
Abstract
Climate change is introducing several challenges to agriculture’s sustainability. The drought stress is hampering cereal crop productivity. There is an increasing interest in exploring how well physiological flexibility in plants can buffer drought stress. Our study investigated how plant growth-promoting bacteria (PGPB) and SORGAAB, which is a water-based extract of Sorghum bicolor L. (10 g/100 ml), affect the physiological, biochemical, and vegetative profiles of Zea mays L. under drought conditions (50% field water capacity). Among all treatments, the application of drought + PGPB + SORGAAB spray at the three-leaf phase (V3) showed significant improvement in growth. We found that the drought + PGPB + SORGAAB treatment increased the amount of amylase (58%), chlorophyll-a and b (67 and 83%, respectively), catalase (58%), superoxide dismutase (45%), peroxidase (52%), and crude protein (69%), compared to the drought treatment alone. However, H2O2 levels went down by 57%. Our results also showed a 90.5% and 95% increase in macro- and micronutrient concentrations, respectively, under PGPB + SORGAB treatment. The combined application of PGPB + SORGAAB at the V3 phase was more effective in enhancing the vegetative biomass by yielding 2.41 and 5.67 g/pot of the shoot and root dry weights, thereby impacting the root-to-shoot ratios, and enabling maize plants to improve growth under drought conditions.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The maize (Zea mays L.) crop is a fundamental cereal for food security (Sohail et al. 2022), and among other cereal crops, it has the third highest ranking as a primary cereal crop. Due to its protein source, it is used as food and feed (Ray et al. 2022). Maize is the backbone of the starch industry and nowadays paved the way for edible oil production (Krishna Motukuri 2020). In comparison to wheat (Triticum aestivum L.), which produces 3.7 million calories per hectare, and rice (Oryza sativa L.), which produces 1.7 million calories per hectare, maize is predicted to produce 6.9 million calories (Krishna Motukuri 2020; FAOSTAT 2021). In terms of output, maize was produced on 202 million hectares (Mha) worldwide and came in first place among cereals with a production of 1128 million metric tonnes in 2020, followed by wheat and rice (FAOSTAT 2021). In Pakistan, it is the third most important cereal crop and is planted across 1.016 Mha (65% irrigated and 35% rainfed), with a total yield of 7.8 million metric tonnes (GOP 2021).
Because of global climate change, maize production is subjected to a variety of abiotic stresses. Among abiotic stresses, drought (water deficiency) has become a severe threat to cereals’ productivity, especially to maize in arid and semi-arid regions (Sabagh et al. 2018). A 40% global decline in maize production in the previous three decades was attributed to drought, as per a meta-analysis study by Daryanto et al. (2016). Using a vector autoregression model, Shakoor et al. (2017) reported that maize yield will further decrease by 6% up to 2030 because of climate change. The International Monetary Fund (IMF) assessment has already declared Pakistan the third-worst country in the world for having a severe water deficit. The yearly water availability per person in Pakistan is 1017 cubic meters, which is dangerously close to the 1000 cubic meter threshold of scarcity. Pakistan had roughly 1500 cubic meters of water per person available in 2009 (Nabi et al. 2019). Pakistan is unfortunate to be positioned in an arid and semi-arid climate. Moreover, the spatial drought (rainfall variability) events are more alarming for water and, therefore, food security.
Under incessant drought conditions, production loss of maize is unavoidable in agriculture (Farooqi et al. 2020). Productivity is a function of gas exchange (CO2) from the atmosphere to water inside the plant (transpiration); therefore, under drought stress, plants must compromise on many metabolic functions (Simova-Stoilova et al. 2020), i.e., reduced transpiration results in poor photosynthesis. Drought is also a precursor to secondary salinization, thus impairing nutrient imbalances and ionic toxicities (Farooqi et al. 2020). Furthermore, dryness stimulates the production of ethylene, which slows root length and growth (Carlson et al. 2020). Drought is linked to alterations in root system development, osmoregulation, oxidative stress management via phytohormone biosynthesis and metabolism, or the creation of antioxidants in response to reactive oxygen species (ROS) production (Farooqi et al. 2020; Khaliq et al. 2022). There are three main ways that a lack of water in the soil lowers crop production: (1) less photosynthetically active radiation absorption by the canopy (PAR); (2) less radiation utilization efficiency (RUE); and (3) a lower harvest index (HI) (Tripathi et al. 2018). Plants have evolved various adaptation systems to deal with drought stress. Restoring drought-induced damage and plant development requires the reorganization of several metabolic pathways, which is a highly complex process. It is much more complicated than simply going back to how you were before the stress started.
Various approaches are proposed for reducing the negative impacts of drought stress. These strategies include using traditional breeding and genetic engineering to create drought-tolerant varieties that depend on biotechnological development, time, and resources. Yet, water management practices, including irrigation techniques and agronomic enhancement of water use efficiency through soil and foliar amendments like soil mulches, moisture absorbents, stress signaling molecules, osmo-protectants, plant growth promoting bacteria (PGPB), and crop/plant osmolytes (Niu et al. 2018), are more viable and economical approaches to combat the drought challenge in crop production. Being environmentally safe and economically feasible, crop enhancement under abiotic stressors has huge potential due to PGPB and phytochemicals (Ibrahim et al. 2022; Rouphael et al. 2021). In the case of PGPB bacteria quickly attack plant roots in the rhizosphere and form a strong and close living relationship with them. They make the rhizosphere richer and safer for host plants by producing exo-polysaccharides, phytohormones, and volatile compounds; increasing the production of osmolytes and antioxidants; and changing the shape of roots, the structure of the soil, and how much water they hold (Kour et al. 2020).
Contrary to traditional agrochemicals, phytochemicals have already paved their way in crop production as efficient biostimulants and plant protectants (Tudu et al. 2022). Sorghum (Sorghum bicolor L.) is a rich source of exclusive phytochemicals with potential implications as a bio-stimulant. Sorghum contains a variety of bioactive chemicals like dhurrin, cyanogenic glycosides, and various soluble phenolic compounds, including antioxidants, making it an important allelopathic crop and source of natural biostimulants (Rouphael et al. 2021). Foliar application with sorghum water extract (SORGAAB) improves leaf growth and helps to increase the tolerance of the plant against water scarcity (Farooq et al. 2018). The phytochemistry of SORGAAB is dependent on sorghum age and genotype (Bajwa et al. 2018). Sorghum water extracts improved wheat crop tolerance to heat stress when given in low quantities (Afzal et al. 2020). The effect of the application of allelochemicals, either for growth stimulation or retardation, is dose-dependent (Khaliq et al. 2022). As far as we know, not much has been written about how PGPBs can work together with phytoextractants applied through the foliar route during drought. We hypothesized that the combination of two strategies could have great potential to mitigate drought impacts.
