
Abstract
Dissolved oxygen dynamics in estuarine and coastal environments are complex and highly variable, which highlight the need to compile information from many different types of estuaries. Small estuaries where euhaline habitats dominate are particularly ill represented in the literature. As a contribution to fill this gap, a study on dissolved oxygen dynamics was conducted in the small estuary of Urdaibai (inner Bay of Biscay). Spatial and temporal variations in the percentage saturation of dissolved oxygen (DO-saturation) along the salinity gradient of the estuary and the role of hydro-climatic factors and eutrophication as drivers of those variations at seasonal and inter-annual time scales were analyzed (period 1998–2008). DO-saturation showed an inner to outer estuary increasing gradient. DO-saturation below the salinity gradient layer was either significantly higher than or not significantly different from that in the salinity gradient layer. DO-saturation showed summer minima, but hypoxia was rarely observed. At the outermost estuary, seasonal variations of DO were small and it is hypothesized that DO dynamics were governed mainly by tidal exchange and turbulence. In the intermediate and inner estuary, seasonal variations of DO-saturation were best explained by river discharge, and to a lesser extent by chlorophyll a and temperature, each factor gaining relevance in different periods of the year. In intermediate and inner zones, river discharge exerted a positive effect on DO-saturation, likely via an increase in the renewal rate of DO. At the inter-annual time scale, unlike at the seasonal scale, temperature did not show a significant negative relationship with DO-saturation.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Dissolved oxygen (DO) is a useful metric for aquatic ecosystem health because of its influence in a number of biogeochemical processes that affect the well-being of organisms (Moore et al. 2009; Brown and Power 2011). In the last decades, the number of hypoxic (<2 mg l−1 of DO or approximately 30 % saturation, see Rabalais et al. 2010) sites and the extension and intensity of coastal and estuarine hypoxia appear to be on the rise (Diaz and Rosenberg 2008; Vaquer-Sunyer and Duarte 2008; Gilbert et al. 2010), and DO depletion has become one of the most serious environmental problems of these ecosystems worldwide (Rabalais et al. 2002; Hagy et al. 2004; Chen et al. 2007; Vaquer-Sunyer and Duarte 2008; Meier et al. 2011). Spatiotemporal variations in DO in estuarine and coastal systems are the result of complex interactions of physical and biogeochemical factors, such as nutrient loadings that fuel photosynthetic production of organic matter, as well as direct loadings of reduced substrates that promote oxygen demand, and hydrodynamic and climatic factors like water temperature, river discharge, winds, tidal currents, upwelling processes, vertical stratification, and estuarine circulation (see Kemp et al. 2009; Meire et al. 2013). In some aquatic systems, low DO levels are naturally formed (Helly and Levin 2004; Hagy and Murrell 2007). However, since the mid-twentieth century eutrophication and global climate change, which are two major environmental changes with strong potential to negatively affect coastal and estuarine DO levels, are accelerating (Meire et al. 2013). Indeed, the general decline in DO observed in coastal and estuarine ecosystems in the past decades has been largely attributed to anthropogenic activities which have enhanced nutrient and organic matter loadings (Diaz 2001; Conley et al. 2009). As a consequence, management plans conducive to reductions in nutrient loadings and improvement of sewage treatment have been implemented around many coastal and estuarine areas, which have caused DO levels to increase in some cases (Villate et al. 2013), but not in others (Hagy et al. 2004; Conley et al. 2009), because the linkage between nutrient loads and DO is highly nonlinear (Jonasson et al. 2012). Recent work is showing that in some coastal and estuarine systems climate factors are now emerging as more influential in the decline of DO (Voynova and Sharp 2012; Capet et al. 2013). This highlights the need to investigate how eutrophication and climate change are affecting DO dynamics in coastal and estuarine waters at present and how they can affect them in the future (Meier et al. 2011; Meire et al. 2013). Furthermore, in the case of climate forcing, there is also much uncertainty about whether temperature or rainfall (and related hydrodynamic factors) will be the dominant climatic driver of DO variations (Meire et al. 2013). A further complication arises from the enormous variability across and within estuaries and coasts in the geomorphologic and hydrodynamic features, as well as in the degree and type of anthropogenic disturbance (McLusky and Elliot 2004). These differences are responsible for the high variability across and within estuaries and coastal systems in the way in which different potential drivers of DO dynamics interplay and, therefore, in their dominance (Codiga et al. 2009; Kemp et al. 2009; Villate et al. 2013). As a consequence, a high place-based variability in DO patterns at different spatial and temporal scales can be found (Kemp et al. 2009). In addition to the inter-annual trend of variation, the seasonality of DO depletion is also important from a management point of view. DO depletion can be episodic, seasonal (mostly during spring-summer months), or permanent, depending on estuarine characteristics and the mode of interplay of the different DO drivers (see Kemp et al. 2009). Considering this high variability, compilation of place-based information from many different estuarine and coastal areas is now necessary in order to obtain a general picture of the combined effect of eutrophication and climate change on DO, both at seasonal, but particularly at inter-annual time scales. To this purpose in our research group, the effect of hydro-climatic and anthropogenic factors on DO variability in Basque coast estuaries (southeastern Bay of Biscay) is being investigated (Iriarte et al. 2010; Villate et al. 2013). Results for the polluted estuary of Bilbao revealed significant spatial differences in the percentage saturation of dissolved oxygen (DO-saturation) trends and drivers, related to site-specific differences in the intensity of stratification and flushing (Villate et al. 2013). In the salinity gradient layer of the intermediate estuary, a great improvement in DO-saturation (40 to 80 %) was observed from 1998 to 2008 driven mainly by sewage pollution abatement. However, no parallel recovery took place in bottom waters of the inner estuary, where seasonal hypoxia continues to be a consistent feature. Here, hydrodynamic factors related to river discharge were the factors that best explained inter-annual variations in DO. In contrast, in the outer less stratified, more open estuary, normoxic conditions dominated and photosynthetic oxygen production was the main driver of DO-saturation (Villate et al. 2013).
The nearby (ca. 25 km apart) estuary of Urdaibai is a less polluted, shallower, and less stratified system. A preliminary work (Iriarte et al. 2010) in which seasonal drivers of variations in DO-saturation were assessed showed that both hydro-climatic and chlorophyll a concentration make a contribution to variations in DO-saturation in the inner estuary, whereas in the outer estuary water column ventilation processes related to strong water column mixing and very high tidal flushing were more important than oxygen production or consumption processes. However, DO-saturation patterns were studied at a single depth and no analysis of the effect of hydro-climatic factors and eutrophication at an inter-annual time scale was conducted. The hypothesis is that the hierarchy (mainly on a qualitative basis) and the effect of hydro-climatic and trophic factors governing DO dynamics would differ not only axially and vertically but also from seasonal to inter-annual scales at a given salinity zone. For this reason, the present work aimed to analyze the spatial (both axial and vertical) variations in temporal changes (both seasonal and inter-annual trends) of DO-saturation in the estuary of Urdaibai during the period 1998–2008 and assess the contribution of hydro-climatic factors and phytoplankton biomass to variations in DO-saturation, both at seasonal and inter-annual time scales, at different depths and salinity zones along the longitudinal axis of the estuary. Since information in the literature on estuarine DO dynamics is somewhat biased towards large estuaries with a marked freshwater influence, where oligo-, meso-, or polyhaline habitats dominate (e.g., Gironde estuary, Chesapeake Bay estuary, Changjiang estuary, etc.), results from the present study can be a very useful contribution to fill such a typology gap, since they can illustrate DO dynamics in small estuaries with relatively small river discharges, where euhaline habitats dominate, but subject to occasional torrential river discharges, a type of estuary common for instance in the northern Iberian peninsula (on the central and eastern southern Bay of Biscay), but for which there is more limited information in the literature.
