
Abstract
The global agricultural productivity has been significantly impaired due to the extensive use of heavy metal. Cadmium (Cd) is now recognized as a significant soil and environmental contaminant that is primarily spread by human activity. This study investigates the possible impact of melatonin (ME) in mitigating the toxicity caused by Cd in pepper (Capsicum annuum L.) seedlings. There were three groups of plants used in the experiment: control (CK) plants, Cd-stressed plants and ME-pretreated + Cd-stressed plants. The concentration of ME and Cd was 1 µM and 0.1 mM, respectively, and applied as root application. The results described that Cd treatment remarkably reduced growth parameters, impaired pigment concentration, hindered gas exchange traits. In contrast, ME supplementation significantly recovered these parameters by increase in growth and biomass production of pepper seedlings under Cd toxicity. In addition, ME application considerably increased osmolyte production and protein level in pepper leaves and roots. Furthermore, ME positively upregulated the antioxidant enzymes activity and effectively decreased the oxidative damage in pepper leaves and roots. The enhanced antioxidant enzymes activity performed a significant role in the reduction of H2O2 and MDA concentration in plants under Cd stress. The findings indicated that the application of ME to plants effectively alleviates the stress caused by Cd exposure. Moreover, ME demonstrates significant efficacy in mitigating the adverse impacts of Cd on pepper plants.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The issue of heavy metal contamination in agricultural soils has garnered global recognition in recent times owing to its adverse effects on environmental well-being, crop integrity, and food safety (Adrees et al. 2015; Qin et al. 2022; Altaf et al. 2022a). Eliminating heavy metal contamination in agricultural soils and heavy metal toxicity in plants is the aim of worldwide agricultural and environmental research. Heavy metal pollution of soil has increased in recent years as a result of the continuous transit, deposition, and transformation of heavy metals within the environment (Altaf et al. 2021a; Zhang et al. 2021; Ghuge et al. 2023). Cadmium (Cd) is now more widely recognized as a hazardous heavy metal due to its negative effects on agricultural yield and the potential health risks associated with food chain contamination (Benavides et al. 2005). Cd is well recognized as an environmental risk due to its extended half-life in soil, elevated solubility in water, and the escalating levels of human activity on a global scale (Keltjens and Van Beusichem 1998; Wani et al. 2007). The presence of an excessive amount of Cd in the soil system has been shown to impede plant development and results in various physiological, morphological, anatomical, and metabolic issues (Zhu et al. 2021). Nonetheless, the adverse impacts of Cd may persist through successive plant generations, resulting in reduced seed germination and viability in plants cultivated in soils polluted with Cd (Carvalho et al. 2018). The toxicity of Cd is often associated with its concentration in plant tissues. It is plausible to hypothesize that variations in the absorption, transportation, and accumulation of Cd might contribute to the diverse levels of tolerance seen across different cultivars of a closely related species when subjected to Cd exposure (Altaf et al. 2022b).
Plants have evolved effective systems to regulate the absorption of Cd and alleviate the adverse effects associated with Cd toxicity (Gallego et al. 2012). The roots are the main elements subjected to adverse effects heavy metals such as Cd (Sigel et al. 2013). The increase of reactive oxygen species (ROS) induced by Cd stress may serve a dual role in plant cells, acting both as a detrimental result of metabolic processes and as significant modulators of growth, development, and defense mechanisms (Piqueras et al. 1999; Sharma and Dietz 2009; Li et al. 2022). For example, the presence of ROS has been seen to induce lipid peroxidation, leading to oxidative damage of various macromolecules and subsequent cellular demise (Foy et al. 1978; DalCorso et al. 2008). Plant cells possess enzymatic antioxidant defense systems that are responsible for scavenging ROS to maintain redox equilibrium. To ameliorate the adverse consequences of Cd toxicity, one potentially effective approach might include the use of phytohormones. These substances have the potential to enhance the ability of plants to withstand and recover from detrimental stressors.
Melatonin (ME) is a naturally occurring novel plant growth regulator (Ahmad et al. 2023). ME regulates seedling growth, photosynthetic apparatus, osmolytes accumulation, root architecture system, balanced redox homeostasis, mineral nutrient uptake, ripening, decreasing heavy metals uptake, reducing oxidative damage, and increasing antioxidant defense mechanism (Tiwari et al. 2021; Liang et al. 2018; Altaf et al. 2023). Recent research in plant physiology has focused on the physiological role of ME, specifically in plants grown with synthesized ME molecules or elevated endogenous melatonin levels (Kaya et al. 2019). ME has been shown to enhance plant resilience to many abiotic stressors, such as salinity, drought, cold, heat, heavy metal toxicity, and (UV) radiation (Nawaz et al. 2018; Devi et al. 2021; Altaf et al. 2022c). Exogenous ME application efficiently protected leaf photosynthetic system, biomass production, and antioxidant profile in tomato plants under Cd stress (Hasan et al. 2015). In another study, under Cd toxicity, ME supplementation increased nutrient accumulation and modified the root structure of tomato species (Altaf et al. 2022b). Furthermore, ME restricts Cd uptake in the shoot of tomato, Arabidopsis, and strawberry (Hasan et al. 2015; Gu et al. 2017; Saqib et al. 2023). Under boron toxicity, ME application remarkably improved pepper growth, net photosynthetic rate, photosynthetic pigment concentration, and ion homeostasis level in pepper leaves (Sarafi et al. 2017).