Materials and methods
Experimental site description
A pot experiment was conducted in September–October 2021 at the research area of the University of Okara, Okara, Punjab, Pakistan (latitude = 30° 48′ N, longitude = 73° 27′ E, altitude = 183.5 m). The site falls under the subtropical climate region classification, with mean temperatures ranging from 23.39 °C to 35.42 °C, an annual rainfall average of 22.35 mm, and an average humidity of 33.66%. The laboratory analysis was performed at High-Tech Lab-II, Sub-campus Depalpur Okara, University of Agriculture Faisalabad, and Plant Ecology Laboratory, Department of Botany, University of Okara, Punjab, Pakistan. The experimental set-up was built under a transparent glass-shaded and side-fenced open-wire house with an average temperature of 34 ± 5 °C, and air humidity remained at 40% ± 7 during the experiment period.
Amendments sourcing and preparation
Maize seeds
The maize variety “Sahiwal Gold,” which is currently under cultivation with a yield potential of 9100 kg/ha (https://aari.punjab.gov.pk/crop_varities_maize_millets), was used for this experiment, and it was collected from the Maize and Millets Research Institute, Yusaf Wala, Sahiwal, Punjab, Pakistan.
Sorghum water extract (SORGAAB)
Sorghum water extract (SORGAAB) was prepared from mature and disease-free whole sorghum plants (excluding roots), which were collected from the field and washed in running water once. Plants were dried by spreading them on a polyethene sheet under shade. The dried plants were chopped (~ 1 cm) and soaked in distilled water (1:10, w/v) for 24 h at room temperature (27 ± 2 °C) (Scavo et al. 2019). The extract was initially filtered through a porous stainless-steel sieve and then filtered through Whatman filter paper grade 40 (8 μm medium flow filter paper) to remove sorghum debris. The filtrate was diluted with distilled water to achieve a final concentration of 5% (w/v). The extract was stored at 4 °C. As a control treatment, distilled water was used. The foliar spray was applied using a hand shower with a 1-L capacity.
Application of plant growth promoting bacteria (PGPB)
PGPB was collected from the Soil and Microbiology Laboratory, Ayub Agricultural Research Institute (AARI), Punjab, Pakistan. The PGPB strain was Bacillus subtilis HAS31 (16S rRNA gene partial sequence accession = MT658521; available at (https://www.ncbi.nlm.nih.gov/nuccore/MT658521). This strain was originally isolated from the potato producing region (district Okara). The PGPB and drought stress tolerance has already been tested by Batool et al. (2020). The seed priming with inoculant was performed following the method described by (Paravar et al. 2023). The maize seed surfaces were sterilized before coating by immersing them in 70% (v/v) ethanol for 1 min and then in a 1% sodium hypochlorite solution for 3 min. The maize seeds were rinsed with distilled water and dried using paper towels. Weighing the seeds that had bacterial suspension treatment was the first step in making the suspension of bacteria. According to the total weight of the seeds, the bacterial suspension was prepared (100 g sugar + 500 ml distilled water + bacterial inoculum). After the preparation of bacterial suspension, the seeds were treated with the suspension in a pre-washed dried glass jar by shaking and rotating. Treated seeds were kept for 30 min in the shade to ensure stickiness and drying.
Experimental design
The experiment was arranged under a complete randomized design (CRD). Each treatment had four replicates and pots with different treatments arranged in a CRD layout. Pots were filled with clay loam soil, which was collected from arable land with pH 7.8, EC 2.55 dS m−1, organic matter 0.65%, nitrogen (N) 0.21%, phosphorus (P) 6.75 mg kg−1 and potassium (K) 120.98 mg kg−1. Each 50 cm tall by 30 cm wide plastic pot included ten PGPB-coated seeds. Thinning was done to maintain the five healthy seedlings per pot while germination was constant. Five treatments were used in the experiment: T1 was normal irrigation with 100% field capacity moisture (CK1); T2 was drought stress conditions with 50% moisture contents (CK2); T3 was drought stress plus PGPB; T4 was drought stress plus PGPB plus SORGAAB (10% w/v) spray at the three-leaf stage (V3); and T5 was drought stress plus SORGAAB (10% w/v) spray at the V3 stage. From planting to harvesting, the moisture contents were maintained throughout the study period. The field capacity was determined using Nachabe’s proposed gravimetric approach. The plants were harvested after 25 days of sowing along the soil sampling, and the experiment was terminated. The plant’s agronomic parameters—shoot/root fresh weights, number of leaves, leaf length, leaf weight, and stem diameter—were recorded on the day of harvesting.
Measurements
Germination indices
The defined formula was used to compute the germination index (GI), as given below:
The germination energy (GE%) was calculated by dividing the proportion of seeds, that germinated at the first count (4 days after sowing) by the total number of tested seeds and the proposed equation is given below:
Growth/biomass
Plants were picked 25 days after sowing to estimate seedling growth and conduct biochemical analysis. The root and shoot lengths of three randomly selected plants from each container were measured. The roots and shoots of the seedlings from each treatment were separated from the tops (separate the root from the top; cut at the soil line and carefully transfer the soil and root samples to a sieve and separate the soil from the roots using fresh water), and dry weights were recorded after oven-drying the samples at 75 ± 2 °C for 48 h until the constant weights were achieved. The root/shoot dry weight ratio and root: shoot length ratio for each treatment having three replicates (n = 1) were calculated by following the formulas, respectively.
Stress tolerance (%) determination
The stress tolerance was proposed for yield-related attributes. Here, we have adapted the stress tolerance (with slight modifications) for the plant height from Fernandez (1992). It defines the plant’s ability to maintain sufficient growth in the stress environment corresponding to the control conditions in terms of the overall performance of the plant. The stress tolerance was determined using the following formula:
where Y in the equation represents the biomass yield. Ycontrol represents the CK1 treatment. Ydrought denotes the average biomass yield under CK2 condition, and Ycontrol denotes the average biomass yield under CK1. To assess the response of dry mass to salinity, a regression model was used as a function of the best-fit line and the highest R2.
Biochemical parameters
The root samples (0.1 g for each of the three replicates) were broken up with a mortar and pestle and mixed with 100 mL of distilled water. They were then left alone at 4 °C for 24 h to test the a-amylase activity by filtering and separating the precipitates. The enzyme activity in the supernatant was then estimated using the dinitrosalicylic acid (DNS)-based technique using spectrophotometer analysis at an absorbance wavelength of 545 nm, as described by Bernfeld (1955). This assay has been widely employed for the estimation of reducing sugars derived from the pretreatment of lignocellulosic biomass. This assay tests for the presence of free carbonyl groups (C=O), so-called reducing sugars.
