
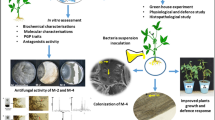
Abstract
Bacillus spp. being rhizospheric bacteria has the potential to provide protection from biotic stresses and also to enhance plant growth. The present study was carried out on bacterial isolate, Bacillus subtilis strain BS87 which has the potential to inhibit several phytopathogens (Bipolaris sorokiniana, Fusarium oxysporum, Fusarium spp., Aspergillus niger, Aspergillus sp., Penicillium sp. and, Rhizoctonia solani) in vitro and is also able to secrete cell-wall-degrading enzymes including protease and chitinase. Isolate BS87, also exhibited several plant growth-promoting traits, such as nutrient solubilization, siderophore, indole-3-acetic acid (IAA), and hydrogen cyanide (HCN) production. Hence, a pot experiment was carried out to evaluate the efficacy of bacterial isolate on plant growth and spot blotch disease in wheat. Results demonstrated that BS87 significantly (p < 0.05) enhanced plant growth and yield in wheat. Chlorophyll content was significantly (p < 0.05) improved when treated with BS87. The spot blotch disease was significantly (p < 0.05) inhibited in wheat by isolate BS87. Lower levels of antioxidant enzymes, including superoxide dismutase, peroxidase, polyphenol oxidase, and phenylalanine ammonia-lyase in the plants infected with spot blotch were observed in bacterial-treated plants. BS87 also significantly reduces malondialdehyde and proline contents in diseased plants. BS87 could successfully colonize and persist in the wheat rhizosphere, providing an advantage to the host plant. Therefore, the study suggested that Bacillus subtilis strain BS87 may be used as an effective bioinoculant, enhancing plant growth and controlling spot blotch disease in wheat caused by Bipolaris sorokiniana.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Wheat (Triticum aestivum L.) is one of the important staple cereal crops of the world, which provides nearly 55% of the carbohydrates and daily proteins for 85% of the population worldwide (FAO 2021) and directly relates to food security and the economy of the globe. This crop is one of India’s primary staple foods; currently, wheat production is around 107.18 million tons, increasing yearly (Singh et al. 2020). Further, due to the exponential growth of the population, there is a need to increase its production through sustainable, eco-friendly strategies. However, some plant diseases in several areas pose a major constraint in wheat production. Predominantly in warmer areas and wheat-rice cropping systems, spot blotch, caused by Bipolaris sorokiniana (Sacc.), is a widespread disease and one of this crop’s most important biotic stresses. In the past 2 decades, spot blotch has lost grain yield from 6.3 to 50% (Juliana et al. 2022). Spot blotch affects the seed quality and thus leads to a loss in its economic value. Chemical control using a triazole group is commonly practiced (Gupta et al. 2018). However, exploring eco-friendly options are required to maintain soil health and compensate for the toxic nature of these chemicals. The biological control of phytopathogens through beneficial plant growth-promoting rhizobacteria (PGPR) is one of the environmentally sustainable options for enhancing soil fertility and crop productivity. The plant–microbial interactions in soil are vital ecological processes (Chandra et al. 2022), and the dynamics of these interactions depend on several factors, e.g., release of microbial metabolites, which may lead to changes in the physiological characteristics of plants. Such plant-associated microbes play an essential role in plant growth and also improve plant nutritional status. PGPRs are soil microbes that colonize the rhizosphere of plants and improve nutrient availability through the solubilization of unavailable phosphorus and zinc. They stimulate plant growth through several mechanisms, including the production of plant hormones (Indole-3-acetic acid (IAA) and several metabolites (Olanrewaju et al. 2017). PGPR can also biocontrol and alleviate/prevent the detrimental effects of phytopathogens. These beneficial microbes also release biocontrol substances including hydrogen cyanide (HCN), siderophores, and cell-wall degrading enzymes (Rosier et al. 2018). Siderophores are ferric iron-chelating compounds and under iron-limited conditions, make them unavailable to the phytopathogens (Sah et al. 2017). Cell-wall-degrading enzymes can suppress diseases by inhibiting pathogens through disintegration of cell walls (Jiao et al. 2021).
Pathogen attack may alter the activity of metabolic enzymes and the contents of non-enzymatic compounds associated with antioxidant metabolism. Thus, the enzymes, including superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD), may be good biochemical markers of oxidative damage. Further, enzymes like polyphenol oxidase (PPO), POD, and phenylalanine ammonia-lyase (PAL) are involved in the phenylpropanoid pathway and play a significant role in plant resistance to stress. Thus, the level and activity of these enzymes could be a sign of resistance or stress (Rosier et al. 2018).
Bacillus spp. is one of the most common rhizospheric dwelling bacteria which have the potential to induce systemic resistance, enhancing the biotic stress tolerance of plants. Bacillus spp. has shown the ability to stimulate various defense reactions in the host plants (Cantoro et al. 2021; Palazzini et al. 2022), specifically under stress conditions including response to pathogen infection. Elevated levels of the activity of antioxidant defense enzymes help the host plants to mitigate the adverse effect of several reactive oxidative species (ROS). Treatment with Bacillus spp. has enhanced the antioxidant defense activities in fungal-infected sugarcane and rice plants, thus alleviating pathogen-induced oxidative damage. The elevated production of different anti-oxidative enzymes and proline content may induce systemic resistance against pathogenic infections in the crops (Rais et al. 2017). Besides the antioxidant defense system, Bacillus treatment may suppress the disease symptoms and enhance plant growth (Miljaković et al. 2020).
