
Abstract
The present study was carried out to understand the interaction between fibroblast and 3T3-L1 preadipocyte cells under H2O2-induced oxidative stress condition. H2O2 (40 μM) was added in co-culture and monoculture of fibroblast and 3T3-L1 cell. The cells in the lower well were harvested for analysis and the process was carried out for both cells. The cell growth, oxidative stress markers, and antioxidant enzymes were analyzed. Additionally, the mRNA expressions of caspase-3 and caspase-7 were selected for analysis of apoptotic pathways and TNF-α and NF-κB were analyzed for inflammatory pathways. The adipogenic marker such as adiponectin and PPAR-γ and collagen synthesis markers such as LOX and BMP-1 were analyzed in the co-culture of fibroblast and 3T3-L1 cells. Cell viability and antioxidant enzymes were significantly increased in the co-culture compared to the monoculture under stress condition. The apoptotic, inflammatory, adipogenic, and collagen-synthesized markers were significantly altered in H2O2-induced co-culture of fibroblast and 3T3-L1 cells when compared with the monoculture of H2O2-induced fibroblast and 3T3-L1 cells. In addition, the confocal microscopical investigation indicated that the co-culture of H2O2-induced 3T3-L1 and fibroblast cells increases collagen type I and type III expression. From our results, we suggested that co-culture of fat cell (3T3-L1) and fibroblast cells may influence/regulate each other and made the cells able to withstand against oxidative stress and aging. It is conceivable that the same mechanism might have been occurring from cell to cell while animals are stressed by various environmental conditions.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Cell-to-cell interactions play a crucial role in both normal and pathological conditions. The direct (physical) contact between two different cells might have influence on cell function and cell proliferation [1]. Co-culture techniques have long been used to study the interactions between cell populations (homotypic or heterotypic) and can influence cell differentiation as well as gene expression. It has been used in various fields of experimental biology and medicine, and it can be applied to elucidate the role of cellular communication in tissue formation and regeneration [2]. More specifically, co-culture between endothelial cells and C2C12 develops prevascularized tissue [3, 4]. Other studies, using co-cultures of neural cells with myoblasts, showed myoblast cell-induced neuronal acetylcholinesterase expression, notably for studying the neuromuscular junction [5]. Our previous study indicated that the co-culture effect of C2C12 and 3T3-L1 preadipocyte cells could influence calpain, caspase, and HSPs [6]. Similarly, the triculture system of osteoblasts, adipocytes, and mesenchymal stem cells is used to understand hyperglycemic condition and may be used to direct future research into cell-based therapies for diabetes and its secondary pathologies [7]. Thus, it is believed that the interaction between different types of cells such as fat cell, fibroblast cell, chondrocytes, and osteoblasts is essential for organogenesis, morphogenesis, and differentiation.
Reactive oxygen species (ROS) are a series of metabolic intermediates, produced during oxidative metabolism in cells. ROS encompass a variety of free radicals including the superoxide anion (O2−), hydroxyl radical (OH•), singlet oxygen (1O2), and hydrogen peroxide (H2O2) [8]. ROS can cause damage to the cells or tissue by reacting with nucleic acids, protein, and lipids [9]. H2O2 is one of the major sources for releasing ROS [10], which has been extensively used to induce oxidative stress in in vitro models [11, 12]. Various environmental stresses lead to enhanced ROS production which may cause inconsistency in meat tenderness and affect meat quality due to irregular synthesis of collagen and decreasing collagen solubility that the meat industry is facing nowadays [13]. The addition of antioxidant in fibroblast cells produced more collagen and increased collagen solubility in some muscles [14].
Oxidative stress affects fibroblast cell proliferation leading to a decrease in collagen synthesis and collagen solubility through the activation of MMPs, NF-κB, and TNF-α [15]. Similarly, oxidative stress also affects adipocyte proliferation, adiponectin, and PPAR-γ through the activation of NF-κB. Moreover, overexpression of collagen, PPAR-γ, and adiponectin protects the cell and suppresses tissue inflammation in adipocytes [16]. Treatment of recombinant full-length adiponectin could reduce the oxidative stress and superoxide production by increasing antioxidant capacity [17]. Almost all aerobic cells possess the antioxidant enzymes such as superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx), and the chain breaking antioxidant such as α-tocopherol, β-carotene, and ascorbic that detoxify ROS or reduce their effects [18]. Each cell is capable of exchanging chemical signals, ions, and enzymes with other neighboring cells into the intracellular space through the pores in the cell membrane which can influence the behavior of neighboring cells [19]. However, the molecular mechanisms behind the interactions between the fibroblast cell and fat cell are not well understood. Therefore, the present study was carried out to understand the co-culturing effects on cell growth, apoptosis, and inflammatory and adipogenic pathways in H2O2-induced fibroblast and 3T3-L1 cells and also to compare the variation with monoculture. The present study will provide an understanding of the molecular mechanisms between fibroblast and 3T3-L1 preadipocytes in oxidative stress condition, essential for biological events in animals.
