
Abstract
This study characterized the benthic algal communities at the mouth of a large eutrophic-hypertrophic river, specifically during the vegetation period of 2019 and the summer-autumn period of 2020. Emphasis was placed on the composition, taxonomic structure, and the seasonal and interannual variability of abundance and diversity indicators. Detailed analysis was conducted on communities from bottom sediments, rocky substrates, and higher aquatic and coastal-water plants. Results show significant variations in composition and taxonomic structure across different substrate types and time frames. The dominant species complexes exhibited both temporal and substrate heterogeneity. Key findings highlight the importance of considering various substrate types and time periods, and they underline the observed changes in taxonomic group abundance. Such insights are crucial for comprehensive assessments of benthic algae communities in large eutrophic-hypertrophic rivers.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Benthic algal communities (microphytobenthos and periphyton) are an integral component of aquatic ecosystems and take an active part in the cycle of organic matter and biogenic elements, retaining various chemical components of water masses in the benthic zone as a result of various physical and chemical and biological processes. Benthic algae can account for up to 50% of the total primary production of a water mass and play an important role in both benthic and pelagic trophic nets, constituting a significant nourishing source for benthic animals and detritivores that feed on bottom sediments (Montagna et al. 1995; Perissinotto et al. 2002). Periphyton can remove nutrients, toxic components and heavy metals from open water (Vymazal 1988; Hill et al. 2000) and is widely used in ecotoxicological studies and bioindication (Khedairia and Okhapkin 2021).
The communities of benthic algae respond quickly and predictably, to changes in the composition and amount of nutrients in water masses; their composition and structure analysis helps to identify key factors that affect the organization and ecology of communities of water masses. Structural and functional characteristics of benthic algal communities can be used as a preliminary indicator of abrupt changes in the ecosystem (Gaiser et al. 2005), disturbances and deterioration of conditions in continental waters (Wu 2016).
Until the mid-20th century, most research were focused on the role of biotic components such as macrophytes, plankton (zooplankton and phytoplankton) and invertebrates (benthos, nekton and neuston) in the aquatic nutrient cycle. While microphytobenthos and phytoperiphyton have not been given sufficient attention due to their role in aquatic ecosystems. In 1963, Wetzel, in his fundamental review, empathized the importance of periphyton in aquatic ecosystems as a complexly organized component in the system of primary producers.
The Oka River is one of the largest rivers in the Russian Federation; it has a significant impact on the Volga River downstream of its confluence. It has immense economic importance and plays a crucial role in the country infrastructure. The length of the river reaches more than 1400 km, the basin area on the territory of 7 regions of the European part of Russia is 245 000 km2 (Tockner et al. 2022). On the banks of the mouth area of the river is the largest city of the Volga region located – Nizhny Novgorod (1.3 million inhabitants), above the mouth area there are a number of cities with a total population of more than 0.3 million people. The river catchment suffers from a significant anthropogenic load, its waters are heavily polluted with biogenic and organic substances, heavy metals and oil products (Okhapkin et al. 2015; Dzhamalov et al. 2017).
Confluence areas of large rivers significantly affect the local ecology while changing water temperature, quality and quantity of suspended sediments, bottom sediments, nutrient concentrations, water chemistry and organic matter content (Zheng et al. 2020; Yuan et al. 2022). They are changing the habitat conditions of fish due to unique distribution characteristics of hydrodynamic parameters and nutrients (Shen et al. 2022).
Microphytobenthic communities of the largest watercourse in the European part of Russia – the Oka River – is still being only in the first stages of study, as the focus of algal research was focused on phytoplankton (Tockner et al. 2022). For the estuarine section of the river, there is information given about diatom microphytobenthos of bottom sediments, periphyton of stony substrate and higher aquatic and coastal aquatic plants and their indicator properties as indicators of water quality; spatial distribution of indicators of the structure of benthic algae communities in the period after the recession of the flood (Khedairia 2021; Khedairia and Okhapkin 2021; Khedairia et al. 2022a, b). The dynamics of the composition, abundance and general organization of benthic algae communities in seasonal and interannual aspects in connection with environmental factors have not been studied.
Purpose of the research
To characterize the species composition, taxonomic and ecological structure, changes in the complex of dominant species, indicators of abundance, and species diversity of the cyanobacteria and algae communities in the benthic zone, and to determine the seasonal and interannual variations of these parameters in the coastal area of the eutrophic-hypertrophic Okа River estuary.
Materials and methods
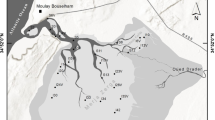
Samples of algal communities from the benthic zone of the Oka River mouth near Nizhny Novgorod were collected monthly from May to October 2019, in summer and autumn (August – October) 2020 (68 quantitative samples). The material was collected at one station located on the right bank of the river above the Kanavinsky bridge (56.301329° N, 43.966726° E) (Fig. 1) simultaneously with chemical analysis of water by the Upper Volga Department for Hydrometeorology and Control of Natural environment (Table 1). Samples were collected from the surface of bottom sediments, specifically epipelon, which consisted of silts and heavily silted sands mixed with a rocky substrate (epilithon) primarily composed of limestones. The aquatic and semiaquatic vegetation (epiphyton) found in the area showed limited growth during the spring season, mainly consisting of scattered instances of arrowhead plants, a few water lilies, and pondweed individuals. The methods for collecting benthic algae followed those prescribed by using the accepted methodological approaches (Kholodnoho and Vasser 1989; Komulainen 2003; Nevrova et al. 2015; Okhapkin et al. 2019). Epilithic algae were removed with a scalpel from a measured area marked on the surface of each stone. The scalpel was used because some stones contained encrusted growths of algae due to the hardness of the river. The sample area on each stone was also brushed vigorously with a toothbrush in a plastic tray with water to remove any remaining algal film, and the sample was then poured into labeled plastic vials. Regarding epiphytic algae, the surface of submerged macrophytes underwent gentle brushing with a toothbrush while being submerged in a cuvette filled with distilled water. Subsequently, the resulting algal suspensions were carefully poured into labeled vials. Samples of the bottom sediments (epipelon) were taken using a plastic tube with a capture area of 14.5 cm2. The tube was inserted into a column of bottom sediments collected using a Petersen grab, and a surface layer 5–7 mm thick was extracted. The collected material was placed in a labeled vial. At least three samples were taken for each substrate, followed by their microscopy in a living and fixed state (iodine-formalin fixative) under a Meiji Techno light microscope (Japan) at a 1000x magnification using immersion oil. Samples were taken in the shallow coastal zone at a depth of (40–70 cm) of the Secchi disk transparency value. Environmental parameters were recorded on-site, with water temperatures ranging from 9.4 to 24.1 °C across different seasons, and electrical conductivity fluctuating between 532 and 583 µS/cm. The pH levels, indicating the active environmental response, were found to be between 8.1 and 9.2. Diatom species were identified through light microscopy and scanning electron microscopy (utilizing the JSM-IT300LV microscope from JEOL, Japan). For this process, diatoms were treated with hot oxidation using hydrogen peroxide. Permanent slides were prepared using Naphrax medium (refractive index 1.74, Brunel Microscopes Ltd). Guidelines for identifying the species composition of algae have been described earlier (Okhapkin 1998; Nevrova et al. 2015), the names of corresponding systematic groups of algae are given according to Algabase (Guiry and Guiry 2019).