The chlorophyll (Chl) contents were quantified according to the methods of Arnon (1949), and the formula explained below. Using a mortar and pestle, the fresh leaves (0.05 g) were crushed, and 10 mL of acetone (80%) was used for the extraction process. The extract was then kept in the dark all night. Following centrifugation at 5000 rpm, the supernatant was withdrawn. The absorbance was measured using a spectrophotometer (Specord 200 Plus-Analytik Jena) at 645 and 663 nm wavelengths. Additionally, an acetone (80%) blank was run. By applying the provided formulas, the contents of Chl-a and Chl-b were calculated (n = 3).
The Giannopolitis and Ries (1977) method was used to measure the superoxide dismutase (SOD) activity. The 25-day-old fresh leaf tissue (1 g) was thoroughly ground with a cold mortar and pestle in an ice bath until no fibrous residue could be seen. The grinding medium (4–6 ml/g fresh weight) consisted of 0.1 M K-phosphate and 0.1 mM EDTA (pH 7.8), plus homogenizing glass beads. The homogenate was centrifuged twice at 13,000 g for 10 min in a refrigerated centrifuge at 0 to 5 °C. 20–50 µL of enzyme extract (supernatant), potassium phosphate (K-phosphate) buffer (pH 7.8), 1.3 µM riboflavin, 13 mM methionine, and 63 µM NBT were combined to create the reaction mixture. Distilled H2O was added to reach the final volume of 3 ml. The test tubes were exposed to fluorescent light for 15 min, and a spectrophotometer was used to measure the absorbance at 560 nm (UV-4000). The enzymes and protein concentrations were reported as mg g−1 FW, as described by Amrutha et al. (2019).
where: * without SOD extract; # with SOD extract.
The activities of catalase (CAT) and peroxidases (POD) were measured using the procedure outlined by Chance and Maehly (1955). 0.5 g of plant material was extracted in 0.01 M K-phosphate buffer, pH 7.0. To measure CAT, an assay solution was made with 50 mM K-phosphate buffer (pH 7.0), 0.1 mL of enzyme extract, and 5.9 M H2O2, and 50 mM K-phosphate buffer (pH 7.0) was produced. A drop in absorbance at 240 nm was noted every 20 s. The CAT activity was calculated according to the following formula:
where \(\Delta A\) represent the absorbance of mean values that change at 240 nm over 20 s, V is the measured aliquot volume (1 mL), Vt is the volume of the sample solution (mL), 0.1 is the catalase concentration (µg/ml), T is the measurement duration, and W is the sample’s fresh weight. For measuring the POD activity, a reaction mixture containing 0.1 mL of enzyme extract, 40 mM H2O2, 20 mM guaiacol, and 50 mM K-phosphate was used to measure the POD activity. After every 20 s, the absorbance at 470 nm was measured. The POD activity was calculated according to the following formula.
where V is the measured liquid volume (mL), Vt is the volume of the sample solution (mL), 0.1 is the catalase concentration (µg/ml), T is the measurement duration, and W is the sample’s fresh weight.
The determination of hydrogen peroxide (H2O2) was executed by adopting the method proposed by Velikova et al. (2000). A sample of 2 g of fresh leaves was extracted with 10 mL of trichloroacetic acid (TCA) (0.1%, w/v). After centrifuging the TCA-containing extract, 1 mL of the supernatant was mixed with 2 mL of 1 M potassium iodide and 1 mL of 0.05 M sodium phosphate buffer (0.05 M, pH 7.0) to measure the amount of H2O2. Using a spectrophotometer (Specord 200 Plus-Analytik Jena), the mixture’s absorbance was measured at a wavelength of 390 nm, and TCA was used as the control. The unit mg/g FW was used to express the values.
Crude proteins were measured by determining N contents in dried leaf samples. The leaf tissues were air dried and then ground to a powder-size fraction. The value of crude protein was calculated from the total nitrogen content (TN) of the sampled leaves analyzed through acid digestion (TCA) followed by Kjeldahl distillation (Bremner and Mulvaney 1982) and multiplied by a factor of 6.25.
Soil parameters
The microbial counts in soil samples were assessed for colony-forming units using the dilution spread plate method. Half-strength R2A nutrient agar, which was utilized as a growth medium for colonies (Aslam et al. 2018). 100 µl of 10–1 to 10–5 dilutions of soil samples (1 g) were put on nutritional agar after being serially diluted from 1 g of soil samples. The plates were carefully sealed with plastic film once the dilutions had spread, and they were then incubated at 30 °C for 24 h. To find out how many culturable bacteria populations there were on R2A (half-strength) agar plates, the methods described by Janssen et al. (2002) were used.
The processes described by Walkley and Black (1934) and Helmke and Sparks (2018) were used to assess soil organic matter (SOM) contents and available potassium (K) and phosphorus (P), respectively. The widely adopted DTPA-TEA (di-ethylene triamine penta acetic acid tri-ethanolamine) extract was used to assess the bio-available Fe, Cu, Mn, and Zn concentrations in the post-harvested. The micronutrients were analyzed using an atomic absorption spectrophotometer (Analytik Jena novAA 350) available at High-Tech Lab-II, Sub-campus Depalpur Okara, University of Agriculture Faisalabad.
Statistical analysis
The data organization, calculation, and graphical presentation were performed with Microsoft Excel 2016 (Redmond, Washington, DC, USA). With the help of the CRD model, statistical analysis was carried out using Statistix 8.1 (Analytical Computer Software, Statistix 8.1; Tallahassee, FL, USA, 1985–2003) to evaluate the significant differences between the treatment means. Analysis of variance (ANOVA) was used to analyze data gathered from diverse observations. The least significant differences (LSD) test with 5% probability was used to compare the treatment means. Pearson’s correlation analysis and graphics were performed through OriginPro v. 2021 (Origin Lab Corp., Northampton, MA, USA) and Sigmaplot 10.0 (Systat Software Inc., San Jose, CA, USA).
Results
Germination indices. Morphological traits and stress tolerance index
This study clearly explains the drought stress mitigation through the above-ground (plant growth promoting rhizobacteria) strategy and above-ground (foliar sorghum water extract application), either alone or in combination, in Zea maize L. plants. The physiological and biochemical evidence supports our hypothesis. Except for the germination index, other physiological traits, including germination percentage, energy, and the plant’s morphological parameters, were significantly affected by treatments. This study depicted that reducing the soil moisture contents (CK2) not only hindered the germination percentage by 34% (Fig. 1) but also resulted in a 65–99% decrease in the morphological traits of maize plants compared to CK1 (Fig. 2). The germination energy was decreased by 65% in CK2 (Fig. 1). Yet, the PGPB treatments, either alone (T3) or in combination with SORGAAB (T4), had the highest germination percentage (95%), and germination energy (64%), compared to other treatments (Fig. 1). The growth parameters—shoot fresh and dry weights (g), root fresh and dry weights (g), shoot/root lengths (cm), and stem diameter (mm) (Fig. 2A–D, respectively)—differed significantly (P ≤ 0.05) between different PGPB and SORGAAB treatments. The results showed that statistically maximum (P ≤ 0.05) germination (95%), root dry weight (5.67 g), shoot length (10.10 cm), root length (40.00 cm), and stem diameter (6.75 mm), were observed with T4 treatment where both the PGPB and SORGAAB were applied under drought conditions. Interestingly, among all treatments, CK1 had the highest shoot fresh weight (11.7 g), which was 40% and 77% higher than T4 and CK2 treatments.