Materials and Methods
Materials
The iScript TM cDNA synthesis kit (170-8891) and SsoFast Eva-Green Supermix (172-5202) were procured from Bio-Rad (Bio-Rad Laboratories, Inc., Hercules, CA). Primers were obtained from Genotech (Daejeon, South Korea). Dulbecco’s modified Eagle’s medium (DMEM) and fetal bovine serum were purchased from (Gibco® Life Technologies, Grand Island, NY). All the laboratory wares were purchased from Falcon Labware (Becton Dickinson, Franklin Lakes, NJ).
Isolation of Intramuscular Connective Tissue Fibroblasts from Hanwoo Cattle
The entire work involving the use of animals was approved by the Institutional Animal Care and Use Committee (CBNU 2015-048 revised in 2015) at Chonbuk National University. Semimembranosus (SM) muscles were excised from Hanwoo cattle (20–30 months old) immediately after slaughter at a commercial abattoir (located in Jeonju Province, South Korea). Immediately, the sample was washed once with 70 % ethanol and then quickly soaked in 1× phosphate buffered saline (PBS) containing 1 % antibiotics (500 mL PBS in 800 mL sample beaker) to remove ethanol. Then, the sample was soaked in DMEM containing 5 % antibiotics and subsequently placed in an icebox before transferring to the laboratory. After reaching the laboratory, the sample is transferred to a laminar flow hood. The muscle is excised and the top layers of tissue are trimmed and minced with fine sharp scissors [the samples are cut into small fragments (about 3 mm) using 60 mm petri dish with DMEM]. Five grams of minced muscle sample was incubated in DMEM (without antibiotics) + pronase (1 mg/mL, final concentration of the enzyme is 0.1 %) solution in a 15-mL sterile tube for 60 min with shaking (80 rpm) at 37 °C. At the end of the digestion period, the muscle fragments were transferred to the pronase solution to a 15-mL conical tube using a wide-bore Pasteur pipette. Subsequently, using a pipette with a narrow tip, the solution containing tissue fragments in pronase solution was pipetted until no clumps were visible. The suspension was collected and centrifuged at 400×g for 10 min. After centrifugation, the supernatant was discarded and the pellets were washed three times with 1× PBS and centrifuged, and at the end of each step, the supernatant is discarded. After the third washing step, sediments were resuspended in DMEM and supplemented with 1 % antibiotic/antimycotic solution and 10 % fetal bovine serum (FBS). Fibroblasts were then allowed to grow until about 95 % confluence in 75-cm2 flasks in a humidified atmosphere at 37 °C and 5 % CO2. When the cells reached 95 % confluence, cultures were split in a 1:10 ratio into new flasks with fresh media. At this ratio, confluent monolayers were obtained within 7 to 10 days after seeding. For selective detachment, the medium was aspirated from the flask, washed with PBS with a milliliter of cold trypsin-ethylenediaminetetraacetic acid (EDTA) (0.25 %), and incubated for 2 min in the incubator and gently wrapped with open palm to release the fibroblasts and aspirate the trypsin-EDTA. This was added to a 15-mL centrifuge tube already containing 5 mL of DMEM + FBS solution and centrifuged. After centrifugation, the pellets were collected and seeded into culture flasks.
Magnetic Assorted Cell Sorting (MACS) of Fibroblasts
To isolate myoblasts from the primary muscle cells, the cells were exposed to a magnetic cell sorting system (AutoMACS, Milteny Biotech, Germany). Particularly, when the cells reached 80 % confluence, they were collected and resuspended in phosphate-buffered saline (1× PBS, Gibco) supplemented with 0.5 % bovine serum albumin (BSA) and 2 mmol/L EDTA. After centrifugation (400×g for 5 min), the cell pellet was resuspended in PBS containing 200 μL of primary rabbit IgG antifibroblast-specific protein and then incubated with 20 μL of antirabbit IgG microbeads at 4 °C for 30 min. Finally, the cell suspension (approximately 107 cells in 2 mL PBS) was loaded into the magnetic cell sorting system to isolate the fibroblast. After collection of fibroblasts from the MACS separator, the purified fibroblasts could be cultured further for downstream experiments or stored in liquid nitrogen. The isolated cells were cultured in growth medium and subcultured when they reached confluence (approximately 80 %), and cells from the fourth passage were used for the present study.
Co-culture of Fibroblast Cells and 3T3-L1 Preadipocytes
Fibroblast and 3T3-L1 cells were incubated at a density of 6000 cells/cm2 and grown in DMEM containing 10 % FBS and 1 % penicillin/streptomycin at 37 °C in 5 % CO2. Then, the cells were co-cultured using trans-well inserts with a 0.4-μm porous membrane to separate fibroblast cells and 3T3-L1 preadipocytes. Each cell type was grown independently on the trans-well plates. Following cell differentiation, inserts containing adipocytes were transferred to separate fibroblast plates and inserts containing fibroblast transferred to adipocyte plates. Following co-culture for 24 and 48 h after the incubation period, the lower well cells only treated with H2O2 for 4 h were then harvested for further analysis.
