
Abstract
The prevalence of chronic kidney disease continues to outpace the development of effective treatment strategies. For patients with advanced disease, renal replacement therapies approximate the filtration functions of the kidney at considerable cost and inconvenience, while failing to restore the resorptive and endocrine functions. Allogeneic transplantation remains the only restorative treatment, but donor shortage, surgical morbidity and the need for lifelong immunosuppression significantly limit clinical application. Emerging technologies in the fields of regenerative medicine and tissue engineering strive to address these limitations. We review recent advances in cell-based therapies, primordial allografts, bio-artificial organs and whole-organ bioengineering as they apply to renal regeneration. Collaborative efforts across these fields aim to produce a bioengineered kidney capable of restoring renal function in patients with end-stage disease.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The prevalence of chronic kidney disease (CKD) continues to outpace the development of effective treatment strategies. Best-practice guidelines for CKD suggest supportive measures that minimize comorbidities, but fail to arrest progression. Although tight blood pressure control may help, fewer than 40 % of patients achieve and maintain therapeutic targets [1, 2]. Patients with advanced disease will eventually require life-long renal replacement therapy in the form of peritoneal or haemodialysis. Although lifesaving, dialysis approximates the filtration functions of the kidney at considerable cost and inconvenience, and fails to restore the homoeostatic, resorptive, metabolic, endocrine and immunomodulatory functions. As a result of these shortcomings, patients with CKD remain at an increased risk of cardiovascular and all-cause mortality[3].
Allogeneic transplantation remains the only restorative treatment available for advanced CKD with progression to end stage renal disease (ESRD). Renal transplantation, pioneered by Dr. Joseph Murray [4], is associated with reduced healthcare costs, longer life expectancy and improved quality of life compared to dialysis [5, 6]. The majority of patients are unable to realize these advantages, however, due to the critical shortage of organs that has emerged over the last decades (Fig. 1). Furthermore, the side effects of lifelong immunosuppressive therapy dramatically impact the overall outcome and significantly limit clinical application.

Official Organ Procurement and Transplantation Network data as of August 24, 2012, reporting on the number of patients in the waiting list for a deceased donor renal transplant and the effective number of transplant performed. Importantly, steepness of the curve of the patient in the waiting list is increasing, while the curve of the number of transplants performed is flattening
Emerging technologies in the field of regenerative medicine (RM) seek to address the limitations of current treatment strategies. Strategies that are currently being implemented to bioengineer or regenerate kidneys for transplant purposes can be broadly classified into the following categories: (1) cell-based strategies; (2) developmental biology strategies; (3) bio-artificial kidneys strategies; and (4) whole-organ bioengineering strategies.
Cell-Based Strategies
The natural regenerative capacity of the kidney was first demonstrated by Oliver in 1953 using an animal model of acute renal failure [7]. Cuppage and Tate [8] proceeded to identify a population of squamoid cells attached to the basement membrane that were capable of multiplying and migrating to regenerate the tubular epithelium in response to acute injury. These pioneers first recognized that the intact basement membrane plays a crucial role in the regenerative process [7–9]. Renal regeneration after injury requires an up-regulation of gene expression and protein synthesis in response to the local release of inflammatory mediators and growth factors [10]. Up-regulation of C-myc, HSP-70 and urokinase-type plasminogen activator has been demonstrated following mechanical injury of cultured renal cells [11]. Likewise, several cytokines have been shown to influence the rate and efficacy of renal regeneration, including epidermal growth factor [12], hepatocyte growth factor [13], fibroblast growth factor-1 [14], interleukin 1 [15], and TGF-B1 [12, 16].
Cell-based technologies capitalize on these native repair mechanisms by using progenitor and stem cells to induce renal regeneration after injury. Grobstein first demonstrated the presence of renal progenitor cells in the metanephric blastema, a tissue layer of the embryonic kidney [17–20]. Discrete populations of these multipotent renal progenitors have been shown to persist into adulthood in the mesenchyme of the proximal tubule [21, 22], tubular epithelial cells [23], Bowman’s capsule [24], renal papilla [25], and cortical stroma [26]. These cells demonstrate high cloning efficiency and self-renewal potential, suggesting a role in renal regeneration [24]. The transcriptional regulators Six2 [27, 28] and Foxd+1 [26] are essential for the persistence of these progenitor populations beyond development, while Pax2 is required for later differentiation. Bussolati et al. [29] demonstrated that these renal progenitors could be reliably harvested and isolated from adult human kidneys, expanded in vitro, and reimplanted. Intravenously injected renal progenitors were shown to preferentially migrate to the site of tubular injury and contribute to renal repair [29]. The regenerative utility of adult-derived renal progenitors has been similarly demonstrated using cells harvested from Bowman’s capsule [24, 30].
In addition to these resident populations of progenitor cells, extrarenal stem cells can also be induced to differentiate into cells capable of promoting renal recovery. These include embryonic stem cells [31•, 32–38], bone marrow–derived stem cells [39–59], amniotic fluid–derived stem cells [60–63] and induced pluripotent stem cells [64–68].
Embryonic Stem Cells
Embryonic stem cells (ESCs) represent a pluripotent population of cells with the capacity to proliferate in undifferentiated form, while retaining the capacity to develop along ectodermal, mesodermal or endodermal lines [31•], as well as extraembryonic and somatic lineages [37]. The differentiation of ESCs is strongly influenced by elements of the extracellular miroenvironement, including cell–cell contacts, cell–matrix contacts and the paracrine signals [69]. With relation to the kidney, ESCs injected into primordial kidneys develop into primitive renal structures [34, 35, 38], suggesting that nephrogenic factors promote differentiation down renal cell lines. Researchers have attempted to recreate these nephrogenic factors in vitro, and have identified specific growth factors capable of inducing renal differentiation in cultured ESCs [32, 33, 36]. Despite these advantages, the clinical application of ESCs is limited by ethical and legal obstacles, as well as a high potential for teratoma formation [70].