Impact of PGPB and SORGAAB treatments on germination indices of maize plants growing under drought conditions. The means of treatments not sharing a common letter are significantly different at the 5% level, according to the LSD test. PGPB = plant growth-promoting bacteria, SORGAAB = sorghum water extract. The germination rate and germination energy are calculated as % and presented on Y-bar while germination index is the ratio and present only numbers

Impact of PGPB and SORGAAB treatments on morphology and growth of maize plants under drought conditions at the 25th day of growth; shoot fresh and dry weight (A), maize root fresh & dry weight (B), shoot & root length (C), stem diameter (D). The means for treatments not sharing a common letter are significantly different at the 5% level, according to the LSD test. PGPB = plant growth promoting bacteria, SORGAAB = sorghum water extract
For plants, the root-to-shoot ratio is significant to study under drought conditions; therefore, based on weight and length, the root-to-shoot ratios were calculated. We found that in both cases, a similar trend was observed. The highest root: shoot on weight basis was observed with T4 treatment (2.35), which was then followed by T5 (1.62) > T3 (1.49) > CK1 (0.97) > CK2 (0.59). Similarly, on length basis, highest ratio was also observed with T4 (3.96) followed by CK1 (3.48) > T5 (3.24) > T3 (2.75) > CK (2.64). Similarly, the stress tolerance index, measured in percentage, was plotted against total dry mass yield as an exponential model. The obtained regression equation (Y = 0.09x + 0.25), presented in (Fig. 3) indicated a linear relationship between two variables (R2 = 0.97). With every increase of 1% in stress tolerance achieved in maize plants, it would contribute to a 9% increase in the total dry mass yield of maize plants in a drought-stress environment. The exponential equations also present the other factors of variability, which share 25% of dry mass yield.

Exponential model for accounting for the variability and prediction of plant dry mass yield response against stress tolerance % as described in the equation from the methodology section
Soil nutrients and microbial analysis
The SOM, macronutrients (TN, K, and P), micronutrients (Cu, Fe, and Zn), and CFU differed significantly (P ≤ 0.05) among various applications of PGPB and SORGAAB as compared to CK2 (Table 1). The SOM contents were highest (1.25%) with T4. The application of drought + PGPR + SORGAAB (10% w/v) spray at the three-leaf phase increased the TN, K, P, Cu, Fe, and Zn, and CFU by 90, 65, 44.2, 95, 44, 73, and 71% respectively, compared to CK2.
Maize plant biochemical and enzymes profile
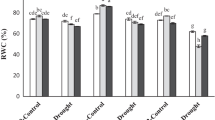
In the maize plant, the amylase, chlorophyll-a and b, H2O2, CAT, SOD, POD, and crude protein contents differed significantly (P ≤ 0.05) among varying applications of PGPB and SORGAAB at the V3 phase under drought conditions (Table 2). Decreasing the water availability by 50% (CK2), the amylase, chlorophyll a, chlorophyll b, and crude protein contents of maize leaf were decreased by 28.8, 24.6, 18.3, and 28.8%, respectively, while the H2O2, SOD, and POD were increased under CK2 by 31.3, 14, and 11.4% respectively, compared to CK1. On the other hand, application of drought + PGPB + SORGAAB at V3, enhanced (P ≤ 0.05) the amylase (58%), chlorophyll-a (67%), chlorophyll-b (83%), catalase (58%), superoxide dismutase (45%), peroxidase (52%), and crude protein (69%) concentrations compared to CK2. Mitigation of drought through either 100% irrigation (CK1) only, the sole application of PGPB (T3), or the sole application of SORGAAB (T5) enabled the sequestration of only 30–31% of H2O2 compared to CK2 while the application of PGPB in combination with SORGAAB spray at V3 mitigated the H2O2 contents by 57% as compared to CK2.
We used Pearson’s correlations between the mean values of each treatment to show the variation and relationship between the traits we measured. The results were shown as a heatmap diagram (Fig. 4). Among the investigated parameters, we found correlations among plant biochemical traits (Amyl, H2O2, CAT, SOD, POD, CP, Chl a and b), plant physiological traits (total fresh weight, root and shoot dry weight, plant height, and total dry biomass yield), and theoretical indices (salt tolerance%, root-to-shoot ratios). Our results showed that most of the parameters were significantly positively correlated at both confidence levels (p ≤ 0.05 and p ≤ 0.01). Only H2O2 was found to be strongly but negatively correlated with all traits except the SOD, POD, shoot dry weight, and plant height (the blue circles in Fig. 4). This finding is significantly supporting information for the explanation of our empirical results. Important yield and drought tolerance traits (drought stress tolerance% and total dry mass yield) were positively associated in a strong relationship (the red circles in Fig. 4).

Pearson’s correlation among plant biochemicals, physiological, agronomic traits and empirical indices; Here; AmyL = amylases, H2O2 = hydrogen peroxides, CAT = catalase, SOD = superoxide dismutase, POD = peroxidases, CP = crude protein, Chl a,b = chlorophyll-a, b, TFW = total fresh weight, RDW = root dry weight, SDW = shoot dry weight, R:S/L = root:shoot ratio on length bases, R:S/W = root:shoot ratio on weight bases, PH = plant height and TDW = total dry biomass yield. Red and blue color show positive and negative association respectively
Discussion
Considering the food security challenge, drought stress is the most determinantal factor in maize yield productivity, impairing the growth, developmental stages, morphology, and physiology of maize plants (Sabagh et al. 2018). The impact of water scarcity becomes more staggering when it comes to maize production in arid or semi-arid climates (Ojuederie et al. 2019), like in Pakistan, which is already on the verge of water scarcity and extreme climatic shifts (Shakoor et al. 2017). In times of limited water, germination and the shape of plants are severely harmed because of problems with their physiological functions (Farooq et al. 2018). These problems are mostly caused by an imbalance in the rate of carbon fixation and utilization, which changes the photosynthetic metabolism and causes too many ROS to be produced. As we have observed in this study, all germination indices were at their lowest values under 50% drought conditions (Fig. 1 and Table 2). It is a well-known fact that seeds need moisture to germinate and grow; yet, without enough water, seedling development is either slowed down or stopped altogether (Ojuederie et al. 2019). As depicted in Fig. 1, lowering the water availability to 50% field capacity (FC) compared to 100% FC resulted in a 53% lower germination percentage of plants due to 16% less GE. The reason for such low GE is the low availability of soil moisture (Hafez et al. 2021). Ultimately, this resulted in poor seedlings with poor agronomic and morphological traits, compared to plants growing at 100% FC. Similarly, the root-to-shoot ratio, which is an important morphological characteristic of maize plants, was also severely affected, and this phenomenon has recently been demonstrated by Khaliq et al. (2022) under 30 and 60% field capacity in wheat.