Experimental Groups
The experimental groups were designed as follows: group 1: fibroblast (monoculture), group 2: fibroblast (co-culture), group 3: fibroblast (monoculture) + H2O2, group 4: fibroblast (co-culture) + H2O2, group 5: 3T3-L1 (monoculture), group 6: 3T3-L1 (co-culture), group 7: 3T3-L1 (monoculture) + H2O2, and group 8: 3T3-L1 (co-culture) + H2O2. The H2O2 concentration used was at 40 μM for 4 h.
Cell Viability
Cell viability was measured by 2 % trypan blue staining. The number of viable cells was estimated in each group by counting in a Neubauer chamber.
Estimation of Oxidative Stress Markers and Antioxidant Enzymes
The experimental group of cells was harvested by trypsinization and washed with PBS. The cells were suspended in 130 mM KCl plus 50 mM PBS containing 10 μM dithiothreitol then centrifuged at 20,000×g for 15 min (4 °C). The supernatant was collected and used for the biochemical estimations.
The oxidative stress markers such as lipid hydroperoxides (LHP) [20] and protein carbonyl (PC) [21] content were measured. The activities of enzymatic antioxidants such as SOD and CAT were measured in all experimental groups by standard methods presented elsewhere [22].
Immunocytochemistry
Cells grown on a glass coverslip in 12-well plates were cultured in DMEM. At the indication time, the cells were fixed for 10 min in 3 % paraformaldehyde in PPS and then washed twice in PBS. Cells were permeabilized in 0.1 % Triton X-100 in PBS for 10 min and then washed twice in PBS. Blocking was performed in 3 % BSA in PBS for 30 min and the cells were then incubated with collagen type I and collagen type III primary antibody (1:500 dilution, abcam® #ab34710 and abcam® #ab7778, Gyeonggi-do, 465-834, Seoul, South Korea) overnight. After three washes in PBS, the cells were incubated with secondary fluorescein isothiocyanate (FITC) conjugated antibody for 1 h at room temperature and then washed three times in PBS. The coverslip was mounted on fluorescent mounting medium (Dako, Trappes, France) and visualized under a fluorescence microscope (Carl Zeiss, Jena, Germany).
Isolation of RNA and Quantitative Real-Time Polymerase Chain Reaction
The cells were lysed using RNA extraction reagent kit (TRIzol, Invitrogen) and total RNA was extracted according to the manufacturer’s protocol. Complementary DNA (cDNA) was synthesized using 2 μg of total RNA by iScript TM cDNA Synthesis Kit from Bio-Rad. The quantitative real-time PCR (qRT-PCR) assays were performed using CFX96™ Real-Time PCR detection system (Bio-Rad). cDNA was amplified with each gene such as LOX, BMP-1, CYS, TRAP-1, TNF-α, NF-κB, caspase-7, caspase-9, PPAR-γ, adiponectin, and housekeeping gene GAPDH (Table 1). The reaction was carried out in 10 μL using SsoFast™ EvaGreen® Supermix (Bio-Rad) according to the manufacturer’s instructions. The statistical analysis of the qRT-PCR results was calculated by using the ΔCt value (Ct gene of interest − Ct reporter gene). Relative gene expression was obtained by ΔΔCt methods (ΔCt sample − −ΔCt calibrator), with the use of the sham-operated group as a calibrator for comparison of all unknown sample gene expression levels. The conversion between ΔΔCt and relative gene expression levels is as follows: Fold induction = 2−ΔΔCt, where 2−ΔΔCt is the relative gene expression [23].
Statistical Analysis
Data are expressed as mean ± SD (n = 6). Statistical evaluation was carried out by one-way analysis of variance (ANOVA) followed by Duncan’s multiple range test (DMRT) for multiple comparisons. A value of p < 0.05 was considered statistically significant.
Results
Figure 1 shows the percentage of cell viability of mono- and co-culture of 3T3-L1 and fibroblast cells under normal and oxidative stress conditions. The percentage of cell viability was significantly decreased up to 40.54 and 41.46 % in H2O2-induced monoculture of 3T3-L1 and fibroblast cells, respectively. Although the cell viability was significantly increased up to 54.05 and 53.65 % in the co-culture of 3T3-L1 and fibroblast cells, respectively. there were no significant differences between the monoculture and co-culture of 3T3-L1 with fibroblast cells and fibroblast cells with 3T3-L1 cells in normal conditions. The activities of enzymatic antioxidants and oxidative stress markers are given in Table 2. Decreased activities of SOD and CAT in the monoculture of fibroblast cell (50 and 48.07 %, respectively) and 3T3-L1 cells (49.60 and 59.52 %, respectively) were observed in H2O2-induced cells as compared to control. In the co-culture of both cells with H2O2, the levels were significantly increased compared to H2O2-induced monoculture cells. The oxidative stress markers such as LHP and PC were significantly increased in H2O2-induced monoculture of 3T3-L1 and fibroblast cells. In H2O2-induced co-culture of those cells, there was a significant decrease when compared to the monoculture.