Pepper (Capsicum annuum L.) is one of the most commonly consumed horticultural crop (Korkmaz et al. 2021). It is considered a worldwide significant cash crop because of its nutritional and economic significance. The pepper fruit is an excellent source of several different types of nutrients, including phenolic compounds, antioxidants, vitamins, proteins, and carbohydrates. (Howard et al. 2000). Pepper plants have a moderate level of sensitivity towards metal toxicity such as Cd (Sarafi et al. 2017; Kaya et al. 2020; Altaf et al. 2021b). A plethora of research has been performed to investigate the diverse impacts of melatonin on mitigating environmental stresses in various agricultural crops (Gu et al. 2017; Chen et al. 2018; Cao et al. 2019; Jahan et al. 2020; Park et al. 2021; Saqib et al. 2023). However, to the best of our knowledge, more research is necessary to determine how ME affects Cd stress in pepper. In light of the aforementioned factors, the primary objective of this study was to investigate the impact of ME on the growth and photosynthetic efficiency of pepper seedlings subjected to Cd-induced stress. Additionally, the study aimed to examine the influence of ME on the antioxidant enzymatic system in both the leaves and roots of Cd-stressed pepper seedlings.
Materials and methods
Experimental setup
Seeds of pepper genotypes (Super Shimla and Ganga) were sown in seedling trays filled with nutrient-rich soil. After 4 weeks of sowing, identical plants were shifted into the black plastic pots and filled with nutrient-rich soil. Following the transplantation, the seedlings were subjected to a 12-day period of adaptation under standard growth conditions to facilitate adjustment to the new environment. The application of ME (1 µM; 200 ml per plant) to the root was conducted at an interval of every 3rd day and persisted for a duration of 12 days. Following a 12-day period of pretreatment with ME, a solution containing CdCl2 at a concentration of 0.1 mM (with a volume of 200 ml per plant) was supplied to the plants. The control (CK) seedlings were subjected to regular irrigation using the Hoagland nutrient solution (pH 5.5 ± 0.1) without the addition of CdCl2 or ME. There were ten plants included in each treatment, and each treatment was replicated three times. The plants were subjected to Cd-induced stress for a duration of 4 weeks, with separate groups receiving either treatment alone or a combination of Cd application and ME. Following this period, samples were collected from the plants to assess a range of growth and physiological parameters. After stress treatment, the collected samples were promptly subjected to treatment with liquid nitrogen and then kept at a temperature of −80 °C for further analysis. The level of Cd and ME used in this experiment was determined based on prior research (Korkmaz et al. 2021; Kaya et al. 2020). The treatments were as follows: (1) CK (control); (2) Cd stress; (3) ME treatment + Cd stress. This study was carried out under controlled environmental conditions as previously described in our study (Saqib et al. 2023).
Plant growth
Different plant growth indicators, such as the fresh and dry weights of the leaves and roots, were measured 4 weeks after the application of Cd. The fresh weight of the plants was promptly measured using an electronic weighing scale immediately after harvesting. To determine the dry weight, samples were subjected to oven-drying at a temperature of 80 °C until a constant weight was achieved (Altaf et al. 2019).
Photosynthesis parameters
The determination of leaf chlorophyll content included the homogenization of fresh leaves (0.5 g) in a solution of cold acetone (80%, 10 ml). Afterward, the resulting mixture was subjected to centrifugation at a speed of 12,000 rpm for a duration of 10 min. The absorbance of the supernatant was measured at wavelengths of 645 nm, 663 nm, and 449 nm using a UV–Vis spectrophotometer (WE, 6000, China) to quantify the levels of chlorophyll a, chlorophyll b, and carotenoids, respectively (Lichtenhaler and Wellburn 1983). The relative chlorophyll content (as measured by the SPAD index) of fully expanded young leaves was assessed using a SPAD-502 m (Minolta, Japan). An infrared gas analyzer (ADC, Bio Scientific Ltd. UK) was used to determine the gas exchange parameters [net photosynthetic rate (PN), transpiration rate (E), stomatal conductance (gs), and intercellular CO2 concentration (ci)]. These gas exchange parameters were measured according the procedure of Jahan et al. (2020).
Determination of total soluble proteins (TSP) and total phenolic content (TPC)
Total soluble proteins were isolated from pepper leaves and roots using a technique based on that developed by Sambrook and Russell (2006), with some modifications. The quantification of total soluble protein content was conducted using the approach described by Bradford (1976). The quantification of the total phenolic content in pepper leaves was conducted using the methodology outlined by Singleton and Rossi (1965).
Determination of H2O2 and MDA content
The plant sample weighing 0.1 g was subjected to extraction using a 0.1% TCA solution in a volume of 1 ml. The resulting mixture was then centrifuged at a speed of 12,000 rpm for a duration of 10 min. The determination of H2O2 concentration in pepper leaves and roots was conducted using the methods established by Velikova et al. (2000). The determination of MDA levels in pepper leaves and roots was conducted using the TBA technique, which was originally described by Heath and Packer (1968).
