
Abstract
Memory deficits with aging are related to the neurodegeneration in the brain, including a reduction in arginine vasopressin (AVP) in the brain of patients with Alzheimer’s disease (AD). AVP(4-8), different from its precursor AVP, plays memory enhancement roles in the CNS without peripheral side-effects. However, it is not clear whether AVP(4-8) can improve cognitive behaviors and synaptic plasticity in the APP/PS1 mouse model of AD. Here, we investigated for the first time the neuroprotective effects of AVP(4-8) on memory behaviors and in vivo long-term potentiation (LTP) in APP/PS1-AD mice. The results showed that: (1) APP/PS1-AD mice had lower spontaneous alternation in the Y-maze than wild-type (WT) mice, and this was significantly reversed by AVP(4-8); (2) the prolonged escape latency of APP/PS1-AD mice in the Morris water maze was significantly decreased by AVP(4-8), and the decreased swimming time in target quadrant recovered significantly after AVP(4-8) treatment; (3) in vivo hippocampal LTP induced by high-frequency stimulation had a significant deficit in the AD mice, and this was partly rescued by AVP(4-8); (4) AVP(4-8) significantly up-regulated the expression levels of postsynaptic density 95 (PSD95) and nerve growth factor (NGF) in the hippocampus of AD mice. These results reveal the beneficial effects of AVP(4-8) in APP/PS1-AD mice, showing that the intranasal administration of AVP(4-8) effectively improved the working memory and long-term spatial memory of APP/PS1-AD mice, which may be associated with the elevation of PSD95 and NGF levels in the brain and the maintenance of hippocampal synaptic plasticity.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Alzheimer’s disease (AD) is an insidious degenerative disease of the brain characterized by progressive cognitive deficits, memory loss, and specific neuropsychiatric anomalies [1]. Multiple pathological characteristics of AD have been identified in the brain, including amyloid-beta (Aβ) deposits, tau hyperphosphorylation, neurotrophic factor dysregulation, and synaptic deficits [2,3,4]. Interestingly, a significant decrease in arginine vasopressin (AVP) has been found in the cerebrospinal fluid and many brain regions, especially in the hippocampus [5, 6] of AD patients.
AVP, traditionally associated with the regulation of water balance and blood pressure in the periphery [7], has been considered as a neurotransmitter and/or neuromodulator in the central neural system (CNS) and affects diverse aspects of cognitive ability such as the consolidation and retrieval of memory [8]. Previous research has shown that AVP-deficient rats have poor social discrimination, object discrimination, and conditioned learning [2, 9]; mice with V1a and V1b receptor knockout have deficits in mnemonic function [10,11,12]; and administration of AVP receptor antagonists leads to memory impairment in rats [13]. On the contrary, administration of AVP has been reported to facilitate learning and memory [14,15,16], and enhance synaptic plasticity and the expression of synapse-related protein in the hippocampus [15]. We have reported that AVP rescues Aβ-induced impairments of spatial memory [17], hippocampal long-term potentiation (LTP) [18], and spontaneous discharges [17] in rats.
Unfortunately, the effects of systemic administration of AVP are more complicated [19,20,21] due to the widely distributed receptors and more peripheral side-effects. AVP(4-8), a major metabolic fragment of AVP, differs from its precursor and has specific receptors and distribution in the CNS including the hippocampus [22] and cortex [23]. There is evidence that the hippocampus may be an important target site for the effect of AVP(4-8), and its receptor may be a link between AVP and cognitive behavior [24]. More importantly, AVP(4-8) has a 1000-times more potent effect on memory processes than AVP in a passive avoidance experience [25, 26]. Besides, another advantage of AVP(4-8) in the CNS is that it lacks the peripheral side-effects of its parent hormone AVP, like pressor and/or antidiuretic activity [25, 26].
However, it is not clear whether AVP(4-8) can improve the cognitive behaviors and synaptic plasticity in the APP/PS1 mouse model of AD. We supposed that the decreased AVP, especially its intermediate product AVP(4-8), in the brain may contribute to the cognitive impairment of AD, and exogenous application of AVP(4-8) might be beneficial for improving synaptic plasticity and cognitive behaviors in AD. Therefore, we investigated for the first time the neuroprotective effects of AVP(4-8) on the cognitive behaviors in APP/PS1-AD mice using several behavioral techniques. We also examined memory-related activity (in vivo hippocampal LTP) and proteins [post-synaptic density-95 (PSD95) and nerve growth factor (NGF)] in the hippocampus to clarify the possible electrophysiological and molecular mechanisms.
Material and Methods
Animals and Grouping
Male heterozygous APP/PS1 transgenic mice (APPswe/PS1dE9, Beijing Huafukang Bioscience Co., Inc.) and wild-type (WT) mice (C57BL/6J, Beijing Vital River Laboratory Animal Technology Co., Ltd.) were maintained in an animal room (20 °C ± 2 °C, 12 h light/dark cycle) with enough food and water. All operations on mice were approved by the Ethical Committee of Shanxi Medical University. At 8 months of age, the mice were randomly divided into 4 groups (n = 10–13/group): WT+Saline, WT+AVP(4-8), APP/PS1+Saline, and APP/PS1+AVP(4-8).