Co-culture of fibroblast cell (a) and 3T3-L1 (b) inhibits cell death in H2O2-induced cells. Single asterisk, significant as compared with control; two asterisks, significant as compared with H2O2-induced cells. Values are mean ± SD of three independent determinations (p < 0.05, ANOVA followed by DMRT)
We analyzed the messenger RNA (mRNA) expressions of caspase-3 and caspase-7 that are related to the apoptotic pathway. Compared to control cells, the mRNA expressions of caspase-3 and caspase-7 were significantly upregulated in the monoculture of H2O2-induced cells. H2O2-induced co-culture of 3T3-L1 and fibroblast cells downregulated the expression of these genes at 24 and 48 h, as recorded in Figs. 2 and 3.

Representative RT-PCR of caspase-3 expression in co-culture of fibroblast (a) and 3T3-L1 (b) cell under basal and H2O2-stimulated oxidative stress conditions. Values are mean ± SD of three independent experiments. Single asterisk, significant as compared to CON; two asterisks, significant as compared to H2O2 (p < 0.05, one-way ANOVA followed by DMRT)

Representative RT-PCR of caspase-7 expression in co-culture of fibroblast (a) and 3T3-L1 (b) cell under basal and H2O2-stimulated oxidative stress conditions. Values are mean ± SD of three independent experiments. Single asterisk, significant as compared to CON; two asterisks, significant as compared to H2O2 (p < 0.05, one-way ANOVA followed by DMRT)
Figure 4a, b shows the mRNA abundance of NF-κB by RT-PCR analysis. The expression of NF-κB was significantly higher in H2O2-induced monocells than in the control. Upon co-culture of 3T3-L1 and fibroblast cell with H2O2 treatment cells, mRNA levels were significantly reduced when compared to H2O2-induced monoculture alone, even though the mRNA levels were significantly downregulated in the co-culture of fibroblast cells with 3T3-L1 cells. In contrast, it is downregulated in the co-culture of fibroblast cells with 3T3-L1 cells than in the monoculture of fibroblast cells. Figure 5a, b depicts the mRNA expression of TNF-α in the experimental groups. H2O2 induction of monoculture cells exhibited increased mRNA expressions of TNF-α when compared to control cells. Co-culture of fibroblast cells with 3T3-L1 cell in H2O2-treated cells significantly decreased the expression than in the monoculture of H2O2-induced cells at both 24 and 48 h. Only the 48-h incubation of co-culture of 3T3-L1 cells with H2O2-induced fibroblast cells downregulated the mRNA expressions of TNF-α but not in 24 h. Without induction of H2O2 in the co-culture cell (fibroblast with 3T3-L1 and 3T3-L1 with fibroblast), the mRNA expressions show significant upregulation compared to those in the monoculture.

Representative RT-PCR of NF-κB expression in co-culture of fibroblast (a) and 3T3-L1 (b) cell under basal and H2O2-stimulated oxidative stress conditions. Values are mean ± SD of three independent experiments. Single asterisk, significant as compared to CON; two asterisks, significant as compared to H2O2 (p < 0.05, one-way ANOVA followed by DMRT)

Representative RT-PCR of TNF-α expression in co-culture of fibroblast (a) and 3T3-L1 (b) cell under basal and H2O2-stimulated oxidative stress conditions. Values are mean ± SD of three independent experiments. Single asterisk, significant as compared to CON; two asterisks, significant as compared to H2O2 (p < 0.05, one-way ANOVA followed by DMRT)
Figure 6a, b shows the mRNA abundance of adiponectin and PPAR-γ in the monoculture and co-culture of 3T3-L1 and fibroblast cells under oxidative stress conditions. The expressions of adiponectin and PPAR-γ were significantly increased in the co-culture of fibroblast cell with 3T3-L1 cell compared to the monoculture. On the other hand, the monoculture of H2O2-induced fibroblast cells shows significant downregulation in those expressions than the control. However, under the co-culture of H2O2-induced fibroblast cells, the PPAR-γ expressions were improved than those in the monoculture cells. This confirms the increased translocation of adiponectin and PPAR-γ into the fibroblast.