Materials and Methods
Study Area
The temperate estuary of Urdaibai, also known as the estuary of Mundaka, is located on the Basque coast, in the inner Bay of Biscay, within the middle latitudes (43°22′N, 2°43′W) of the eastern North Atlantic (Fig. 1). It is a relatively short (12.5 km) and shallow (mean depth of 3 m) meso-macrotidal system, with a maximum and minimum width of 1.2 km and <20 m in the outer and inner areas, respectively, having a mean volume of ~3.3 × 106 m3 (Villate et al. 1989). Salt marshes are abundant at its upper and middle reaches and the central channel is bordered by relatively extensive intertidal flats (mainly sandy) and sandy beaches at its lower reaches. The main drainage branch in the inner estuary is an artificial channel, which connects the Oka River in the estuary head with the natural channel in the middle estuary. The watershed area is ~142 km2 and river discharges are usually low in relation to the estuarine volume. In consequence, most of the estuary exhibits marine dominance, with high salinity waters in the lower half and a stronger axial gradient of salinity towards the head, where it receives most of the freshwater inputs from its main tributary, i.e., the Oka River. However, this is a torrential type of river and at times of very high river flow lower salinity waters can dominate in the estuary (de Madariaga et al. 1992). At the upper reaches, the estuary also receives large amounts of nutrients (mainly ammonia and phosphate, see Iriarte et al. 1997) and organic matter from the Gernika waste water treatment plant (WWTP) that works with a rather low efficiency (Franco et al. 2004). At the outer zone, tidal flushing is high, to the extent that waters of salinities >34 are flushed out of the estuary with each tidal cycle (Villate et al. 1989). In the outer half of the estuary, the water column is well mixed whereas at the inner half it is partially stratified (see Iriarte et al. 2010).

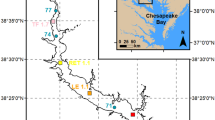
Map of the estuary of Urdaibai showing the location of the salinity zones (26, 30, 33, and 35) at high tide. (WWTP, Gernika wastewater treatment plant)
Sampling Scheme and Data Set Acquisition
The data set (1998–2008) used in the present study was obtained in a plankton monitoring program of Basque coast estuaries in which samplings were carried out monthly, during neap tides at high water, in four selected salinity zones of the estuary of Urdaibai in which at ca. mid depth, below the halocline, there are waters of salinities of 26 (±1), 30 (±1) 33 (±0.5), and 35 (±0.5). All samplings were carried out in the morning and effort was made to sample at a similar time of the day, although this was not always possible due to tidal variations. The estuary stretch within which each salinity zone was sampled is shown in Fig. 1. These salinity zones were selected because the monitoring program aimed to describe variations in DO-saturation in the water masses that define the main pelagic habitats supporting zooplankton in the estuary of Urdaibai (Uriarte and Villate 2004). The spatial zonation of salinity in the estuary of Urdaibai varies due to the effect of tides and river discharge (de Madariaga et al. 1992; Villate 1997), and therefore, instead of sampling at spatially fixed stations, a Lagrangian-like sampling strategy at selected salinity sites was adopted (see Kimmerer et al. 1998; Moderán et al. 2010).
At each sampling site, vertical profiles (every 0.5 m) of salinity, temperature, and percentage saturation of dissolved oxygen (DO-saturation) were obtained in situ using a WTW multi 350i Multi-Parameter Water Quality Meter, and water samples were collected for chlorophyll a analysis from below the halocline. Chlorophyll a was measured spectrophotometrically following the monochromatic method with acidification (Jeffrey and Mantoura 1997). Secchi disk depths were also measured.
Monthly mean values of river discharge for the period of study, measured at the hydro-meteorological station of Muxika (OK01), were obtained from the Provincial Council of Bizkaia (http://www.bizkaia.net/home2/Temas/DetalleTema.asp?Tem_Codigo=2684&idioma=CA&bnetmobile=0&dpto_biz=9&codpath_biz=9|347|2679|2684).
Data Analyses
Missing data gaps (ca. 10 %) for individual months were filled with the corresponding monthly mean for the whole period of study. In the case of river discharge data, however, missing values were estimated from regression models with the complete series from the nearest gauging station, which was the series for the Lea stream measured at Oleta (LE02) station (y = 0.3892x; R 2 = 0.9333).
From the analysis of the vertical profiles of salinity, it was concluded that in the 35 salinity zone the water column was well mixed, with occasional weak gradients in surface, but in the 33, 30, and 26 salinity zones, the water column usually showed vertical salinity gradients. Since DO-saturation, salinity, and temperature data were obtained at 0.5 m depth intervals, the average water column values were used for the 35 salinity zone. However, for the 33, 30, and 26 salinity zones, two layers were distinguished: the top layer that extended from the surface down to the depth of maximum salinity difference in a 0.5-m interval, which in the present work has been termed as the salinity gradient layer (SGL), and the layer below it, which in the present work has been termed as the layer below the salinity gradient (BSGL).
Water column stratification has been claimed to be an important factor affecting bottom water ventilation (Codiga 2012). In the Urdaibai estuary, stratification is mainly caused by depth differences in salinity, which varies at both spatial and temporal scales. Testing the influence of salinity stratification in the spatial and temporal variations of DO-saturation was, therefore, considered important. Salinity stratification indices were estimated in two ways. A salinity stratification index named salinity stratification index 1 in the present work was calculated as the difference between bottom and surface salinities divided by the mean water column salinity, similarly to the n s stratification parameter described in Haralambidou et al. (2010). The term salinity stratification index 2 as used in the present work corresponds to the maximum difference in salinity at 0.5 m depth intervals obtained in the water column. This second stratification index was also used in a previous work (Villate et al. 2013) as an index to reflect the sharpening of the salinity gradient associated to the narrowing of the halocline layer.
Prior to data analysis, chlorophyll a data were transformed to log (x + 1) to achieve homogeneity of variance. In order to visualize the seasonal and inter-annual trends in variables at each salinity zone, the temporal sequences of the corresponding monthly data and their 12-month moving averages were plotted. To determine annual DO-saturation anomalies, yearly medians and the grand median (i.e., the median of the yearly medians) were estimated and then the deviations of yearly medians from the grand median were calculated.
To analyze the relationships between DO-saturation and the rest of environmental variables, we used Spearman rank correlation analysis, and in order to estimate the percentage of the variability in DO-saturation that could be explained by each of these environmental variables, we performed stepwise regression analyses. We used the stepwise regression with a combination of the forward and backward selection techniques. It is a modification of the forward selection so that after each step in which a variable is added, all candidate variables in the model are checked to see if their significance has been reduced below the specified tolerance level. If a nonsignificant variable is found, it is removed from the model. The specified tolerance levels were that the F probability to enter a variable had to be ≤0.05, and ≥0.1 to be removed. Also, in order to distinguish the inter-annual component of variation, the correlation and regression analyses were performed using both raw data and deseasonalized data. Following Lehman (2004) and Villate et al. (2013), the deseasonalized data were obtained by calculating the difference between the monthly value and the average for all years for each month divided by the standard deviation. The correlation and regression analyses were carried out using the IBM SPSS Statistics 20 package. The Durbin–Watson test was performed to detect the presence of autocorrelation in the residuals from the regressions analysis, using the IBM SPSS Statistics 20 package.
