pakistan. United States Geological Survey, Reston Professional Paper; 716–D
Merkl N, Schultze-Kraft R, Infante C (2004) Phytoremediation in the tropics—the effect of crude oil on the growth of tropical plants. Bioremediation J 8:177–184
Merkl N, Schultze-Kraft R, Infante C (2005) Assessment of tropical grasses and legumes for phytoremediation of petroleum-contaminated soils. Water Air Soil Pollut 165:195–209
Micallef SA, Shiaris MP, Colón-Carmona A (2009) Influence of Arabidopsis thaliana accessions on rhizobacterial communities and natural variation in root exudates. Journal of Experimental Botany:erp053
Mishra S, Jyot J, Kuhad RC, Lal B (2001) Evaluation of inoculum addition to stimulate in situ bioremediation of oily-sludge-contaminated soil. Appl Environ Microbiol 67:1675–1681
Muthuswamy S, Arthur RB, Sang-Ho B, Sei-Eok Y (2008) Biodegradation of crude oil by individual bacterial strains and a mixed bacterial consortium isolated from hydrocarbon contaminated areas. Clean 36:92–96
Nelson DW, Sommers L (1983) Total carbon, organic carbon, and organic matter. In: Methods of soil analysis: part 2 chemical and microbiological properties, vol 9, pp 539–579
Nie M, Wang Y, Yu J, Xiao M, Jiang L, Yang J, Fang C, Chen J, Li B (2011) Understanding plant-microbe interactions for phytoremediation of petroleum-polluted soil. PLoS One 6:e17961
Palanisamy N, Ramya J, Kumar S, Vasanthi N, Chandran P, Khan S (2014) Diesel biodegradation capacities of indigenous bacterial species isolated from diesel contaminated soil. J Environ Health Sci Eng 12:142
Phillips LA, Greer CW, Farrell R, Germida JJ (2009) Field-scale assessment of weathered hydrocarbon degradation by mixed and single plant treatments. Appl Soil Ecol 42:9–17
Plohl K, Leskovsek H, Bricelj M (2002) Biological degradation of motor oil in water. Acta Chim Slov 49:279–290
Quadri, Viqar-un-Nisa, Shuaib SM, (1986) Hydrocarbon prospects of southern Indus Basin, Pakistan: American Association of Petroleum Geologists Bulletin, v. 70, no. 4, p. 396–414
Reeve RN (1994) Environmental analysis. Wiely, Chichester, pp 149–151
Reshma J, Thanga VSG, Mathew A (2014) Phenol degradation by bacteria isolated from coir retting beds of South Kerala coast. J Glob Biosci 3:935–940
Schwab S, Ramos HJ, Souza EM, Pedrosa FO, Yates MG, Chubatsu LS, Rigo LU (2007) Identification of NH 4+-regulated genes of Herbaspirillum seropedicae by random insertional mutagenesis. Arch Microbiol 187(5)379–386
Shah SMI (1977) Stratigraphy of Pakistan: Geological Survey of Pakistan, Memories, 12, 137
Shahzad A, Siddiqui S, Bano A (2015) The response of maize (Zea mays L.) plant assisted with bacterial consortium and fertilizer under oily sludge. Int J Phytoremediat 18(5):521–526. https://doi.org/10.1080/15226514.2015.1115964
Siciliano SD, Germida JJ, Banks K, Greer CW (2003) Changes in microbial community composition and function during a polyaromatic hydrocarbon phytoremediation field trial. Appl Environ Microbiol 69:483–489
Siddiqui S, Adams WA, Schollion J (2001) The phytotoxicity and degradation of diesel hydrocarbons in soil. J Plant Nutr Soil Sci 164:631–635
Silva-Castro G, Uad I, Gónzalez-López J, Fandino C, Toledo F, Calvo C (2012) Application of selected microbial consortia combined with inorganic and oleophilic fertilizers to recuperate oil-polluted soil using land farming technology. Clean Techn Environ Policy 14:719–726
Song H-G, Bartha R (1990) Effects of jet fuel spills on the microbial community of soil. Appl Environ Microbiol 56:646–651
Tang JC, Wang RG, Niu XW, Wang M, Chu HR, Zhou QX (2010) Characterization on the rhizoremediation of petroleum contaminated soil as affected by different influencing factors. Biogeosciences 7:3961–3969
Tang J, Lu X, Sun Q, Zhu W (2012) Aging effect of petroleum hydrocarbons in soil under different attenuation conditions. Agric Ecosyst Environ 149:109–117
Vieira PA, Faria S, Vieira RB, De Franca FP, Cardoso VL (2009) Statistical analysis and optimization of nitrogen, phosphorus, and inoculum concentrations for the biodegradation of petroleum hydrocarbons by response surface methodology. World J Microbiol Biotechnol 25(3):427–438
Weronika G, Piotr L, Alicja S, Łukasz C, JoannaW K (2013) Bio-augmentation with petroleum-degrading consortia has a selective growth-promoting impact on crop plants germinated in diesel oil-contaminated soil. Water Air Soil Pollut 224:1676
Yousefi K, Khodadadi A, Ganjidoust H, Badkoubi A, Amoozegar MA (2009) Isolaton and characterization of a novel native Bacillus strain capable of degrading diesel fuel. Int J Environ Sci Technol 6:435–442
Funding
This project received financial support from the Government of Pakistan and Higher Education Commission (HEC).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Amjad Kallel
Rights and permissions
About this article

Cite this article
Shahzad, A., Siddiqui, S., Bano, A. et al. Hydrocarbon degradation in oily sludge by bacterial consortium assisted with alfalfa (Medicago sativa L.) and maize (Zea mays L.). Arab J Geosci 13, 879 (2020). https://doi.org/10.1007/s12517-020-05902-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12517-020-05902-w