Bacillus spp. has significant potential to suppress various diseases and has also been considered a promising biocontrol agent (BCA) (Andrić et al. 2020). Several isolates of Bacillus spp. such as Bacillus subtilis, B. amyloliquefaciens B. licheniformis, B. megaterium, B. velezensis, B. cereus, and B. thuringiensis had successfully colonized the roots of crops and significantly suppressed several pathogens, including Phytophthora capsici, Fusarium oxysporum, Penicillium sp., and Rhizopus stolonifer (Alvindia and Natsuaki 2009; Rao et al. 2017; Miljaković et al. 2020; Mácha et al. 2021). The potential of Bacillus spp. to colonize the plant roots and enhance their growth has also been well documented along with their biocontrol properties (Sukkasem et al. 2018; Hashem et al. 2019). Hence, we hypothesize that inoculation of Bacillus subtilis strain BS87 in wheat may develop induced systemic resistance against spot blotch disease and improve plant growth, as well as it can persist after in the wheat rhizosphere.
Materials and methods
Biocontrol agents and their characterization
Bacillus subtilis strain BS87 has been used as a biocontrol agent in the present study. Based on the earlier study, the culture was isolated from the rhizospheric soil of fields where sugarcane has been grown for > 50 years (ICAR-Indian Institute of Sugarcane Research, Lucknow, India, 26° 56′ N, 80° 52′ E and 111 m above sea level), and was further characterized and identified as B. subtilis (Chandra et al. 2018). Soils of these fields were largely sandy loam (13.3% clay, 24.5% silt, and 62.2% sand) of Indo-Gangetic alluvial origin and are classified as non-calcareous mixed with hyperthermic udic ustochrept. The 16sRNA was submitted to NCBI Genbank, and the Accession No. (KY604948.1) was obtained. Keeping the importance of the isolate in terms of its plant growth-promoting potential in view, the culture was deposited to the repository (National Agriculturally Important Microbial Culture Collection, ICAR-National Bureau of Agriculturally Important Microorganisms), and Accession No. (NAIMCC-B-02187) was obtained. A whole genome sequence of Bacillus subtilis strain BS87 has been reported, which showed that this isolate could be a preferable candidate as a biofertilizer (Chandra et al. 2021a). The genome sequence database of isolate BS87 has been deposited in the NCBI SRA module and is accessible through Accession No. SRR11870891 (PRJNA634628) (Chandra et al. 2021a). The BS87 culture was maintained by regular subculturing on nutrient agar medium (NAM) (peptone 5.0; yeast extract 2.0; sodium chloride 5.0; agar 15.0) at 30 ºC for 24 h and stored at 4 ºC. Growth and morphological attributes of the biocontrol agent were recorded. The BS87 culture was characterized by various biochemical tests like citrate and D-glucose utilization, urease, oxidase, catalase, and Gram staining (Chandra and Chandra 2017).
In vitro biocontrol potential
In vitro biocontrol potential was evaluated against fungal pathogens, namely Bipolaris sorokiniana (BP), Fusarium oxysporum (FO), Fusarium spp. (FS), Aspergillus niger (AN), Aspergillus sp. (AS), Penicillium sp. (PS), and Rhizoctonia solani (RS). In vitro screening for antagonistic BS87 culture activity was performed by dual culture technique on potato dextrose agar (PDA) (potatoes 200.0; dextrose 20.0; agar 15.0) plates. PDA medium was prepared and poured (20 ml) into sterile Petri dishes. A 5-mm agar disc of actively growing pathogens was placed in the center of each plate, and BS87 culture was streaked 3 cm away from the agar disc toward the edge of the Petri dish. In the control plate, no bacterial isolate was inoculated. Plates were covered with parafilm and incubated at 27 ± 2 °C for 5 days until the fungal mycelia reached the edge in the control plates. Mycelial growth inhibition indicates antagonistic activity. The percentage (%) of radial mycelial growth inhibition was calculated.
In vitro PGPR properties
The BS87 was screened for phosphate solubilization on Pikovskaya’s agar medium. It was also checked for their Zn solubilizing ability based on halo formation on solid basal medium supplemented with ZnO (Kamran et al. 2017). The results were represented as the diameter of the solubilized area (mm) and then the solubilization index for zinc and phosphate was calculated using the following formula. Solubilization index (SI) = (colony diameter + halo zone diameter)/ colony diameter.
The indole-3-acetic acid (IAA) content was estimated using Salkowski reagent (Gang et al. 2019). Siderophore production was assayed using tertiary complex Chrome Azurol S (CAS)/ Fe hexadecyl trimethyl ammonium bromide. The development of a yellow–orange halo around the growth was considered positive for siderophore production (Gang et al. 2019). The diameter of the resultant zone was measured and the siderophore production index was calculated with the following formula.
The BS87 was also tested for the production of ammonia in peptone water as reported by Chandra and Chandra 2016. The development of faint yellow to dark brown color was considered a positive test for ammonia production. Hydrogen cyanide (HCN) production was tested on nutrient agar supplemented with glycine by Kamran et al. (2017). Briefly, qualitative phosphorus-solubilizing potential was also analyzed by measuring soluble P concentration colorimetrically using the molybdenum blue method. Titrable acidity (g tartaric acid L−1) and change in pH of the medium were also measured (Panda et al. 2016).
Estimation of cell-wall-degrading enzymes
Protease production by BS87 was determined using 1% skimmed milk agar. The protease activity was indicated by forming clear zones around the inoculated site on the agar plates (Chandra and Vij 2018). Similarly, the chitinase production was assayed on chitin agar plates (using powdered chitin as the sole carbon source). A clear halo zone around the colonies developed after 48–72 h of incubation demonstrated chitinase production.
In vitro germination test
For the in vitro germination test, the seeds of wheat varieties Raj 3765 and DBW 90 were surface sterilized with 0.1% mercuric chloride (HgCl2), washed with distilled water properly, and then treated with BS87 culture (109 cells/ml) for 30 min and then dried aseptically. Seeds treated with an uninoculated medium were considered as a control. The dried seeds were incubated on sterilized wet blotting paper in pre-sterilized petri dishes. The germination percentage and growth parameters were recorded at 2–7 days. Based on the growth parameters, the vigor index was calculated.