Drought stops the production of chlorophyll and the uptake of minerals by the roots and their movement to the parts above ground. This lowers seed yield because the plants can’t assimilate as much (Yang et al. 2021). It has been reported that plants reduce the biosynthesis of certain proteins and increase the biosynthesis of defensive proteins to overcome drought stress trauma in plants, which is also dependent on the intensity and duration of stress (Ahmad et al. 2019). In our investigation, all growth and germination parameters were significantly affected by the PGPB and SORGAAB applications. However, for the three germination parameters, the trend remained the same for all parameters, where the PGPB effect is more pronounced than the PGPB + SORGAB. The first reason for such a non-significant difference is the application time of SORGAB. The germination indices were measured 10 days after sowing, while V3 clearly demonstrated them on the 19th day after sowing. That’s the reason CK2 and Drought + SORGAAB were also statistically non-significant. Such results have been previously explained by Khaliq et al. (2022) and Ahmad et al. (2019), where the interactive effect of either foliar extract or PGPB remained non-significant. Moreover, the distribution of photosynthates under drought stress is the primary focus of plants that depend on hormonal signaling. In this study, high root biomass or stem diameter may have resulted from the maize plant’s ability to help the plants increase their above-ground and below-ground growth through nutrient acquisition, assimilation, enhanced root networking, and metabolite production (Ahmad et al. 2019). This kind of metabolite assimilation in roots may have resulted in lower accumulation in above-ground tissue, resulting in a reduction in cell division, elongation, and expansion (Khaliq et al. 2022; Farooq et al. 2018; Osakabe et al. 2014).
More people are becoming aware of the use of PGPR as microbial seed inoculants as an environmentally benign alternative to the ongoing use of harmful chemical fertilizers for crop enhancement (Ojuederie et al. 2019). Similarly, phytochemicals have been widely used as bio-weedicides, insecticides, fungicides, etc. in crop production and crop protection (Bajwa et al. 2018), but their exogenous use in promoting plant growth and enhancing various modes of stress tolerance owing to their antioxidant, bio-stimulation, and nutritional features, is currently scaling in crop production (Tudu et al. 2022). Therefore, the use of phytochemicals is also deemed an eco-friendly, efficient technology or complement to their synthetic counterparts for enhancing stress tolerance (Rouphael et al. 2021). Though the efficacy of both PGPB and phytochemicals is demonstrated in various field studies, their interactive effect is rarely reported. The results of this investigation elucidated that PGPR + foliar applied SORGAAB at V3 were highly effective in improving morphological (Fig. 2) and physiological traits (Table 2) and significantly helped the maize plants counter the biochemical oxidative stress, i.e., excessive H2O2 production, and subsequent mitigation through promoting the antioxidant machinery (45–58% higher production than CK2).
Moreover, the correlation analysis also confirms that H2O2 production has a highly strong association with other antioxidant phytochemicals (Fig. 4). Even the role of individual rhizobacterial strain effects on plant growth promotion has never been questionable. Recently, Ahmad et al. (2019) showed that Pseudomonas fluorescens (Pf1) and P. putida (Pp) KT2440 strains effectively lowered the drought-induced ROS in Zea mays during the first three to six days of growth compared to control treatment. Their real-time quantitative PCR analysis showed that 6 days after sowing (DAS) of drought stress significantly down-regulated the cold-related WRKY18 gene (dehydrin 410 gene). These genes are responsive to drought stress and are regulated by 7 days of continuous drought. Lowering of lipoxygenase genes in plants that had been infected on day six after infection showed that Pf1 and Pp help remove ROS during drought-stress conditions. Similarly, the individual treatment of phytochemicals has also recently proven to be highly effective for the modulation of crop physiology and morphology under drought stress. According to Khaliq et al. (2022), the application of 2.0% brassica water extract improved the morphological characteristics (seedling length, fresh and dry weights) of both wheat cultivars in the range of 2–160% under 30% FC.
To maintain the homeostatic balance, plants must adapt to environmental hazards (drought, heat, salinity etc.) by changing their morphological, physiological, and molecular responses (Ojuederie et al. 2019). In this study, it is evident that maize plants went through significant morphological adaptations to counter water scarcity. The root-to-shoot ratio was the highest for the plants stimulated with PGPB + SORGAAB treatments under drought conditions. On both sides, either on weight bases or length bases, the root-to-shoot ratio was higher, inferring that under drought conditions, plants’ roots were more stimulated to grow extensively in search of water. Under drought conditions, this morphological adaptation is a plant’s response to boost primary root growth and produce little roots with root hairs, which will give a larger absorptive surface and increase the uptake of available water (Carlson et al. 2020). Our results also inferred that PGPB + SORGAAB resulted in extensive stimulation, which thus enhanced the root and shoot biomass due to excessive nutrition, hormones, and water uptake, as evident by higher fresh and dry biomass. A study by Carlson et al. (2020) also showed that PGPB strains could change the shape of sorghum plants’ roots by increasing the production of gibberellic acid, indole acetic acid, and cytokinin during three weeks of drought stress.
As a key rhizospheric player in crop production, the PGPB treatments have also been able to promote soil fertility levels that might be due to the acidification of rhizoplane soil (Pereira et al. 2020), which resulted in higher availability of macronutrients (44–90% higher) and micronutrients (44–95% higher) than the control treatment. The enhanced availability, and uptake, especially of the micronutrients, were due to nutrient recycling, high water absorption by the modified root’s structure, and polysaccharide associations that ultimately supported plant growth and activation of the antioxidant system (Pereira et al. 2020, Saberi-Riseh et al. 2021). This is also evident from the pictorial illustration that plants growing under Drought + PGPB + SORGAAB treatment did not show any sign of retorted or stunted growth or any nutritional deficiency symptoms. The interactions of different plant species, root exudates, soil characteristics, and many other elements occur in the rhizosphere, a special ecological niche that determines the composition of microbial communities (Niu et al. 2018). When compared to the microbial communities in the bulk soil, the rhizosphere microbial communities may be more active and unique due to the organic compounds generated from roots’ high microbial biomass and metabolic activity. This seems true, as the CFU measurement shows that the microbial population in the Drought + PGPB + SORGAAB treatment was 71% in the soil. It has been previously documented how useful bacterial strains can be for crop productivity in maintaining proper levels of mineral minerals, particularly phosphorus (Kour et al. 2020). It was found that adding PGPR microbes to the roots of Avena sativa, Medicago sativa, and Cucumis sativus increased the activity of soil-urease (SURE), invertase (SIN), alkaline phosphatase (SAKP), catalase (SCAT), available nitrogen (AN), available phosphorus (AP), available potassium (AK), and organic carbon (SOC) (Li et al. 2020). In another study, Çığ et al. (2021) reported that commercial Bacillus subtilis and Paenibacillus azotofixans PGPR applications increased K, Ca, Mg, Fe, and Ni contents in wheat variety (Triticum monococcum L.). Recently, Pereira et al. (2020) demonstrated that PGPB application of two strains, PGPR Cupriavidus necator 1C2 (B1) and Pseudomonas fluorescens S3X (B2), resulted in an 89% increase in shoot biomass due to 32.41% and 35.21% higher nitrogen use efficiency (NUE) and phosphorus use efficiency (PUE) in maize plants growing at 60% FC, respectively, compared to non-inoculated plants.