Determination of antioxidant enzymes
The sample (leaf or root) 0.3 g was fully pulverized in a solution of 50-mM sodium phosphate buffer (3 ml) at pH 7.8. The resulting mixture was then transferred to an Eppendorf tube. Subsequently, the samples were subjected to centrifugation at a speed of 12,000 rpm for a duration of 10 min. SOD, CAT, and POD activity was determined by adopting the Chance and Maehly’s (1955) methodology.
Statistical analysis
The program Origin Pro 2021 was used to statistically evaluate the collected data. The significant differences between the various treatments and cultivars were estimated using Duncan’s multiple range tests (DMRT). For each treatment, the means and related standard errors were computed for biological triplicates.
Results
Plant growth
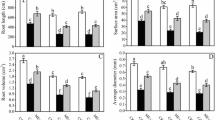
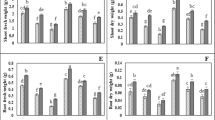
To investigate the impact of exogenous application of ME on Cd stress in two pepper genotypes, the roots were watered with a nutrient solution containing ME for a period of 12 d prior to Cd treatment. The findings shown in Fig. 1 demonstrate a distinct disparity seen in the growth of pepper seedlings when subjected to Cd toxicity, both with and without treatment with ME. When compared to CK plants, it was noticed that Super Shimla’s fresh shoot weight, dry shoot weight, fresh root weight, and dry root weight were all decreased by 60.89%, 56.45%, 63.41%, and 62.51%, respectively, while the Ganga seedlings grown under Cd-stress conditions saw reductions of 56.42%, 55.45%, 59.69%, and 61.27, respectively. In contrast, ME supplementation efficiently increased these growth characteristics in both pepper species under Cd stress (Fig. 2).

Investigation of the impact of exogenously applied melatonin on two pepper genotypes exposed to cadmium toxicity

Effect of melatonin on the growth characteristics in two pepper genotypes exposed to cadmium stress. Results are means ± standard error for n = 3. Furthermore, small alphabet letters show significant differences at P < 0.05 (DMRT)
Leaf photosynthesis parameters
The present results described that Cd application considerably reduced photosynthetic pigment concentration in pepper leaves (Fig. 3A–C). Conversely, ME supplementation profoundly increased the photosynthetic pigment level in both pepper species under Cd toxicity (Fig. 3A–C). Further, the relative chlorophyll content (SPAD index) was decreased by 59.64% and 56.51%; whereas ME supplementation increased the relative chlorophyll content by 64.61% and 55.02%, respectively, in Super Shimla and Ganga seedlings (Fig. 3D).

Effect of melatonin on pigment content and SPAD index in two pepper genotypes exposed to cadmium stress. Results are means ± standard error for n = 3. Furthermore, small alphabet letters show significant differences at P < 0.05 (DMRT)
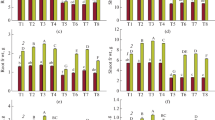
The current findings stated that gas exchange characteristics were dramatically damaged during Cd stress. Compared CK group, the gas exchange elements were reduced under Cd toxicity in the Super Shimla and Ganga seedlings, respectively. In contrast, ME application enhanced the Pn, Gs, Ci, and Tr by 95.81%, 64.01%, 49.68%, and 89.31%, respectively, in the Super Shimla cultivar and decreased by 75.87%, 55.81%, 75.21%, and 98.22%, respectively, in Ganga cultivar, compared to only Cd treatment (Fig. 4).

Effect of melatonin on gas exchange parameters in two pepper genotypes exposed to cadmium stress. Results are means ± standard error for n = 3. Furthermore, small alphabet letters show significant differences at P < 0.05 (DMRT)
Phenolic and protein content
Compared to the CK treatment, Cd treatment significantly improved the total phenolic content in leaves and roots of both pepper species. Importantly, ME supplementation further increased the total phenolic content in both species under Cd toxicity (Fig. 5A, B). Further, protein concentration decreased in leaves and roots of both species under Cd toxicity. On the other hand ME supplementation remarkably increased the protein concentration on leaves and roots of both pepper species under Cd toxicity, when compared with only Cd treatment (Fig. 5C, D).

Effect of melatonin on total phenolic and protein content in two pepper genotypes exposed to cadmium stress. Results are means ± standard error for n = 3. Furthermore, small alphabet letters show significant differences at P < 0.05 (DMRT)
H2O2 and MDA content
During Cd stress, the H2O2 and MDA concentrations were enhanced in the leaves and roots of both pepper species (Fig. 6). Conversely, ME supplementation dramatically decreased H2O2 11.88/17.21% and 19.37/18.93% in the leaves/roots of Super Shimla and Ganga seedlings. In addition, it reduced the concentration of MDA 21.16/20.39% and 25.19/20.97% in the leaves/root of Super Shimla and Ganga plants, respectively, when compared with Cd treatment (Fig. 6).