Bone Marrow–Derived Stem Cells
Bone marrow–derived stem cells (BMDCs) allow researchers to circumvent some of the ethical and legal dilemmas inherent to ESC technology. BMDCs, especially mesenchymal stem cells (MSCs), are multipotent, produced throughout life, easily harvested, migrate across tissues, and contribute to the repair of various organs [71]. The administration of BMDCs has been shown to promote neovascularisation, reduce inflammation, inhibit apoptosis, and stimulate differentiation and proliferation in multiple systems [42]. Poulsom et al. first demonstrated that circulating BMDCs engraft into the damaged kidney, promoting the turnover and repair of renal tissues following acute injury [40]. BMDCs have since been shown to contribute to the repair of multiple renal cell types, including tubular epithelia [39, 53, 59], mesangial cells [46, 49, 56, 57], podocytes [51, 52, 56] and endothelial cells [41, 45, 55]. Despite these established benefits, controversy surrounds the mechanism of BMDC-mediated renal repair, as researchers debate whether BMDCs directly repopulate injured nephrons via transdifferentiation, or indirectly through paracrine signalling [43, 44, 48, 54, 58]. Support for the paracrine hypothesis comes from studies showing that BMDC-conditioned medium contains microvesicles and growth factors that reduce inflammation and accelerate renal repair through interactions with renal progenitors [50]. Regardless of the mechanism, injection of exogenous BMDCs has been shown to enhance renal recovery in animal models of acute kidney injury, although a therapeutic benefit in CKD has yet to be demonstrated.
However, the clinical application of BMDCs has been questioned by studies suggesting that these cells may also be involved in kidney disease. BMDCs have been shown to contribute to the development of interstitial fibrosis in mouse models of CKD [47]. Furthermore, direct renal injection of BMDCs has been associated with the development of angiomyeloproliferative lesions of unknown neoplastic potential [72]. Clearly, further research is required before the regenerative utility of BMDCs can be safely and effectively harnessed for clinical applications.
Amniotic Fluid–Derived Stem Cells
Amniotic-derived stem cells are easily harvested and cultured, have low risk for tumour development, and show a breadth of pluripotency similar to BMDCs. Perin et al. demonstrated that human amniotic fluid–derived stem cells (hAFCSs) microinjected into murine embryonic kidneys differentiated along renal cell lines, contributing to the development of primordial kidney structures [60]. Further studies have demonstrated that mTOR plays an essential part in the signalling pathways involved in the renal differentiation of hAFCs [62]. The therapeutic potential of hAFSCs in renal disease has been recently demonstrated by Rota et al. [61], who showed a pro-regenerative effect of hAFSC infusion in an animal model of acute kidney injury. Engrafted hAFSC localised primarily to the peritubular region, improving renal function, limiting tubular damage and prolonging animal survival. Again, the regenerative effect appears to result from a combination of hAFSC transdifferentiation and paracrine signalling including IL-6, VEGF, and SDF-1. The importance of local growth factors and signalling molecules was further demonstrated by experiments showing that hAFSCs preconditioned with GDNF enhanced renal function and tubular repair through increased homing of stem cells to the site of injury. Furthermore, comparative studies have demonstrated that hAFSC treatment normalises renal function more rapidly than BMDCs in an animal model of acute kidney injury [63]. Although BMDCs showed higher potential for proliferation, hAFSCs were more antiapoptotic and persistent within the peritubular capillaries. Importantly, the authors isolated different cytokines and growth factors from BMDCs vs. hAFSCs, suggesting different modes of action.
Induced Pluripotent Stem Cells
Induced pluripotent stem cells (IPCSs) are generated via retroviral infection of somatic cell lines with transcription factors [66]. IPSCs have been successfully generated from human kidney mesangial cells [65] as well as urine-derived renal cells [68]. IPSCs have garnered increasing interest in the field of regenerative medicine, as they may potentially provide an inexhaustible source of patient- and tissue-specific stem cells. Furthermore, the IPSCs have been shown to retain the epigenetic pattern of the parent cell, which may facilitate targeted, organ-specific differentiation [64] with less potential for abnormal tissue formation than ESCs or BMDCs. Despite these advantages, IPSCs have also been shown to induce an immune reaction in syngeneic mice [67], which may compromise the clinical utility. To date, the therapeutic benefit of IPSCs from acute or chronic renal disease has not been demonstrated.
Developmental Biology Strategies
While advances in stem cell technology hold promise for the future of renal regeneration and repair, investigators have further advanced the cellular approach by incorporating principles of developmental biology. This expanding field of renal regenerative medicine endeavours to create, implant and maintain a tissue structure that mimics the physical and physiological characteristics of the native kidney.
Early research in the field sought to add new nephrons to developing kidneys by implanting embryonic metanephric tissue into the renal cortex of neonatal mice [73]. Metanephric tissue is the embryological precursors of the adult kidney, characterised by ureteric buds and mesodermal blastema [74]. Several advantages have been proposed for the transplantation of primitive metanephric tissue over fully developed organs, including reduced immunogenicity due to the absence of native vasculature [74] and antigen-presenting cells [75]. The primitive donor tissue successfully differentiated and developed into functional nephrons in the neonatal host kidney, but these results were not initially reproducible in adult hosts due to lack of differentiation and acute graft rejection [76]. After protocol modifications, Rogers et al. [74] reported the successful engraftment and differentiation of metanephroi into adult hosts, with demonstrated glomerular filtration and plasma clearance. Furthermore, the metanephroi were poorly immunogenic, and were capable of persisting in vivo without host immunosuppression.