Representative RT-PCR of adiponectin (a) and PPAR-γ (b) expression in co-culture of 3T3-L1 cell under basal and H2O2-stimulated oxidative stress conditions. Values are mean ± SD of three independent experiments. Single asterisk, significant as compared to CON; two asterisks, significant as compared to H2O2 (p < 0.05, one-way ANOVA followed by DMRT)
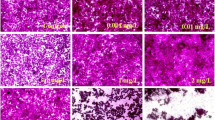
The mRNA expressions of collagen synthesizing genes such as LOX and BMP-1 are shown in Fig. 7a, b. These mRNA expressions were significantly altered in the co-culture of 3T3-L1 cells when compared to those in the monoculture cells. However, H2O2-induced monoculture of 3T3-L1 cells significantly downregulated those mRNA expressions than in the monoculture. Furthermore, the presence of 3T3-L1 cells with fibroblast cells in the same medium containing H2O2 resulted in the upregulation of the mRNA expressions of those genes in 3T3-L1 cells compared to the monoculture of H2O2-induced 3T3-L1 cells. To examine changes in the collagen type I and type III in the fibroblast and 3T3-L1 adipocytes, we used immunofluorescent staining with collagen type I and type III. The H2O2-induced monoculture fibroblast cell and 3T3-L1 showed decreased fluorescence of collagen type I and type III expression. Moreover, the H2O2-induced co-culture of 3T3-L1 and fibroblast cells increased the fluorescence of collagen type I and type III expressions. The distribution of collagen type I and type III in fibroblast cells and 3T3-L1 cells is presented in Fig. 8.

Representative RT-PCR of LOX (a) and BMP-1 (b) expression in co-culture of 3T3-L1 cell under basal and H2O2-stimulated oxidative stress conditions. Values are mean ± SD of three independent experiments. Single asterisk, significant as compared to CON; two asterisks, significant as compared to H2O2 (p < 0.05, one-way ANOVA followed by DMRT)

a–p Immunohistochemistry of collagen type I and type III on co-culture of fibroblast and 3T3-L1 cell under normal and H2O2-induced conditions at 48 h
Discussion
Excessive ROS including H2O2 inherently involved in the pathogenesis of chronic cell death and different cell types may be differentially sensitive to H2O2. They are also due to changes in the cellular composition of inflammatory cells and apoptotic cells infiltrating the wounds. It is well known that H2O2 reduces cell viability and apoptosis through activation of caspase-3 which requires the activation of inflammatory pathways such as TNF-α, TGF-β, and NF-κB [24]. In fibroblast, the higher level of H2O2 could promote cell death by reducing connective tissue formation and collagen synthesis [9]. Similarly, H2O2 induces cell death in adipocytes by inhibiting antioxidant enzymes such as catalase. Moreover, collagen protects brain cells against amyloid beta protein, the accumulation of which leads to Alzheimer’s disease [25]. Thus, we can suggest that with the co-culturing of 3T3-L1 cell with fibroblast, collagen might have penetrated from the fibroblast cell to the 3T3-L1 cell protecting from cell death at both normal and oxidative stress conditions. It is evident from our study that cell viability significantly increased in H2O2-induced co-culture of 3T3-L1 and fibroblast cells compared to that in the monoculture. Our results agreed with Holen et al. [26] who proposed the cross talk between two cell types by secreted molecules and that this influences the gene transcription and protein expression as well as the cell viability into the cell media.
SOD and CAT are antioxidant enzymes abundant in various tissues and play a vital role in modulating ROS generation. Faraci et al. [27] reported that SOD is a major antioxidant enzyme that protects cells from the harmful effects of superoxide and neutralizes free radicals. Catalase is an antioxidant enzyme highly expressed in adipose tissue than in various tissues and it protects cells from hydrogen peroxide (H2O2) by converting it to oxygen and water. Adipose tissues have highly expressed antioxidant enzymes such as SOD and CAT comparable to the liver and kidney [28]. Also, healthy fibroblast cells are produced in the dermis and epidermis which are more responsible for the production of SOD. The fibroblast is the most common cell for collagen synthesis and SOD production when the cells injured the fibroblast cells producing collagen and SOD to protect the cells from oxidative stress. The maintenance of connective tissue and collagen formation in fibroblast cells and the maintenance of antioxidant enzymes in adipocytes both contribute to the protection of cells from oxidative stress by the exchange of collagen, SOD, and CAT. However, no studies have been made regarding the role of fibroblast cell and 3T3-L1 cells in H2O2-induced co-culture. In our current experiment, the co-culture of H2O2-induced oxidative stress in 3T3-L1 cell may provide protection by localization of collagen from fibroblast cells (Fig. 8). Similarly, the co-culture of fibroblast cells with 3T3-L1 adipocytes cells may reduce cell injury or cell death by localization of SOD and CAT from 3T3-L1 cell (Table 2). This implies that the fibroblast cells contained a higher level of collagen that may infiltrate into the adipocyte and other neighboring cells which may cover the cell from cell injury. Similarly, adipocyte cells containing antioxidant enzymes mainly CAT may penetrate the fibroblast cell to protect cells from oxidative stress and may increase the enzymatic antioxidants such as SOD, CAT, GPx, etc. Also, we suggested that fibroblast and 3T3-L1 cells may not produce excessive ROS which is effectively ameliorated by the endogenous antioxidant defense system. This is in accordance with Loo et al. [9] who reported that increased neutrophil infiltration reduced the oxidative damage by decreasing lipid peroxidation.