Drug Administration
Before behavioral tests, vasopressin metabolite neuropeptide [pGlu4, Cystine6]-AVP(4-8) (2 μg/kg, Phoenix Pharmaceuticals, Inc., USA) or an equivalent volume of saline was administered intranasally three times a day for 4 weeks. The application of AVP(4-8) or saline was continued throughout the behavioral test period. Fig. 1 shows the timeline of the experimental procedures.

Timeline of the experimental procedures. After 1 day of adaptation to the lab setting and 4 weeks of treatment with AVP(4-8) (2 μg/kg) or equivalent saline, the Y-maze test (YMT), Morris water maze (MWM) test, in vivo hippocampal long-term potentiation (LTP) recording, Western blotting, and ELISA were performed sequentially.
Y-Maze Test
After 4 weeks of drug administration (Fig. 1), the Y-maze was first used to measure short-term working memory in mice. This was a three-arm equiangular maze, and the arms were 30 cm long, 7 cm high, and 15 cm wide. Each animal (n = 9/group) was placed in the central triangular region and allowed to travel freely in the maze for 8 min. The Smart 3.0 software system was used to record the number and order of arm entries. A correct spontaneous alternation was identified as a set of three different arm entries. The percentage of correct spontaneous alternation was calculated as (number of correct alternations)/(total arm entries − 2) × 100%. A higher percentage of correct spontaneous alternation indicates better working memory.
Morris Water Maze (MWM) Test
Hippocampus-dependent spatial learning and reference memory were assessed with the MWM test, as performed previously in our lab. The pool (diameter, 120 cm; height, 50 cm) filled with opaque water (temperature 20 °C ± 2 °C), was conceptually divided into four equal quadrants with the escape platform submerged in the center of the first quadrant (target quadrant) (1 cm below the water surface). Different-shaped black marks were displayed at the perimeter of each quadrant. During the acquisition phase, mice (n = 9–10/group) were trained in 4 sessions (60 s per session) per day over 5 consecutive days in searching for the underwater platform. The escape latency and swimming trials were recorded by a video-tracking system (Ethovision 3.0, Noldus Information Technology, Wageningen, Netherlands). In probe trials (day 6), mice were allowed to swim freely for 1 min without the platform. The swimming time in the target quadrant and the swimming speed were recorded. After that, a visible platform test was performed to assess the visual and motor ability of mice by recording the time when they arrived at the escape platform.
In Vivo Hippocampal LTP Recording
Hippocampal LTP is a synaptic model of memory with the closest connection to long-term memory due to its long duration. Therefore, field excitatory postsynaptic potentials (fEPSPs) in the hippocampal CA1 region were recorded after the MWM test was finished. Mice were anesthetized with chloral hydrate (5%, i.p., 0.08 mL/kg) and placed in a stereotaxic apparatus (RWD Life Science, Shenzhen, China). Then, a hole (2.0 mm posterior to bregma and 1.5 mm from midline) was drilled through the skull for a self-made bound stimulating/recording electrode (FHC, USA) inserted into the Schaffer collateral/CA1 region and recording fEPSPs. The distance between the tips of the recording and stimulating electrodes was 0.5 mm horizontally, with a 0.3 mm of vertical difference. The tips of the stimulating and recording electrodes were located in the Schaffer collateral/commissural pathway and the stratum radiatum of the hippocampal CA1 region. The orientation of the tips was determined by electrophysiological criteria (appearance of an evoked fEPSP) and final histological verification. Test stimuli from Master-9 stimulator (AMPI, Jerusalem, Israel) were delivered to the Schaffer-collateral/commissural pathway at 0.033 Hz and an intensity that induced 40%–50% of the maximum fEPSP response. Baseline synaptic transmission was recorded for 30 min to ensure stability. Paired pulse facilitation (PPF) was initiated by paired stimuli at an interval of 50 ms, and calculated as fEPSP2/fEPSP1. Then, LTP was induced by high-frequency stimulation (HFS) (three trains of 20 pulses at 200 Hz with an interval of 30 s) at an intensity that evoked 80% of the maximum fEPSP. After that, fEPSPs were evoked and recorded again for at least 60 min with the same intensity as the test stimuli. The voltage signals were acquired at 40 kHz and filtered with a 1-kHz cutoff frequency and a 1-s time constant. The slopes of the raw fEPSP traces were obtained automatically by the system software (RM-6240 Biological Signal Collecting and Processing System, Chengdu Instruments Ltd, China), and every four fEPSPs were averaged.