Wheat cultivars
Two wheat (Triticum aestivum L.) varieties, Raj 3765 and DBW 90, have been used in the study. The seeds were obtained from the ICAR-Indian Institute of Wheat and Barley Research, Karnal, India. These two varieties were selected as they are suitable for different pedoclimatic areas in India and are late-sown varieties in both areas. The wheat variety DBW 90 is more suitable for the irrigated conditions of the North Western Plains Zone, while Raj 3765 is highly suitable for the North West and North Eastern Plain Zone.
Preparation of bacterial inoculum
For the preparation of bacterial inoculum, BS87 culture inoculum was grown in nutrient broth and incubated at 28 °C for 48 h by shaking at 150 rpm. The bacterial cells were centrifuged for 10 min at 12,000 rpm. The pellet formed was resuspended in 10% sterilized glycerol with a 108 CFU/ml concentration. The bacterial inoculum’s colony-forming unit (CFU) was calculated using the serial dilution method. The CFU count of the bacterial inoculum was 4.0 × 108 mL−1.
Seed biopriming
The prepared bacterial inoculum was mixed in 1 kg sterilized talc powder for seed biopriming. The pH was adjusted to 7 by adding calcium carbonate. The talc-based bacterial inoculum was stored at 4 °C after maintaining 35% moisture content. The talc-based bacterial inoculum was serially diluted and plated on NAM at 28 °C for 24 h to test viable bacterial colonies. Five-gram seeds of Raj 3765 and DBW 90 genotypes were surface sterilized by 95% ethanol following 0.2% HgCl2 solution for 3 min. Then seeds were thoroughly washed with sterile distilled water and air dried aseptically. Biopriming of the surface-sterilized wheat seeds was carried out by coating with 0.2% (w/v) sterilized carboxymethyl cellulose (CMC) and talc-based bacterial inoculum prepared earlier. The seeds were left for 6 h and dried aseptically. The seeds treated with only CMC suspension in distilled water served as control (Chandra et al. 2021b).
Experimental set-up
Bioprimed five seeds of both the wheat varieties were sown in each pot. The capacity of the pots was 12 kg, and was filled with sterilized soil collected from the fields of ICAR-Indian Institute of Wheat and Barley Research, Karnal, Haryana, India, situated in the Indo-Gangetic plains (29o43ʹ N, 76o58ʹ E, 245 m above mean sea level). The physiochemical properties of soils collected were as follows—texture: sandy loam; sand: 61.2%; silt: 25.1%; clay: 10.2%; pH (1:2): 7.2; electrical conductivity (1:2 soil: water) (dS−1): 0.48 dS m−1; soil organic carbon: 0.41%; available nitrogen: 198.8 kg ha−1; available phosphorus: 16.2 kg ha−1; available potassium: 242 kg ha−1. The experiments were laid out during October–March with 60–75% relative humidity under an 11/13 h light/dark photoperiod. Moisture content (at field capacity, 60%) in the pots was maintained by sprinkling sterilized water on every alternate day. The experiment was designed in a completely randomized design (CRD) with three replications and eight treatments, as described in Table 1.
Pathogenic inoculum preparation
Conidia of BP were used as inoculum to initiate spot blotch disease in wheat plants. The conidia of BP were produced by inoculating the pathogen in potato dextrose broth. After 7 days of incubation at 25 °C at 120 RPM, the grown culture was filtered through sterile gauze to obtain a conidial suspension. The conidia concentration was adjusted to 1 × 105 conidia ml−1 with sterile distilled water plus Tween 80 (0.05%) using a Neubauer chamber for inoculations in plants. The conidial suspension prepared was artificially inoculated during the stem extension stage of wheat. The growing stems of wheat were infected by injecting the conidial suspension with help of a sterile syringe. After 2 days of inoculation, the disease symptoms were visible.
Biocontrol foliar spray
After 24 h of pathogen inoculation, biocontrol foliar spray consisting BS87 culture inoculum was applied on plants. Briefly, foliar spray was prepared from the bacterial inoculum described earlier. The bacterial inoculum was diluted by adding sterile water with 0.05% v/v Tween 80 and maintained upto 1.0 × 108 CFU mL−1. The diluted bacterial inoculum was applied as a foliar application at the rate of 5 mL per plant during evening hours (16:00 h) to avoid the direct effect of sunlight.
Plant growth and nutrient status
Chlorophyll content was recorded at the seedling stage using SPAD 502. Plant height (cm) and spike length (cm) were recorded at the physiological maturity of the plants. After harvesting, the total grain weight was recorded by the electronic weighing balance. The number of grains was measured manually, while 1000 grain weight was recorded by the seed counter (CONTADOR). The Kjeldahl method was used to estimate the nitrogen (Michałowski et al. 2013), while phosphorus was determined colorimetrically using the ascorbic acid reductant method (Jackson 1967). A flame photometer was used to estimate the potash. Fe, Zn, Mn, and Cu in grains and straw were digested by di-acid (HNO3–HClO4) and estimated by atomic absorption spectrophotometer (Analytika Jena, ZEEnit 700p; Germany). The uptake of nutrients was computed by multiplying the concentrations with the dry weight of the respective plant parts.
Antioxidant defense enzymes
The antioxidant defense-related enzymes viz. superoxide dismutase (SOD), peroxidase (PO), polyphenol oxidase (PPO), and phenylalanine ammonia-lyase (PAL) were estimated in all the treatments after the emergence of spot blotch in infected plants. Two grams of the sample were crushed and mixed with 4 mL of phosphate buffer (0.1 M, pH 7) in a pre-chilled mortar and pestle. This mixture was centrifuged at 10,000 rpm, 4 °C for 15 min, and the supernatant was used as crude enzyme extract, according to Chandra et al. (2018). For PO, PPO, and PAL, the absorbance was recorded using a spectrophotometer at 280 nm, 450 nm, and 460 nm, respectively. The enzyme activity was expressed as a change in absorbance min/g (FW). The activity of superoxide dismutase was measured at 560 nm, and the activity was presented as the enzyme unit, defined as the amount of enzyme needed to inhibit a 50% reduction of NBT. This was presented in unit min−1 mg−1. Total proline content and estimation of the level of lipid peroxidation in terms of malondialdehyde were carried out as described earlier (Chandra et al. 2018).