Effect of melatonin on oxidative stress biomarkers in two pepper genotypes exposed to cadmium stress. Results are means ± standard error for n = 3. Furthermore, small alphabet letters show significant differences at P < 0.05 (DMRT)
Antioxidant enzymes
Cd toxicity remarkably improved the antioxidant enzymes (SOD, CAT, and POD) activity in both leaves and roots of pepper species. In contrast, pepper leaves/roots antioxidant enzymes activity further increased by the application of ME under Cd toxicity (Fig. 7). The current results suggested that ME supplementation enhanced the plant defense system and reduced the oxidative damage in pepper species.

Effect of melatonin on antioxidant enzymes activity in two pepper genotypes exposed to cadmium stress. Results are means ± standard error for n = 3. Furthermore, small alphabet letters show significant differences at P < 0.05 (DMRT)
Discussion
The growth attributes of the pepper plants were significantly diminished as a result of exposure to Cd treatment. The use of ME has been shown to significantly enhance the growth characteristics of pepper plants. (Figs. 1, 2). The findings of our study agree with previous research, which confirmed that the growth characteristics of tomato, cucumber, and fava bean were significantly impaired by the presence of hazardous metals such as vanadium, copper, and arsenic, respectively. On the other hand, the addition of ME greatly strengthened the growth parameters (Cao et al. 2019; Siddiqui et al. 2020). Under iron stress, the growth of cucumber was increased by the application of ME (Ahammed et al. 2020). In addition, ME increased the growth rate of many plant species such as wheat, radish, and tomato (Tang et al. 2016; Hasan et al. 2018; Al-Huqail et al. 2020) under heavy metals toxicity. ME has been shown to impede the absorption of Cd in plants and mitigate the adverse effects of Cd toxicity (Li et al. 2016). These results indicate that the use of ME, might potentially serve as an effective approach for mitigating the detrimental impact of heavy metals.
Photosynthetic pigment and relative chlorophyll content were reduced by Cd treatment, On the other hand, ME supplementation restored the photosynthetic pigment concentration in pepper leaves under stress condition (Fig. 3). Similarly, ME foliar application significantly improved the photosynthetic pigment content in several plant species under stress environment (Zhang et al. 2017; Chen et al. 2018; Manafi et al. 2021; Altaf et al. 2022c). The findings of our study align with a previous published work (He et al. 2020). The findings of our study indicate a decrease in chlorophyll a and chlorophyll b levels in plants subjected to Cd-induced stress. However, the application of ME showed noticeable increase in chlorophyll content.Wu et al. (2021) and Saqib et al. (2023) reported a similar trend in strawberry seedlings under Cd toxicity. Previous research has shown that the decrease in chlorophyll content caused by stress conditions may be due to the increased accumulation of H2O2 inside the leaf tissues (Liang et al. 2015; Ni et al. 2018). Photosynthesis is regarded as a fundamental metabolic process in plants. Chlorophyll plays an essential role as the main ingredient in the process of photosynthesis (Jahan et al. 2021). In this study, ME supplementation profoundly increased the gas exchange parameters in both pepper species under Cd toxicity (Fig. 4A–D). Similarly, under lanthanum toxicity, net photosynthetic rate was increased by the application of ME (Siddiqui et al. 2019). Further, ME application enhanced gas exchange characteristics in pea plants (Ahmad et al. 2023). Nawaz et al. (2018) reported that vanadium toxicity effectively damage photosynthetic system in watermelon seedlings. However, ME application remarkably protected photosynthetic efficiency of watermelon seedlings under vanadium stress. Jahan et al. (2020), Ahammed et al. (2020), and Saqib et al. (2023) also reported similar findings.
Phenols are a class of secondary metabolites with antioxidant characteristics, which function as a supplementary mechanism in combating the harmful effects of free radicals. The total TPC in leaves and roots exhibited improvement under Cd toxicity conditions in both pepper species, particularly in the control group (CK). The supplementation of ME under Cd toxicity conditions resulted in an increased content of TPC in both pepper species (Fig. 5). Sarafi et al. (2017) confirmed that under boron toxicity, ME application significantly improved phenolic content in pepper leaves. In addition, ME supplementation decreased Cd accumulation in apple and increased phenolic concentration in apple (He et al. 2020). Under nickel toxicity, the phenolic concentration was increased in the leaves of tomato by the application of ME (Jahan et al. 2020). The protein content of plants is regulated by photosynthetic mechanisms in response to adverse environmental circumstances (Simkin et al. 2019). Furthermore, a significant correlation was established between the net photosynthetic rate and the levels of photosynthetic pigments and proteins in pepper plants that were pre-treated with ME and exposed to Cd toxicity. Altaf et al. (2022a) also documented similar findings in rice under vanadium toxicity.