Prompted by these early successes, researchers soon endeavoured to expand the technology, investigating the possibility of xenotransplantation and human hosts. The reduced immunogenicity of metanephric transplants could potentially overcome the humoral rejection that has hindered whole-organ xenotransplantation [77–79]. Rogers et al. [80] demonstrated the successful allogeneic transplantation of porcine metanephroi into adult pigs, as well as the xenogeneic transplantation into adult mice. Both types of transplant proved successful, although xenotransplantation required the additional administration of a co-stimulatory blockade including anti-CDR45RB, anti-CD154 and anti-CD11a. Dekel et al. [81] have similarly demonstrated the successful xenotransplantation of human metanephroi into immune-deficient mice, resulting in a pattern of gene expression similar to that seen in human kidney development. Although metanephric xenotransplantation into human hosts has not yet been reported, several potential risks have been hypothesized, including cross-species infection and neoplastic transformation [82].
Researchers seeking to circumvent the limitations of xenotransplantation have sought develop a similarly inexhaustible source of primordial kidney tissue of human origin. Clearly, ethical considerations would prevent the large-scale harvesting of metanephroi from human embryos. Researchers have thus sought a means of culturing and propagating nephrons in vitro, which would allow for the generation of multiple kidney-like tissues from a single metanephroi [83]. Cells derived from both Wolffian duct/ureteric bud and metanephric mesenchyme progenitor tissues have been successfully cultured alone or in co-culture [84•]. Investigators have additionally sought to culture these cell lines on extracellular matrix gels in an attempt to reconstitute essential three-dimensional (3D) relationships [84•]. These efforts have culminated in the successful in vitro construction of renal organoids from single-cell suspensions of embryonic cell lines [85]. Following implantation under the kidney capsule of rodents, these organoids formed vascularized glomeruli and fully differentiated capillary walls that performed physiological functions, including tubular reabsorption. The generation of vascularized nephrons from single-cell suspensions signifies important progress towards the long-term goal of restoring renal function with a bioengineered kidney. However, studies reporting the long-term functiom and viability of these organoids have yet to be published.
Bio-artificial Kidney Strategies
The development of bio-artificial kidneys (BAKs) represents the intersection of regenerative medicine and renal replacement therapy. BAKs combine a hemofilter used in conventional dialysis with a bioreactor unit containing human primary renal proximal tubule cells, termed a renal tubule assisted device (RAD) [86]. The addition of the bioreactor is intended to provide the homoeostatic, resorptive, metabolic, endocrine and immunomodulatory functions of the kidney that are lacking from current renal replacement therapies. Furthermore, these units are designed to be portable or implantable, providing continuous renal support with minimal lifestyle disruption.
Since BAKs were first proposed by Aebischer et al. [87], the technology has matured from concept to clinical trial. Humes et al. reported the safe and efficacious use of BAKs in patients with acute renal failure (ARF) in the ICU setting [88]. The BAK demonstrated metabolic, endocrine and immunomodulatory activities, including glutathione degradation, hydroxylation of calcifediol, and reduction of pro-inflammatory cytokines. Subsequent Phase II trials showed improved 180-day survival in patients with ARF treated with RAD support compared to continuous renal replacement therapy alone [89]. However, these clinical trials have been mired in controversy, and the results have been called into question (see Tasnim et al. [86] for full review).
Whole Organ Bioengineering Strategies
Since Grobstein’s early characterisation of renal organogenesis, researchers have recognized that the extracellular matrix (ECM) plays a crucial role in kidney development and repair [7–9]. The extracellular matrix (ECM) is a three-dimensional (3D) framework of structural and functional proteins in a state of dynamic reciprocity with intracellular cytoskeletal and nuclear elements [90]. ECM molecules and their receptors influence organogenesis and repair by (1) providing a 3D scaffold for the spatial organisation of cells, (2) secreting and storing growth factors and cytokines, and (3) regulating signal transduction [91]. The components of the ECM include type IV collagen, entactin, proteoglycans and the laminins, a family of trimeric glycoproteins that interact with cell-surface integrins. The integrins, especially α8β1, have been shown to be crucial for metanephric proliferation, branching and epithelialization during organogenesis [92]. The importance of the ECM for normal renal function is highlighted by studies suggesting that changes to the ECM underlie common renal pathologies, including diabetic nephropathy [93].
Cutting-edge technologies in RM have recently allowed researchers to exploit and appreciate the advantages of preserving the innate ECM for organ bioengineering investigations [94–97, 98•]. Indeed, innate ECM represents a biochemically, geometrically and spatially ideal platform for such investigations, because it is biocompatible [99•], it has both basic components (proteins and polysaccharides) and matrix-bound growth factors and cytokines preserved, and at physiological levels [100], it retains an intact and patent vasculature, which sustains physiologic blood pressure when implanted in vivo [99•] and is able to drive differentiation of progenitor cells into an organ-specific phenotype [101, 102]. In other words, in natura, innate ECM represents the requisite environment for cell welfare because it contains all indispensable information for growth and function [103]. RM is now exploring the possibility of using intact ECM from animal or human whole organs for bioengineering purposes.
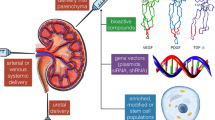
ECM scaffolds from whole animal or human-cadaveric organs can be generated through detergent-based decellularization [95–97, 104] (Fig. 2). Current decellularization protocols are capable of removing DNA, cellular material and cell surface antigens from the ECM scaffold, while preserving attachment sites, structural integrity and vascular channels [105]. Decellularization protocols involve the repeated irrigation of cadaveric tissues with detergents or acids through the innate vasculature, although organs with higher fat content, like the pancreas, often require the addition of lipid solvents, such as alcohol [106]. It remains unclear whether detergent decellularization damages essential components of the ECM, although irrigation through the existing vasculature is thought to limit potentially disruptive ECM exposure. Complete decellularization is essential as residual cellular material may contain antigenic epitopes that trigger inflammatory responses [107] and compromise subsequent recellularization [108]. Following decellularization, ethylene oxide [109] or paracetic acid [110] have been shown to effectively sterilize the ECM without denaturing the ECM proteins or growth factors, although the risk of viral contamination remains [94].