Schematic map of the mouth of the Oka River. (st) sampling station
The abundance (N) of phytobenthos and the predominant components of the benthic algal communities were assessed using an Uchinskaya camera (Najotte counting chamber) with a volume of 0.01 cm³. The samples contained high amounts of organic and mineral particles, which could interfere with the identification process. To mitigate this issue and improve the visibility of the algal cells, samples with high levels of particles were diluted. In each sample, a minimum of 400 cells were counted. The results were expressed in millions of cells per 10 cm², calculated using the following formula:
where A is the total number of counted cells, n is the number of calculated bands of the chamber, V0 is the initial volume of a sample of algae (ml), V1 is a volume of the examined sample, and S is the total substrate surface from which the sample was taken.
Biomass (B, mg/10 cm2) was determined by the volumetric counting method in accordance with published recommendations (Komulainen 2003; Meteleva 2013). The average biovolume for each species in every sample was determined by measuring the dimensions of 20–30 cells and applying the geometrical equations suggested by Hillebrand et al. (1999) to calculate their cell volume. Taxa whose biomass or abundance were counted as ≥ 10% of the total values were considered dominant. For them, the occurrence frequency and dominance frequency were determined. The species diversity of communities was assessed using the Shannon – Weaver index (Magurran 2004).
To ensure accuracy and reliability, a range of statistical methods and software tools were applied to analyze and interpret the data. The data analysis was conducted in the R environment (Team R 2022), which is an open-source software environment for statistical computing and modeling. Standardization of the data was carried out using the “hell” method of the “decostand” function, a unique feature that emphasizes the need for careful review and interpretation of the results.
To classify the communities, the species variety of the benthic algal communities of the Oka River mouth was subjected to a cluster analysis using the appropriate Bray Curtis index (qualitative for presence/absence) by the Ward method. The results are presented in the form of ordination diagrams (PCA, principal component analysis). Differences between communities were tested for significance using permutation and multivariate analysis of variance, with distance matrices being utilized for this purpose. The functions “adonis2” and “pairwise. adonis2” were employed to compare species variety of differences between communities.
To explore the effects of substrate and season on the biological indicators and diversity indexes, a two-way analysis of variance was conducted. The model reliability was checked by performing an ANOVA test using the “anova()” function, and a TukeyHSD test was used to compare group means via the “TukeyHSD()” function.
Additionally, the hydrochemical composition data of the waters of the Oka River, provided by the Upper Volga Department for Hydrometeorology and Environmental Control, were analyzed to characterize the abiotic habitat conditions of benthic communities. The coefficient of variation (CV) was used in this study as a measure of relative variability. Calculated by dividing the standard deviation by the mean and expressing it as a percentage, the CV was computed using Microsoft Excel software. This analysis provided insights into the relationships between the different variables in the study and helped to better understand the variation in the data.
The Oka River, the second largest tributary of the Volga in terms of the channel size and the runoff volume over the past decades has had a stable low water quality, estimated as class 4. As mentionned before (Okhapkin et al. 2015; Dzhamalov et al. 2017; Tockner et al. 2022), the waters of its mouth section were characterized by high turbidity, high concentrations of nitrogen and phosphorus in mineral form, organic substances, heavy metals and other pollution components.
Results
Species composition of benthic algoflora
In the benthic algal communities studied, 163 taxa of algae (species, including the nomenclatural type of species, varieties, and forms, and defined only to the genus, IT), these taxa spanned 4 phyla, 68 genera were identified. An analysis of the taxa numbers on different substrates and in different periods of the hydrological cycle showed the predominance of Bacillariophyta (diatoms), which were the most numerous phyla in terms of species variety: 87 IT (53.87%) of the total list. They were followed by green algae (67 taxa, 41.10%) and cyanobacteria, (7 species, 4.29%) mostly filamentous. Euglenoids are presented singly – 1.2%. Thus, the Oka algoflora of microphytobenthos on the bottom sediments, fouling of stony substrate, and higher aquatic and coastal aquatic plants during the study period can be characterized as diatom green algae with an insignificant participation of cyanoprokaryotes.
The composition of the epilithon community (117 taxa) turned out to be somewhat richer than that of the epiphyton (106 species) and epipelon (105 species). The floristic similarity analysis of different types of algocenose substrates (Fig. 2) showed that three large groups are clearly distinguished according to the type of substrate (Permanova, F value = 3.91, P < 0.001). This picture emerged at the level of the overall species composition of algae and the list of dominant taxa (Fig. 2, a–e), and the composition of diatoms showed noticeable differences between epipelon and periphyton (Fig. 2, b) (epilithon and epiphyton) (Permanova, F value = 4.10, P < 0.001), while between the periphyton communities (epilithon and epiphyton), this difference was not recorded (Permanova, P > 0.05). For the list of Chlorophyta (Fig. 2, c), a great similarity between all types of communities was found (Permanova, P > 0.05). The distribution of the composition of the dominant species and the composition of cyanobacteria corresponded to the type of substrate (Fig. 2, d–e) (Permanova, F value = 12.58, P < 0.001; F value = 4.84, P < 0.01).