Western Blotting
Mice (n = 6/group) were sacrificed to obtain whole brains and the hippocampus was quickly dissected on ice to determine the level of PSD95. All samples were snap-frozen in liquid nitrogen and stored at – 80 °C until use. The hippocampal tissue was homogenized in the tissue protein extraction reagent and phenylmethylsulfonyl fluoride (PMSF). After centrifugation (13,000 rpm, 15 min, 4 °C), protein in the supernatant was quantified using a BCA Protein Assay Kit. Each sample (20 μg) was separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis for detecting β-actin, PSD95, and NGF. Separated proteins were then transferred onto polyvinylidene difluoride membranes, which were blocked with 5% bovine serum albumin (BSA) for 2 h and then incubated with primary rabbit antibodies against β-actin (1:5,000, ZSGB-BIO, Beijing), PSD95 (1:500, Abcam), or mouse antibody against NGF (1:200, Santa Cruz Biotechnology) in TBST containing 5% BSA at 4 °C overnight. After washing with TBST 3 times for 15 min, 10 min, and 5 min, the blots were incubated with horseradish peroxidase-conjugated goat anti-rabbit IgG (1:5,000, Abcam) and goat anti-mouse IgG (1:5,000, Abcam) for 2 h at 4 °C. After washing as above, the blots were assessed by an ECL assay system. Densitometric analysis of the Western blots was performed using Alpha View software.
Enzyme-Linked Immunosorbent Assay (ELISA)
To further verify the result of hippocampal NGF measurement in Western blots, we used a pre-coated Mouse NGF/NGF Beta ELISA Kit (Boster Biological Technology, Wuhan, China. Catalog #EK0470) (n = 4–5/group) to measure the NGF content in the hippocampus according to the manufacturer’s instructions. The absorbance was measured at 450 nm using an Enspire™ multilabel reader 2300 (Perkin Elmer, Turku, Finland).
Statistical Analysis
All data are reported as the mean ± SEM, and analyzed with SPSS 16.0 and SigmaPlot 12.3 statistical packages. Three-way repeated measure analysis of variance (ANOVA) with time, genotype, and treatment as within-subject factors was used to analyze the escape latency in MWM acquisition and the fEPSP slopes from in vivo hippocampal LTP. The other data were analyzed with two-way ANOVA followed by Tukey’s post hoc test. Statistical significance was considered to be P < 0.05.
Results
AVP(4-8) Improves Working Memory of Both WT and APP/PS1 Mice in the Y-Maze Test
The total arm entries did not show any significant difference among the four groups (P > 0.05), which revealed that genotype and AVP(4-8) had no influence on the locomotor activity and motor function (Fig. 2A). In contrast, a significantly less spontaneous alternation was seen in the APP/PS1+Saline group (59.67% ± 4.98%) than in the WT+Saline group (76.56% ± 2.01%, P < 0.001). AVP(4-8) treatment caused significantly more spontaneous alternation in the WT+AVP(4-8) (81.44% ± 1.16%, P < 0.05) and APP/PS1+AVP(4-8) groups (71.78% ± 1.14%, P < 0.001) than in their respective saline control groups (Fig. 2B; two-way ANOVA: genotype: F(1,32) = 74.449, P < 0.001; AVP(4-8): F(1,32) = 30.510, P < 0.001; genotype × AVP(4-8): F(1,32) = 5.507, P < 0.05), indicating a working memory enhancement in normal mice by AVP(4-8), an impairment in APP/PS1 mice, and its reversal by AVP(4-8).

AVP(4-8) improved the working memory of APP/PS1 mice in the Y-maze test. A Total arm entries during 8 min, with no significant difference between groups (n = 9/group). B Percentage of correct spontaneous alternation, with a significant decrease in the APP/PS1 mice compared with controls, and a significant reversal after AVP(4-8) treatment (*P < 0.05, ***P < 0.001).
AVP(4-8) Rescued Spatial Learning and Reference Memory Impairments of APP/PS1 Mice in MWM
The MWM test was used to assess the spatial learning and memory of mice. The three-way repeated measure ANOVA showed no interaction among the three within-subject factors (time × genotype × treatment: F(4,32) = 1.015, P > 0.05), but genotype and treatment had significant main effects and an interaction on the escape latency (genotype: F(1,8) = 112.694, P < 0.001; treatment: F(1,8) = 5.755, P < 0.05; genotype × treatment: F(1,8) = 448.594, P < 0.05). Throughout the acquisition session there was an overall gradual decrease in escape latency (day: F(4, 32) = 45.195, P < 0.001; Table 1, Fig. 3A). What is more, the APP/PS1+Saline group spent more time on finding the hidden platform than the WT+Saline group on days 3–5 (P < 0.001), while treatment with AVP(4-8) decreased the escape latency in the APP/PS1+AVP(4-8) group on days 4–5 (P < 0.05). Representative swimming traces of mice on navigation training day 5 are shown in Fig. 3B. This result indicated a spatial learning impairment in APP/PS1 mice, which was partly rescued by AVP(4-8) treatment. In the probe test (Fig. 3C, D), a reference memory deficit also occurred in the APP/PS1+Saline group, with a lower percentage of swimming time in the target quadrant than its WT control (P < 0.001). After treatment with AVP(4-8), the APP/PS1+AVP(4-8) group showed a significantly higher percentage of swimming time (P < 0.05) than the APP/PS1+Saline group (two-way ANOVA: genotype: F(1,35) = 31.43, P < 0.001; AVP(4-8): F(1,35) = 6.62, P < 0.05; genotype × AVP(4-8): F(1,35) = 0.586, P = 0.450). Meantime, all mice in the four groups showed similar swimming speeds in the probe test (Fig. 3E) and similar swimming times to the visible platform (Fig. 3F). These results suggested that the differences in spatial learning and memory did not result from alterations of motor ability and visual acuity.