Principles of whole-organ bioengineering. Animal or human organs are processed with detergent-based solutions to remove cells. In the case of the kidney, the higher density of the renal texture requires treatment with very strong detergents like SDS, whereas less-dense organs like the heart, liver, intestine, pancreas and lung may be successfully decellularized with milder detergents like Triton or sodium deoxycholate. Once cells are removed, the so-obtained extracellular matrix scaffold represents a formidable platform for organ bioengineering and regeneration investigations. Next step is the reconstitution of the different cellular compartments, namely the parenchymal compartment to allow function and the endothelium to allow implantation. Unfortunately, while the decellularization process is quite doable for all organs of all sizes and species, the repopulation of the scaffolds remains far from the realm of the possible and represents the greatest challenge for the years to come
The decellularized, sterilized ECM serves as the scaffold on which stem cells or renal progenitors are seeded with the intent to reconstitute the cellular compartment (recellularization). The successful recellularization of ECM scaffolds has been reported in several organ systems, including liver [111], respiratory tract [112], nerve [106], tendon [113], valve [114], bladder [115] and mammary gland [116]. These results demonstrate the potential of regenerative medicine to dramatically impact organ transplantation, with the possibility of upscaling to more complex, modular organs. Bioengineered organs from autologous cells may enable surgeons to successfully address the two major obstacles of organ transplantation, namely, the need for a new, ideally inexhaustible source of organs, and the achievement of an immunosuppression-free state post-transplantation.
Recent advances have allowed researchers to apply the principles of decellularization-recellularization technology (DRT) to complex organs, including the kidney [99•, 101, 117–119]. Ross et al. [101] were the first to report the successful recellularization of intact rat kidney scaffolds with xenotransplanted murine ESCs perfused through the innate vasculature. ESC cells were chosen as a seeding population due to their high doubling capacity, pluripotency, and potential to differentiate and integrate into primordial kidney cultures. The decellularized kidney scaffold successfully supported the growth and migration of the xenotransplanted ESCs within glomerular, vascular, and tubular structures while inducing differentiation down renal cell lines. Within 10 days of seeding, the ESCs showed gross morphological changes consistent with epithelial maturation, as well as immunohistochemical markers of renal differentiation, including Pax-2 [120], Ksp-cadherin [121] and pan-cytokeratin [122]. Although the importance of their seminal findings cannot be overstated, Ross et al. identified several lines of investigation that must be satisfied in order to advance the field. The authors theorize that pretreatment of the ESC seed population with prodifferentiation agents, such as retinoic acid, activin-A and bone morphogenetic protein 7 (BMP7) [34], may promote engraftment and proliferation by providing a more kidney-specific lineage. Furthermore, although seeding via innate vascular channels provides even cell distribution in the renal cortex, the renal collecting system is omitted. Retrograde seeding through the ureter was attempted to address this problem, but this route was associated with uneven cell dispersion, possibly due to the papillary architecture of the rat kidney. Furthermore, murine scaffolds do not adequately reflect the size and structure of the adult human kidney. The authors suggest that the kidney architecture of higher-order mammals may thus be better suited as a platform for kidney bioengineering. Furthermore, larger pig and primate organs have the physiological and structural capacity to support the critical mass of nephrons required to meet human renal requirements.
The application of DRT protocols to larger, complex organs can be challenging, as perfusion decellularization relies in part on diffusion. Larger organs with greater parenchymal mass require higher perfusion pressures, stronger detergents, and prolonged detergent exposure that may damage the native vasculature and ECM proteins [99•]. Nakayama et al. [118] first performed decellularization studies on sectioned non-human primate kidneys. Microscopy, histology and immunohistochemistry confirmed effective bath decellularization with preservation of native ECM architecture and protein complement. Interestingly, their results showed that decellularization rate correlated inversely with the animal age, due in part to the greater presence of polysaccharide chains in fetal ECM. The authors further performed recellularization via direct extension by culturing scaffolds layered with explants from unrelated donors. Immunohistochemistry showed effective local migration of renal cells to the explant-scaffold border through subsidiary Pax-2 positive and vimentin-positive cell attachment and migration. These findings served as effective proof-of-principle, demonstrating that primate kidneys were amenable to DRT protocols.
More recently, intact, whole-organ ECM scaffolds have been successfully produced from porcine kidneys [99•, 119] (Fig. 3). Orlando et al. [99•] reported effective decellularization of the porcine kidney via detergent perfusion of the innate vasculature. Decellularization was confirmed by histological staining and scanning electron microscopy, which showed total cell clearance and preservation of the scaffolding architecture of the glomeruli, tubules and vessels. Importantly, the decellularization protocol preserved the patency and hierarchical branching structures of the vascular network, which is imperative for subsequent transplant, perfusion and recellularization. The acellular, whole-organ scaffolds were then implanted and surgically reconnected, and maintained for 2 weeks in vivo. Blood pressure was sustained throughout the study, and no blood extravasation was observed, proving the integrity of the intrinsic vasculature. At retrieval, the renal artery and vein were obstructed by massive thrombi despite strong anticoagulation prophylaxis. The observed coagulation was an expected by-product of the interaction of whole blood with the de-endothelialized vascular network. Further dissection showed non-specific inflammatory infiltrate, though the animals did not show any observable signs of fever or infection during the recovery period.