Ordination diagram of species composition of benthic communities, a all the algae classes, b Bacillariophyta, c Chlorophyta, d Cyanophyta, e Dominants species
On a time, scale, the specific species variety (the number of species in a sample) varied from 10 to 40 IT, the smallest number of species was noted in May 2019, August and September 2020 on macrophytes (10 species), and at other times the number of species on different substrates was comparable. The mean species richness of all types of benthic algae communities differed significantly in different months (two.way ANOVA, F value = 2.02, P < 0.05). At the same time, more noticeable seasonal differences in the species richness of periphyton communities were established (plants – two.way ANOVA, F value = 5.85, P < 0.001; stones – F value = 4.09, P < 0.05) with the similarity of the overall curve of its dynamics within the growing season with a rise in late summer – early autumn (August – September). For the late summer – early autumn of 2019, the algal abundance of the periphyton communities was significantly higher than in the autumn of 2020 (TukeyHSD, two.way P < 0.001), and for the epipelon communities it turned out to be almost the same over the entire study period (two.way ANOVA, P > 0.05).
Complex of dominant species
35 taxa of algae (or 21.5% of quantitative samples) (Table 2) (Fig. 3), were assigned as prevailing species in terms of abundance and biomass. As for assessing dominance by biomass, diatoms are more diverse (75.8% of the list of all dominants), and the role of Cyanobacteria is more noticeable in the formation of abundance (17.1%); only four species (11%) were noted among Chlorophyta. The number of dominant microphytobenthos in bottom sediments (20 IT) did not differ from other types of communities and representatives of benthic and periphyton cenoses (Cocconeis pediculus Ehrenberg, C. placentula Ehrenberg, Gomphonella olivacea (Hornemann) Rabenhorst, Surirella librile (Ehrenberg) Ehrenberg. and Cymbella spp.) including potamoplankton components (various Stephanodiscus, Cyclotella and Skeletonema). Planktic-benthic species (Melosira varians C. Agardh, Merismopedia tenuissima Lemmermann, Oscillatoria simplicissima Gomont and Monoraphidium contortum (Thuret) Komárková-Legnerová) are sporadically noted among the dominants of the epipelon. The maximum occurrence and frequency of dominance are typical for planktic forms (mainly Stephanodiscus hantzschii Grunow) and foulers (Cocconeis spp.). The components of the benthic assemblage itself (species of the genera Cymatopleura, Cymbella, Fragilaria, Gomphonema, Leptolyngbya and Phormidium) developed less frequently and with lower or insignificant dominance and abundance indicators.

Predominant species of Bacillariophyta. a Nitzschia dissipata, b Navicula cryptotonella, c Navicula tripunctata, d N. capitatoriata, e Cocconeis placentula, f Encyonema cespitosum Kütz, g Diatoma vulgaris Bory, h Surirella librile, i Aulacoseira ambiguaj Stephanodiscus neoastraea and k various diatom species
The composition of the dominant epilithon species (19 taxa) (Table 2) is characterized by representatives of the genus Navicula (N. tripunctata (OF Müller) Bory, N. cryptotenella Lange-Bertalot, N. cryptocephala Kützing and N. capitatoradiata H.Germain ex Gasse, Nitzschia dissipata (Kütz.) Rabenhorst, Gomphonella olivacea and Oscillatoria simplicissima) Other cyanoprokaryotes (Phormidium breve (Kützing ex Gomont) Anagnostidis & Komárek, Leptolyngbya foveolarum (Gomont) Anagnostidis & Komárek and Limnococcus limneticus (Lemmermann) Komárková, Jezberová, O.Komárek & Zapomelová. predominated only in terms of cell number. The dominant composition of epiphyton complexes (20 taxa) was formed by foulers (Cocconeis placentula., Gomphonella olivacea and G. parvulum Kütz., less frequently – Cocconeis pediculus), plankton-benthic forms (Ulnaria ulna), green algae (Chlamydopodium pluricoccum (Korshikov) Ettl & Komárek – by biomass, Coelastrum microporum Nägeli – by abundance). Cyanoprokaryotes (Merismopedia tenuissima, Oscillatoria simplicissima, Phormidium breve, and Leptolyngbya foveolarum) are less common among the dominant species.
The highest frequencies of occurrence were found in the planktic species Stephanodiscus hantzschii (average for all types of substrates 92%) and planktic-benthic Cocconeis placentula (91%). They developed on all substrates, with the former almost always dominating on bottom sediments (94%), and to a lesser extent on stones (39%) and plants (18%). For Cocconeis placentula an opposite trend was observed: on plants, it prevailed in 82% of samples; on stones and bottom sediments, the frequency of its dominance is much lower (11–12%). Species of the genus Navicula are characterized by the presence and predominance of epilithon communities in the samples (Navicula cryptotenella occurrence frequency is measured as 100% and dominance – as 39%, N. tripunctata as 85 and 28%, respectively). For phytogrowths, these species are also typical components of cenoses (89–94% of findings), but prevailed only in 6–12% of samples. Filamentous cyanoprokaryotes, including motile ones (Oscillatoria simplicissima, Leptolyngbya foveolarum) were present in more than 33% of the samples and often created an aspect of epilithon communities and fouling of aquatic plants (Table 2).