Results
Spatial and Temporal Variations of the Percentage Saturation of Dissolved Oxygen (DO-saturation)
DO-saturation values showed an outer to inner estuary decreasing gradient, both in the SGL and in the BSGL (significant differences at p < 0.001, Kruskal–Wallis H test for each depth layer) (see Figs. 2 and 3). Mean ± SD of DO-saturation (%) values were 98.5 % ± 9.9 in the 35 salinity zone, and 88.6 ± 12.7, 82.4 ± 15.1, and 74.3 ± 20.1 in the SGL of the 33, 30 and 26 salinity zones, respectively, and 91.7 ± 12.9, 84.4 ± 14.5, and 75.7 ± 18.1 in the BSGL of the 33, 30, and 26 salinity zones, respectively. In the 35 salinity zone, DO values were at or near saturation values throughout the water column, and in the rest of salinity zones, values in the BSGL were, in general, slightly higher than in the SGL (Figs. 2 and 3). However, differences between depth layers were only statistically significant in the 33 salinity zone (Mann–Whitney U test, p < 0.05).

Time series of DO-saturation (%) at the 26, 30, 33, and 35 salinity zones from 1998 to 2008 in the SGL (dashed line) and BSGL (continuous line). Thicker lines represent moving averages. For the 35 salinity zone, data are whole water column mean values (see text) and the continuous line represents moving averages

Monthly values of temperature, stratification index 1, stratification index 2, DO-saturation (%), chlorophyll a concentration, and Secchi disk depth in the 26, 30, 33, and 35 salinity zones from 1998 to 2008. Temperature and DO-saturation values for the 26, 30, and 33 salinity zones are shown as mean values in the SGL and BSGL, and for the 35 salinity zone as mean water column values. No Secchi disk depth data are shown for the 35 salinity zone (see text)
Seasonal variations in DO-saturation showed similar patterns in the two depth layers along the estuary, with maximum values in late winter-spring and minimum in late summer, but variations of DO-saturation throughout the annual cycle were most marked in the inner estuary and least marked in the outer estuary (Fig. 3). A closer examination of these patterns revealed that in the intermediate and inner estuary (except in the SGL of the 26 salinity zone) the annual maxima of DO-saturation occurred, in general, later than in the 35 salinity zone. Also, in the 26 and 30 salinity zones, the summer decline in DO-saturation started earlier than in the 33 and 35 salinity zones. Hypoxia, with DO-saturation values slightly lower than 30 %, was only observed occasionally in August or September in the 26 salinity zone (1998, 1999, 2003, and 2005), generally at surface (Fig. 2).
The analysis of the year-to year variations revealed that DO-saturation anomalies were <±15 % in all salinity zones, being highest and lowest in the 26 and 35 salinity zones, respectively (Fig. 4), but no significant increasing nor decreasing trend was detected in the time series. Axial differences were evident. In the inner estuary (26 and 30 salinity zones), the highest positive anomalies occurred in 2003 and the lowest in 2006, but in the outer estuary (35 salinity zone) these were observed in 2008 and 2007, respectively. It is worth noting, for example, that 2003 was the year of maximum DO-saturation in the entire estuary. It coincided with unusually high sustained chlorophyll a values for most of the year (see Fig. 7). In contrast, the year with the maximum negative anomaly in DO-saturation at the 30 and 26 salinity zones was 2006, which was the year that showed the longest spring-summer dry period of the series (see Fig. 6).

Inter-annual variations in DO-saturation depicted as yearly deviations from the annual median for the time series in the 30, 33, 34, and 35 salinity zones from 1998 to 2008. Data for the SGL and BSGL are shown, except for the 35 salinity zone, for which whole water column values are shown
Spatial and Temporal Variations of Other Environmental Factors
Hydro-climatic Factors
The seasonal pattern of water temperature was the typical for temperate regions with minima in winter and maxima in summer (Fig. 3). In the year-to-year variations, 2002 stands out as the year with the coldest summer of the series, whereas the summers of 2003 and 2006 were the warmest (Fig. 5).

Time series of water temperature at the 26, 30, 33, and 35 salinity zones from 1998 to 2008 in the SGL (dashed line) and BSGL (continuous line). Thicker lines represent moving averages. For the 35 salinity zone, data are whole water column mean values (see text) and the continuous line represents moving averages
As it corresponds to an area with a temperate oceanic climate, river flows were highest during late autumn and winter and lowest during summer (Fig. 3). During late autumn 2001 and winter 2002, river flows were unusually low (with regards to other years of the time series analyzed in the present work) and they were higher than usual during the summer of 2002 (Fig. 6). The highest values of the series were recorded in autumn 2008.

Time series of monthly mean values of river discharge and of stratification index 1 and stratification index 2 at the 26, 30, 33, and 35 salinity zones (26 salinity in black and 30 to 35 salinities from darker to progressively lighter gray) from 1998 to 2008. Thicker lines represent moving averages
In the outer estuary, the water column was well mixed throughout the year and salinity stratification increased progressively towards the inner estuary (Figs. 3 and 6). The seasonal patterns of stratification index 1 and stratification index 2 were quite similar and they were broadly in agreement with the seasonal pattern of river flow, showing maxima in late autumn and winter and minima in summer (Fig. 3). Nevertheless, the stratification index 2 showed more irregular spatial and temporal patterns than the stratification index 1.
Chlorophyll a and Turbidity
Throughout most of the annual cycle, chlorophyll a concentrations showed an outer to inner estuary increasing pattern (Figs. 3 and 7). Furthermore, the seasonal cycle of chlorophyll a showed clear differences along the longitudinal axis of the estuary. In the outer estuary (35 salinity zone), the seasonal pattern was bimodal with a larger peak in late-winter to early spring and a smaller one in late summer-early autumn (Fig. 3). In the intermediate and inner estuarine zones, however, chlorophyll a concentration peaked in summer, but the more landward, the period of high chlorophyll a values was longer, starting in spring and extending throughout the summer until August in the 30 and 26 salinity zones (Fig. 3). There was no clear inter-annual trend of variation for chlorophyll a concentration in any salinity zone (Fig. 7). Annual averaged chlorophyll a values in the estuary were highest in 2003, and the second highest in 2001.

Time series of Secchi disk depth and chlorophyll a concentration at the 26, 30, 33, and 35 salinity zones (26 salinity in black and 30 to 35 salinities from darker to progressively lighter gray). Thicker lines represent moving averages
Secchi disk depth showed an outer to inner estuary decreasing gradient (Figs. 3 and 7). Data from the 35 salinity zone is not shown because most of the times the disk was visible at the bottom, in which cases the real magnitude of water transparency could be underestimated. No clear common seasonal trend of variation was apparent for any of the salinity zones (Fig. 3); however, year-to year variation tendencies coincided for all the salinity zones (Fig. 7).
Relationships Between DO-saturation and Other Environmental Factors
Tables 1 and 2 contain respectively the correlation and stepwise regression analyses results performed with the raw data. In the outer estuary (35 salinity zone), where seasonal and inter-annual variations of DO-saturation were smallest, no significant correlation (p > 0.05) between DO-saturation and any of the rest of environmental factors under study was observed.
In the intermediate and inner estuary reaches, the factors that showed statistically significant correlations with DO-saturation were essentially the same for the SGL and BSGL. In these intermediate and inner estuary salinity zones, river discharge was the variable that best correlated with DO-saturation, showing a positive correlation. In the 30 and 26 salinity zones, other hydrological parameters closely related to river discharge, such as the stratification indices, were also positively correlated to DO-saturation. In addition, temperature showed negative correlation with DO-saturation in the 30 and 26 salinity zones.