Microbial enzymes and colonization analysis
The plate count technique estimated bacterial colonization in the rhizosphere and expressed it as the number of colony-forming units (CFU) per g of dry soil (Chandra et al. 2020). The microbial enzymes estimated in the soil were dehydrogenase (Casida et al. 1964), and alkaline and acidic phosphatase (Tabatabai and Bremner 1969). Alkaline and acidic phosphatase activity was expressed as mg p-nitrophenol released g−1 soil h−1. Dehydrogenase activity was expressed as mg of triphenyl formazan (TPF) g−1 soil d−1.
Statistical analysis
Samples in three replications consisted of three independent plants to generate all experimental data. To test the significance of treatments, data were analyzed by one-way analysis of variance (ANOVA), and treatment means were compared using DUNCAN’s multiple range test at a 5% level (p < 0.05). Statistical analysis of experimental data was performed online on Indian NARS Statistical Computing Portal (http://stat.iasri.res.in/sscnarsportal) using General Linear Model (GLM) procedure in SAS (SAS Institute Inc.).
Results
Biochemical characterization and plant growth-promoting properties
The biochemical characterization revealed that BS87 culture is an oxidase negative, can hydrolyze starch, and utilizes citrate and D-glucose as carbon sources (Table 2). Optical density showed that culture could grow up to 45 ºC, while the optimal growth temperature was 30 ºC. The isolate could tolerate salt (NaCl) concentrations up to 7%, while the optimum growth was observed at a 4% NaCl level. Similarly, pH tolerance was up to pH 11 (Table 2). BS87 produced chitinase, protease, and cellulase enzymes, which was confirmed by the halo-zone around the culture growth on the medium supplemented by chitin, skim milk, and carboxymethyl cellulose (Table 2). BS87 produced indole-3-acetic acid, which was confirmed by the conversion to the pink-colored medium supplemented with tryptophan. The appearance of an orange-halo zone (22 mm) on the CAS-agar plate confirmed the siderophore production. BS87 isolate was also found positive for ammonia and HCN production. The formation of a clear zone around the bacterial colonies on media supplemented with an insoluble form of phosphorus (tricalcium phosphate) and zinc (zinc oxide) indicated phosphorus- and zinc-solubilizing activity (Table 2). BS87 solubilized 61.7 mg L−1 phosphorus in 5 days, increasing to 81.3 mg L−1 in 15 days. In support of phosphorus- and zinc-solubilizing activity, organic acid secretion by BS87 was also observed, which decreased pH to 4.1 (Table 3).
Biocontrol potential
The biocontrol potential of BS87 culture was significantly highest against FS (51.05%) and FO (47.96%) (Table 4) (Fig. 1a, b) following AN (43.65%) and BP (39.77%). However, it was significantly the least in the case of AS (36.02%), PS (31.85%), and RS (29.93%) (Table 4). After wheat inoculation of BP alone, lesions were successfully formed, as shown in Fig. 2a and after the application of BS87, the spot blotch disease in wheat was impeded. The number of lesions/cm2 formed in each pot in the different treatments was noted and presented in the heat map (Fig. 2b). In BS87-treated pots, lesions due to the infection of BP were significantly lower in comparison to the non-treated plants. Non-treated varieties Raj 3765 (T5) and DBW 90 (T6) showed 8.33 and 8 lesions/cm2, which were reduced to 4.67 and 5.33 lesions/cm2, respectively, when plants were treated with BS87 (Fig. 2c, d) (supplementary Table 1).

Growth inhibition demonstrated by biocontrol agent, Bacillus subtilis strain BS87: a. Bipolaris sorokiniana, b. Fusarium oxysporum

a. Arrow showing lesions of spot blotch on the leaves. b. Heat map expressing number of lesions/cm2 in each Bipolaris sorokiniana inoculates pots of different treatments ((i) pot 1 (ii) pot 2 (iii) pot 3). c. Graph representing number of lesions/cm2 in different treatments. d. Leaves showing symptoms (necrosis and chlorosis) due to spot blotch in control and BS87 applied treatments (T5: wheat variety DBW 90 infected with BP; T6: wheat variety Raj 3765 infected with BP; T7: wheat variety DBW 90 infected with BP and treatment of BS87; T8: wheat variety Raj 3765 infected with BP and treatment of BS87.) Means with different capital letters within same column are significantly different (p < 0.05) using DUNCAN’s multiple range test
Plant growth and nutrient status
In vitro experiment demonstrated significant improvement in seedling vigor of wheat when treated with BS87. The germination percentage in both varieties significantly increased from 83–86 to 91–92% after treatment. The vigor index of the non-treated seed variety Raj 3765 was 317.89, while after treatment with BS87, it significantly increased to 604.4. However, the vigor index for variety DBW 90 was 260.58, which increased to 504 after treatment with BS87 (Fig. 3, supplementary Table 2). Similarly, in pot experiments, the growth and yield of the wheat plants were enhanced by treating the seeds with BS87 culture compared to the non-treated control. Chlorophyll content was also significantly higher in wheat varieties treated with BS87. Significantly higher grain yield was observed in DBW 90 (7.28 g/pot), which was followed by wheat variety Raj 3765 (5.36 g/pot). In BP-infected plants, the yield was similar to non-infected crops when treated with BS87 culture (Table 5). A similar pattern of results was found for straw yield also. Inoculation of BS87 significantly boosted nutrient uptake in wheat varieties, further increasing the concentration of both macronutrient and micronutrients (Table 6). The nitrogen content in both straw and grain was significantly higher than in control. A similar pattern was observed for phosphorus and potassium content. The micronutrient content in grain and straw was also found to be significantly higher in the BS87-treated plants (Table 6).