Porcine kidneys were decellularized with SDS-based solution. Cell clearance was quite spectacular. After decellularization, the renal ECM scaffold acquires a whitish gross appearance (a, b). H&E shows total clearance of nuclear and cellular material (c), while collagen (stained pink) remains intact. Masson trichrome (d) confirms cell clearance and persistence of collagen fibers. Scaffolds were implanted in pigs (e, f). The renal vein of the scaffold was implanted on the vena cava right above the iliac bifurcation, while the renal artery was implanted on the aorta before the bifurcation of the iliac axis
Despite these recent advances, kidney regeneration still lags behind other organs in the field. Although effective protocols have been established for whole-organ kidney decellularization in pigs and rats, similar studies involving non-human primates have not been attempted. Non-human primate studies will become increasingly important as the field advances, as porcine kidneys show marked differences in vascular architecture [123]. Secondly, the attempted recellularization of higher-order mammalian kidneys has not been reported. It thus remains unclear whether larger kidneys can support the retrograde ureteric perfusion needed to recellularize the renal collecting system. These studies are currently underway at our institute. Additionally, the most effective seeding population remains to be determined, although we have reviewed several candidates (see Section 1). Likewise, it is unclear whether the decellularized kidney ECM can support the proliferation and differentiation of stem cells into the approximately 26 requisite cell types that comprise the mature human kidney [124]. Even after reliable recellularization protocols have been established, the homoeostatic, resorptive, metabolic, endocrine and immunomodulatory functions must be re-established by the regenerated kidney. Finally, it is unclear whether perfusion recellularization is sufficient to prevent the thrombogenic effects of the collagen scaffold observed upon re-implantation.
Conclusion
We have reviewed key aspects of RM technologies as they relate to the kidney. Advances in stem cell technology, renal replacement therapy and organ bioengineering and regeneration are intersecting with promise to resolve the dire shortage of transplantable organs. Despite steady progress, renal bioengineering lags behind other organs in the field due to the complex architecture and physiology of the native kidney and to the lack of in-depth knowledge on the interactions between ECM, cells and growth factors. While current research endeavours are promising, the transition to safe and effective clinical implementation faces significant obstacles. Collaborative efforts and investigation are required to drive the field towards the production of a bioengineered kidney capable of restoring renal function in patients with end-stage disease.
References
Papers of particular interest, published recently, have been highlighted as: • Of importance
Peralta CA et al. Control of hypertension in adults with chronic kidney disease in the United States. Hypertension. 2005;45(6):1119–24.
Sarafidis PA et al. Hypertension awareness, treatment, and control in chronic kidney disease. Am J Med. 2008;121(4):332–40.
Tonelli M et al. Chronic kidney disease and mortality risk: a systematic review. J Am Soc Nephrol. 2006;17(7):2034–47.
Guild WR et al. Successful homotransplantation of the kidney in an identical twin. Trans Am Clin Climatol Assoc. 1955;67:167–73.
Jassal SV et al. Kidney transplantation in the elderly: a decision analysis. J Am Soc Nephrol. 2003;14(1):187–96.
Laupacis A et al. A study of the quality of life and cost-utility of renal transplantation. Kidney Int. 1996;50(1):235–42.
Oliver J. Correlations of structure and function and mechanisms of recovery in acute tubular necrosis. Am J Med. 1953;15(4):535–57.
Cuppage FE, Tate A. Repair of the nephron following injury with mercuric chloride. Am J Pathol. 1967;51(3):405–29.
Haagsma BH, Pound AW. Mercuric chloride-induced tubulonecrosis in the rat kidney: the recovery phase. Br J Exp Pathol. 1980;61(3):229–41.
Nony P, Boissel JP. Use of sensitivity functions to characterise and compare the forgiveness of drugs. Clin Pharmacokinet. 2002;41(5):371–80.
Pawar S, Kartha S, Toback FG. Differential gene expression in migrating renal epithelial cells after wounding. J Cell Physiol. 1995;165(3):556–65.
Counts RS et al. Nephrotoxicant inhibition of renal proximal tubule cell regeneration. Am J Physiol. 1995;269(2 Pt 2):F274–81.
Kawaida K et al. Hepatocyte growth factor prevents acute renal failure and accelerates renal regeneration in mice. Proc Natl Acad Sci U S A. 1994;91(10):4357–61.
Ichimura T et al. FGF-1 in normal and regenerating kidney: expression in mononuclear, interstitial, and regenerating epithelial cells. Am J Physiol. 1995;269(5 Pt 2):F653–62.
Haq M et al. Role of IL-1 in renal ischemic reperfusion injury. J Am Soc Nephrol. 1998;9(4):614–9.
Nowak G, Schnellmann RG. Renal cell regeneration following oxidant exposure: inhibition by TGF-beta1 and stimulation by ascorbic acid. Toxicol Appl Pharmacol. 1997;145(1):175–83.
Grobstein C. Morphogenetic interaction between embryonic mouse tissues separated by a membrane filter. Nature. 1953;172(4384):869–70.
Grobstein C. Inductive epitheliomesenchymal interaction in cultured organ rudiments of the mouse. Science. 1953;118(3054):52–5.
Grobstein C. Trans-filter induction of tubules in mouse metanephrogenic mesenchyme. Exp Cell Res. 1956;10(2):424–40.
Grobstein C. Inductive tissue interaction in development. Adv Cancer Res. 1956;4:187–236.
Kitamura S et al. Establishment and characterization of renal progenitor like cells from S3 segment of nephron in rat adult kidney. FASEB J. 2005;19(13):1789–97.
Lindgren D et al. Isolation and characterization of progenitor-like cells from human renal proximal tubules. Am J Pathol. 2011;178(2):828–37.
Maeshima A, Yamashita S, Nojima Y. Identification of renal progenitor-like tubular cells that participate in the regeneration processes of the kidney. J Am Soc Nephrol. 2003;14(12):3138–46.
Sagrinati C et al. Isolation and characterization of multipotent progenitor cells from the Bowman's capsule of adult human kidneys. J Am Soc Nephrol. 2006;17(9):2443–56.
Oliver JA et al. The renal papilla is a niche for adult kidney stem cells. J Clin Invest. 2004;114(6):795–804.
Kobayashi A, Valerius MT, Mugford JW, et al. Pax2 maintains a selfrenewing nephron progenitor population by repressing interstitial cell fates during mammalian kidney development. San Diego: 2009 American Society of Nephrology Meeting; 2009.