Plants under stress environment exhibit an overproduction of ROS, which disrupt crucial metabolic pathways, resulting in cellular toxicity and ultimate cell death (Tiwari et al. 2020). In the present study, pepper plants subjected to Cd treatment exhibited elevated levels of H2O2 and MDA in both their leaves and roots. Higher accumulation of oxidative stress biomarkers caused oxidative damage, whereas ME application reduced these in pepper seedlings (Fig. 6). Similarly, these results are also reported in many plant species such as pepper, wheat, and strawberry (Altaf et al. 2021a, b; Al-Huqail et al. 2020; Saqib et al. 2023). Nawaz et al. (2018) reported that ME application reduced the excessive accumulation of H2O2 and MDA in watermelon leaves and roots under vanadium stress. In another study, Jahan et al. (2020) and Ahammed et al. (2020) revealed that ME foliar application considerably decreased the ROS level in cucumber and tomato under iron and nickel toxicity. A prior study conducted on mallow shown that the application of ME effectively mitigates the adverse effects of Cd-induced phytotoxicity and enhances the growth and development of plants. In addition, ME significantly improved the pigments contents, increased antioxidant enzymes activity, and reduced oxidative damage in plants (Tousi et al. 2020). The antioxidant system plays a crucial role in plants by effectively reducing ROS and facilitating the development of metal tolerance (Imtiaz et al. 2015).
ME is a strong antioxidant molecule, and scientific studies have shown its ability to boost enzyme activity in several plant species when subjected to stress (Arnao and Hernández-Ruiz 2019; Park et al. 2021; Zhao et al. 2021). The current findings indicate that in pepper plants exposed to Cd toxicity, there was regulation of ROS overproduction and an increase in the activity of antioxidant enzymes due to the application of ME (Fig. 7). The findings presented in this study are consistent with the results published by Ahammed et al. (2020) and Nawaz et al. (2018), who similarly observed that the supplementation of ME enhanced the activity of antioxidant enzymes and mitigated the excessive generation of ROS in cucumber and watermelon seedlings. Furthermore, ME application efficiently maintained the antioxidant enzymes defense system in many plant species such as Vicia faba, radish, and pepper (Tang et al. 2016; Sarafi et al. 2017; Siddiqui et al. 2020) under heavy metals toxicity. Numerous studies have provided evidence for the significance of ME in the plant’s defense mechanism, and the application of exogenous ME has been shown to mitigate oxidative damage caused by stress. In agriculture, exogenous ME is effective for preventing the decrease of stress-related crop damage. ME is a cost-effective, chemically stable, ecologically sustainable, and readily obtainable compound that has the potential to safeguard plants against many environmental threats, primarily by mitigating the accessibility of contaminants, notably heavy metals.
Conclusion
The Cd treatment exhibited significant reductions in growth and photosynthetic activity, accompanied by an excessive accumulation of ROS. In contrast, ME application potentially improved seedling growth, photosynthetic pigment content, photosynthetic assimilation rate, and total phenolic content (Fig. 8). Further, ME supplementation hindered ROS accumulation and increased antioxidant enzymes activity in pepper species. Exogenous ME pretreatment have the potential to mitigate the Cd toxicity in plant. To optimize plant performance under Cd stress and promote the production of nutritious food, it is essential to conduct a comprehensive investigation of the impact of melatonin on reducing the translocation of Cd from roots to shoots. In conclusion, the use of ME treatment effectively mitigated the adverse effects of Cd-induced phytotoxicity in both pepper genotypes, leading to enhanced tolerance to Cd-induced stress.

An overview of the pepper seedlings’ response to ME-induced cadmium stress
Data availability
The datasets are available on reasonable request.
References
Adrees M, Ali S, Rizwan M, Zia-ur-Rehman M, Ibrahim M, Abbas F, Farid M, Qayyum MF, Irshad MK (2015) Mechanisms of silicon-mediated alleviation of heavy metal toxicity in plants: a review. Ecotoxicol Environ Saf 119:186–197
Ahammed GJ, Wu M, Wang Y, Yan Y, Mao Q, Ren J, Ma R, Liu A, Chen S (2020) Melatonin alleviates iron stress by improving iron homeostasis, antioxidant defense and secondary metabolism in cucumber. Sci Hortic 265:109205