Results of in vitro experiment demonstrating the effect of Bacillus subtilis strain BS87 on germination and seedling growth of two wheat varieties (C1: control of wheat variety DBW90 (without bacterial treatment); C2: wheat variety Raj 3765 (without bacterial treatment); B1: seed of wheat variety DBW 90 treated with BS87; B2: wheat variety Raj 3765 treated with BS87
Antioxidant enzymes
An increased level of superoxide dismutase in T6 (7.87 min−1 mg−1) followed closely by T5 (6.8 min−1 mg−1) was observed in BP-infected plants. Bacterial treatment significantly reduced the SOD activity in infected plants (Fig. 4i) (Supplementary Table 3). Following the pattern of SOD, BP-infected plants (T6 and T5) showed a significant increase in MDA (15.8 µM−1 g) and proline content (30.13 µg−1 g). Bacterial treatment significantly reduced these contents, while MDA and proline content were much less in control plants (Fig. 4ii, iii) (supplementary Table 3). PO was maximum in the plants infected with BP and treated by BS87 (T7 and T8) (supplementary Table 3). Non-inoculated control wheat variety Raj 3765 (T2) and wheat variety DBW 90 treated with BS87 (T3) did not show the presence of PO (Fig. 4iv). Similarly, the plants infected with BP and treated with BS87 (T7 and T8) showed significantly higher PAL levels, while control and BS87-treated varieties did not exhibit PAL in a detectable range (Fig. 4v). PPO was also highest in T7 and T8, while it was lower in BS87-treated plants (T3 and T4). T1 did not show any presence of PPO (Fig. 4vi) (supplementary Table 3).

(i) Superoxide dismutase (SOD) activity, (ii) malondialdehyde (MDA) content, (iii) proline content, (iv) peroxidase (PO), (v) polyphenol oxidase (PPO), and (vi) phenylalanine ammonia-lyase (PAL) activity in the leaves of Bacillus subtilis strain BS87-treated and non-treated wheat varieties. T1: non-inoculated control of wheat variety DBW 90; T2: non-inoculated control of wheat variety Raj 3765; T3: wheat variety DBW 90 treated with BS87; T4: wheat variety Raj 3765 treated with BS87; T5: wheat variety DBW 90 infected with BP; T6: wheat variety Raj 3765 infected with BP; T7: wheat variety DBW 90 infected with BP and treatment of BS87; T8: wheat variety Raj 3765 infected with BP and treatment of BS87. Means with different capital letters within same column are significantly different (p < 0.05) using DUNCAN’s multiple range test
Bacterial colonization and soil microbial enzymes
The bacterial colonization was significantly high in T7 (1463.67 × 103 CFU) and T4 (1254 × 103 CFU) (Table 7). BS87 treatment significantly enhanced the soil microbial enzymatic activity. Dehydrogenase activity was significantly high in T4 (799 μg TPF g − 1 soil d-1) followed by T8 (664.67 μg TPF g−1soil d−1) and T7 (599.67 μg TPF g−1 soil d−1). Alkaline phosphatase activity significantly increased in soils inoculated with BS87. However, acidic phosphatase activity was high in T7 (172.33 μgp NPP g−1 soil h−1) in T8 and T3 (Table 7).
Discussion
The microbial community that resides in the rhizosphere has been reported to promote plant growth and suppress the disease through several biochemical and molecular mechanisms (Chandra et al. 2021c). In the present study, plant growth-promoting properties of BS87 exerted a positive influence on wheat growth and productivity. Improvement in seed germination and seedling vigor in vitro is likely attributable to IAA produced by BS87. IAA promotes plant growth and development of roots contributing to their rapid establishment, facilitating their elongation, and the proliferation of lateral and adventitious roots (Olanrewaju et al. 2017). BS87 was able to enhance the growth and yield of wheat and also to increase the ability of seedlings to establish in soil, in turn helping them to extract water and nutrients more efficiently. PGPR has also been reported to modulate plant’s root architecture, enhancing nutrient uptake and increasing crop yield (Vacheron et al. 2013; Ambreetha et al. 2018; Grover et al. 2021). Present results are in consonance with earlier studies wherein an increase in length and weight of the root and stem of Dioscorea rotundata L was reported when they were treated with IAA-producing B. subtilis (Swain et al. 2007).
The ability to synthesize IAA is widely associated with PGPR strains, which can be used as bioinoculants for plant crops to enhance root and shoot biomass. IAA production may result in overall root development, including root growth, and increase root surface area and metabolism (Gang et al. 2019). For example, inoculating rapeseed seeds with Pseudomonas putida increased primary root elongation. Therefore, the enhanced plant growth in the presence of PGPR inoculants is likely primarily attributed to the secretion of auxin (Souza et al. 2015). A few other significant features of the BS87 culture are the ability to solubilize phosphorus and produce siderophore, ammonia, and HCN. The increased biomass and overall growth in the case of seeds treated with BS87 culture may be attributed to the collective effect of phosphate solubilization and the production of plant growth regulators. Bacillus species have been tested on various crops and reported to increase growth, biomass, and yield (Aloo et al. 2019). Phosphorus solubilizing potential is the ability of microbes to produce metabolites such as organic acids. Their hydroxyl and carboxyl groups chelate the phosphate ion and convert them to soluble forms (Alori et al. 2017; Rai et al. 2021). The BS87 strain has effectively converted the insoluble inorganic phosphate into a soluble form in the present study. The results align with the earlier reports in which solubilization of phosphorus in a liquid medium has been demonstrated by PGPR (Gupta et al. 2012; Panda et al. 2016).