PPAR-γ was implicated in many age-related inflammatory responses and tissue injury. H2O2-induced oxidative stress leads to the downregulation of the expression of PPARs, whereas the activation of PPAR can reduce oxidative stress [29]. PPAR-γ is highly expressed in adipose tissue and is considered to be the “master switch” of adipocyte differentiation. Overexpression of PPAR-γ in SIPS HDF cells reduced the levels of inflammatory molecules; inhibiting the NF-κB signaling pathway also induced cell survival capacity both in vitro and in vivo. Several studies reported that PPAR-γ and PPAR-α inhibit the expression of inflammatory genes, such as TNF-α, TGF-β, and MMPs [30]. PPAR-γ expression was greater in salmon liver cells, and transcription was significantly improved when liver cells were co-cultured with head kidney cells [26]. In our study, the co-culture of fibroblast cells with 3T3-L1 cells showed significantly increased PPAR-γ expression at both normal and stress conditions when compared to the monoculture. Thus, during the co-culture, 3T3-L1 cells secrete PPAR-γ which may voluntarily migrate into fibroblast cells which may protect the cell from stress.
Adipogenesis is a complex process regulated by several transcriptional factors, including PPARs. The beneficial effects of adiponectin include suppressing ROS generation and apoptosis through inhibition of endothelial NF-κB signaling and AMPK pathway [5]. Adiponectin also acts as an endogenous antioxidant and antiapoptotic agent against palmitate-induced cells [31]. Fat and fibroblast growth regulation mechanism is not yet completely understood. Our co-culture study shows that the adiponectin level was significantly upregulated in H2O2-induced co-culture of fibroblast with 3T3-L1 cell when compared to H2O2-induced monoculture cells. In line with this, fibroblast cells may recover from oxidative stress by translocation of adiponectin from 3T3-L1 in stress condition. Co-culture of fibroblast and 3T3-L1 cells provides fundamental information to understand differences between cell-to-cell effects and alteration of discrete pathways. The antiapoptotic effects of adiponectin might be promoting cell viability [31]; thus, we can suggest that adiponectin might be possible to penetrate from adipocytes to other neighboring cells to improve cell growth at stress condition.
Oxidative stress can influence the expression/activation of NF-κB while NF-κB can induce the synthesis of TNF-α and IL-6. The activation of NF-κB increases the expression of a large number of proinflammatory cytokines, stimulation of the inflammatory cascade, and caspase activation. Suppression of TNF-α activity by a TNF-α converting enzyme inhibitor that blocks TNF-α secretion improves cell proliferation [32]. The biochemical hallmarks of apoptosis were essentially activated by caspases-3 through the stimulation of TNF-α and NF-κB [33]. Thus, the H2O2-induced oxidative stress cell of monoculture cells causes inflammation through increased activation of NF-κB and TNF-α, whereas it was reduced in the co-culture through penetration of antioxidant enzymes from 3T3-L1 to fibroblast or fibroblast to 3T3-L1 cells. The production of antioxidant enzymes from adipose tissue can protect the cells from apoptosis through downregulation of caspases. There was an increased gene expression of caspase-3, TNF-α, and NF-κB in H2O2-stressed cells of monoculture, whereas it was downregulated in H2O2-induced co-culture of both 3T3-L1 and fibroblast cells. Thus, the co-cultured cells of 3T3-L1 and fibroblast cells may share the secreted molecules with each other through a medium including SOD, adiponectin, and PPAR-γ, which can downregulate apoptosis and inflammation through attenuation of the caspases, TNF-α, and NF-κB.
Recently, the co-culture of liver cells with kidney cells influences the protein levels such as arginine, and the liver cells in co-culture showed elevated or reduced gene transcription when compared to the monocultures. Co-culture of liver cells can elevate or reduce gene transcription when compared to the monocultures [26]. In our study, the expression of LOX and BMP-1 genes was higher in the co-culture of 3T3-L1 cell than in the monoculture cells. Moreover, in H2O2-induced cells in co-culture, the levels of LOX were significantly increased than in the H2O2-induced monoculture cells. From this, collagen secretion infiltrates from fibroblast cells to the 3T3-L1 adipocyte cell at both normal and oxidative stress conditions. This might have happened through contact of 3T3-L1 cells with fibroblast cells by translocation of collagen. Thus, our co-culture study indicates the alteration in LOX and BMP-1, probably the invasiveness of collagen from fibroblast cells to 3T3-L1 cells in both normal and oxidative stress conditions. LOX and BMP-1 genes were highly expressed by fibroblast cells.