The stepwise regression analyses results were mainly an aid to discern the hierarchy of the effects on DO-saturation. It has to be pointed out that we tested for autocorrelation in the residuals from the regressions (Durbin–Watson test) and no autocorrelation was observed. In the 35 salinity zone, the regression model selected stratification index 2 as the single explicative variable. The coefficient was negative, likely denoting that when the effect of increased river discharge (which causes some degree of stratification) reached this outer estuary, it resulted in small decreases of DO-saturation. In the 33 salinity zone, a statistically significant regression model was only obtained for the SGL and this showed Secchi disk depth as the single explicative variable, with a positive effect, that is to say that the more turbid the water column, the lower the value of DO-saturation. In the 30 and 26 salinity zones, the variable that explained the largest proportion of variations in DO-saturation was river discharge, followed by chlorophyll a and temperature. This reflects the seasonal variations in the factors controlling DO-saturation in these two salinity zones. In winter, from January to March, river discharge was high and DO-saturation was also high. However, from March to September, river discharge decreased, but DO-saturation started to decrease only from May onwards, and this seems to be linked with the fact that chlorophyll a concentrations increased considerably from March onwards, which would indicate the positive effect of autotrophic processes on oxygen production. However, despite chlorophyll a concentrations remained high until August in these salinity zones, DO-saturation decreased considerably from May to September, which appears to be related to the increase in temperature.
Results of the correlation and stepwise regression analyses performed with the deseasonalized data have been presented in Tables 3 and 4, respectively. It has to be born in mind that the year-to-year variations in DO-saturation were smaller than the seasonal variations. In the 35 salinity zone and in the SGL of the 33 salinity zone, hydrological factors such as stratification indices showed negative correlations with DO-saturation, which would be indicative of the negative effect of increased river discharge, at times when this would cause slight salinity variations with depth in the 33 and 35 salinity zones. This is in agreement with regression analyses results obtained for the raw data. In the 30 and 26 salinity zones, river discharge and chlorophyll a showed the highest correlations with DO-saturation; however, no significant negative relationship with temperature was observed. Instead, in one case temperature appeared to be positively correlated with DO-saturation. Regarding the stepwise regression analyses, no statistically significant models were obtained for the 35 and 33 salinity zone data. For the 30 and 26 salinity zones, in agreement with the correlation analyses results, river discharge and chlorophyll a appeared as important explicative variables with positive effects in both depth layers, whereas the models did not include negative effects of temperature. In one case the model included (with a marginal percentage of variability explained) temperature, but with a positive effect.
A comparison of all the spatial and temporal variations in DO-saturation considered in the present work shows that the percentage of variation explained by the stepwise regression models was higher for the SGL than for the BSLG, that it was much lower in the outer estuary than in the intermediate and inner estuarine zones, and that it was higher for the seasonal variations than for the year-to-year variations (Tables 2 and 4).
Discussion
Spatial Variations of DO-saturation
The pattern of decrease in DO-saturation from the outer to the inner sites, observed in the estuary of Urdaibai, has also been described for other estuaries on the Basque coast (e.g., estuary of Bilbao (Iriarte et al. 2010; Villate et al. 2013)) and conforms with the most common, albeit not universal (e.g., Yakina estuary (Brown and Power 2011)), condition observed in bottom waters of estuaries (e.g., Westerschelde estuary (Soetaert and van Rijswik 1993); Thames estuary (Trimmer et al. 2000); Long Island Sound (Lee and Lwiza 2008); lower Saint Lawrence estuary (Lefort et al. 2012); Gironde estuary (Lanoux et al. 2013)). Several factors can contribute to this decrease in DO-saturation levels in the inner reaches of the estuary of Urdaibai. The inner estuary receives larger amounts of organic matter and inorganic nutrients of fluvial origin, as well as from discharges from the Gernika WWTP, which fuel both primary production and respiration (Revilla et al. 2000). In addition, in the outer estuary, tidal flushing is high, to the extent that in this zone the estuary almost empties itself with each tidal cycle (Villate et al. 1989). As a consequence, water renewal rates are higher and inorganic nutrient, organic matter, chlorophyll a concentrations, and primary production rates are generally much lower than in the inner estuary (Ruiz et al. 1994; Iriarte et al. 1997; Revilla et al. 2000), waters at the outer estuary generally being normoxic and inner estuary waters having lower levels of DO-saturation.
In the outer estuary (35 salinity zone), DO-saturation distributed homogeneously with depth, in agreement with the uniform distributions of temperature and salinity, which is due to this estuarine zone being a high tidal and wave energy zone where the water column is well mixed. In the intermediate and inner estuary, however, mean values in the SGL were, in general, slightly lower than in the BSGL, being these differences statistically significant in the 33 salinity zone. This is in contrast with conditions found in most estuaries and coastal systems experiencing DO depletion, where water column stratification restricts aeration of bottom waters, thus causing depletion of bottom water DO and, therefore, causing a surface to bottom decreasing gradient in DO (Hagy et al. 2004; Wild-Allen et al. 2013). In the 33, 30, and 26 salinity zones of the estuary of Urdaibai, the water column is only partially mixed (see Iriarte et al. 2010) and in such type of estuarine zones the advective transport is primarily due to gravitational circulation, with a net up-estuary flow of denser, saltier water near the bottom (Pritchard 1952). In this type of estuaries with large longitudinal salinity gradients, gravitational circulation has been shown to replenish DO in bottom waters causing lower DO concentration near the surface and higher near the bottom (Kuo and Neilson 1987; Lin et al. 2006).
Although low DO-saturation levels (<50 %) were a common feature in the SGL of the 26 salinity zone in late summer, in agreement with previous results (Revilla et al. 2000), hypoxic waters (<30 % DO-saturation, Rabalais et al. 2010) were only occasionally detected in this area. However, in a small more landward stretch of estuary of lower salinity and higher stratification, close to the WWTP discharge point, hypoxia has been reported to occur intermittently from spring through summer in bottom waters (Franco et al. 1996).
Seasonal Variations of DO-saturation
The seasonal pattern of variation of DO-saturation with summer minima observed in the estuary of Urdaibai is the most common seasonal pattern observed in estuaries and other coastal waters (Rabalais et al. 2002; Lin et al. 2008; Codiga et al. 2009; Lanoux et al. 2013; Villate et al. 2013). In some systems, hypoxia starts already by early spring (Justić and Wang 2014), whereas in the estuary of Urdaibai, the initiation of the seasonal depletion of DO is delayed until May-June. Furthermore, in some permanently stratified estuaries or estuarine zones which are very deep or with high organic loading, bottom waters are consistently hypoxic or anoxic throughout the entire seasonal cycle (Dyrssen et al. 1996; Gilbert et al. 2005).
In the outer estuary (35 salinity zone), no significant relationship was detected between DO-saturation and the possible driving factors that we tested, except for stratification index 2, which explained only a very small percentage of the variability in DO-saturation. It has to be born in mind that DO-saturation was generally over 90 % and that seasonal variations were small (9.6 %) in this outer estuary as compared to the inner estuary (43.2 and 36.2 % in the SGL and BSGL, respectively). The increase of stratification in this high-salinity vertically homogenous zone can only be due to the seaward spread of lower density surface waters modified through dilution with freshwater. Therefore, the negative relationship of DO-saturation with stratification index 2 is likely due to the increase of organic matter in the outer estuary with the arrival of lower salinity waters richer in organic matter, which would bring about a drawdown of DO-saturation resulting from the decomposition of such organic substrates. Drops in DO due to increases in the concentration of oxygen consuming compounds that occur immediately after rainfall events have also been observed in other estuaries (Pearce and Schumann 2003). In any case, given the high tidal flushing (Villate et al. 1989) and well mixed condition of the water column, it can be hypothesised that physical processes of tidal exchange and turbulence are the main mechanisms controlling DO-saturation in the outer estuary of Urdaibai and responsible for keeping it usually at normoxic levels (see also Iriarte et al. 2010). High physical energy estuarine zones are generally less prone to DO depletion (Diaz 2001; Wang 2009) and this is the case for the outer estuary of Urdaibai. The high physical energy in the outer estuary of Urdaibai is not only due to tidal forcing, but it is also attributable to currents or winds (Borsuk et al. 2001). It is interesting to note that in the estuary of Bilbao, a nearby system located ca. 25 km westward, seasonal dynamics of DO-saturation in the outer estuary are mainly driven by photosynthetic processes, but pollution levels (inorganic nutrients and organic matter concentrations) are higher and hydrodynamic conditions are quite different in the outer zone of the estuary of Bilbao, where the water column is deeper and not well mixed, but partially stratified, and tidal flushing is not as high as in the outer estuary of Urdaibai (Iriarte et al. 2010; Villate et al. 2013).