Similarly, a decrease in the pH of the culture medium was also observed in the present study, possibly due to the culture’s secretion of organic acids. Organic acids produced by microbes are reported to be responsible for their P-solubilization efficiency (Panda et al. 2016). BS87 not only improved the growth of seedlings but also felicitated the nutrient uptake due to a better nutrient solubilization ability which also led to a more root-shoot ratio. A combination of PGPR strains (Azotobacter vinelandii, Pantoea agglomerans, Pseudomonas putida) showed enhancement of nutrient uptake in Onobrychis sativa (Delshadi et al. 2017). Bacillus and Pseudomonas possessing zinc-solubilizing capacity were also found to enhance the uptake of nutrients in maize (Goteti et al. 2013). Similarly, an increase in the Fe and Mg content in the Pseudomonas- and Bacillus-treated strawberries was reported (Esitken et al. 2010). Improved chlorophyll content was also observed which can be attributed to the better growth of BS87-treated plants.
Beneficial microbes in the rhizosphere produce various hydrolytic enzymes responsible for the degradation of various cell components of fungal pathogens. These enzymes usually break glycosidic linkages in the polysaccharides of the cell wall of pathogens leading to cell death, hence playing an important role in plant disease management (Singh and Gaur 2017). BS87 culture successfully control the spread of spot blotch disease in wheat, possibly as a consequence of hydrolytic enzymes production that would impair the adhesion of the conidial spores on the surface of leaves or the formation of germ tube, its penetration and growth. (Gupta et al. 2018). Moreover, BS87 is fast growing (Table 2), can swiftly colonize the phyllosphere of the wheat, and is tolerant to variable circumstances (temperature, pH, salt). It can easily control the adhesion of the conidial spores of BP on the phyllosphere, thus reducing the infection. It has been reported that the B. subtilis group can produce various metabolites that possess various biological control activities such as antibiotics, biosurfactants, lytic enzymes, and siderophores (Singh and Gaur 2017). Similarly, inhibition of BP in Triticum turgidum L. subsp. durum by native rhizospheric B. subtilis strain was also reported (Villa-Rodríguez et al. 2019). The inhibition of pathogenic fungal growth by BS87 may also be attributed to metabolites such as siderophore, ammonia, and hydrogen cyanide secreted by it. Microbial strains such as B. cereus, B. subtilis, and B. thuringiensis have been reported to inhibit several phytopathogens through hydrolytic enzyme secretion (Miljaković et al. 2020). BS87 also secretes organic acid that leads to the lowering of pH of the surrounding environment, which is also found responsible for the growth inhibition of phytopathogens (Table 2).
In the present study, BS87 significantly enhanced the activity of antioxidant enzymes, possibly contributing to spot blotch control. As also previously observed, Bacillus sp was found to control the rice blast caused by Pyricularia oryzae (Rais et al. 2017). Streptomyces spp. improves the activity of antioxidant enzymes by secreting a siderophore and decreases the incidence and progression of Sclerotium rolfsii in chickpea (Singh and Gaur 2017).
The ability to colonize the root system is crucial for rhizobacteria to be an effective plant growth promoter (Weisskopf et al. 2011). The biochemical characteristics like starch hydrolytic property and utilization of citrate as the carbon source of BS87 culture (Table 2) showed that they could sustain around plant roots and survive competitive root colonization. The total CFU counts obtained from the analysis, carried out from the rhizosphere of the treated plants, were high compared to the control, demonstrating that BS87 culture successfully colonized the rhizosphere of wheat plants. However, the colonization differs with the wheat variety as the population of the same bacteria varies in both of the wheat varieties used in the present study. These results demonstrated specific interactions between wheat varieties and the BS87 culture. Microbial enzymes reflect their microbiological activity. Indeed, the microbial enzymatic activity, namely dehydrogenase, and phosphatases, was higher in the rhizospheric soil of the BS87 culture-inoculated plants. The results of microbial enzymes and the total CFU count were found to be well correlated, which indicated that the BS87 strain has persisted and well colonized the rhizosphere of both the wheat varieties.
Conclusion
The present results suggested that the biocontrol agent B. subtilis strain BS87 possesses growth-promoting attributes along with the potential to control several fungal pathogens. Inoculation of B. subtilis strain BS87 led to improved wheat plant growth, grain yield, and nutrient status. It also controlled the spot blotch disease in wheat varieties of different regions and could colonize and persist in their rhizosphere. Further investigation aimed to test the efficiency of this strain in the field under different climatic conditions would undoubtedly improve its applicability, along with its bulk production and exploration/ knowledge of associated metabolites.
Data availability
The datasets generated during and/or analysis the current study are available from the corresponding author on reasonable request.