The structure of the abundance and biomass of benthic communities
Abundance and biomass of benthic communities in the mouth area of the Oka River on various substrates (Fig. 4) varied (N – from 0.08 to 4360 million cells/10 cm2; B – from 0.01 to 145.3 mg/10 cm2) with significant differences in the seasonal aspect (N – F value = 2.80, P < 0.01; B – two.way ANOVA, F value = 3.98, P < 0.001). The highest mean values of biomass are typical for the epilithon, the lowest for the epiphyton (two.way ANOVA, P < 0.01). The development rate of benthic algae and cyanobacteria in the epipelon, estimated from the biomass, was on average almost the same as in the epilithon (two.way ANOVA, P > 0.05).

Mean abundance (million cells/10 cm2) (a, b and c) and biomass, mg/10 cm2 (d, e and f) of communities in the benthic zone. (a, d) Epipelon, (b, e) Epilithon, and (c, f) Epiphyton
Changes in the average density and biomass of benthic algae communities in different months of the study on different substrates were approximately the same (Fig. 4). In epipelon communities in late spring and early summer 2019, when the water warmed up to (17.5 °C), the indicators of the abundance of cenoses changed insignificantly (Fig. 4, a–d). At the same time, more than 80% of the biomass of the communities is formed by components of potamoplankton (mainly Stephanodiscus hantzschii), and less to planktic-benthic and true benthic species (Melosira varians, Surirella librile). In summer, with an increase in water temperature (23.5 °C), the abundance of communities increased markedly, especially in August due to the vegetation of diatoms (species of the genus Cocconeis, Gomphonema, Cymbella by biomass) and Cyanobacteria (Leptolyngbya foveolarum, Phormidium breve by number of cells). The basis of communities was formed by true benthic and plankton-benthic components (more than 80% of the biomass). In autumn, the vegetation intensity of microphytobenthos decreased markedly with the predominance of Oscillatoria simplicissima, Leptolyngbya foveolarum and Fragilaria aequalis Heiberg Østrup, as well as Cocconeis placentula, Surirella librile and Stephanodiscus hantzschii.
The second half of the growing season in 2020 was characterized by a slight increase in abundance and biomass from late summer to early autumn and a noticeable decrease in October (N – F value = 16.69, P < 0.001; B – two.way ANOVA, F value = 8.64, P < 0.01). The period from August to October was characterized by a decrease in water temperature from 19.8 °C to 13.9 °C and, according to the Upper Volga UGCS, was characterized by cool weather with a noticeable amount of precipitation. This was reflected in the predominance of the planktic complex of diatoms (Stephanodiscus hantzschi, S. neoastraea Håkansson & Hickel., Stephanocyclus meneghinianus (Kützing) Kulikovskiy, Genkal & Kociolek. form 80–85% of the biomass), the absence of cyanoprokaryotes in the list of dominant benthic biocenosis and the insignificant development of Cocconeis, Fragilaria and Cymatopleura.
For epilithon communities, the maximum value of abundance and biomass (Fig. 4, b–e) was recorded in summer, in July 2019, then they gradually decreased with a minimum in October. In 2020, higher levels of algae abundance were noted in summer than in autumn, but their decrease on the stony substrate was faster (already in September) than in bottom sediments community (N – F value = 5.51, P < 0.05; B – two.way ANOVA, F value = 4.62, P < 0.05). At the same time, the influence of the planktic complex of algae on the indexes of epilithon abundance turned out to be much less (from 5 to 65% of the biomass) than that of the epipelon and was more clearly manifested in spring and autumn. The role of true benthic species is much more pronounced than in epipelon community (10–90% of biomass).
The may cenoses of fouling of the stony substrate were formed by diatoms (Cocconeis placentula, Navicula cryptotenella, Ulnaria ulna (Nitzsch) Compère, Gomphonella olivacea) and to a lesser extent cyanoprokaryotes (Oscillatoria simplicissima). In summer, massive vegetation of cyanoprokaryotes (besides the above, various Phormidium and Leptolyngbya) was noted on the stones, which formed the basis of the abundance and more than half of the biomass of the communities. The set of dominants of the summer epilithon in 2019 is diverse due to diatoms (except for the May components – Navicula tripunctata, N. capitatorradiata, Cocconeis pediculus, much less frequently – Stephanodiscus hantzschii). In autumn, under the dominance of the Navicula spp. complex – Cocconeis pediculus, among diatoms – Amphora copulata (Kützing) Schoeman & R.E.M.Archibald., A. pediculus (Kützing) Grunow and Nitzschia dissipata. Algocenoces in summer – autumn 2020 are poorer in composition of dominants and are formed mainly by the same species as in the corresponding months of 2019, but with a smaller participation of cyanoprokaryotes.
Epiphyton community showed similar dynamics of abundance and biomass (Fig. 4, c f), but their growth began earlier than the previous ones and was noted in late spring – early summer. Compared to other substrates, phytoperiphyton on macrophytes was relatively lower in biomass (N – F value = 5.98, P < 0.001; B – two.way ANOVA, F value = 5.39, P < 0.001).
In spring, at the juvenile stage of development of aquatic plants, the epiphyton is poorly developed, and among the predominant species, except for diatoms (Melosira varians, Cocconeis placentula, Navicula cryptotenella) green algae (Chlamydopodium pluricoccum, Lemmermannia triangularis (Chodat) C.Bock & Krienitz and cyanobacteria (Oscillatoria simplicissima). The rate of algae ecological groups in cenoses is approximately the same as in epilithon communities. In summer, communities are formed by diatoms (in addition to spring forms, there are also Gomphonella olivacea, Cocconeis pediculus, Navicula cryptocephala presented), including planktic forms (Aulacoseira ambigua (Grunow) Simonsen, Stephanodiscus hantzschii). The number of Cyanobacteria has sharply increased, reaching a maximum in July, as well as their species variety (Oscillatoria simplicissima, Phormidium breve, Merismopedia tenuissima and Aphanocapsa delicatissima West & G.S.West). The relative abundance of benthic forms after the June maximum by the end of the summer season has decreased due to the greater contribution of eurytopic planktic -benthic representatives. A similar picture was also observed in the summer of 2020. At the beginning of autumn, the importance of the planktic components of benthic biocenosis (Stephanodiscus hantzschii) became more noticeable than in spring and summer (more than 40% of the total biomass), the abundance of foulers and true benthic algae (Amphora, Gomphonema, Cocconeis, Navicula) has noticeably decreased. In August and September 2020, with lower abundance indicators, the composition of communities turned out to be poorer than in the same period of 2019 with the predominance of diatoms and insignificant vegetation of cyanoprokaryotes (Leptolyngbya foveolarum).