Kobayashi A et al. Six2 defines and regulates a multipotent self-renewing nephron progenitor population throughout mammalian kidney development. Cell Stem Cell. 2008;3(2):169–81.
Self M et al. Six2 is required for suppression of nephrogenesis and progenitor renewal in the developing kidney. EMBO J. 2006;25(21):5214–28.
Bussolati B et al. Isolation of renal progenitor cells from adult human kidney. Am J Pathol. 2005;166(2):545–55.
Ronconi E et al. Regeneration of glomerular podocytes by human renal progenitors. J Am Soc Nephrol. 2009;20(2):322–32.
Martin GR. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. Proc Natl Acad Sci U S A. 1981;78(12):7634–8. First, groundbreaking report on the isolation of embryonic stem cells in mice. This paper came out almost at the same time as another paper from Evans and Kaufman..
Mae S et al. Combination of small molecules enhances differentiation of mouse embryonic stem cells into intermediate mesoderm through BMP7-positive cells. Biochem Biophys Res Commun. 2010;393(4):877–82.
Schuldiner M et al. Effects of eight growth factors on the differentiation of cells derived from human embryonic stem cells. Proc Natl Acad Sci U S A. 2000;97(21):11307–12.
Kim D, Dressler GR. Nephrogenic factors promote differentiation of mouse embryonic stem cells into renal epithelia. J Am Soc Nephrol. 2005;16(12):3527–34.
Vigneau C et al. Mouse embryonic stem cell-derived embryoid bodies generate progenitors that integrate long term into renal proximal tubules in vivo. J Am Soc Nephrol. 2007;18(6):1709–20.
Kobayashi T et al. Wnt4-transformed mouse embryonic stem cells differentiate into renal tubular cells. Biochem Biophys Res Commun. 2005;336(2):585–95.
Reubinoff BE et al. Embryonic stem cell lines from human blastocysts: somatic differentiation in vitro. Nat Biotechnol. 2000;18(4):399–404.
Steenhard BM et al. Integration of embryonic stem cells in metanephric kidney organ culture. J Am Soc Nephrol. 2005;16(6):1623–31.
Fang TC et al. Exogenous bone marrow cells do not rescue non-irradiated mice from acute renal tubular damage caused by HgCl2, despite establishment of chimaerism and cell proliferation in bone marrow and spleen. Cell Prolif. 2008;41(4):592–606.
Poulsom R et al. Bone marrow contributes to renal parenchymal turnover and regeneration. J Pathol. 2001;195(2):229–35.
Hayakawa M et al. Role of bone marrow cells in the healing process of mouse experimental glomerulonephritis. Pediatr Res. 2005;58(2):323–8.
Burdon TJ et al. Bone marrow stem cell derived paracrine factors for regenerative medicine: current perspectives and therapeutic potential. Bone Marrow Res. 2011;2011:207326.
Camussi G, Deregibus MC, Tetta C. Paracrine/endocrine mechanism of stem cells on kidney repair: role of microvesicle-mediated transfer of genetic information. Curr Opin Nephrol Hypertens. 2010;19(1):7–12.
He J et al. Bone marrow stem cells-derived microvesicles protect against renal injury in the mouse remnant kidney model. Nephrology (Carlton). 2012;17(5):493–500.
Ikarashi K et al. Bone marrow cells contribute to regeneration of damaged glomerular endothelial cells. Kidney Int. 2005;67(5):1925–33.
Ito T et al. Bone marrow is a reservoir of repopulating mesangial cells during glomerular remodeling. J Am Soc Nephrol. 2001;12(12):2625–35.
Li J et al. The contribution of bone marrow-derived cells to the development of renal interstitial fibrosis. Stem Cells. 2007;25(3):697–706.
Lindoso RS et al. Paracrine interaction between bone marrow-derived stem cells and renal epithelial cells. Cell Physiol Biochem. 2011;28(2):267–78.
Masuya M et al. Hematopoietic origin of glomerular mesangial cells. Blood. 2003;101(6):2215–8.
McTaggart SJ, Atkinson K. Mesenchymal stem cells: immunobiology and therapeutic potential in kidney disease. Nephrology (Carlton). 2007;12(1):44–52.
Perry J et al. Type IV collagen induces podocytic features in bone marrow stromal stem cells in vitro. J Am Soc Nephrol. 2006;17(1):66–76.
Prodromidi EI et al. Bone marrow-derived cells contribute to podocyte regeneration and amelioration of renal disease in a mouse model of Alport syndrome. Stem Cells. 2006;24(11):2448–55.
Qian H et al. Bone marrow mesenchymal stem cells ameliorate rat acute renal failure by differentiation into renal tubular epithelial-like cells. Int J Mol Med. 2008;22(3):325–32.
Reis LA et al. Bone marrow-derived mesenchymal stem cells repaired but did not prevent gentamicin-induced acute kidney injury through paracrine effects in rats. PLoS One. 2012;7(9):e44092.
Rookmaaker MB et al. Bone-marrow-derived cells contribute to glomerular endothelial repair in experimental glomerulonephritis. Am J Pathol. 2003;163(2):553–62.
Sugimoto H et al. Bone-marrow-derived stem cells repair basement membrane collagen defects and reverse genetic kidney disease. Proc Natl Acad Sci U S A. 2006;103(19):7321–6.
Suzuki A et al. Platelet-derived growth factor plays a critical role to convert bone marrow cells into glomerular mesangial-like cells. Kidney Int. 2004;65(1):15–24.
Togel F et al. VEGF is a mediator of the renoprotective effects of multipotent marrow stromal cells in acute kidney injury. J Cell Mol Med. 2009;13(8B):2109–14.
Yadav N et al. Bone marrow cells contribute to tubular epithelium regeneration following acute kidney injury induced by mercuric chloride. Indian J Med Res. 2012;136(2):211–20.
Perin L et al. Renal differentiation of amniotic fluid stem cells. Cell Prolif. 2007;40(6):936–48.