AVP(4-8) rescued the spatial learning and memory of APP/PS1 mice in MWM tests. A Average escape latencies of mice in the place navigation test during 5 training days (***P < 0.001 vs WT+Saline mice, #P < 0.05 vs APP/PS1+Saline mice). B Representative swimming traces of mice on navigation training day 5. C Percentages of swimming time spent in the target quadrant in the spatial probe test (*P < 0.05, ***P < 0.001). D Representative swimming traces in the probe test. E, F Average swimming speed in the probe test (E) and swimming time to the visible platform (F) (n = 9).
AVP(4-8) Partly Reversed the Suppression of Hippocampal Synaptic Plasticity in APP/PS1 Mice
The hippocampal fEPSP slope differed before and after HFS (Fig. 4A, B). During 30 min of fEPSP recording before HFS, the basic synaptic transmission was stable in each group. After HFS, LTP was induced successfully with a rapid increase in the fEPSP slope and then gradual attenuation with time in the four groups (time: F(44, 176) = 293.464, P < 0.001). At 30 and 60 min after HFS, the fEPSP slope in the APP/PS1+Saline group was significantly lower (n = 6; 30 min: 139.09% ± 2.69%, P < 0.01; 60 min: 116.69% ± 5.09%, P < 0.001) than in the WT+Saline group (n = 8; 30 min: 176.19% ± 8.86%; 60 min: 166.72% ± 3.68%). After treatment with AVP(4-8), the fEPSP slopes in the APP/PS1+AVP(4-8) group significantly increased compared to that in the APP/PS1+Saline group (n = 7; 30 min: 157.19% ± 3.52%, P < 0.05; 60 min: 145.06% ± 3.38%, P < 0.001) (genotype: F(1,24) = 23.604, P < 0.01, AVP(4-8): F(1,24) = 4.204, P < 0.05; genotype × AVP(4-8): F(1,24) = 0.507, P < 0.05). Representative fEPSP traces before and 60 min after HFS in different groups are shown in Fig. 4C. Besides, we did not find any significant difference in the PPF ratio among the four groups. These results demonstrated that AVP(4-8) partly rescued the LTP maintenance in APP/PS1 mice, and this could involve a postsynaptic mechanism.

AVP(4-8) partly reversed the suppression of synaptic plasticity in the hippocampus of APP/PS1 mice. A Changes in the slope of fEPSPs before and after high-frequency stimulation (HFS) in the CA1 region. B fEPSP slopes before HFS, and 1, 30, and 60 min after HFS in the four groups (*P < 0.05, **P < 0.01, ***P < 0.001). C Representative fEPSP traces before (black) and 60 min after (red) HFS in the different groups. D Paired-pulse facilitation (PPF) in the different groups. Inset: a sample PPF trace.
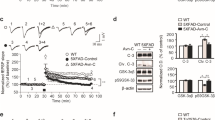
AVP(4-8) Up-Regulated the Levels of PSD95 and NGF in the Hippocampus of APP/PS1 Mice
To further clarify the molecular mechanisms underlying the improving effects of AVP(4-8) on behaviors and LTP in APP/PS1 mice, the expression levels of PSD95 and NGF were measured using Western blot (n = 6/group). The data (Fig. 5A) showed that the level of PSD95 in the APP/PS1+Saline group was significantly lower (P < 0.001) than in the WT+Saline group, while AVP(4-8) application significantly up-regulated the level of PSD95 in the APP/PS1+AVP(4-8) group (P < 0.001). Two-way ANOVA showed significant main effects and an interaction (genotype: F(1,23) = 128.605, P < 0.001, AVP(4-8): F(1,23) = 32.890, P < 0.001; genotype × AVP(4-8): F(1,23) = 7.319, P < 0.05). In addition, the NGF data (Fig. 5B) showed a distinct increase in the NGF level in the WT+AVP(4-8) (P < 0.05) and APP/PS1+AVP(4-8) (P < 0.05) groups compared to their respective saline controls (two-way ANOVA: genotype: F(1,17) = 2.937, P = 0.109; AVP(4-8): F(1,17) = 12.116, P < 0.01; genotype × AVP(4-8): F(1,17) = 0.117, P = 0.738), but without any significant difference between the WT+Saline and APP/PS1+Saline groups. Similarly, the result of ELISA measurement of NGF (data not shown) was in accord with that from the Western blots. These together indicated that the neuroprotection of AVP(4-8) on the cognitive behavior and synaptic plasticity in APP/PS1 mice is closely associated with the up-regulation of PSD95 and NGF levels in the hippocampus.

AVP(4-8) up-regulated the levels of PSD95 and NGF in the hippocampus of APP/PS1 mice. A Representative Western blots of PSD95 (upper) and their quantitative plots (lower) in the different groups, showing that the decreased level of PSD95 in the hippocampus of APP/PS1 mice was partially reversed in the APP/PS1+AVP(4-8) group. B Representative Western blots of NGF (upper) and corresponding quantitative plots (lower), showing significantly increased levels of NGF in both the AVP(4-8)-treated groups (***P < 0.001).