The maximal cell density was found in epilithon, it significantly exceeds the densities found in epiphyton (105.27 million/10 cm2) and epipelon (22.96 million /10 cm2). The biomass of epilithon benthic biocenosis (18.66 mg/10 cm2) is also higher than that of epipelon (14.62 mg/10 cm2) and epiphyton (5.59 mg/10 cm2) (Fig. 4). Cyanoprokaryotes were numerically the leaders on stones, in the epipelon they were inferior to diatoms and greens.
When analyzing epilithic communities, stones with pronounced fouling of green filamentous algae (mainly Cladophora glomerata (L.) Kütz.), it was noted that it was first discovered at a depth of 75 cm at the end of May, when the surface water temperature was 17 °C. In early June, its biomass rapidly increased, reaching 19.25 g/10 cm2 (wet biomass). The peak of the biomass of the Cladophora complex was observed in June. After this rise, the intensity of fouling of stones with threads decreased significantly, with visual signs of aging (yellowing and degradation of chloroplasts) and death at the end of the growing season. As the biomass of filamentous green algae grew, the intensity of development of epilithic communities also tended to increase, mainly due to Cocconeis pediculus.
Structural diversity of benthic algae communities
The values of the Shannon – Weiner diversity index by abundance (HN) ranged from 0.21 to 3.54. The average value of the diversity index in epilithon communities (2.51) is higher than in the communities of epipelon (2.30) (TukeyHSD, two.way P < 0.01) and epiphyton (1.91) (TukeyHSD, two.way P < 0.001). Seasonal changes in the values of the diversity index of different epipelon and periphyton communities were almost the same with a maximum in September 2019) (Fig. 5). The difference in Shannon – Weiner index between seasons is statistically significant both for the epipelon community (two.way ANOVA, F value = 9.09, P < 0.01) and for the epiphyton community (two.way ANOVA, F value = 3.67, P < 0.01), however, there were no statistically significant differences for epilithon cenoses (two.way ANOVA, P > 0.05). The interannual differences in the structural characteristics of benthic algal communities turned out to be significant, and the diversity index was higher in the epipelon and epiphyton communities in autumn 2019 (TukeyHSD, two.way P < 0.001, P < 0.05) than in autumn 2020.

Seasonal variations in the Shannon–Wiener diversity index across different substrates of benthic algal communities in the Oka River
Discussion
The development and optimization of the water quality monitoring system for large eutrophic rivers of the Russian Federation in accordance with modern approaches requires the use of data not only on phytoplankton, but also on benthic communities, which first require a thorough study of their composition and structure. This is due to the fact that benthic algae are important components of river ecosystems, which play a key role in self-purification processes, as well as in the exchange of matter and energy with the environment. The structure and composition research of benthic algae in Russian rivers, such as the Oka River, allows to expand significantly the knowledge about functioning of river ecosystems and how these systems can be protected from anthropogenic impacts.
The Oka River, being one of the largest rivers in Europe, was characterized in the first decades of the 21st century as a watercourse of the eutrophic-hypertrophic type with hydrocarbonate-calcium waters, a high content of sulfates, organic substances, and components of the mineral nutrition of algae (Table 1). At the same time, weak seasonal changes (CV up to 15%) during the years of research were found in pH of waters, oxygen content, hydrocarbonates, and total mineralization. A somewhat greater variability of indicators (20–50%) is typical for the color, transparency and temperature of water, the amount of suspended solids, the concentration of magnesium ions and sulfates, total phosphorus and the easily oxidized fraction of organic matter (BOD5). The most labile components of chemical composition of the Oka waters (CV = 50–100% or more) include the content of monovalent alkaline metal ions, chlorine, as well as silicon, iron, the sum of nitrogen mineral forms, which determine the productivity of planktic benthic biocenosis. The peculiarities of the climatic indicators of the study years (a noticeably colder autumn of 2019 with heavy precipitation and rainfall compared to 2020) were reflected in the variability dynamics of water chemistry. More significant interannual changes in the autumn of different years of research differed in water temperature, the content of suspension in it, ions of alkaline earth metals and bicarbonates, mineral nitrogen, and silicon concentration. The variability of other indicators remained at the level of off-season values. In general, the general hydrochemical background of the medial of the Oka waters, contributing to the intensive development of phytoplankton in the river (eutrophy-hypertrophy) (Tockner et al. 2022) apparently weakly controls seasonal changes in the structural and functional characteristics of benthic communities. In conducting statistical analysis to explore the potential relationships between the structure of benthic algal communities and the chemical indicators representative of the medial Oka river’s, no significant correlations emerged. Possibly, they will be revealed by comparing biotic parameters with the composition and dynamics of abiotic characteristics in the very bottom layer of riparian zone waters 1–2 mm thick from the substrate (epiphyton and epilithon) and the composition of interstitial waters (epipelon), which requires special additional studies.
The relatively low total species richness of benthic communities (163 IT) in the researched area of the river, in our opinion, is determined by its ecotopic homogeneity (a straightened section of the right bank’s riparian zone, with sandy-silty soils, the absence of bays protected from runoff, wave and wind currents etc., very weak development of aquatic and coastal aquatic vegetation, insignificant presence of rocky substrates). The predominance of diatom diversity in benthic algal communities (53.87% of the total composition in the Oka River) is a characteristic feature of the phytobenthos of large and small rivers in the temperate zone and subtropical regions (Vladimirova 1978; Komulainen 2004, 2019; Allan and Castillo 2007; Gómez et al. 2009; Majewska et al. 2012 and others). Less noticeable is the role of green algae (41.1% of the list of benthic communities species), which are distinguished by the diversity of the river phytoplankton composition (45.1%) (Tockner et al. 2022). Possibly, this is due to the fact that diatoms are covered with a silica valve, which gives them protection from adverse environmental factors. They are also capable of rapid reproduction and adaptation to different environmental conditions, which may be one of the reasons for the diversity of this group.