Rota C et al. Human amniotic fluid stem cell preconditioning improves their regenerative potential. Stem Cells Dev. 2012;21(11):1911–23.
Siegel N et al. Contribution of human amniotic fluid stem cells to renal tissue formation depends on mTOR. Hum Mol Genet. 2010;19(17):3320–31.
Hauser PV et al. Stem cells derived from human amniotic fluid contribute to acute kidney injury recovery. Am J Pathol. 2010;177(4):2011–21.
Polo JM et al. Cell type of origin influences the molecular and functional properties of mouse induced pluripotent stem cells. Nat Biotechnol. 2010;28(8):848–55.
Song B et al. Generation of induced pluripotent stem cells from human kidney mesangial cells. J Am Soc Nephrol. 2011;22(7):1213–20.
Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006;126(4):663–76.
Zhao T et al. Immunogenicity of induced pluripotent stem cells. Nature. 2011;474(7350):212–5.
Zhou T et al. Generation of induced pluripotent stem cells from urine. J Am Soc Nephrol. 2011;22(7):1221–8.
Bratt-Leal AM, Carpenedo RL, McDevitt TC. Engineering the embryoid body microenvironment to direct embryonic stem cell differentiation. Biotechnol Prog. 2009;25(1):43–51.
Blum B et al. The anti-apoptotic gene survivin contributes to teratoma formation by human embryonic stem cells. Nat Biotechnol. 2009;27(3):281–7.
Phinney DG, Prockop DJ. Concise review: mesenchymal stem/multipotent stromal cells: the state of transdifferentiation and modes of tissue repair–current views. Stem Cells. 2007;25(11):2896–902.
Thirabanjasak D, Tantiwongse K, Thorner PS. Angiomyeloproliferative lesions following autologous stem cell therapy. J Am Soc Nephrol. 2010;21(7):1218–22.
Woolf AS et al. Creation of a functioning chimeric mammalian kidney. Kidney Int. 1990;38(5):991–7.
Rogers SA et al. Transplantation of developing metanephroi into adult rats. Kidney Int. 1998;54(1):27–37.
Statter MB et al. Correlation of fetal kidney and testis congenic graft survival with reduced major histocompatibility complex burden. Transplantation. 1989;47(4):651–60.
Abrahamson DR et al. Glomerular development in intraocular and intrarenal grafts of fetal kidneys. Lab Invest. 1991;64(5):629–39.
Samstein B, Platt JL. Physiologic and immunologic hurdles to xenotransplantation. J Am Soc Nephrol. 2001;12(1):182–93.
Dai Y et al. Targeted disruption of the alpha1,3-galactosyltransferase gene in cloned pigs. Nat Biotechnol. 2002;20(3):251–5.
Cascalho M, Platt JL. Xenotransplantation and other means of organ replacement. Nat Rev Immunol. 2001;1(2):154–60.
Rogers SA, Talcott M, Hammerman MR. Transplantation of pig metanephroi. ASAIO J. 2003;49(1):48–52.
Dekel B et al. Engraftment and differentiation of human metanephroi into functional mature nephrons after transplantation into mice is accompanied by a profile of gene expression similar to normal human kidney development. J Am Soc Nephrol. 2002;13(4):977–90.
Hammerman MR. Renal organogenesis from transplanted metanephric primordia. J Am Soc Nephrol. 2004;15(5):1126–32.
Steer DL et al. A strategy for in vitro propagation of rat nephrons. Kidney Int. 2002;62(6):1958–65.
Rosines E et al. Constructing kidney-like tissues from cells based on programs for organ development: toward a method of in vitro tissue engineering of the kidney. Tissue Eng Part A. 2010;16(8):2441–55. Pioneering paper in which renal bioengineering is attempted through a developmental biology approach in which the ureteric bud is manipulated to obtain kidney-like structures..
Xinaris C et al. In vivo maturation of functional renal organoids formed from embryonic cell suspensions. J Am Soc Nephrol. 2012;23(11):1857–68.
Tasnim F et al. Achievements and challenges in bioartificial kidney development. Fibrogenesis Tissue Repair. 2010;3:14.
Aebischer P et al. The bioartificial kidney: progress towards an ultrafiltration device with renal epithelial cells processing. Life Support Syst. 1987;5(2):159–68.
Humes HD et al. Initial clinical results of the bioartificial kidney containing human cells in ICU patients with acute renal failure. Kidney Int. 2004;66(4):1578–88.
Tumlin J et al. Efficacy and safety of renal tubule cell therapy for acute renal failure. J Am Soc Nephrol. 2008;19(5):1034–40.
Bissell MJ, Hall HG, Parry G. How does the extracellular matrix direct gene expression? J Theor Biol. 1982;99(1):31–68.
Lelongt B, Ronco P. Role of extracellular matrix in kidney development and repair. Pediatr Nephrol. 2003;18(8):731–42.
Muller U et al. Integrin alpha8beta1 is critically important for epithelial-mesenchymal interactions during kidney morphogenesis. Cell. 1997;88(5):603–13.
Mason RM, Wahab NA. Extracellular matrix metabolism in diabetic nephropathy. J Am Soc Nephrol. 2003;14(5):1358–73.
Song JJ, Ott HC. Organ engineering based on decellularized matrix scaffolds. Trends Mol Med. 2011;17(8):424–32.
Orlando G et al. Regenerative medicine as applied to solid organ transplantation: current status and future challenges. Transpl Int. 2011;24(3):223–32.
Orlando G et al. Regenerative medicine and organ transplantation: past, present, and future. Transplantation. 2011;91(12):1310–7.
Orlando G et al. Regenerative medicine as applied to general surgery. Ann Surg. 2012;255(5):867–80.
Badylak SF et al. Engineered whole organs and complex tissues. Lancet. 2012;379(9819):943–52. Seminal review on the cell-scaffold technology which seems to offer the most promising approach to renal bioengineering, while granting the quickest route to clinical application..