In the intermediate 33 salinity zone, and in a stronger way in the inner salinity zones of 30 and 26, river discharge and related factors such as turbidity or stratification were positively correlated with DO-saturation. In these cases, high river discharges had a positive influence on DO-saturation, likely via the increase in water renewal rate and advection of oxygen-rich freshwater. Positive correlations between river discharge and DO-saturation, related to an increase in water renewal rate, have also been reported for inner estuarine sites in the Gironde estuary (Lanoux et al. 2013) and positive effects have also been reported for the case of extreme river discharges in other systems (Lee and Lwiza 2008; Sato et al. 2012). Thus, the estuary of Urdaibai appears to be a system in which the prevailing effect of river discharge on DO-saturation (positive via increases in water renewal rate and oxygen replenishment or negative via increased respiratory substrates) varies at different sites and this is related to the typology of the estuarine zone. In the outer well-mixed zone with high tidal flushing, high river discharges promote drops in DO, but in more stratified and less tidally flushed sites located landward from this area, high river discharges cause DO-saturation to increase. This outer to inner estuary variation in the prevailing effect (negative to positive) of river discharge on DO-saturation was also observed in the nearby estuary of Bilbao, in which there is also an outer to inner estuary increasing gradient of stratification intensity (Villate et al. 2013).
In the salinity zones of 30 and 26, however, in addition to river discharge, which was the factor that showed the highest correlation with DO-saturation, chlorophyll a (positively), and temperature (negatively) also exerted an influence, each factor gaining relevance in different periods of the year. During autumn and winter, high river discharge and low temperature can both contribute to maintain DO-saturation levels high, but in spring (April and May), autotrophic production of oxygen would make DO-saturation levels to increase, despite lower river discharges. It has to be born in mind that our analysis focused on the rather immediate (non-lagged) effect on DO-saturation, and algal blooms have been shown to be accompanied with high DO values in other estuaries too (Khangaonkar et al. 2012). Furthermore, a close coupling of DO with primary production in coastal waters has been observed even under non-bloom conditions (Yoshikawa et al. 2007). However, in these salinity zones of the estuary of Urdaibai, in summer high photosynthetic production cannot counteract the decrease in DO-saturation caused by the combined effect of low river discharge and elevated temperature, the latter likely acting on DO-saturation mainly through increased rates of respiration (Revilla et al. 2000), as has been shown elsewhere (Vaquer-Sunyer et al. 2012; Bendtsen and Hansen 2013; Nydhal et al. 2013), which would make these inner waters net heterotrophic. This has been reported for other estuaries too (Smith and Kemp 1995; Caffrey 2003; Russell and Montagna 2007). Iriarte et al. (1997) found that respiration rates based on substrates not associated with the autochthonous pelagic primary production increased greatly from the outer to the inner estuary of Urdaibai. However, high temperature is not an imperative for DO depletion in estuaries, since this occurs also in the cold months of the year in some estuarine zones with high enough reserves of stored reduced substrates in sediments (Buzzelli et al. 2002; Villate et al. 2013). Overall, the findings for the estuary of Urdaibai support what was suggested by Zhang and Li (2010) when comparing DO dynamics in different estuaries, about longer residence time of estuarine water resulting in biochemical processes dominating over physical processes in the DO budget.
In a previous study on DO-saturation dynamics conducted in the estuary of Urdaibai, but which covered a different period of years, river discharge did not appear to play such a fundamental role in DO-saturation dynamics (Iriarte et al. 2010). A comparison of the time series of river discharge used in the present work and in the work by Iriarte et al. (2010) shows that the latter contained proportionately more “odd” years (i.e., with river discharges different from usual for a given season of the year), which seems to have masked its potential role as an important environmental driver of DO-saturation. This underscores the need to maintain monitoring programs, in order to obtain time series long enough to draw sound environmental conclusions.
Given that the percentage variance of DO-saturation explained by environmental factors increased from the outer to the inner estuary, and from the BSGL to the SGL layers, it can be said that DO-saturation was more sensitive to the seasonal effects of environmental factors in inner waters and SGL waters than in outer waters, and this was the case also for the inter-annual effects, a feature also observed for other Basque coast estuaries (Villate et al. 2013).
Inter-annual Variations of DO-saturation
Over the past decades, DO levels have been generally declining in coastal and estuarine waters around the world (Diaz and Rosenberg 2008; Vaquer-Sunyer and Duarte 2008; Gilbert et al. 2010), and eutrophication has been claimed to have been the main driver of this DO drop (Diaz 2001). Predictions are now that DO will generally continue to decline owing to the combined effect of climate forcings and eutrophication (Vaquer-Sunyer and Duarte 2008; Meire et al. 2013). In the estuary of Urdaibai, there were some inter-annual variations and these were controlled primarily by hydro-climatic factors. In agreement with this finding, it has been shown that in shelf waters of the Black Sea climatic predictors have been mainly responsible for the increased intensity of hypoxia during the period 2000–2009, eutrophication having had a lower impact (Capet et al. 2013). In the estuary of Urdaibai, river discharge (or related factors such as salinity stratification) was the most influential factor on variations in DO-saturation, which, as for seasonal variations, affected differently from the outer (negative effect) to the inner (positive effect) estuary. An analysis of the literature for different estuaries also reveals these positive and negative effects. River discharge is the main driver of inter-annual variations in DO in estuaries such as Narragansett Bay, but here, river discharge has a predominantly negative effect on DO via the strengthening of water column stratification (Codiga 2012). In other systems, year-to-year variations in DO levels also appear to be negatively linked to variations in stratification, but both salinity and temperature are important drivers of stratification (Capet et al. 2013). However, positive effects have also been observed in some estuaries. As mentioned above, extreme river discharges (floods) may contribute to enhance bottom water DO through increased estuarine circulation in estuaries like Long Island Sound (Lee and Lwiza 2008). Nevertheless, in other estuaries, it is believed that an increased frequency of extreme rainfall events (particularly those occurring in warm months) will increase the frequency of hypoxia/anoxia events because floods can mobilize significant quantities of organic matter and suspended solids which are subsequently deposited in estuaries (Wong et al. 2010) and/or because of more frequent/intense stratification, as observed already in systems such as the Delaware estuary (Voynova and Sharp 2012). Given that precipitation variability has mainly a zonal nature (Dettinger et al. 1998), predictions of climate change driven rainfall and river flow variations are highly region- or site-specific (IPCC 2007). For the Basque Country region, only preliminary analyses of this type have been conducted, but results point toward general reductions in river flow (Zabaleta and Antiguedad 2012), which on their own would tend to lower DO-saturation levels.