Sorghum water extract is rich in phenolics, which have been shown to operate as a balancing agent for redox homeostasis. Most studies support the reducing power of SORGAAB, a hydrogen donor and quencher of singlet oxygen in cellular compartments, thereby acting as an efficient ROS scavenger (Khaliq et al. 2022; Tudu et al. 2022). When used as antioxidants, phenolics promote oxygenation processes and inhibit the activity of several oxidizing enzymes (Rouphael et al. 2021). PGPB are good for helping plants grow better when they are stressed by drought. This is because rhizobacteria can make exopolysaccharides like cellulose and alginate, which have been shown to make plants more resistant to drought (Kour et al. 2020). Exopolysaccharides may therefore be essential in reducing the impacts of stress on both plant and microbial populations under drought circumstances (Kour et al. 2020).
In this study, PGPB plus foliar SORGAAB application significantly increased the amylase activity, chlorophyll a, b, hydrogen peroxide, catalysis, superoxide, dismutase, and peroxidase under drought stress (Tables 1 and 2). We investigated how changes in the photosynthetic pigments, biochemical responses, and nutritional linkages in the leaves of drought-stressed maize could lead to better growth characteristics (Tables 1 and 2). For optimum plant growth, the light and dark responses of photosynthesis must function optimally, and these phenomena are substantially hampered by drought stress in this study results. Antioxidants like SOD, POD, and CAT work faster when SORGAAB is applied as foliar spray in addition to PGPB as soil inoculant, and consequently the ROS’ negative impacts on cellular organelles are decreased. Previous studies shows that due to the phenolic-based scavenging effect, foliar-applied SORGAAB dramatically increased antioxidant activity and decreased the H2O2 levels in wheat leaves (Ibrahim et al. 2022; Zhang et al. 2018). Therefore, the detoxification of ROS that are controlled by the action of phenolic as a singlet oxygen quencher and hydrogen donor are linked to the process of increasing stress tolerance in grass species (Tudu et al. 2022).
Similarly, the phytochemicals like application of brassica water extract triggered the activities of antioxidant enzymes, including SOD, CAT, and POD (11–159%), decreased the H2O2 content (14–30%), and enhanced chlorophyll a and b and carotenoid contents (19–154%), as compared to the control, in 2 wheat cultivars under 30% field capacity (Khaliq et al. 2022). Chiappero et al. (2019) also demonstrated 30% and 60% enhancement of SOD and POD activity in Mentha piperita at 35% FC by two PGPR species (Pseudomonas fluorescens WCS417R and Bacillus amyloliquefaciens GB03). In both studies, the maintenance of redox homeostasis supported the plants’ ability to grow optimally under drought conditions. Our correlation analysis also confirms that the stress tolerance percentage is conferred by the improvement in physiological and biochemical traits, and the stress tolerance further confirms the highest yield of maize. Similarly, the application of two drought-adaptive P-solubilizing strains (EU-LWT3-69 and EU-F10) enhanced the Chl. a and b contents by a 2–4-fold increase in the sorghum plant under 50% drought conditions (Kour et al. 2020). In comparison to untreated plants and those with PGPR-HAS31, they retained higher levels of the photosynthetic process, chlorophyll content, soluble proteins, total soluble sugars, and enzyme activity of CAT, POD, and SOD (Batool et al. 2020).
Moreover, the exponential model has explained that 75% yield is attributed to maize plants’ ability to adapt to drought stress (Fig. 3), which is eventually conferred by the mitigation of ROS species (H2O2) through the highest production of AmyL, Chl. a, Chl. b, and CP (Table 2 and Fig. 4). Yet, the drought stress adoption strategy of maize plants is highly linked to the genotypic characteristics of maize (Saberi-Riseh et al. 2021; Chen et al. 2016). The literature is also corroborated by our findings on the ROS mitigation potential of PGPB and phytochemicals’ bio-stimulation. The application of PGPR + exogenously applied SORGAAB was more pronounced in drought tolerance, owing primarily to its inherent ability to increase the scavenging action of ROS and alter the plant morphology.
Conclusion
In conclusion, application of PGPB as soil amendment and foliar SORGAAB at third leaf stage of maize under the drought environment showed encouraging results. The enhanced ROS scavenging activity can be considered as leading way of maize plant for drought stress tolerance. The plants were able to gain optimum photosynthesis rate by enhancing the chlorophyll-a and b contents by 67 and 83%, respectively, that resulted in achieving higher agronomic biomass. All other biochemical responses i.e. amylase activity, catalase, superoxide dismutase, peroxidase and crude protein were significantly enhanced. Our results also demonstrates that individual application of PGPB had an influential impact on both soil properties and plant productivity but lesser than the combined application with SORGAAB. Therefore, SORGAAB proved an effective bio-stimulant that triggers the biochemical response in the plant to sequester the ROS generated under drought stress. Therefore, results strongly imply that the application of both PGPB and foliar SORGAAB could alleviate drought stress and help achieve optimum production in arid regions. However, further field verifications of integrated utilization of PGPB and SORGAAB are necessary as the efficacy may vary with crop species, and field conditions. Moreover, economic efficiency comparisons to commercial biostimulants are also imperative to be evaluated.
Data availability
The authors confirm that the whole data supporting the findings of this study is available within the article however, raw data generated from this research can be available on reseanable request to corresponding author.