References
Aloo BN, Makumba BA, Mbega ER (2019) The potential of Bacilli rhizobacteria for sustainable crop production and environmental sustainability. Microbiol Res 219:26–39. https://doi.org/10.1016/j.micres.2018.10.011
Alori ET, Glick BR, Babalola OO (2017) Microbial phosphorus solubilization and its potential for use in sustainable agriculture. Front Microbiol 8:971. https://doi.org/10.3389/fmicb.2017.00971
Alvindia D, Natsuaki K (2009) Biocontrol activities of Bacillus amyloliquefaciens DGA14 isolated from banana fruit surface against banana crown rot-causing pathogens. Crop Prot 28:236–242. https://doi.org/10.1016/j.cropro.2008.10.011
Ambreetha S, Chinnadurai C, Marimuthu P, Balachandar D (2018) Plant-associated Bacillus modulates the expression of auxin-responsive genes of rice and modifies the root architecture. Rhizosphere 5:57–66. https://doi.org/10.1016/j.rhisph.2017.12.001
Andrić S, Meyer T, Ongena M (2020) Bacillus responses to plant-associated fungal and bacterial communities. Front Microbiol. https://doi.org/10.3389/fmicb.2020.01350
Cantoro R, Palazzini JM, Yerkovich N et al (2021) Bacillus velezensis RC 218 as a biocontrol agent against Fusarium graminearum: effect on penetration, growth and TRI5 expression in wheat spikes. Biocontrol 66:259–270. https://doi.org/10.1007/s10526-020-10062-7
Casida LE, Klein DA, Santoro T (1964) Soil dehydrogenase activity. Soil Sci 98:371–376
Chandra P, Chandra A (2016) Isolation and characterization of plant growth promoting bacteria from wheat-sugarcane cropping system. J Cereal Res 8:62–65. https://doi.org/10.13140/RG.2.2.10655.00162
Chandra P, Chandra A (2017) Elucidation of rRNA secondary structure and phylogenetic analysis of Salmonella enterica based on 16S RNA. Int J Curr Microbiol Appl Sci 6:4056–4063
Chandra P, Vij S (2018) Molecular characterization and identification of bioactive peptides producing Lactobacillus sps. Based on 16S rRNA gene sequencing. Food Biotechnol 32:1–14. https://doi.org/10.1080/08905436.2017.1413657
Chandra P, Tripathi P, Chandra A (2018) Isolation and molecular characterization of plant growth-promoting Bacillus spp. and their impact on sugarcane (Saccharum spp. hybrids) growth and tolerance towards drought stress. Acta Physiol Plant 40:199. https://doi.org/10.1007/s11738-018-2770-0
Chandra P, Dhuli P, Verma P et al (2020) Culturable microbial diversity in the rhizosphere of different biotypes under variable salinity. Trop Ecol 61:291–300. https://doi.org/10.1007/s42965-020-00089-3
Chandra A, Chandra P, Tripathi P (2021a) Whole genome sequence insight of two plant growth-promoting bacteria (B. subtilis BS87 and B. megaterium BM89) isolated and characterized from sugarcane rhizosphere depicting better crop yield potentiality. Microbiol Res 247:126733. https://doi.org/10.1016/j.micres.2021.126733
Chandra P, Khobra R, Sundha P et al (2021b) Plant growth promoting Bacillus-based bio formulations improve wheat rhizosphere biological activity, nutrient uptake and growth of the plant. Acta Physiol Plant 43:139. https://doi.org/10.1007/s11738-021-03310-5
Chandra P, Wunnava A, Verma P et al (2021c) Strategies to mitigate the adverse effect of drought stress on crop plants—influences of soil bacteria: A review. Pedosphere 31:496–509. https://doi.org/10.1016/S1002-0160(20)60092-3
Chandra P, Singh A, Prajapat K et al (2022) Native arbuscular mycorrhizal fungi improve growth, biomass yield, and phosphorus nutrition of sorghum in saline and sodic soils of the semi–arid region. Environ Exp Bot 201:104982. https://doi.org/10.1016/j.envexpbot.2022.104982
de Souza R, Ambrosini A, Passaglia LMP (2015) Plant growth-promoting bacteria as inoculants in agricultural soils. Genet Mol Biol 38:401–419. https://doi.org/10.1590/S1415-475738420150053
Delshadi S, Ebrahimi M, Shirmohammadi E (2017) Influence of plant-growth-promoting bacteria on germination, growth and nutrients’ uptake of Onobrychis sativa L. under drought stress. J Plant Interact 12:200–208. https://doi.org/10.1080/17429145.2017.1316527
Esitken A, Yildiz HE, Ercisli S et al (2010) Effects of plant growth promoting bacteria (PGPB) on yield, growth and nutrient contents of organically grown strawberry. Sci Hortic (amsterdam) 124:62–66. https://doi.org/10.1016/j.scienta.2009.12.012
FAO (2021) Cereal Supply and Demand Brief, Food and Agriculture. Organization of the United Nations, Washington DC
Gang S, Sharma S, Saraf M et al (2019) Analysis of indole-3-acetic acid (IAA) production in Klebsiellaby LC-MS/MS and the Salkowski Method. Bio-Protoc 9:e3230. https://doi.org/10.21769/BioProtoc.3230
Goteti PK, Emmanuel LDA, Desai S, Shaik MHA (2013) Prospective zinc solubilising bacteria for enhanced nutrient uptake and growth promotion in maize (Zea mays L.). Int J Microbiol. https://doi.org/10.1155/2013/869697
Grover M, Bodhankar S, Sharma A et al (2021) PGPR mediated alterations in root traits: way toward sustainable crop production. Front Sustain Food Syst. https://doi.org/10.3389/fsufs.2020.618230
Gupta M, Kiran S, Gulati A et al (2012) Isolation and identification of phosphate solubilizing bacteria able to enhance the growth and aloin—a biosynthesis of Aloe barbadensis Miller. Microbiol Res 167:358–363. https://doi.org/10.1016/j.micres.2012.02.004
Gupta PK, Chand R, Vasistha NK et al (2018) Spot blotch disease of wheat: the current status of research on genetics and breeding. Plant Pathol 67:508–531. https://doi.org/10.1111/ppa.12781
Hashem A, Tabassum B, Fathi AbdAllah E (2019) Bacillus subtilis: a plant-growth promoting rhizobacterium that also impacts biotic stress. Saudi J Biol Sci 26:1291–1297. https://doi.org/10.1016/j.sjbs.2019.05.004
Jackson ML (1967) Soil chemical analysis. Printice Hall of India Pvt. Ltd., New Delhi