Discussion
AD brings about a number of neurophysiological and neurochemical dysfunctions, like decreased AVP in the rat hippocampus [6], which partly accounts for the impairment of learning and memory. AVP exerts complicated effects in the periphery and CNS, such as a classical neuroendocrine role, and neurotransmitter or neuromodulator functions. Interestingly, the central behavioral effects of AVP are dissociated from its traditional peripheral effects [26]. AVP(4-8), a more potent peptide than its parent molecule, is considered to be a mediator of the central behavioral effects of AVP [24]. Moreover, it has been established that there are specific receptors for AVP(4-8) in regions such as the hippocampus, cerebral cortex, and amygdaloid nucleus [22]. These regions are closely associated with learning and memory. Thus, AVP(4-8) receptors have been thought to link AVP with cognitive behaviors.
In the present study, we investigated for the first time the effects of AVP(4-8) on learning and memory in the APP/PS1 mouse model of AD. Because the hippocampus is essential for place learning and cognitive mapping, we first used the Y maze and MWM to study spatial reference memory. The behavioral tests showed that intranasal administration of AVP(4-8) for > 4 weeks improved the deficits in learning and memory of APP/PS1 mice. In the Y-maze test, AVP(4-8) increased the percentage of correct spontaneous alternation in both WT and APP/PS1 mice. This suggested that AVP(4-8) not only facilitated working memory in WT mice but also rescued the working memory impairment due to the gene mutation in APP/PS1 mice. Afterwards, the classical MWM test showed that the APP/PS1+AVP(4-8) group had a decreased escape latency in the acquisition phase and an increased swimming time in the target quadrant compared with the APP/PS1+saline group. These results indicated that AVP(4-8) partly improved the long-term spatial learning and memory in APP/PS1 mice. We noted that the escape latency of the APP/PS1+AVP(4-8) group on training day 3 had an unexpected increase over that on training day 2. We suppose this resulted from internal or external influences. The acquisition phase involves many complex memory processes, including collecting visual information about spatial location, and processing and remembering it to find the hidden platform. In the AD model treatment group, the drug effect might be another influencing factor. So, the escape latency is vulnerable to many internal or external factors. Maybe environmental disturbances on training day 3 affected the learning behaviors.
The mechanisms underlying the neuroprotective effects of AVP(4-8) are uncertain. It is known that hippocampal synaptic failure is the best correlate of cognitive decline in AD patients and in animal models of the disease. Hippocampal LTP, defined as long-lasting modifications of synaptic potentiation, is believed to be the basis of memory processes in the mammalian brain [27]. Thus, the improvement of cognitive behavior in APP/PS1 mice might be due to the recovery of hippocampal synaptic function [28]. Similar to our in vivo results for PPF and LTP, a recent in vitro brain slice study showed that APP/PS1 mice (8–10 months old) exhibit reduced basal synaptic transmission and LTP in the CA1 area of the hippocampus [29]. It has been reported that AVP and AVP(4-8) potentiate the excitatory postsynaptic potential and elicit the LTP of synaptic transmission of CA1/subiculum neurons in rat hippocampal slices [30]; AVP potentiates or rescues LTP in the hippocampal DG region of rats [18, 31, 32]. The present study further showed that AVP(4-8) treatment effectively rescued the hippocampal LTP in the CA1 region of APP/PS1 mice, adding to the research on AVP(4-8). The electrophysiological results were also consistent with and supported our findings in the cognitive behavioral tests, further clarifying the relevance of hippocampal LTP to spatial learning and memory.
Similar to our results, Gelman et al. also found that the reduction of basal synaptic transmission and LTP in the CA1 area is accompanied by invariant PPF [29]. This suggests that the impairments in basal synaptic transmission and LTP may result from a postsynaptic mechanism. It has been reported that hippocampal mutant APP and Aβ significantly reduce the level of PSD [33]. Moreover, AD patients with mild cognitive impairment already demonstrate a decline of PSD95 [34]. Thus, a dysfunction of PSD95 may be the beginning of the process of synaptic damage prior to synaptic loss in the pathology of AD [35]. PSD95 is closely associated with synaptic plasticity because the neuronal scaffolding protein is associated with NMDA- and AMPA-type glutamate receptors and their downstream signaling molecules [36]. In accordance with previous studies, our Western blot results showed that the expression level of PSD95 in the hippocampus of APP/PS1 mice was markedly decreased. Interestingly, AVP(4-8) administration significantly up-regulated the level in APP/PS1 mice, which might partly account for the improvement of behavior and the rescue of LTP.