In comparison with the watercourses of the taiga zone in the North-West of the Russian Federation (small and medium rivers), there is a tendency for the algoflora in the benthic zone of the Oka River to show depletion already at the level of large taxa (such as the absence of Rhodophyta, Chrysophyceae, Tribophyceae, Zygnemophyceae among the dominants). Among Cyanoprokaryota, the diversity of composition is shifted from Nostocales and Stigonematales towards Oscillatoriales (Oscillatoria, Phormidium, Pseudanabaena, Limnothrix). The dominance of Stigeoclonium, Oedogoniun, Ulothrix, Mougeotia, Spirogyra, Zygnema and Desmidiales was not established among green and charophytic algae due to the predominance of Cladohora glomerata, which develops sporadically on some stones. Among the dominant taxa in abundance, there are no representatives of oligoacid-neutral, meso-polyhumous oligotrophic-mesotrophic waters, such as Tabellaria fenestrata (Lyngbye) Kützing and T. flocculosa (Roth) Kützing, Eunotia, Pinnularia, Ceratoneis etc., which form the basis for the quantitative development of the epilithon and epiphyton of many rivers of the North-West of Russia (Komulainen 2004, 2005; Stanislavskaya 2006; Rusanov et al. 2012). At the mouth of the river among the benthic algae communities prevailing in the Oka, components of the eutrophic-hypertrophic complex of representatives of the genera Stephanodiscus, Aulacoseira, Melosira, Ulnaria, Navicula, Nitzschia, Gomphonema, Cocconeis and Oscillatoria were distinguished. A similar picture was noted for the river Vistula, flowing through the territory of Poland. Among the dominant genera in the composition of the benthic algoflora of the river, Aulacoseira, Cyclotella, Diatoma, Navicula, Nitzschia and Stephanodiscus were identified, while plankton representatives dominated in half of the studied samples (Majewska et al. 2012).
A multivariate analysis of the results obtained showed that the floristic composition of benthic algae and the complex of dominant species of benthic communities was mainly influenced by the type of substrate with a clear separation of all communities (epipelon and periphyton), which was also noted by other authors (Cox 1988; Rothfritz et al. 1997; Rusanov et al. 2012). At the same time, at the mouth of the Oka River composition of green algae on different substrates turned out to be more similar than that of cyanoprokaryotes, which showed more marked substrate specificity. Comparison of the taxonomic composition of diatoms from different substrates clearly separated the epipelon and periphyton communities; their composition turned out to be more similar in epilithon and epiphyton communities.
Microphytobenthic communities are closely related to the environment and habitat conditions determining the features of their structure and productivity (Majewska et al. 2012; Wu 2016). As prior research has shown, the species composition of benthic algae and cyanoprokaryotes at the Oka River mouth includes a wide range of ecological characteristics (Khedairia et al. 2022a), the presence of species with specific requirements for the quality and quantity of nutrients, the dynamics of light or other environmental conditions, which are more common on certain types of substrates, form features of their composition and structure. In addition, the development of certain species on a certain substrate may be associated with the influence of biotic factors, such as competition, grazing, fungal or viral infections. (Lamberti et al. 1987; Bernot and Turner 2001).
Seasonal variability in the abundance of communities in Oka River turned out to be higher in the communities of epiphyton (up to 273%) and epilithon (233%) due to the variability of this indicator in cyanoprokaryotes (280 and 207%, respectively). The population of the epipelon is more stable in the seasonal aspect (90%). The biomass of both the total and individual taxonomic groups varied less than the abundance and changed more strongly in communities of stony substrate (180%) and epiphyton (138%), also due to the variability in the development of cyanoprokaryotes (249 and 267%, respectively). The variability of the indicators of the abundance of benthic communities, as a rule, increased in summer and autumn, regardless of the type of substrate and in all taxonomic groups of algae. This may indirectly indicate an increase in the spatial differentiation of communities of different types during the seasonal succession of benthic communities as a result of a clearer isolation of the microhabitat system within individual substrates, especially stony and vegetative ones.
The ecological structure of the abundance and biomass of communities (the ratio of planktic, benthic, and benthic-planktic forms) (Barinova et al. 2019) reflected both the biotopic features of various substrates and trends in temporal changes. Lesser variability of indicators (average for the period of research CV = 27 and 50 – abundance and 38 and 56% – biomass) is typical for the eurybiont component of epiphyton and epilithon, while for the epipelon the abundance of true plankters turned out to be more stable (41 and 43%, respectively). Typically, benthic species showed a large variability in the indicators of quantitative development in spring (155% abundance, 197% biomass) and autumn (80–99 and up to 201%, respectively), following the dynamics of the composition and abundance of diatoms.
The abundance of communities can be determined by the type of substrate on which they grow (Rothfritz et al. 1997; Gómez et al. 2009; Majewska et al. 2012). In the current research, it was found that the average biomass of communities was the highest on the epilithon (stony substrate) and the lowest on the epiphyton (aquatic macrophytes).
According to Magurran (2004), the values of the species diversity of biotic communities fluctuate greatly, and the maximum values (Shannon index) rarely exceed 4.5. The diversity of benthic communities turned out to be at the level of average values and below average for epiphyton. The community structure, as a rule, became more complex by the end of summer and beginning of autumn. In addition, the results of the study confirm the hypothesis that the type of substrate plays a significant role in the formation of the diversity of communities of the bental of the Vistula River. The Shannon indices for epiphytic, epilithic, and epipsammic specimens were 1.22, 2.38, and 2.93, respectively (Majewska et al. 2012).