Ahmad R, Alsahli AA, Alansi S, Altaf MA (2023) Exogenous melatonin confers drought stress by promoting plant growth, photosynthetic efficiency and antioxidant defense system of pea (Pisum sativum L.). Sci Hortic 322:112431
Al-Huqail AA, Khan MN, Ali HM, Siddiqui MH, Al-Huqail AA, Al Zuaibr FM, Al-Muwayhi MA, Marraiki N, Al-Humaid LA (2020) Exogenous melatonin mitigates boron toxicity in wheat. Ecotoxicol Environ Safe 201:110822
Altaf MA, Shahid R, Altaf MA, Ren MX, Tan K, Xiang WQ, Qadir A, Shakoor A, Altaf MM (2019) Effect of NPK, organic manure and their combination on growth, yield and nutrient uptake of chilli (Capsicum annum L.). Hortic Int J 3:217–222
Altaf MM, Diao XP, Shakoor A, Imtiaz M, Altaf MA, Khan LU (2021a) Delineating vanadium (V) ecological distribution, its toxicant potential, and effective remediation strategies from contaminated soils. J Soil Sci Plant Nutr 22:121–139
Altaf MA, Shu H, Hao Y, Zhou Y, Mumtaz MA, Wang Z (2021b) Vanadium toxicity induced changes in growth, antioxidant profiling, and vanadium uptake in pepper (Capsicum annum L.) seedlings. Horticulturae 8:28
Altaf MM, Diao XP, Wang H, Khan LU, Shakoor A, Altaf MA, Farooq TH (2022a) Salicylic acid induces vanadium stress tolerance in rice by regulating the AsA-GSH cycle and glyoxalase system. J Soil Sci Plant Nutr 22:1983–1999
Altaf MA, Shahid R, Ren MX, Naz S, Altaf MM, Khan LU, Lal MK, Tiwari RK, Shakoor A (2022b) Melatonin mitigates cadmium toxicity by promoting root architecture and mineral homeostasis of tomato genotypes. J Soil Sci Plant Nutr 22:1112–1128
Altaf MA, Shahid R, Ren MX, Naz S, Altaf MM, Khan LU, Tiwari RK, Lal MK, Shahid MA, Kumar R, Nawaz MA (2022c) Melatonin improves drought stress tolerance of tomato by modulating plant growth, root architecture, photosynthesis, and antioxidant defense system. Antioxidants 11:309
Altaf MA, Sharma N, Srivastava D, Mandal S, Adavi S, Jena R, Bairwa RK, Gopalakrishnan AV, Kumar A, Dey A, Lal MK (2023) Deciphering the melatonin-mediated response and signalling in the regulation of heavy metal stress in plants. Planta 257:115
Arnao MB, Hernández-Ruiz J (2019) Melatonin: a new plant hormone and/or a plant master regulator. Trends Plant Sci 24:38–48
Benavides MP, Gallego SM, Tomaro ML (2005) Cadmium toxicity in plants. Braz J Plant Physiol 17:21–34
Bradford M (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal Biochem 72:248–254
Cao YY, Qi CD, Li S, Wang Z, Wang X, Wang J, Ren S, Li X, Zhang N, Guo YD (2019) Melatonin alleviates copper toxicity via improving copper sequestration and ROS scavenging in cucumber. Plant Cell Physiol 60:562–574
Carvalho MEA, Piotto FA, Franco MR, Borges KLR, Gaziola SA, Castro PRC, Azevedo RA (2018) Cadmium toxicity degree on tomato development is associated with disbalances in B and Mn status at early stages of plant exposure. Ecotoxicol 27:1293–1302
Chen YE, Mao JJ, Sun LQ, Huang B, Ding CB, Gu Y, Yuan M (2018) Exogenous melatonin enhances salt stress tolerance in maize seedlings by improving antioxidant and photosynthetic capacity. Physiol Plant 164:349–363
DalCorso G, Farinati S, Maistri S, Furini A (2008) How plants cope with cadmium: staking all on metabolism and gene expression. J Integr Plant Biol 50:1268–1280
Devi R, Behera B, Raza MB, Mangal V, Altaf MA, Kumar R, Kumar A, Tiwari RK, Lal MK, Singh B (2021) An insight into microbes mediated heavy metal detoxification in plants: a review. J Soil Sci Plant Nutr 22:914–936
Foy CD, Chaney RT, White MC (1978) The physiology of metal toxicity in plants. Annu Rev Plant Physiol 29:511–566
Gallego SM, Pena LB, Barcia RA (2012) Unravelling cadmium toxicity and tolerance in plants: insight into regulatory mechanisms. Environ Exp Bot 83:33–46
Ghuge SA, Nikalje GC, Kadam US, Suprasanna P, Hong JC (2023) Comprehensive mechanisms of heavy metal toxicity in plants, detoxification, and remediation. J Hazard Mat 450:131039
Gu Q, Chen Z, Yu X, Cui W, Pan J, Zhao G, Xu S, Wang R, Shen W (2017) Melatonin confers plant tolerance against cadmium stress via the decrease of cadmium accumulation and reestablishment of microRNA-mediated redox homeostasis. Plant Sci 261:28–37
Hasan M, Ahammed GJ, Yin L, Shi K, Xia X, Zhou Y, Yu J, Zhou J (2015) Melatonin mitigates cadmium phytotoxicity through modulation of phytochelatins biosynthesis, vacuolar sequestration, and antioxidant potential in Solanum lycopersicum L. Front Plant Sci 6:601
Hasan M, Liu CX, Pan YT, Ahammed GJ, Qi ZY, Zhou J (2018) Melatonin alleviates low-sulfur stress by promoting sulfur homeostasis in tomato plants. Sci Rep 8:1–12
He J, Zhuang X, Zhou J, Sun L, Wan H, Li H, Lyu D (2020) Exogenous melatonin alleviates cadmium uptake and toxicity in apple rootstocks. Tree Physiol 40:746–761