Previous studies [37] have confirmed the molecular basis of AVP(4-8) in cognitive protection, including: binding to and stimulating G protein-coupled receptors [38, 39], accumulating inositol 1,4,5-triphosphate, activating Ca2+/CaM-dependent kinase II [40] and protein kinase C [41], facilitating the phosphorylation of growth-associated protein and mitogen-activated protein kinase [42], enhancing the gene expression of NGF [43, 44] and brain-derived neurotrophic factor [45], and finally, strengthening synaptic plasticity and LTP [46]. In accordance with the above, our experimental results from Western blotting and ELISA showed that the expression level of NGF was greatly up-regulated in both WT and APP/PS1 mice by AVP(4-8). NGF is a neurotrophin particularly necessary for the survival, differentiation, maintenance and plasticity of forebrain cholinergic neurons. The degeneration of these neurons has been thought to be responsible for cognitive impairments in AD patients [47, 48]. Early studies reported that anti-NGF antibody impairs cognitive behavior in rats, while NGF synthesis stimulators restore it [49]. The function of NGF has also been demonstrated in AD11 mice. These mice express recombinant anti-NGF monoclonal antibodies and thus show marked neuronal degeneration, memory impairments, and Aβ accumulation in the cortex and hippocampus, while these deficits can be rescued by exogenous NGF [50, 51]. Besides, NGF stimulates nerve fiber growth in cultured spinal ganglia and increases the cell body size of septal neurons [47]. Likewise, AVP(4-8) also affects nerve growth in cultured hippocampal neurons [52]. Together, these findings imply that increased NGF content in the hippocampus plays an important role in improving learning and memory in APP/PS1 mice. It is interesting that AVP(4-8) enhanced working memory but not spatial memory in WT mice while the NGF level was dramatically increased. This probably involves the time-dependence of memory development. Correct alternation in the Y maze reflects working memory, which is a component of short-term memory. On the other hand, spatial memory in the probe test of the MWM represents long-term memory. The establishment of long-term memory needs more and longer associative learning. Four weeks of treatment with AVP(4-8) in the present study may be enough for working memory, but not enough for the long-term spatial memory in WT mice. In fact, the WT mice still expressed an increasing trend (although not significant) in spatial memory induced by AVP(4-8).
In addition, the present study also indicated that intranasal administration is a convenient and effective drug-delivery approach for central effects. De Wied et al. showed that central administration (intracerebroventricular; icv) of AVP(4-8) is 2,300 times more potent than peripheral administration (subcutaneous) [16, 25]. Because icv injection is impractical for patients, nasal administration may be the only drug-delivery route for easy entry into the brain. By bypassing liver metabolism and the blood-brain barrier, the half-life of AVP(4-8) in the brain may be much longer and the final concentration in the brain should be much higher. Meantime, intranasal application of AVP(4-8) also avoids peripheral side-effects.
In conclusion, our study reveals for the first time the neuroprotective effects of AVP(4-8) in APP/PS1-AD mice. These results indicate that intranasal administration of AVP(4-8) effectively improves working memory and long-term spatial memory in APP/PS1-AD mice, and this is associated with the elevation of PSD95 and NGF levels in the brain and the maintenance of hippocampal synaptic plasticity.
References
Wuwongse S, Chang RC, Law AC. The putative neurodegenerative links between depression and Alzheimer’s disease. Prog Neurobiol 2010, 91: 362–375.
Varga J, Klausz B, Kálmán ÁD, Pákáski M, Szucs S, Garab D, et al. Increase in Alzheimer’s related markers preceeds memory disturbances: Studies in vasopressin-deficient Brattleboro rat. Brain Res Bull 2014, 100: 6–13.
Strac DS, Muck-Seler D, Pivac N. Neurotransmitter measures in the cerebrospinal fluid of patients with Alzheimer’s disease: a review. Psychiatr Danub 2015, 27: 14–24.
Sun BL, Li WW, Zhu C, Jin WS, Zeng F, Liu YH, et al. Clinical research on Alzheimer’s disease: progress and perspectives. Neurosci Bull 2018, 34: 1111–1118.
Raskind MA, Peskind ER, Lampe TH, Risse SC, Taborsky GJ, Dorsa D. Cerebrospinal fluid vasopressin, oxytocin, somatostatin, and beta-endorphin in Alzheimer’s disease. Arch Gen Psychiatry 1986, 43: 382–388.
Mazurek MF, Bed MF, Bird ED, Martin JB. Vasopressin in Alzheimer’s disease: a study of postmortem brain concentrations. Ann Neurol 1986, 20: 665–670.
Rotondo F, Butz H, Syro LV, Yousef GM, Di Ieva A, Restrepo LM. Arginine vasopressin (AVP): a review of its historical perspectives, current research and multifunctional role in the hypothalamohypophysial system. Pituitary 2016, 19: 345–355.
van Wimersma Greidanus TB, Bohus B, de Wied D. The role of vasopressin in memory processes. Prog Brain Res 1975, 42: 135–141.
Engelmann M, Landgraf R. Microdialysis administration of vasopressin into the septum improves social recognition in Brattleboro rats. Physiol Behav1994, 55: 145–149.
Egashira N, Tanoue A, Higashihara F, Mishima K, Fukue Y, Takano Y, et al. V1a receptor knockout mice exhibit impairment of spatial memory in an eight-arm radial maze. Neurosci Lett 2004, 356: 195–198.