The other important potential climatic driver of DO is temperature. However, in the estuary of Urdaibai, temperature did not appear as a significant driver of variations in DO-saturation on an inter-annual time scale, although in the intermediate-inner estuary, weak positive correlations were occasionally found. These weak positive correlations may be related to the fact that temperature was positively correlated with chlorophyll a biomass on an inter-annual basis in the intermediate-inner estuary, particularly because in years with colder than usual summers chlorophyll a biomass was low (Villate et al. 2008). However, global warming scenarios generally predict that the risk of hypoxia will increase in coastal waters, not only through the decrease in oxygen solubility but also through the increase in metabolic rates and, therefore, in the biological oxygen demand (Conley et al. 2009; Bendtsen and Hansen 2013; Meire et al. 2013), as well as in the intensification of vertical stratification and subsequent reduction in ventilation of bottom waters (Kemp et al. 2009). It has to be taken into account, though, that in the estuary of Urdaibai the inter-annual variations in river discharge were larger than those occurred in temperature during the period of the time series we analyzed. The difference in the contribution of temperature to variations in DO-saturation between seasonal and inter-annual time scales observed in the present work for the estuary of Urdaibai has also been reported for the nearby estuary of Bilbao in the same period of years (Villate et al. 2013). If warming intensifies as predicted (Christensen et al. 2007), it cannot be ruled out that a future analysis of a longer time series with a larger temperature span may reveal a higher contribution of temperature to inter-annual variations in DO-saturation, but the positive effect of temperature on enhancement of phytoplankton biomass may to some extent counteract the negative effects of temperature on DO-saturation at the inner estuary.
In the 30 and 26 salinity zones of the estuary of Urdaibai, in addition to climate forcing, chlorophyll a concentration was also influential on an inter-annual time scale, having a positive effect on DO-saturation. As mentioned above, at seasonal or shorter time scales, positive immediate relationships have been observed between phytoplankton biomass/production and DO in some coastal waters (Yoshikawa et al. 2007; Khangaonkar et al. 2012), but in other areas, a clear linkage was not detected between chlorophyll and DO not at weekly, nor monthly nor at inter-annual time scales, because of interactions of physical processes and accumulation of nutrients and organic matter in sediments from the prior year (Walker and Rabalais 2006). Phytoplankton production is sensitive to nutrient availability (Paerl et al. 2014). At present, the WWTP of Gernika discharges directly in the upper estuary of Urdaibai supplying large amounts of organic matter and nutrients such as ammonium and phosphate, but future plans are that wastewaters from Gernika will be treated in a new WWTP located in the coastal area between Bermeo and Mundaka, which will be discharging its wastes about 1 km off the coast through a sewage pipe (Eusko Jaurlaritza 2011). It is difficult to predict how this change will affect not only phytoplankton production but also microbial respiration, and consequently, levels of DO-saturation. In any case, data from the present time series will be a baseline for future possible changes to be compared with.
It is clear that climate factors can be major drivers of DO variations in estuaries and coastal waters, but site-specific physical features and anthropogenic disturbances can modulate their impact. An example of this comes from the comparison of the variations in DO-saturation and their driving factors between the estuary of Urdaibai and the nearby estuary of Bilbao (ca. 25 km apart). They both share the same climate, but the estuary of Bilbao is a more polluted and more intensely stratified estuary where hypoxia/anoxia became a serious ecosystem health problem (González-Oreja and Sáiz-Salinas 1998). Indeed, a wastewater treatment scheme for the metropolitan area of Bilbao was initiated by the Water Authorities in the early 1980s (see García-Barcina et al. 2006). As a consequence, a significant trend of increase in DO-saturation occurred (García-Barcina et al. 2006), particularly in the salinity gradient of mid estuarine waters (Villate et al. 2013), which has not been observed in the estuary of Urdaibai. Therefore, in the mid estuarine waters of Bilbao, the anthropogenically mediated pollution mitigation dominated over climatic factors as drivers of inter-annual variations in DO-saturation, whereas in the estuary of Urdaibai, hydro-climatic factors were the most relevant environmental drivers of DO during the same period.
References
Bendtsen, J., and J.L.S. Hansen. 2013. Effects of global warming on hypoxia in the Baltic Sea-North Sea transition zone. Ecological Modelling 264: 17–26.
Borsuk, M.E., C.A. Stow, R.A. Luettich Jr., H.W. Paerl, and J.L. Pinckney. 2001. Modelling oxygen dynamics in an intermittently stratified estuary: estimation of process rates using field data. Estuarine, Coastal and Shelf Science 52: 33–49.
Brown, C.A., and J.H. Power. 2011. Historic and recent patterns of dissolved oxygen in the Yaquina estuary (Oregon, USA): importance of anthropogenic activities and oceanic conditions. Estuarine, Coastal and Shelf Science 92: 446–455.
Buzzelli, C.P., R.A. Luettich Jr., S.P. Powers, C.H. Peterson, J.E. McNinch, J.L. Pinckney, and H.W. Paerl. 2002. Estimating the spatial extent of bottom-water hypoxia and habitat degradation in a shallow estuary. Marine Ecology Progress Series 230: 103–112.
Caffrey, J.M. 2003. Production, respiration and net ecosystem metabolism in U.S. estuaries. Environmental Monitoring and Assessment 81: 207–219.
Capet, A., J.-M. Beckers, and M. Grégoire. 2013. Drivers, mechanisms and long-term variability of seasonal hypoxia on the Black Sea northwestern shelf—is there any recovery after eutrophication? Biogeosciences 10: 3943–3962.
Chen, C.-C., G.-C. Gong, and F.-K. Shiah. 2007. Hypoxia in the East China Sea: one of the largest coastal low-oxygen areas in the world. Marine Environmental Research 64: 399–408.
Christensen, J.H., B. Hewitson, A. Busuioc, A. Chen, X. Gao, I. Held, R. Jones, R.K. Kolli, W.-T. Kwon, R. Laprise, V. Magaña Rueda, L. Mearns, C.G. Menéndez, J. Räisänen, A. Rinke, A. Sarr, and P. Whetton. 2007. Regional climate projections. In Climate Change 2007: the physical science basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change, ed. S. Solomon, D. Qin, M. Manning, Z. Chen, M. Marquis, K.B. Averyt, M. Tignor, and H.L. Miller. Cambridge: Cambridge University Press.
Codiga, D.L. 2012. Density stratification in an estuary with complex geometry: driving processes and relationship to hypoxia on monthly to inter-annual timescales. Journal of Geophysical Research 117. doi:10.1029/2012JC008473.
Codiga, D.L., H.E. Stoffel, C.F. Deacutis, S. Kiernan, and C.A. Oviatt. 2009. Narragansett Bay hypoxic event characteristics based on fixed-site monitoring network time series: intermittency, geographic distribution, spatial synchronicity, and interannual variability. Estuaries and Coasts 32: 621–641.
Conley, D.J., J. Carstensen, J. Aigars, P. Axe, E. Bonsdorff, T. Eremina, B.-M. Haahti, C. Humborg, P. Jonsson, J. Kotta, C. Lännegren, U. Larsson, A. Maximov, M. Rodriguez-Medina, E. Lysiak-Pastuszak, N. Remeikaite-Nikiene, J. Walve, S. Wilhelms, and L. Zillén. 2009. Hypoxia is increasing in the coastal zone of the Baltic Sea. Environmental Science and Technology 45: 6777–6783.
de Madariaga, I., L. González-Azpiri, F. Villate, and E. Orive. 1992. Plankton responses to hydrological changes induced by freshets in a shallow mesotidal estuary. Estuarine, Coastal and Shelf Science 35: 425–434.
Dettinger, M.D., D.R. Cayan, H.F. Diaz, and D.M. Meko. 1998. North–south precipitation patterns in Western North America on interannual-to-decadal timescales. Journal of Climate 11: 3095–3111.