References
Afzal I, Akram MW, Rehman HU et al (2020) Moringa leaf and sorghum water extracts and salicylic acid to alleviate the impacts of heat stress in wheat. S Afr J Bot 129:169–174. https://doi.org/10.1016/j.sajb.2019.04.009
Ahmad I, Zaib S, Alves PCMS et al (2019) Molecular and physiological analysis of drought stress responses in Zea mays treated with plant growth promoting rhizobacteria. Biol Plant 63:536–547. https://doi.org/10.32615/bp.2019.092
Amrutha S, Parveen ABM, Muthupandi M, Sivakumar V et al (2019) Variation in morpho-physiological, biochemical, and molecular responses of two Eucalyptus species under short-term water stress. Acta Bot Croat 78(2):125–134. https://doi.org/10.2478/botcro-2019-0021
Arnon DI (1949) Copper enzymes in isolated chloroplasts. polyphenol oxidase in beta vulgaris. Plant Physiol 24:1–15. https://doi.org/10.1104/pp.24.1.1
Aslam M, Nasim F, Ruhi R, Murad H, Ejaz S, Choudhary M, Mustafa G, Ashraf M, Rehman J (2018) Isolation and characteristics of biotechnologically important antagonistic thermophilic bacteria from rhizosphere of Haloxylon salicornicum. Pol J Microbiol 67(1):49–58. https://doi.org/10.5604/01.3001.0011.6142
Bajwa AA, Farooq M, Nawaz A (2018) Seed priming with sorghum extracts and benzyl aminopurine improves the tolerance against salt stress in wheat (Triticum aestivum L.). Physiol Mol Biol Plants 24:239–249. https://doi.org/10.1007/s12298-018-0512-9
Batool T, Ali S, Seleiman MF et al (2020) Plant growth promoting rhizobacteria alleviates drought stress in potato in response to suppressive oxidative stress and antioxidant enzymes activities. Sci Rep 10:16975. https://doi.org/10.1038/s41598-020-73489-z
Bernfeld P (1955) [17] Amylases, α and β. Methods Enzymol 1:149–158. https://doi.org/10.1016/0076-6879(55)01021-5
Bremner JM, Mulvaney CS (1982) “Nitrogen-Total”-Methods of soil analysis: part 2 chemical and microbiological properties. Agron Monogr 9:595–624. https://doi.org/10.2134/agronmonogr9.2.2ed.c31
Carlson R, Tugizimana F, Steenkamp PA et al (2020) Rhizobacteria-induced systemic tolerance against drought stress in Sorghum bicolor (L.) Moench. Microbiol Res 232:126388. https://doi.org/10.1016/j.micres.2019.126388
Chance B, Maehly AC (1955) [136] Assay of catalases and peroxidases. Methods Enzymol 2:764–775. https://doi.org/10.1016/S0076-6879(55)02300-8
Chen D, Wang S, Cao B et al (2016) Genotypic variation in growth and physiological response to drought stress and re-watering reveals the critical role of recovery in drought adaptation in maize seedlings. Front Plant Sci 6:1241. https://doi.org/10.3389/fpls.2015.01241
Chiappero J, del Rosario Cappellari L, Sosa Alderete LG et al (2019) Plant growth promoting rhizobacteria improves the antioxidant status in Mentha piperita grown under drought stress leading to an enhancement of plant growth and total phenolic content. Ind Crops Prod 139:111553. https://doi.org/10.1016/j.indcrop.2019.111553
Çığ F, Sönmez F, Nadeem MA, El Sabagh A (2021) Effect of biochar and PGPR on the growth and nutrients content of einkorn wheat (Triticum monococcum L.) and post-harvest soil properties. Agronomy 11:2418. https://doi.org/10.3390/agronomy11122418
Daryanto S, Wang L, Jacinthe PA (2016) Global synthesis of drought effects on maize and wheat production. PLoS ONE 11:0156362–0156362. https://doi.org/10.1371/journal.pone.0156362
El Sabagh A, Hossain A, Barutçular C et al (2018) Sustainable maize (Zea mays L.) production under drought stress by understanding its adverse effect, survival mechanism and drought tolerance indices. J Exp Biol Agric Sci 6:282–295. https://doi.org/10.18006/2018.6(2).282.295
FAOSTAT (2021) FAOSTAT-Food and Agriculture Organization Rome. Data Retrieve from: http://www.fao.org/faostat/en/
Farooq M, Nadeem F, Arfat MY et al (2018) Exogenous application of allelopathic water extracts helps improving tolerance against terminal heat and drought stresses in bread wheat (Triticum aestivum L. Em. Thell.). J Agron Crop Sci 204:298–312. https://doi.org/10.1111/jac.12261
Farooqi ZUR, Zia Ur Rehman M, Sohail MI et al (2020) Regulation of drought stress in plants. In: Tripathi DK (ed) Plant life under changing environment; responses management. Academic Press, New York, NY, pp 77–104. https://doi.org/10.1016/B978-0-12-818204-8.00004-7
Fernandez GC (1992) Effective selection criteria for assessing plant stress tolerance. In Proceeding of the international symposium on adaptation of vegetables and other food crops in temperature and water stress, Shanhua, Taiwan, 13–16 Aug, pp 257–270. https://doi.org/10.22001/wvc.72511
Giannopolitis CN, Ries SK (1977) Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol 59:309–314. https://doi.org/10.1104/pp.59.2.309
GOP (2021). Government of Pakistan, Economic survey of Pakistan 2020–2021. Chapter 2-Agriculture. Available at https://www.finance.gov.pk/survey_2021.html Accessed 4 July 2022
Hafez EM, Osman HS, Gowayed SM et al (2021) Minimizing the adversely impacts of water deficit and soil salinity on maize growth and productivity in response to the application of plant growth-promoting rhizobacteria and silica nanoparticles. Agronomy 11:676. https://doi.org/10.3390/agronomy11040676
Helmke PA, Sparks DL (2018) Lithium, sodium, potassium, rubidium, and cesium. In: Sparks DL, Page AL, Helmke PA, Loeppert RH, Soltanpour PN, Tabatabai MA, Johnston CT, Sumner ME (eds) Methods of soil analysis: part 3 chemical methods. Soil Science Society of America, American Society of Agronomy, Madison, WI, pp 551–574. https://doi.org/10.2136/sssabookser5.3.c19
Ibrahim MU, Khaliq A, Hussain S, Murtaza G (2022) Sorghum water extract application mediates antioxidant defense and confers drought stress tolerance in wheat. J Plant Growth Reg 41(2):863–874. https://doi.org/10.1007/s00344-021-10345-y
Janssen PH, Yates PS, Grinton BE et al (2002) Improved culturability of soil bacteria and isolation in pure culture of novel members of the divisions Acidobacteria, Actinobacteria, Proteobacteria, and Verrucomicrobia. Appl Environ Microbiol 68:2391–2396. https://doi.org/10.1128/AEM.68.5.2391-2396.2002
Khaliq A, Ibrahim MU, Hussain S et al (2022) The hermetic effects of a Brassica water extract triggered wheat growth and antioxidative defense under drought stress. Appl Sci 12:4582. https://doi.org/10.3390/app12094582