Conclusion
In summary, H2O2, a free radical generator, which can induce cell death, affects antioxidant enzymes and collagen synthesis through activation of inflammation mainly TNF-α and NF-κB. Our experimental results indicated that cell viability and antioxidants such as SOD and CAT were significantly increased in H2O2-induced co-culture of 3T3-L1 and fibroblast than in the monoculture because the secreting enzymes including the antioxidants can translocate from one cell to another cell. In addition, the oxidative stress markers such as PC and LHP were altered in the co-culture of 3T3-L1 and fibroblast cells. These inferences suggest that the co-culture between two cells exchanging secreted molecules can help each other to prevent cell death from oxidative stress condition. In addition, the co-culture of 3T3-L1 and fibroblast cells can enhance the capability of each other to fight against free radicals. The same mechanism would be occurring in in vivo condition while the animal is affected by oxidative stress.
References
Spink, B. C., Cole, R. W., Katz, B. H., Gierthy, J. F., Bradley, L. M., & Spink, D. C. (2006). Inhibition of MCF-7 breast cancer cell proliferation by MCF-10A breast epithelial cells in co-culture. Cell Biology International, 30, 227–238.
Gerashchenko, B. I., & Howell, R. W. (2013). Flow cytometry-based quantification of cell proliferation in the mixed cell co-culture. Current Protocols in Cytometry Chapter 9:Unit9.40.
Sasagawa, T., Shimizu, T., Sekiya, S., Haraguchi, Y., Yamato, M., Sawa, Y., & Okano, T. (2010). Design of prevascularized threedimensional cell-dense tissues using a cell sheet stacking manipulation technology. Biomaterials, 31, 1646–1654.
van der Schaft, D. W., van Spreeuwel, A. C., van Assen, H. C., & Baaijens, F. P. (2011). Mechanoregulation of vascularization in aligned tissue-engineered muscle: a role for vascular endothelial growth factor. Tissue Engineering, 17, 2857–2865.
Jeong, H. J., Park, S. W., Kim, H., Park, S. K., & Yoon, D. (2010). Coculture with BJ fibroblast cells inhibits the adipogenesis and lipogenesis in 3T3-L1 cells. Biochemical and Biophysical Research Communications, 392, 520–525.
Pandurangan, M., Jeong, D., Amna, T., Van Ba, H., & Hwang, I. (2012). Co-culture of C2C12 and 3T3-L1 preadipocyte cells alters the gene expression of calpains, caspases and heat shock proteins. In Vitro Cellular & Developmental Biology: Animal, 48, 577–582.
Rinker, T. E., Hammoudi, T. M., Kemp, M. L., Lu, H., & Temenoff, J. S. (2014). Interactions between mesenchymal stem cells, adipocytes, and osteoblasts in a 3D tri-culture model of hyperglycemic conditions in the bone marrow microenvironment. Integrative Biology (Cambridge), 6, 324–337.
Kamata, H., & Hirata, H. (1999). Redox regulation of cellular signalling. Cellular Signalling, 11, 1–14.
Loo, A. E., Wong, Y. T., Ho, R., Wasser, M., Du, T., Ng, W. T., & Halliwell, B. (2007). Effects of hydrogen peroxide on wound healing in mice in relation to oxidative damage. PloS One, 7, 49215.
Nohl, H., Kozlov, A. V., Gille, L., & Staniek, K. (2003). Cell respiration and formation of reactive oxygen species: facts and artefacts. Biochemical Society Transactions, 31, 1308–1311.
Xiao, Y., Yuan, T., Yao, W., & Liao, K. (2010). 3T3-L1 adipocyte apoptosis induced by thiazolidinediones is peroxisome proliferator-activated receptor-gamma-dependent and mediated by the caspase-3-dependent apoptotic pathway. FEBS Journal, 277(3), 687–696.
Yang, B., Oo, T. N., & Rizzo, V. (2006). Lipid rafts mediate H2O2 prosurvival effects in cultured endothelial cells. FASEB, 20, 1501–1503.
Ferguson, D. M., & Warner, R. D. (2008). Have we underestimated the impact of preslaughter stress on meat quality in ruminants? Meat Science, 80(1), 12–19.
Maiorano, G., Cavone, C., McCormick, R. J., Ciarlariello, A., Gambacorta, M., & Manchisi, A. (2007). The effect of dietary energy and vitamin E administration on performance and intramuscular collagen properties of lambs. Meat Science, 76, 182–188.
Siwik, D. A., Pagano, P. J., & Colucci, W. S. (2001). Oxidative stress regulates collagen synthesis and matrix metalloproteinase activity in cardiac fibroblasts. American Journal of Physiology - Cell Physiology, 280, C53–C60.
Liu, G. S., Chan, E. C., Higuchi, M., Dusting, G. J., & Jiang, F. (2012). Redox mechanisms in regulation of adipocyte differentiation: beyond a general stress response. Cells, 1(4), 976–993.