DeVito LM, Konigsberg R, Lykken C, Sauvage M, Young WS, 3rd, Eichenbaum H. Vasopressin 1b receptor knock-out impairs memory for temporal order. J Neurosci 2009, 29: 2676–2683.
Bielsky IF, Hu SB, Szegda KL, Westphal H, Young LJ. Profound impairment in social recognition and reduction in anxiety-like behavior in vasopressin V1a receptor knockout mice. Neuropsychopharmacology 2004, 29: 483–493.
Nephew BC, Bridges RS. Arginine vasopressin V1a receptor antagonist impairs maternal memory in rats. Physiol Behav 2008, 95: 182–186.
Weingartner H, Gold P, Ballenger JC, Smallberg SA, Summers R, Rubinow DR, et al. Effects of vasopressin on human memory functions. Science 1981, 211: 601–603.
Yang C, Zhang X, Gao J, Wang M, Yang Z. Arginine vasopressin ameliorates spatial learning impairments in chronic cerebral hypoperfusion via V1a receptor and autophagy signaling partially. Transl Psychiatry 2017, 7: e1174.
de Wied D, Gaffori O, van Ree JM, de Jong W. Central target for the behavioural effects of vasopressin neuropeptides. Nature 1984, 308: 276–278.
Pan YF, Jia XT, Wang XH, Chen XR, Li QS, Gao XP, et al. Arginine vasopressin remolds the spontaneous discharges disturbed by amyloid beta protein in hippocampal CA1 region of rats. Regul Pept 2013, 183: 7–12.
Jing W, Guo F, Cheng L, Zhang JF, Qi JS. Arginine vasopressin prevents amyloid beta protein-induced impairment of long-term potentiation in rat hippocampus in vivo. Neurosci Lett 2009, 450: 306–310.
Hicks C, Ramos L, Reekie T, Misagh GH, Narlawar R, Kassiou M, et al. Body temperature and cardiac changes induced by peripherally administered oxytocin, vasopressin and the non-peptide oxytocin receptor agonist WAY 267,464: a biotelemetry study in rats. Br J Pharmacol 2014, 171: 2868–2887.
Song Z, Albers HE. Cross-talk among oxytocin and arginine-vasopressin receptors: Relevance for basic and clinical studies of the brain and periphery. Front Neuroendocrinol 2018, 51: 14–24.
Alescio-Lautier B, Soumireu-Mourat B. Effects of peripherally administered arginine-vasopressin on learning, retention and forgetting in mice. Behav Brain Res 1990, 41: 117–128.
Wu JH, Du YC. Binding sites of ZNC(C)PR, a pentapeptide fragment of argipressin, in rat brain. Acta Pharmacol Sin 1995, 16: 141–144.
Du YC, Wu JH, Jiang XM, Gu YJ. Characterization of binding sites of a memoryenhancing peptide AVP(4-8) in rat cortical synaptosomal membranes. Peptides 1994, 15: 1273–1279.
Reijmers LGJE, van Ree JM, Spraijt BM, Burbach JP, De Wied D. Vasopressin metabolites: A link between vasopressin and memory? Prog Brain Res 1999, 119: 523–535.
Burbach JP, Kovacs GL, de Wied D, van Nispen JW, Greven HM. A major metabolite of arginine vasopressin in the brain is a highly potent neuropeptide. Science 1983, 221: 1310–1312.
De Wied D, Gaffori O, Van Ree JM, De Jong W. Vasopressin antagonists block peripheral as well as central vasopressin receptors. Pharmacol Biochem Behav 1984, 21: 393–400.
Peineau S, Rabiant K, Pierrefiche O, Potier B. Synaptic plasticity modulation by circulating peptides and metaplasticity: Involvement in Alzheimer’s disease. Pharmacol Res 2018, 130: 385–401.
Ni B, Wu R, Yu T, Zhu H, Li Y, Liu Z. Role of the hippocampus in distinct memory traces: timing of match and mismatch enhancement revealed by intracranial recording. Neurosci Bull 2017, 33: 664–674.
Gelman S, Palma J, Tombaugh G, Ghavami A. Differences in synaptic dysfunction between rTg4510 and APP/PS1 mouse models of Alzheimer’s disease. J Alzheimers Dis 2018, 61: 195–208.
Chepkova AN, French P, De Wied D, Ontskul AH, Ramakers GM, Skrebitski VG, et al. Long-lasting enhancement of synaptic excitability of CA1/subiculum neurons of the rat ventral hippocampus by vasopressin and vasopressin(4-8). Brain Res 1995, 701: 255–266.
Dubrovsky B, Tatarinov A, Gijsbers K, Harris J, Tsiodras A. Effects of arginine-vasopressin (AVP) on long-term potentiation in intact anesthetized rats. Brain Res Bull 2003, 59: 467–472.
Wang M, Chen JT, Ruan DY, Xu YZ. Vasopressin reverses aluminum-induced impairment of synaptic plasticity in the rat dentate gyrus in vivo. Brain Res 2001, 899: 193–200.
Reddy PH, Yin X, Manczak M, Kumar S, Pradeepkiran JA, Vijayan M, et al. Mutant APP and amyloid beta-induced defective autophagy, mitophagy, mitochondrial structural and functional changes and synaptic damage in hippocampal neurons from Alzheimer’s disease. Hum Mol Genet 2018, 27: 2502–2516.