Heath RL, Packer L (1968) Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch Biochem Biophys 125:189–198
Howard LR, Talcott SY, Brenes CH, Villalon B (2000) Changes in phytochemical and antioxidant activity of selected pepper cultivars (Capsicum sp.) as influenced by maturity. J Agric Food Chem 48:1713–1720
Imtiaz M, Rizwan MS, Xiong S, Li H, Ashraf M, Shahzad SM, Shahzad M, Rizwan M, Tu S (2015) Vanadium, recent advancements and research prospects: a review. Environ Int 80:79–88
Jahan MS, Guo S, Baloch AR, Sun J, Shu S, Wang Y, Ahammed GJ, Kabir K, Roy R (2020) Melatonin alleviates nickel phytotoxicity by improving photosynthesis, secondary metabolism and oxidative stress tolerance in tomato seedlings. Ecotoxicol Environ Saf 197:110593
Jahan MS, Shu S, Wang Y, Hasan MM, El-Yazied AA, Alabdallah NM, Hajjar D, Altaf MA, Sun J, Guo S (2021) Melatonin pretreatment confers heat tolerance and repression of heat-induced senescence in tomato through the modulation of ABA- and GA-mediated pathways. Front Plant Sci 12:650955
Kaya C, Okant M, Ugurlar F, Alyemeni MN, Ashraf M, Ahmad P (2019) Melatonin-mediated nitric oxide improves tolerance to cadmium toxicity by reducing oxidative stress in wheat plants. Chemosphere 225:627–638
Kaya C, Ashraf M, Alyemeni MN, Ahmad P (2020) The role of nitrate reductase in brassinosteroid-induced endogenous nitric oxide generation to improve cadmium stress tolerance of pepper plants by upregulating the ascorbate-glutathione cycle. Ecotoxicol Environ Safe 196:110483
Keltjens WG, Van Beusichem ML (1998) Phytochelatins as biomarkers for heavy metal toxicity in maize: single metal effects of copper and cadmium. J Plant Nutr 21:635–648
Korkmaz A, Değer Ö, Szafrańska K, Köklü Ş, Karaca A, Yakupoğlu G, Kocacinar F (2021) Melatonin effects in enhancing chilling stress tolerance of pepper. Sci Hortic 289:110434
Li C, Liang B, Chang C, Wei Z, Zhou S, Ma F (2016) Exogenous melatonin improved potassium content in Malus under diferent stress conditions. J Pineal Res 61:218–229
Li M, Xia Q, Lv S, Tong J, Wang Z, Nie Q, Yang J (2022) Enhanced CO2 capture for photosynthetic lycopene production in engineered Rhodopseudomonas palustris, a purple nonsulfur bacterium. Green Chem 24:7500–7518
Liang C, Zheng G, Li W, Wang Y, Hu B, Wang H, Wu H, Qian Y, Zhu XG, Tan DX (2015) Chen SY (2015) Melatonin delays leaf senescence and enhances salt stress tolerance in rice. J Pineal Res 59(1):91–101
Liang C, Ma C, Zhang Z, Wei Z, Gao T, Zhao Q, Ma F, Li C (2018) Long-term exogenous application of melatonin improves nutrient uptake fluxes in apple plants under moderate drought stress. Environ Exp Bot 155:650–661
Lichtenhaler K, Wellburn AR (1983) Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem Soc Trans 11:591–592
Manafi H, Baninasab B, Gholami M, Talebi M (2021) Effect of melatonin and sodium nitroprusside foliar application on improving high-temperature stress tolerance of ‘Kurdistan’ strawberry. Iran J Hortic Sci Technol 22:383–398
Nawaz MA, Jiao Y, Chen C, Shireen F, Zheng Z, Imtiaz M, Bie Z, Huang Y (2018) Melatonin pretreatment improves vanadium stress tolerance of watermelon seedlings by reducing vanadium concentration in the leaves and regulating melatonin biosynthesis and antioxidant-related gene expression. J Plant Physiol 220:115–127
Ni J, Wang Q, Shah FA, Liu W, Wang D, Huang S, Fu S, Wu L (2018) Exogenous melatonin confers cadmium tolerance by counterbalancing the hydrogen peroxide homeostasis in wheat seedlings. Molecules 23:799. https://doi.org/10.3390/molecules23040799
Park HS, Kazerooni EA, Kang SM, Al-Sadi AM, Lee IJ (2021) Melatonin enhances the tolerance and recovery mechanisms in Brassica juncea (L.) Czern. under saline conditions. Front Plant Sci 12:593717
Piqueras A, Olmos E, Martinez-Solano JR, Hellin E (1999) Cd-induced oxidative burst in tobacco BY2 cells: time course, subcellular location and antioxidant response. Free Radical Res 31:33–38
Qin X, Zhang K, Fan Y, Fang H, Nie Y, Wu XL (2022) The bacterial MtrAB two-component system regulates the cell wall homeostasis responding to environmental alkaline stress. Microbiol Spectr 10:e0231122
Sambrook J, Russell DW (2006) In Vitro mutagenesis using double-stranded DNA templates: selection of mutants with DpnI. Cold Spring Harb Protoc 2006:pdb.prot3813
Saqib M, Shahzad U, Zulfiqar F, Tiwari RK, Lal MK, Naz S, Jahan MS, Awan ZA, El-Sheikh MA, Altaf MA (2023) Exogenous melatonin alleviates cadmium-induced inhibition of growth and photosynthesis through upregulating antioxidant defense system in strawberry. S Afr J Bot 157:10–18