Diaz, R.J. 2001. Overview of hypoxia around the world. Journal of Environmental Quality 30: 275–281.
Diaz, R.J., and R. Rosenberg. 2008. Spreading dead zones and consequences for marine ecosystems. Science 321: 926–929.
Dyrssen, D.W., P.O.J. Hall, C. Haraldsson, M. Chierici, J.M. Skei, and H.G. Östlund. 1996. Time dependence of organic matter decay and mixing processes in Framvaren, a permanently anoxic fjord in South Norway. Aquatic Geochemistry 2: 111–129.
Eusko Jaurlaritza, Ingurumen, Lurralde Plangintza, Nekazaritza eta Arrantza Saila. 2011. Cambio Climático. Impacto y Adaptación en la Comunidad Autónoma Vasca. Eusko Jaurlaritzaren Argitalpen Zerbitzu Nagusia. p. 110.
Franco, J., A. Ruiz, and E. Orive. 1996. Factores y escalas de variabilidad del oxígeno disuelto en el estuario de Urdaibai. Ozeanografika 1: 43–64.
Franco, J., A. Borja, and V. Valencia. 2004. Overall assessment—human impacts and quality status. In Oceanography and marine environment of the Basque country, ed. A. Borja and M. Collins, 581–597. Amsterdam: Elsevier. Elsevier Oceanography Series 70.
García-Barcina, J.M., J.A. González-Oreja, and A. de la Sota. 2006. Assessing the improvement of the Bilbao estuary water quality in response to pollution abatement measures. Water Research 40: 951–960.
Gilbert, D., B. Sundby, C. Gobeil, A. Mucci, and G.-H. Tremblay. 2005. A seventy-two-year record of diminishing deep-water oxygen in the St. Lawrence Estuary: the northwest Atlantic connection. Limnology and Oceanography 50: 1654–1666.
Gilbert, D., N.N. Rabalais, R.J. Díaz, and J. Zhang. 2010. Evidence for greater oxygen decline rates in the coastal ocean than in the open ocean. Biogeosciences 7: 2283–2296.
González-Oreja, J.A., and J.I. Sáiz-Salinas. 1998. Exploring the relationships between abiotic variables and benthic community structure. Water Research 32: 3799–3807.
Hagy, J.D., and M.C. Murrell. 2007. Susceptibility of a northern Gulf of Mexico estuary to hypoxia: an analysis using box models. Estuarine, Coastal and Shelf Science 74: 239–253.
Hagy, J.D., W.R. Boynton, C.W. Keefe, and K.V. Wood. 2004. Hypoxia in Chesapeake Bay, 1950–2001: long-term change in relation to nutrient loading and river flow. Estuaries 27: 634–658.
Haralambidou, K., G. Sylaios, and V.A. Tsihrintzis. 2010. Salt-wedge propagation in a Mediterranean micro-tidal river mouth. Estuarine, Coastal and Shelf Science 90: 174–184.
Helly, J.J., and L.A. Levin. 2004. Global distribution of naturally occurring marine hypoxia on continental margins. Deep Sea Research, Part I 51: 1159–1168.
IPCC. 2007. Climate change 2007. The physical science basis. In Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change, ed. S. Solomon, D. Qin, M. Manning, Z. Chen, M. Marquis, K.B. Averyt, M. Tignor, and H.L. Miller. Cambridge: Cambridge University Press.
Iriarte, A., I. de Madariaga, F. Diez-Garagarza, M. Revilla, and E. Orive. 1997. Primary plankton production, respiration and nitrification in a shallow temperate estuary during summer. Journal of Experimental Marine Biology and Ecology 208: 127–151.
Iriarte, A., G. Aravena, F. Villate, I. Uriarte, B. Ibáñez, M. Llope, and N.C. Stenseth. 2010. Dissolved oxygen in contrasting estuaries of the Bay of Biscay: effects of temperature, river discharge and chlorophyll. Marine Ecology Progress Series 418: 57–71.
Jeffrey, S.W., and R.F.C. Mantoura. 1997. Development of pigment methods for oceanography: SCOR-supported working groups and objectives. In Phytoplankton pigments in oceanography: guidelines to modern methods. Monographs on Oceanographic Methodology, 10, ed. S.W. Jeffrey, R.F.C. Mantoura, and S.W. Wright, 19–36. Paris: UNESCO Publishing.
Jonasson, L., J.L.S. Hansen, Z. Wan, and J. She. 2012. The impacts of physical processes on oxygen variations in the North Sea-Baltic Sea transition zone. Ocean Science 8: 37–48.
Justić, D., and L. Wang. 2014. Assessing temporal and spatial variability of hypoxia over the inner Louisiana–upper Texas shelf: application of an unstructured-grid three-dimensional coupled hydrodynamic-water quality model. Continental Shelf Research 72: 163–179.
Kemp, W.M., J.M. Testa, D.J. Conley, D. Gilbert, and J.D. Hagy. 2009. Coastal hypoxia responses to remediation. Biogeosciences 6: 2985–3008.
Khangaonkar, T., B. Sackmann, W. Long, T. Mohamedali, and M. Roberts. 2012. Simulation of annual biogeochemical cycles of nutrient balance, phytoplankton bloom(s), and DO in Puget Sound using an unstructured grid model. Ocean Dynamics 62: 1353–1379.
Kimmerer, W.J., J.R. Burau, and W.A. Bennett. 1998. Tidally oriented vertical migration and position maintenance of zooplankton in a temperate estuary. Limnology and Oceanography 43: 1697–1709.
Kuo, A.Y., and B.J. Neilson. 1987. Hypoxia and salinity in Virginia estuaries. Estuaries 10: 277–283.
Lanoux, A., H. Etcheber, S. Schmidt, A. Sottolichio, G. Chabaud, M. Richard, and G. Abril. 2013. Factors contributing to hypoxia in a highly turbid, macrotidal estuary (the Gironde, France). Environmental Science: Processes & Impacts 15: 585–595.
Lee, Y.J., and K.M.M. Lwiza. 2008. Characteristics of bottom dissolved oxygen in Long Island Sound, New York. Estuarine, Coastal and Shelf Science 76: 187–200.
Lefort, S., Y. Gratton, A. Mucci, I. Dadou, and D. Gilbert. 2012. Hypoxia in the Lower St. Lawrence Estuary: how physics controls spatial patterns. Journal of Geophysical Research 117, C07018. doi:10.1029/2011JC007751.
Lehman, P.W. 2004. The influence of climate on mechanistic pathways that affect lower food web production in northern San Francisco Bay estuary. Estuaries 27: 311–324.
Lin, J., L. Xie, L.J. Pietrafesa, J. Shen, M.A. Mallin, and M.J. Durako. 2006. Dissolved oxygen stratification in two micro-tidal partially-mixed estuaries. Estuarine, Coastal and Shelf Science 70: 423–437.
Lin, J., H. Xu, C. Cudaback, and D. Wang. 2008. Inter-annual variability of hypoxic conditions in a shallow estuary. Journal of Marine Systems 73: 169–184.
McLusky, D.S., and M. Elliot. 2004. The estuarine ecosystem. Ecology, threats and management. New York: Oxford University Press.
Meier, H.E.M., H.C. Andersson, K. Eilola, B.G. Gustafsson, I. Kuznetsov, B. Muller-Karulis, T. Neumann, and O.P. Savchuk. 2011. Hypoxia in future climates: a model ensemble study for the Baltic Sea. Geophysical Research Letters 38, L24608. doi:10.1029/2011GL049929.