In general, the total species variety, its taxonomic parameters, the abundance and biomass of communities, the composition and abundance of predominant species in the mouth area of the Oka River with a sufficient level of nutrients (eutrophy-hypertrophy) change significantly during different seasons of the hydrological cycle and depend on such factors as type of substrate, and structural and functional characteristics of planktonic communities, The effects of water temperature and hydrochemical factors require additional investigation. The intensity of development of epipelon and epilithon communities were comparable, but the average abundance and biomass of epilithon were higher. These results can be used to better understand and predict the development of algae in the benthic zone of the mouth of the Oka River and optimization of the monitoring system for the quality of its water.
Conclusions
-
1.
The total species variety (163 IT) and indicators of quantitative development (average biomass 5.6–18.7 mg/10 cm2) of the benthic algal communities sampled at the specified station are formed by diatoms and green algae; in epilithon communities, the numerical development of cyanoprokaryotes is more noticeable.
-
2.
The list of the most significant components of communities from the cenotic point of view is formed by eurybiont representatives of mesotrophic and eutrophic-hypertrophic waters. It includes, along with the inhabitants of the bottom and fouling (Navicula, Nitzschia, Gomphonema, Ulnaria, Cocconeis, Amphora, Cymatopleura, Oscillatoria, Phormidium, Leptolyngbya etc.), typical planktonic components (Stephanodiscus, Aulacoseira, Melosira), which reflects a significant its impact on the periphytic and benthic communities.
-
3.
The classification of the composition of benthic algae cenoses using both qualitative and quantitative approaches clearly separated them according to the substrate feature, green algae can tolerate a wider range of environmental conditions than diatoms and cyanobacteria.
-
4.
With the relative stability of the chemical composition of the waters of the mouth area of the river and the absence of biogenic limitation (eutrophy-hypertrophy), noticeable seasonal differences in the composition and structural characteristics of benthic algae communities, apparently, are determined primarily by the type of substrate, the temperature characteristics of individual phases of the hydrological cycle, and the level of development of the planktonic algal communities in the benthic zone, which are more pronounced in spring and autumn.
-
5.
At the beginning and the end of the growing season (May – June, September – October), the cenoses of microphytobenthos of bottom sediments were almost completely determined by the structure of plankton and its quantitative development, and the proportion of autochthonous components (true benthic, foulers) did not exceed 20% of the abundance indicators. The influence of plankton on other types of benthic communities is less pronounced.
-
6.
The maximum abundance of benthic communities in the seasonal aspect is formed in summer, and the values of species diversity are shifted to the end of summer – beginning of autumn (September, for the epipelon – July – August) with a decrease in the intensity of proliferation of cyanoprokaryotes and the degree of dominance of certain diatom species.
-
7.
The intensity of development of epipelon and epilithon communities is practically the same, and the average biomass of epilithic communities is higher than that from bottom sediments and aquatic macrophytes. Filamentous green algae with a maximum development in the first half of summer are noted sporadically on a stony substrate at depths of no more than 1 m, which is determined by low water transparency.
-
8.
The analysis of diversity revealed that epilithic communities have a higher Shannon –Weiner index compared to epipelon and epiphyton, with the diversity peaking in September on a seasonal basis. Despite the same intensity of development between epilithon and epipelon, epilithic communities consistently showed greater species diversity. A notable increase in diversity indices in 2019 compared to 2020 reflects the interannual variability of this parameter within the benthic algal communities of the Oka River.
References
Allan JD, Castillo MM (2007) Stream ecology: structure and function of running waters, 2nd edn. Springer, Dordrecht
Barinova SS, Bilous OP, Tsarenko PM (2019) Algal indication of water bodies in Ukraine: methods and perspectives. University of Haifa, Haifa, Kiev
Bernot RJ, Turner AM (2001) Predator identity and trait-mediated indirect effects in a littoral food web. Oecologia 129:139–146
Cox EJ (1988) Has the role of the substratum been underestimated for algal distribution patterns in freshwater ecosystems? Biofouling 1, 49–63. https://doi.org/10.1080/08927018809378095
Dzhamalov RG, Nikanorov AM, Reshetnyak OS, Safronova TI (2017) The water of the Oka River basin: chemical composition and sources of pollution. Water Ecol 3:114–132
Gaiser EE, Trexler JC, Richards JH, Childers DL, Lee D, Edwards AL, Scinto LJ, Jayachandran K, Noe GB, Jones RD (2005) Cascading ecological effects of low-level phosphorus enrichment in the Florida Everglades. J Environ Qual 34:717–723
Gómez N, Licursi M, Cochero J (2009) Seasonal and spatial distribution of the microbenthic communities of the Rio De La Plata estuary (Argentina) and possible environmental controls. Mar Pollut Bull 58:878–887. https://doi.org/10.1016/j.marpolbul.2009.01.014
Guiry MD, Guiry GM (2019) AlgaeBase. World-wide electronic publication, National University of Ireland, Galway. 2019. http:www.Algaebase.Org.