Mirza FJ, Zahid S. The role of synapsins in neurological disorders. Neurosci Bull 2018, 34: 349–358.
Pham E, Crews L, Ubhi K, Hansen L, Adame A, Cartier A, et al. Progressive accumulation of amyloid-beta oligomers in Alzheimer’s disease and in amyloid precursor protein transgenic mice is accompanied by selective alterations in synaptic scaffold proteins. FEBS J 2010, 277: 3051–3067.
Shinohara Y. Quantification of postsynaptic density proteins: glutamate receptor subunits and scaffolding proteins. Hippocampus 2012, 22: 942–953.
Du YC, Yan QW, Qiao LY. Function and molecular basis of action of vasopressin 4-8 and its analogues in rat brain. Prog Brain Res 1998, 119: 163–175.
Qiao LY, Du YC. Involvement of a putative G-protein-coupled receptor and a branching pathway in argipressin (4-8) signal transduction in rat hippocampus. Acta Pharmacol Sin 1998, 19: 15–20.
Yan QW, Du YC. AVP(4-8) may stimulate a G protein-coupled receptor in rat hippocampal synaptosomal membranes. Acta Biochim Biophys Sin 1998, 30: 505–509.
Qiao LY, Chen XF, Gu BX, Wang TX, Du YC. Effect of AVP(4-8) administration on Ca2+/CaM-dependent protein kinase II autophosphorylation in rat brain. Acta Physiol Sin 1998, 50: 132–138.
Zhen X, Dong M, Du YC. Effect of arginine-vasopressin(4-8) on PKC and PKA activities in rat brain. Chin J Biochem Mol Biol 2000, 16: 529–532.
Zhen XG, Du YC. AVP(4-8) enhances PKC and MAPK activities in SK-N-SH cells. Acta Biochim Biophys Sin 2000, 32: 105–108.
Guo J, Zhou AW, Du YC, Chen XF. AVP(4-8) increases NGF mRNA and protein content in rat hippocampus. Chin J Neurosci 1996, 3: 23–27.
Zhou AW, Guo J, Wang HY, Gu BX, Du YC. Enhancement of NGF gene expression in rat brain by the memory-enhancing peptide AVP(4-8). Peptides 1995, 16: 581–586.
Zhou AW, Li WX, Guo J, Du YC. Facilitation of AVP(4-8) on gene expression of BDNF and NGF in rat brain. Peptides 1997, 18: 1179–1187.
Chen XF, Tang T, Zhang JW, Miao HH, Wang TX, Du YC. ZNC(C)PR affects developmental changes of P46 phosphorylation in rat hippocampus. Mol Reprod Dev 1993, 35: 251–256.
Isaev NK, Stelmashook EV, Genrikhs EE. Role of nerve growth factor in plasticity of forebrain cholinergic neurons. Biochemistry (Mosc) 2017, 82: 291–300.
Tiernan CT, Ginsberg SD, He B, Ward SM, Guillozet-Bongaarts AL, Kanaan NM, et al. Pretangle pathology within cholinergic nucleus basalis neurons coincides with neurotrophic and neurotransmitter receptor gene dysregulation during the progression of Alzheimer’s disease. Neurobiol Dis 2018, 117: 125–136.
Yamada K, Nitta A, Hasegawa T, Fuji K, Hiramatsu M, Kameyama T, et al. Orally active NGF synthesis stimulators: potential therapeutic agents in Alzheimer’s disease. Behav Brain Res 1997, 83: 117–122.
Berardi N, Braschi C, Capsoni S, Cattaneo A, Maffei L. Environmental enrichment delays the onset of memory deficits and reduces neuropathological hallmarks in a mouse model of Alzheimer-like neurodegeneration. J Alzheimers Dis 2007, 11: 359–370.
De Rosa R, Garcia AA, Braschi C, Capsoni S, Maffei L, Berardi N, et al. Intranasal administration of nerve growth factor (NGF) rescues recognition memory deficits in AD11 anti-NGF transgenic mice. Proc Natl Acad Sci U S A 2005, 102: 3811–3816.
Tarumi T, Sugimoto Y, Chen Z, Zhao Q, Kamei C. Effects of metabolic fragments of [Arg(8)]-vasopressin on nerve growth in cultured hippocampal neurons. Brain Res Bull 2000, 51: 407–411.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (31471080), the Scientific Program for “Sanjin Scholars” of Shanxi Province, Shanxi “1331 Project” Key Subjects Construction (1331KSC), and Science Foundation for Excellent Young Scholars of Shanxi Province, China (201801D211005).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors claim no conflict of interest.
Rights and permissions
About this article

Cite this article
Zhang, X., Zhao, F., Wang, C. et al. AVP(4-8) Improves Cognitive Behaviors and Hippocampal Synaptic Plasticity in the APP/PS1 Mouse Model of Alzheimer’s Disease. Neurosci. Bull. 36, 254–262 (2020). https://doi.org/10.1007/s12264-019-00434-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12264-019-00434-0