Sarafi E, Tsouvaltzis P, Chatzissavvidis C, Siomos A, Therios I (2017) Melatonin and resveratrol reverse the toxic effect of high boron (B) and modulate biochemical parameters in pepper plants (Capsicum annuum L.). Plant Physiol Biochem 112:173–182
Sharma SS, Dietz KJ (2009) The relationship between metal toxicity and cellular redox imbalance. Trends Plant Sci 14:43–50
Siddiqui MH, Alamri S, Alsubaie QD, Ali HM, Ibrahim AA, Alsadon A (2019) Potential roles of melatonin and sulfur in alleviation of lanthanum toxicity in tomato seedlings. Ecotoxicol Environ Safe 180:656–667
Siddiqui MH, Alamri S, Khan MN, Corpas FJ, Al-Amri AA, Alsubaie QD, Ali HM, Kalaji H, Ahmad P (2020) Melatonin and calcium function synergistically to promote the resilience through ROS metabolism under arsenic-induced stress. J Hazard Mater 398:122882
Sigel A, Sigel H, Sigel RK (eds) (2013) Cadmium: from toxicity to essentiality, vol 11. Springer, New York
Simkin AJ, López-Calcagno PE, Raines CA (2019) Feeding the world: improving photosynthetic efficiency for sustainable crop production. J Exp Bot 70:1119–1140
Singleton V, Rossi JA (1965) Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am J Enol Viticult 16:144–158
Tang Y, Lei F, Zhang E, Li H (2016) Soaking seeds with exogenous melatonin improve photosynthetic characteristics of radish under aluminum stress. 2016 2nd international conference on advances in energy, environment and chemical engineering (AEECE 2016). Atlantis Press, Amsterdam, pp 64–67
Tiwari RK, Lal MK, Naga KC, Kumar R, Chourasia KN, Subhash S, Kumar D, Sharma S (2020) Emerging roles of melatonin in mitigating abiotic and biotic stresses of horticultural crops. Sci Hortic 272:109592
Tiwari RK, Lal MK, Kumar R, Chourasia KN, Naga KC, Kumar D, Das SK, Zinta G (2021) Mechanistic insights on melatonin-mediated drought stress mitigation in plants. Physiol Plant 172:1212–1226
Tousi S, Zoufan P, Ghahfarrokhie AR (2020) Alleviation of cadmium-induced phytotoxicity and growth improvement by exogenous melatonin pretreatment in mallow (Malva parviflora) plants. Ecotoxicol Environ Safe 206:111403
Velikova V, Yordanov I, Edreva A (2000) Oxidative stress and some antioxidant systems in acid rain-treated bean plants: protective role of exogenous polyamines. Plant Sci 151:59–66
Wani PA, Khan MS, Zaidi A (2007) Impact of heavy metal toxicity on plant growth, symbiosis, seed yield and nitrogen and metal uptake in chickpea. Aust J Exp Agric 47:712–720
Wu S, Wang Y, Zhang J, Gong X, Zhang Z, Sun J, Chen X, Wang Y (2021) Exogenous melatonin improves physiological characteristics and promotes growth of strawberry seedlings under cadmium stress. Hortic Plant J 7:13–22
Zhang R, Sun Y, Liu Z, Jin W, Sun Y (2017) Effects of melatonin on seedling growth, mineral nutrition, and nitrogen metabolism in cucumber under nitrate stress. J Pineal Res 62:12403
Zhang G, Zhao Z, Yin X, Zhu Y (2021) Impacts of biochars on bacterial community shifts and biodegradation of antibiotics in an agricultural soil during short-term incubation. Sci Total Environ 771:14475
Zhao YQ, Zhang ZW, Chen YE, Ding CB, Yuan S, Reiter RJ (2021) Melatonin: a potential agent in delaying leaf senescence. Crit Rev Plant Sci 40:1–22
Zhu T, Li L, Duan Q, Liu X, Chen M (2021) Progress in our understanding of plant responses to the stress of heavy metal cadmium. Plant Signal Behav 16:1836884
Acknowledgements
The authors express their gratitude to the Research Square platform and Springer Nature for publishing the pre-print of this manuscript with https://doi.org/10.21203/rs.3.rs-1651489/v1. The authors would like to extend their sincere appreciation to the Researchers Supporting Project Number (RSP2024R182) King Saud University, Riyadh, Saudi Arabia.
Author information
Authors and Affiliations
Contributions
Material preparation was done by MS, data collection by ZA, data analysis by MS, ZA, and US, original draft preparation by MS, MAA, formal analysis by FZ, WAK, reviewing and editing by RKT, MKL, MAE, and MAA and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
No competing interests have been declared by the authors.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Additional information
Communicated by T. Hoshino.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Saqib, M., Shahzad, U., Abid, Z. et al. Melatonin alleviates cadmium phytotoxicity through regulation of growth, photosynthesis, and antioxidant potential in two pepper genotypes. Acta Physiol Plant 46, 49 (2024). https://doi.org/10.1007/s11738-024-03672-6
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11738-024-03672-6