Kour D, Rana KL, Kaur T et al (2020) Microbe-mediated alleviation of drought stress and acquisition of phosphorus in great millet (Sorghum bicolour L.) by drought-adaptive and phosphorus-solubilizing microbes. Biocatal Agric Biotechnol 23:101501. https://doi.org/10.1016/j.bcab.2020.101501
Krishna Motukuri SR (2020) Quality protein maize: an alternative food to mitigate protein deficiency in developing countries. In: Hossain A (ed) Maize – production and use. IntechOpen, Rijeka, pp 45–72. https://doi.org/10.5772/intechopen.89038
Li H, Qiu Y, Yao T et al (2020) Effects of PGPR microbial inoculants on the growth and soil properties of Avena sativa, Medicago sativa, and Cucumis sativus seedlings. Soil Tillage Res 199:104577. https://doi.org/10.1016/j.still.2020.104577
Nabi G, Ali M, Khan S, Kumar S (2019) The crisis of water shortage and pollution in Pakistan: risk to public health, biodiversity, and ecosystem. Environ Sci Pollut Res 26:10443–10445. https://doi.org/10.1007/s11356-019-04483-w
Niu X, Song L, Xiao Y, Ge W (2018) Drought-tolerant plant growth-promoting rhizobacteria associated with foxtail millet in a semi-arid and their potential in alleviating drought stress. Front Microbiol 8:2580. https://doi.org/10.3389/fmicb.2017.02580
Ojuederie OB, Olanrewaju OS, Babalola OO (2019) Plant growth promoting rhizobacterial mitigation of drought stress in crop plants: Implications for sustainable agriculture. Agronomy 9:712. https://doi.org/10.3390/agronomy9110712
Osakabe Y, Osakabe K, Shinozaki K, Tran L-SP (2014) Response of plants to water stress. Front Plant Sci 5:86. https://doi.org/10.3389/fpls.2014.00086
Paravar A, Piri R, Balouchi H, Ma Y (2023) Microbial seed coating: an attractive tool for sustainable agriculture. Biotechnol Rep 37:00781–00781. https://doi.org/10.1016/j.btre.2023.e00781
Pereira SIA, Abreu D, Moreira H et al (2020) Plant growth-promoting rhizobacteria (PGPR) improve the growth and nutrient use efficiency in maize (Zea mays L.) under water deficit conditions. Heliyon 6:05106–05106. https://doi.org/10.1016/j.heliyon.2020.05106
Ray DK, Sloat LL, Garcia AS et al (2022) Crop harvests for direct food use insufficient to meet the UN’s food security goal. Nat Food 3:367–374. https://doi.org/10.1038/s43016-022-00504-z
Rouphael Y, Corrado G, Colla G et al (2021) Biostimulation as a means for optimizing fruit phytochemical content and functional quality of tomato landraces of the San Marzano area. Foods 10:926. https://doi.org/10.3390/foods10050926
Saberi-Riseh R, Ebrahimi-Zarandi M, Gholizadeh Vazvani M, Skori YA (2021) Reducing drought stress in plants by encapsulating plant growth-promoting bacteria with polysaccharides. Int J Molec Sci 22(23):12979. https://doi.org/10.3390/ijms222312979
Scavo A, Abbate C, Mauromicale G (2019) Plant allelochemicals: agronomic, nutritional and ecological relevance in the soil system. Plant Soil 442:23–48. https://doi.org/10.1007/s11104-019-04190-y
Shakoor U, Rashid M, Saboor A et al (2017) Maize production response to climate change in Pakistan: a time series assessment. Sarhad J Agric 33:320–330. https://doi.org/10.17582/journal.sja/2017/33.2.320.330
Simova-Stoilova L, Pecheva D, Kirova E (2020) Drought stress response in winter wheat varieties–changes in leaf proteins and proteolytic activities. Acta Bot Croat 79(2):121–130. https://doi.org/10.37427/botcro-2020-018
Sohail MI, Zia Ur Rehman M, Aziz T et al (2022) Iron bio-fortification and heavy metal/(loid)s contamination in cereals: successes, issues, and challenges. Crop Pasture Sci 73:877–895. https://doi.org/10.1071/CP21771
Tripathi AM, Pohanková E, Fischer M et al (2018) The evaluation of radiation uses efficiency and leaf area index development for the estimation of biomass accumulation in short rotation poplar and annual field crops. Forests 9:168. https://doi.org/10.3390/f9040168
Tudu C, Dey A, Pandey DK, Panwar JS, Nandy S (2022) Role of plant derived extracts as biostimulants in sustainable agriculture: a detailed study on research advances, bottlenecks and future prospects. In: Singh HB, Vaishnav A (eds) New and future developments in microbial biotechnology and bioengineering. Elsevier, Amsterdam, Netherlands, pp 159–179. https://doi.org/10.1016/B978-0-323-85579-2.00017-4
Velikova V, Yordanov I, Edreva A (2000) Oxidative stress and some antioxidant systems in acid rain-treated bean plants protective role of exogenous polyamines. Plant Sci 151:59–66. https://doi.org/10.1016/S0168-9452(99)00197-1
Walkley A, Black IA (1934) An examination of the DEGTJAREFF method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci 37:29–38. https://doi.org/10.1097/00010694-193401000-00003
Yang C, Zhang D, Li X et al (2021) Drought effects on photosynthetic performance of two wheat cultivars contrasting in drought. New Zeal J Crop Hortic Sci 49:17–29. https://doi.org/10.1080/01140671.2020.1851264
Zhang J, Jiang H, Song X et al (2018) The responses of plant leaf CO2/H2O exchange and water use efficiency to drought: a meta-analysis. Sustain 10:551. https://doi.org/10.3390/su10020551
Acknowledgements
We are indebted to the Higher Education Commission of Pakistan for a grant of PKR 1 million under the Start-Up Research Grant Program (SRGP/412/2020/65).
Author information
Authors and Affiliations
Contributions
Raza Ullah: project administration, conceptualization, funding acquisition, methodology, investigation, formal analysis, Muhammad Irfan Sohail: Conceptualization, methodology, visualization, investigation, formal analysis, writing-original draft, writing—review & editing, Ayesha Siddiqui: writing—review & editing, Muhammad Rahil Afzal: methodology, investigation, writing-original draft, Shoaib ur Rehman: writing—review & editing, Muhammad Kamran: writing—review & editing, Hafiz Tassawar Abbas: writing-original draft, Muhammad Awais Khalid: formal analysis, writing—review & editing, Esha Jabbar: writing—review & editing, visualization.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could influence the work reported in this paper.
Additional information
Communicated by P.K. Nagar.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Ullah, R., Siddiqui, A., Rehman, S.U. et al. Mitigation of drought stress in Zea mays L. through plant growth-promoting rhizobacteria assisted by foliar sorghum water extract. Acta Physiol Plant 46, 65 (2024). https://doi.org/10.1007/s11738-024-03690-4
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11738-024-03690-4