Wang, W. Q., Zhang, H. F., Gao, G. X., Bai, Q. X., Li, R., & Wang, X. M. (2011). Adiponectin inhibits hyperlipidemia-induced platelet aggregation via attenuating oxidative/nitrative stress. Physiological Research, 60, 347–354.
Miller, J. K., Brzezinska-Slebodzinska, E., & Madsen, F. C. (1993). Oxidative stress, antioxidants, and animal function. Journal of Dairy Science, 76, 2812–2823.
Wilson, J. H., & Hunt, T. (2002). Molecular biology of the cell (4th ed.). New York: Garland Science. 712 pp.
Jiang, Z. Y., Hunt, J. V., & Wolf, S. P. (1992). Detection of lipid hydroperoxides using the FOX method. Analytical Biochemistry, 202, 384–389.
Levine, R. L., Garland, D., & Oliver, C. N. L. (1990). Determination of carbonyl content in oxidatively modified proteins. Methods in Enzymology, 186, 464–478.
Kakkar, P., Das, B., & Viswanathan, P. N. (1984). A modified spectrophotometric assay of superoxide dismutase. Indian Journal of Biochemistry Biophysics, 21, 130–132.
Livak, K. J., & Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCt method. Methods, 25, 402–408.
DiPietrantonio, A. M., Hsieh, T., & Wu, J. M. (1999). Activation of caspase 3 in HL-60 cells exposed to hydrogen peroxide. Biochemical and Biophysical Research Communications, 255, 477–482.
Cheng, J. S., Dubal, D. B., Kim, D. H., Legleiter, J., Cheng, I. H., Yu, G. Q., Tesseur, I., Wyss-Coray, T., Bonaldo, P., & Mucke, L. (2010). Collagen VI protects neurons against Aβ toxicity. Nature Neuroscience, 12, 119–121.
Holen, E., Espe, M., Andersen, S. M., Taylor, R., Aksnes, A., Mengesha, Z., & Araujo, P. (2014). A co culture approach show that polyamine turnover is affected during inflammation in Atlantic salmon immune and liver cells and that arginine and LPS exerts opposite effects on p38MAPK signaling. Fish & Shellfish Immunology, 37, 286–298.
Faraci, F. M., & Didion, S. P. (2004). Vascular protection: superoxide dismutase isoforms in the vessel wall. Arteriosclerosis, Thrombosis, and Vascular Biology, 24(8), 1367–1373.
Okuno, Y., Matsuda, M., Kobayashi, H., Morita, K., Suzuki, E., Fukuhara, A., Komuro, R., Shimabukuro, M., & Shimomura, I. (2008). Adipose expression of catalase is regulated via a novel remote PPARgamma-responsive region. Biochemical and Biophysical Research Communications, 366, 698–704.
Lee, Y. H., Lee, N. H., Bhattarai, G., Yun, J. S., Kim, T. I., Jhee, E. C., & Yi, H. K. (2010). PPARgamma inhibits inflammatory reaction in oxidative stress induced human diploid fibloblast. Cell Biochemistry and Function, 28, 490–496.
Xiao, Q. X., Lee, N. T. K., Carlier, P. R., Pang, Y. P., & Han, Y. F. (2000). Bis(7)-tacrine, a promising anti-Alzheimer’s agent, reduces hydrogen peroxide-induced injury in rat pheochromocytoma cells: comparison with tacrine. Neuroscience Letters, 290, 197–200.
Kim, J. E., Song, S. E., Kim, Y. W., Kim, J. Y., Park, S. C., Park, Y. K., Baek, S. H., Lee, I. K., & Park, S. Y. (2010). Adiponectin inhibits palmitate-induced apoptosis through suppression of reactive oxygen species in endothelial cells: involvement of cAMP/protein kinase A and AMP-activated protein kinase. Journal of Endocrinology, 207, 35–44.
Xie, Q. W., Kashiwabara, Y., & Nathan, C. (1994). Role of transcription factor NF-kappa B/Rel in induction of nitric oxide synthase. Journal of Biological Chemistry, 269, 4705–4708.
Sivakumar, A. S., & Hwang, I. (2015). Effects of Sunphenon and Polyphenon 60 on proteolytic pathways, inflammatory cytokines and myogenic markers in H2O2-treated C2C12 cells. Journal of Biosciences, 40, 53–59.
Acknowledgments
It should be acknowledged that this work was carried out with the support of a grant from the Next-Generation BioGreen 21 Program (Project No. PJ011101) and a grant for FTA issues (Project No. PJ01017003) of Rural Development Administration, Republic of Korea.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Subramaniyan, S.A., Kim, S. & Hwang, I. Cell-Cell Communication Between Fibroblast and 3T3-L1 Cells Under Co-culturing in Oxidative Stress Condition Induced by H2O2 . Appl Biochem Biotechnol 180, 668–681 (2016). https://doi.org/10.1007/s12010-016-2123-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12010-016-2123-7