Orlando G et al. Production and implantation of renal extracellular matrix scaffolds from porcine kidneys as a platform for renal bioengineering investigations. Ann Surg. 2012;256(2):363–70. Seminal study providing evidence that porcine kidneys can be decellularized to produce acellular scaffolds. Such scaffolds represent an ideal platform for renal bioengineering investigations..
Wang Y et al. Lineage restriction of human hepatic stem cells to mature fates is made efficient by tissue-specific biomatrix scaffolds. Hepatology. 2011;53(1):293–305.
Ross EA et al. Embryonic stem cells proliferate and differentiate when seeded into kidney scaffolds. J Am Soc Nephrol. 2009;20(11):2338–47.
Ng SL et al. Lineage restricted progenitors for the repopulation of decellularized heart. Biomaterials. 2011;32(30):7571–80.
Hynes RO. The extracellular matrix: not just pretty fibrils. Science. 2009;326(5957):1216–9.
Badylak SF, Taylor D, Uygun K. Whole-organ tissue engineering: decellularization and recellularization of three-dimensional matrix scaffolds. Annu Rev Biomed Eng. 2011;13:27–53.
Gilbert TW, Sellaro TL, Badylak SF. Decellularization of tissues and organs. Biomaterials. 2006;27(19):3675–83.
Crapo PM et al. Biologic scaffolds composed of central nervous system extracellular matrix. Biomaterials. 2012;33(13):3539–47.
Badylak SF, Gilbert TW. Immune response to biologic scaffold materials. Semin Immunol. 2008;20(2):109–16.
Brown BN et al. Macrophage phenotype and remodeling outcomes in response to biologic scaffolds with and without a cellular component. Biomaterials. 2009;30(8):1482–91.
Reing JE et al. The effects of processing methods upon mechanical and biologic properties of porcine dermal extracellular matrix scaffolds. Biomaterials. 2010;31(33):8626–33.
Brown B et al. The basement membrane component of biologic scaffolds derived from extracellular matrix. Tissue Eng. 2006;12(3):519–26.
Baptista PM et al. The use of whole organ decellularization for the generation of a vascularized liver organoid. Hepatology. 2011;53(2):604–17.
Song JJ et al. Enhanced in vivo function of bioartificial lungs in rats. Ann Thorac Surg. 2011;92(3):998–1005. discussion 1005–6.
Martinello, T., et al., Successful recellularization of human tendon scaffolds using adipose-derived mesenchymal stem cells and collagen gel. J Tissue Eng Regen Med, 2012
Honge JL et al. Recellularization of aortic valves in pigs. Eur J Cardiothorac Surg. 2011;39(6):829–34.
Loai Y et al. Bladder tissue engineering: tissue regeneration and neovascularization of HA-VEGF-incorporated bladder acellular constructs in mouse and porcine animal models. J Biomed Mater Res A. 2010;94(4):1205–15.
Wicha MS et al. Extracellular matrix promotes mammary epithelial growth and differentiation in vitro. Proc Natl Acad Sci U S A. 1982;79(10):3213–7.
Liu CX et al. Preparation of whole-kidney acellular matrix in rats by perfusion. Nan Fang Yi Ke Da Xue Xue Bao. 2009;29(5):979–82.
Nakayama KH et al. Decellularized rhesus monkey kidney as a three-dimensional scaffold for renal tissue engineering. Tissue Eng Part A. 2010;16(7):2207–16.
Sullivan DC et al. Decellularization methods of porcine kidneys for whole organ engineering using a high-throughput system. Biomaterials. 2012;33(31):7756–64.
Narlis M et al. Pax2 and pax8 regulate branching morphogenesis and nephron differentiation in the developing kidney. J Am Soc Nephrol. 2007;18(4):1121–9.
Shao X et al. A minimal Ksp-cadherin promoter linked to a green fluorescent protein reporter gene exhibits tissue-specific expression in the developing kidney and genitourinary tract. J Am Soc Nephrol. 2002;13(7):1824–36.
Gupta S et al. Isolation and characterization of kidney-derived stem cells. J Am Soc Nephrol. 2006;17(11):3028–40.
Bagetti Filho HJ et al. Pig kidney: anatomical relationships between the renal venous arrangement and the kidney collecting system. J Urol. 2008;179(4):1627–30.
Al-Awqati Q, Oliver JA. Stem cells in the kidney. Kidney Int. 2002;61(2):387–95.
Compliance with Ethics Guidelines
ᅟ
Conflict of Interest
Dr. Marcus Salvatori, Dr. Andrea Peloso, Dr. Ravi Katari, and Dr. Giuseppe Orlando reported no potential conflicts of interest relevant to this article.
Human and Animal Rights and Informed Consent
This article does not contain any studies with human or animal subjects performed by any of the authors.
Author information
Authors and Affiliations
Corresponding author
Additional information
This article is part of the Topical Collection on Kidney Diseases
Rights and permissions
About this article
Cite this article
Salvatori, M., Peloso, A., Katari, R. et al. Regeneration and Bioengineering of the Kidney: Current Status and Future Challenges. Curr Urol Rep 15, 379 (2014). https://doi.org/10.1007/s11934-013-0379-9
Published:
DOI: https://doi.org/10.1007/s11934-013-0379-9
Keywords
- CKD Chronic Kidney Disease
- ESRD End-Stage Renal Disease
- RM Regenerative Medicine
- ESC Embryonic Stem Cell
- BMDC Bone Marrow-Derived Stem Cell
- MSC Mesenchymal Stem Cell
- Hafsc Human Amniotic Fluid-Derived Stem Cells
- IPSC Induced Pluripotent Stem Cells
- BAK Bio-Artificial Kidney
- RAD Renal Tubule Assisted Device
- ARF Acute Renal Failure
- ECM Extracellular Matrix
- DRT Decellularization-Recellularization Technology