Meire, L., K.E.R. Soetaert, and F.J.R. Meysman. 2013. Impact of global change on coastal oxygen dynamics and risk of hypoxia. Biogeosciences 10: 2633–2653.
Moderán, J., P. Bouvais, V. David, S. Le Noc, B. Simon-Bouhet, N. Niquil, P. Miramand, and D. Fichet. 2010. Zooplankton community structure in a highly turbid environment (Charente estuary, France): spatio-temporal patterns and environmental control. Estuarine, Coastal and Shelf Science 88: 219–232.
Moore, T.S., D.B. Nuzzio, D.M. Di Toro, and G.W. Luther III. 2009. Oxygen dynamics in a well mixed estuary, the lower Delaware Bay, USA. Marine Chemistry 117: 11–20.
Nydhal, A., S. Panigrahi, and J. Wikner. 2013. Increased microbial activity in a warmer and wetter climate enhances the risk of coastal hypoxia. Microbial Ecology 85: 338–347.
Paerl, H.W., N.S. Hall, B.L. Peierls, and K.L. Rossignol. 2014. Evolving paradigms and challenges in estuarine and coastal eutrophication dynamics in a culturally and climatically stressed world. Estuaries and Coasts. doi:10.1007/s12237-014-9773-x.
Pearce, M.W., and E.H. Schumann. 2003. Dissolved oxygen characteristics of the Gamtoos estuary, South Africa. African Journal of Marine Science 25: 99–109.
Pritchard, D.W. 1952. Salinity distribution and circulation in the Chesapeake Bay estuarine system. Journal of Marine Research 11: 106–123.
Rabalais, N.N., R.E. Turner, and W.J. Wiseman. 2002. Gulf of Mexico hypoxia, a.k.a. “The dead zone”. Annual Review of Ecology and Systematics 33: 235–263.
Rabalais, N.N., R.J. Díaz, L.A. Levin, R.E. Turner, D. Gilbert, and J. Zhang. 2010. Dynamics and distribution of natural and human-caused hypoxia. Biogeosciences 7: 585–619.
Revilla, M., A. Iriarte, I. Madariaga, and E. Orive. 2000. Bacterial and phytoplankton dynamics along a trophic gradient in a shallow temperate estuary. Estuarine, Coastal and Shelf Science 50: 297–313.
Ruiz, A., J. Franco, and E. Orive. 1994. Suspended particulate matter dynamics in the shallow mesotidal Urdaibai estuary (Bay of Biscay, Spain). Netherlands Journal of Aquatic Ecology 28: 309–316.
Russell, M.J., and P.A. Montagna. 2007. Spatial and temporal variability and drivers of net ecosystem metabolism in western Gulf of Mexico estuaries. Estuaries and Coasts 30: 137–153.
Sato, C., K. Nakayama, and K. Furukawa. 2012. Contributions of wind and river effects on DO concentration in Tokyo Bay. Estuarine, Coastal and Shelf Science 109: 91–97.
Smith, E.M., and W.M. Kemp. 1995. Seasonal and regional variations in plankton community production and respiration for Chesapeake Bay. Marine Ecology Progress Series 116: 217–231.
Soetaert, K., and P. van Rijswik. 1993. Spatial and temporal patterns of the zooplankton in the Westerschelde estuary. Marine Ecology Progress Series 97: 47–59.
Trimmer, M., D.B. Nedwell, D. Sivyer, and S.J. Malcolm. 2000. Seasonal benthic organic matter mineralisation measured by oxygen uptake and denitrification along a transect of the inner and outer River Thames estuary, UK. Marine Ecology Progress Series 197: 103–119.
Uriarte, I., and F. Villate. 2004. Effects of pollution on zooplankton abundance and distribution in two estuaries of the Basque coast (Bay of Biscay). Marine Pollution Bulletin 49: 220–228.
Vaquer-Sunyer, R., and C.M. Duarte. 2008. Thresholds of hypoxia for marine biodiversity. Proceedings of the National Academy of Sciences of the United States of America 105: 15452–15457.
Vaquer-Sunyer, R., C.M. Duarte, G. Jorda, and S. Ruiz-Halpern. 2012. Temperature dependence of oxygen dynamics and community metabolism in a shallow Mediterranean macroalgal meadow (Caulerpa prolifera). Estuaries and Coasts 35: 1182–1192.
Villate, F. 1997. Tidal influence on zonation and occurrence of resident and temporary zooplankton in a shallow system (Estuary of Mundaka, Bay of Biscay). Scientia Marina 61: 173–188.
Villate, F., J. Franco, A. Ruiz, and E. Orive. 1989. Caracterización geomorfológica e hidrológica de cinco sistemas estuáricos del País Vasco. Kobie 18: 157–170.
Villate, F., G. Aravena, A. Iriarte, and I. Uriarte. 2008. Axial variability in the relationship of chlorophyll a with climatic factors and the North Atlantic Oscillation in a Basque coast estuary, Bay of Biscay (1997–2006). Journal of Plankton Research 30: 1041–1049.
Villate, F., A. Iriarte, I. Uriarte, L. Intxausti, and A. de la Sota. 2013. Dissolved oxygen in the rehabilitation phase of an estuary: influence of sewage pollution abatement and hydro-climatic factors. Marine Pollution Bulletin 70: 234.246.
Voynova, Y.G., and J.H. Sharp. 2012. Anomalous biogeochemical response to a flooding event in the Delaware Estuary: a possible typology shift due to climate change. Estuaries and Coasts 35: 943–958.
Walker, N.D., and N.N. Rabalais. 2006. Relationships among satellite chlorophyll a, river inputs, and hypoxia on the Louisiana Continental Shelf, Gulf of Mexico. Estuaries and Coasts 29: 1081–1093.
Wang, B. 2009. Hydromorphological mechanisms leading to hypoxia off the Changjiang estuary. Marine Environmental Research 67: 53–58.
Wild-Allen, K., J. Skerratt, J. Whitehead, F. Rizwi, and J. Parslow. 2013. Mechanisms driving estuarine water quality: a 3D biogeochemical model for informed management. Estuarine, Coastal and Shelf Science 135: 33–45.
Wong, V.N.L., S.G. Johnston, R.T. Bush, L.A. Sullivan, C. Clay, E.D. Burton, and P.G. Slavich. 2010. Spatial and temporal changes in estuarine water quality during a post-flood hypoxic event. Estuarine, Coastal and Shelf Science 87: 73–82.
Yoshikawa, T., O. Murata, K. Furuya, and M. Eguchi. 2007. Short-term covariation of dissolved oxygen and phytoplankton photosynthesis in a coastal fish aquaculture site. Estuarine, Coastal and Shelf Science 74: 515–527.
Zabaleta, A., and I. Antiguedad. 2012. Klimaren seinale hidrologikoak EAEko arroetako ur-emarien joeretan. Gerora begirako proiekzio hidrologikoak. Ekaia 25: 33–59.
Zhang, H., and S. Li. 2010. Effects of physical and biochemical processes on the dissolved oxygen budget for the Pearl River Estuary during summer. Journal of Marine Systems 79: 65–88.
Acknowledgments
This research was funded by the University of the Basque Country (UFI11/37) and the Basque Government (GIC12/03). We thank the Provincial Council of Bizkaia for providing hydro-meteorological data.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Margaret R. Mulholland
Rights and permissions
About this article

Cite this article
Iriarte, A., Villate, F., Uriarte, I. et al. Dissolved Oxygen in a Temperate Estuary: the Influence of Hydro-climatic Factors and Eutrophication at Seasonal and Inter-annual Time Scales. Estuaries and Coasts 38, 1000–1015 (2015). https://doi.org/10.1007/s12237-014-9870-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12237-014-9870-x