Jiao X, Takishita Y, Zhou G, Smith DL (2021) Plant associated rhizobacteria for biocontrol and plant growth enhancement. Front Plant Sci 12:634796. https://doi.org/10.3389/fpls.2021.634796
Juliana P, He X, Poland J et al (2022) Genome-wide association mapping indicates quantitative genetic control of spot blotch resistance in bread wheat and the favorable effects of some spot blotch loci on grain yield. Front Plant Sci. https://doi.org/10.3389/fpls.2022.835095
Kamran S, Shahid I, Baig DN et al (2017) Contribution of zinc solubilizing bacteria in growth promotion and zinc content of wheat. Front Microbiol. https://doi.org/10.3389/fmicb.2017.02593
Mácha H, Marešová H, Juříková T et al (2021) Killing effect of Bacillus Velezensis FZB42 on a Xanthomonas campestris pv. Campestris (Xcc) strain newly isolated from Cabbage Brassica Oleracea Convar. Capitata (L.): a metabolomic study. Microorganisms 9:10. https://doi.org/10.3390/microorganisms9071410
Michałowski T, Asuero AG, Wybraniec S (2013) The titration in the Kjeldahl method of nitrogen determination: base or acid as titrant? J Chem Educ 90:191–197. https://doi.org/10.1021/ed200863p
Miljaković D, Marinković J, Balešević-Tubić S (2020) The significance of Bacillus spp. in disease suppression and growth promotion of field and vegetable crops. Microorganisms. https://doi.org/10.3390/microorganisms8071037
Olanrewaju OS, Glick BR, Babalola OO (2017) Mechanisms of action of plant growth promoting bacteria. World J Microbiol Biotechnol 33:197. https://doi.org/10.1007/s11274-017-2364-9
Palazzini J, Reynoso A, Yerkovich N et al (2022) Combination of Bacillus velezensis RC218 and chitosan to control fusarium head blight on bread and durum wheat under greenhouse and field conditions. Toxins (basel). https://doi.org/10.3390/toxins14070499
Panda B, Rahman H, Panda J (2016) Phosphate solubilizing bacteria from the acidic soils of Eastern Himalayan region and their antagonistic effect on fungal pathogens. Rhizosphere 2:62–71. https://doi.org/10.1016/j.rhisph.2016.08.001
Rai AK, Dinkar A, Basak N et al (2021) Phosphorus nutrition of oats genotypes in acidic soils: exploiting responsive plant-microbe partnership. Appl Soil Ecol 167:104094. https://doi.org/10.1016/j.apsoil.2021.104094
Rais A, Jabeen Z, Shair F et al (2017) Bacillus spp., a bio-control agent enhances the activity of antioxidant defense enzymes in rice against Pyricularia oryzae. PLoS ONE 12:e0187412
Rao M, Kamalnath M, Umamaheswari R et al (2017) Bacillus subtilis IIHR BS-2 enriched vermicompost controls root knot nematode and soft rot disease complex in carrot. Sci Hortic (amsterdam) 218:56–62. https://doi.org/10.1016/j.scienta.2017.01.051
Rosier A, Medeiros FHV, Bais HP (2018) Defining plant growth promoting rhizobacteria molecular and biochemical networks in beneficial plant-microbe interactions. Plant Soil 428:35–55. https://doi.org/10.1007/s11104-018-3679-5
Sah S, Singh N, Singh R (2017) Iron acquisition in maize (Zea mays L.) using Pseudomonas siderophore. 3 Biotech 7:121. https://doi.org/10.1007/s13205-017-0772-z
Singh SP, Gaur R (2017) Endophytic Streptomyces spp. underscore induction of defense regulatory genes and confers resistance against Sclerotium rolfsii in chickpea. Biol Control 104:44–56. https://doi.org/10.1016/j.biocontrol.2016.10.011
Singh G, Tyagi B, Gupta A et al (2020) Progress Report of AICRP on Wheat and Barley 2019–20, crop Improvement. ICAR-Indian Institute of Wheat and Barley Research, Karnal
Sukkasem P, Kurniawan A, Kao T-C, Chuang H (2018) A multifaceted rhizobacterium Bacillus licheniformis functions as a fungal antagonist and a promoter of plant growth and abiotic stress tolerance. Environ Exp Bot 155:541–551. https://doi.org/10.1016/j.envexpbot.2018.08.005
Swain MR, Naskar SK, Ray RC (2007) Indole-3-acetic acid production and effect on sprouting of yam (Dioscorea rotundata L.) minisetts by Bacillus subtilis isolated from culturable cowdung microflora. Polish J Microbiol 56:103–110
Tabatabai MA, Bremner JM (1969) Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol Biochem 1:301–307. https://doi.org/10.1016/0038-0717(69)90012-1
Vacheron J, Desbrosses G, Bouffaud M-L et al (2013) Plant growth-promoting rhizobacteria and root system functioning. Front Plant Sci 4:356. https://doi.org/10.3389/fpls.2013.00356
Villa-Rodríguez E, Parra-Cota F, Castro-Longoria E et al (2019) Bacillus subtilis TE3: A promising biological control agent against Bipolaris sorokiniana, the causal agent of spot blotch in wheat (Triticum turgidum L. subsp. durum). Biol Control 132:135–143. https://doi.org/10.1016/j.biocontrol.2019.02.012
Weisskopf L, Heller S, Eberl L (2011) Burkholderia species are major inhabitants of white lupin cluster roots. Appl Environ Microbiol 77:7715–7720. https://doi.org/10.1128/AEM.05845-11
Acknowledgements
Authors are thankful to the Director ICAR-Indian Institute of Wheat and Barley Research, Karnal, India for providing necessary facilities to carry out the research work.
Author information
Authors and Affiliations
Contributions
PC, RK: conceptualization, analysis, and writing; PS, AC, RKS: review and editing; GPS: supervision.
Corresponding author
Ethics declarations
Conflict of interest
No conflicts of interest declared.
Additional information
Communicated by M.J. Reigosa.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Chandra, P., Khobra, R., Sundha, P. et al. Bacillus subtilis strain BS87 as a biocontrol agent against spot blotch disease: effect on growth, nutrient status, and antioxidant enzymes in wheat. Acta Physiol Plant 46, 35 (2024). https://doi.org/10.1007/s11738-024-03657-5
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11738-024-03657-5