Hildebrand H, Durselen C, Pollingher U, Zohary T (1999) Biovolume calculation for Pelagic and Benthic Microalgae. J Phycol 35:403–424
Hill BH, Willingham WT, Parrish LP, McFarland BH (2000) Periphyton community responses to elevated metal concentrations in a Rocky Mountain stream. Hydrobiologia 428:161–169
Khedairia T (2021) Ecological characteristics of the composition of diatoms of benthic algocenoses in the mouth of the Oka River. Issues Mod Algol Вопросы Современной Альгологии 131–133. https://doi.org/10.33624/2311-0147-2021-1(25)-131-133
Khedairia T, Okhapkin AG (2021) Diatoms in algocenoses of bental as indicators of water quality in the mouth of the Oka River (Russia). Princ Ecol 41:99–115. https://doi.org/10.15393/j1.art.2021.11583
Khedairia T, Okhapkin AG, Fukina DG (2022a) Composition and structure of Benthal Algocenoses of a large Eutrophic River: I. taxonomic characteristics and Dominant species of communities (Oka River, Russia). Inland Water Biol 15:555–565. https://doi.org/10.1134/S1995082922050121
Khedairia T, Okhapkin AG, Genkal SI, Gluschenko AM, Kulikovskiy MS (2022b) Composition and Ecological Characteristics of Diatoms of algocenoses in the Benthal of the Mouth of a large eutrophic-hypertrophic river (Oka River, Russia). Inland Water Biol 15:415–423. https://doi.org/10.1134/S1995082922040320
Kholodnoho I, Vasser SP (1989) Vodorosli Spravochnik. Naukova dumka
Komulainen SF (2003) Metodicheskie rekomendatsii po izucheniyu fitoperifitona v malykh rekakh (guidelines for the study of Phytoperiphyton in Small Rivers), Petrozavodsk: Karel. Nauchn Tsentr Ross Akad Nauk
Komulainen SF (2004) Phytoperiphyton in rivers of Republic of Karelia. Bot J 89:18–35
Komulainen SF (2005) Structure and functioning of phytoperiphyton in small rivers of Eastern Fennoscandia. Dissertation, St. Petersburg
Komulainen SF (2019) Phytoperiphyton of the Kem River and its tributaries (Republic of Karelia, Russia). Int J Algae 21
Lamberti GA, Ashkenas LR, Gregory SV, Steinman AD (1987) Effects of three herbivores on periphyton communities in laboratory streams. J North Am Benthol Soc 6:92–104
Magurran AE (2004) Measuring biological diversity blackwell science. Biol Divers Front Meas Assess Oxf 105
Majewska R, Zgrundo A, Lemke P, De Stefano M (2012) Benthic diatoms of the Vistula River estuary (Northern Poland): seasonality, substrata preferences, and the influence of water chemistry: benthic diatoms of the Vistula estuary. Phycol Res 60:1–19. https://doi.org/10.1111/j.1440-1835.2011.00637.x
Meteleva NY (2013) Structure and productivity of phytoperiphyton in water bodies of the Upper Volga basin. Dissertation, Institute of Biology of Inland Waters named after I.D. Papanin of the Russian Academy of Sciences
Montagna PA, Blanchard GF, Dinet A (1995) Effect of production and biomass of intertidal microphytobenthos on meiofaunal grazing rates. J Exp Mar Biol Ecol 185:149–165
Nevrova EL, Snigireva AA, Petrov AN, Kovaleva GV (2015) Rukovodstvo Po Izucheniyu morskogo mikrofitobentosa i ego primeneniyu dlya kontrolya kachestva sredy (guidelines for the study of Marine Microphytobenthos and its application for Environmental Quality Control). N. Orianda, Sevastopol’
Okhapkin AG (1998) Species composition of phytoplankton as an indicator of living conditions in streams of different types. Bot Zh 83:1–13
Okhapkin AG, Andriyanova NV, Maksimova VA (2015) Dynamics of hydrochemical composition in the lower reaches of the Oka River. Khim. Ekol, Voda
Okhapkin AG, Vodeneeva EL, Startseva NA, Khedairia T (2019) Ekologicheskii monitoring. Metody Izucheniya Fitobentosa Presnykh Vodoemov Posobie (Environmental Monitoring. Methods for Studying Phytobenthos of Fresh Water Bodies, Nizhny Novgorod
Perissinotto R, Nozais C, Kibirige I (2002) Spatio-temporal Dynamics of Phytoplankton and Microphytobenthos in a South African temporarily-open Estuary. Estuar Coast Shelf Sci 55:47–58. https://doi.org/10.1006/ecss.2001.0885
Rothfritz H, Jüttner I, Suren AM, Ormerod SJ (1997) Epiphytic and epilithic diatom communities along environmental gradients in the Nepalese Himalaya: implications for the assessment of biodiversity and water quality. Arch Für Hydrobiol 138:465–482. https://doi.org/10.1127/archiv-hydrobiol/138/1997/465
Rusanov AG, Stanislavskaya EV, Ács É (2012) Periphytic algal assemblages along environmental gradients in the rivers of the Lake Ladoga basin, Northwestern Russia: implication for the water quality assessment. Hydrobiologia 695:305–327. https://doi.org/10.1007/s10750-012-1199-5
Shen X, Li R, Cai H, Feng J, Wan H (2022) Characteristics of secondary flow and separation zone with different junction angle and flow ratio at river confluences. J Hydrol 614:128537. https://doi.org/10.1016/j.jhydrol.2022.128537
Stanislavskaya EV (2006) Periphyton structure as indicator of ecological conditions in the Lake Ladoga tributaries and the Neva River. Assessment of Ecological Condition in Rivers of the Lake Ladoga Basin using hydrochemical characteristics and structure of Hydrobiocenosis. Lema, St. Petersburg, p 91103
Team R RC (2022) A language and environment for statistical computing. 2022. R Foundation for Statistical Computing, Vienna, Austria. R Found Stat Comput. Austria
Tockner K, Zarfl C, Robinson CT (2022) Rivers of Europe, Second edition. ed. Elsevier, Amsterdam Oxford Cambridge, MA
Vymazal J (1988) The use of periphyton communities for nutrient removal from polluted streams. Hydrobiologia 166:225–237
Wetzel RG (1963) Primary productivity of periphyton. Nature 197:1026–1027
Wu Y (2016) Periphyton: functions and application in environmental remediation, 1st edition. ed. Elsevier, Waltham, MA
Yuan S, Xu L, Tang H, Xiao Y, Gualtieri C (2022) The dynamics of river confluences and their effects on the ecology of aquatic environment: a review. J Hydrodyn 34:1–14
Zheng L, Xu J, Tan Z, Xu G, Xu L, Wang X (2020) A thirty-year Landsat study reveals changes to a river-lake junction ecosystem after implementation of the three gorges dam. J Hydrol 589:125185
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The authors did not receive support from any organization for the submitted work. The authors have no financial or proprietary interests in any material discussed in this article.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Khedairia, T., Okhapkin, A. & Yakimov, V. Succession, composition and structure of microphytobenthic communities of the large eutrophic Oka River in Russia, before its confluence with the Volga River. Biologia 79, 1623–1637 (2024). https://doi.org/10.1007/s11756-024-01635-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11756-024-01635-3