
Abstract
The study was accompanied to assess the effect of varying levels (0, 20, 40 or 60 t ha−1) of fly ash (FA) and nitrogen (N) (0, 60 or 80 kg ha−1) on Cd, Cr and Pb accumulation, antioxidant activity and yield traits of two mustard cultivars (Brassica juncea cv. ‘Varuna’ and ‘Pusa Bold’). The results showed that antioxidant activity increased concurrently as the FA levels increases. The dose of FA60 accompanied with N60 and N80 lowers enzymatic activity, photosynthetic pigments level, and yield; however, FA at the rate of 40 t ha−1 together with N60 significantly enhanced the crop productivity followed by N80. These results suggest that FA40N60 proved optimal for enhancing crop growth and yield, whereas a high concentration of FA (FA60) caused oxidative stress in both mustard cultivars. Oleic and linoleic acid contents were increased; however, erucic acid content decreased by the application of FA40N60.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Environmental pollution has increased continuously as a consequence of various anthropogenic activities that adversely affected the flora and fauna leading to loss of agricultural productivity worldwide. The generation of fly ash (FA) from coal-fired thermal power plants is one of the major concerns throughout the Earth primarily in the growing nations. At the present time, more than 80 thermal power plants generated about 118 MT year−1 of coal FA in India and it will cross the figure of 440 MT year−1 by the end of 2030 (Ram et al. 2008). Therefore, disposal of such huge amounts of FA and its management is the major problem. Interestingly, it has an alkaline nature that enhances the mineralization of organic matter and promotes the nutrient supply to plants although it also consists of non-essential heavy metals (HMs) including Cd, Pb, Cr, etc. along with various nutrients like Ca, Mg, Fe, Cu, Zn, Ni, B, Na, N and P (Antonkiewicz 2010). Lower levels of FA amendments to the soil positively affect the physico-chemical status of soil, by increasing soil conductivity, organic carbon, pH, and water holding capacity (Pandey and Singh 2010). Similarly, Singh et al. (1997), Pandey et al. (2009a, b) also stated that low levels of FA (5–10%) have positive effect on plant productivity, while the higher dose (20–30%) substantially altered the plant metabolism. The concentrations of HMs including Cd, Cr, Pb, Ni, Fe, and Cu that are already present in the FA, at supra-optimal concentration exhibit metal toxicity in plants. High metal concentration becomes phytotoxic that results in the generation of reactive oxygen species (ROS). It is well documented that HMs induce oxidative stress with the generation of ROS, such as hydroxyl radical (OH·), superoxide (O2−), singlet oxygen (1O2), and hydrogen peroxide (H2O2), either through electron transfer or by metal-mediated inhibition of metabolic reaction (Andresen and Küpper 2013). However, FA may alters protein, amino acid metabolisms, inhibits enzyme activity, dislocates the energy flow in cells, and adversely affects plant growth and productivity also reduces resistance against plant pathogenic injury (Singh et al. 2016). Plants up-regulate the production of enzymatic and non-enzymatic antioxidant system to scavenge the ROS (Noctor et al. 2012) and maintain the basic metabolism under stress conditions.
An adequate dose of fertilizers contributes to the improvement of both quality and quantity of plant growth and productivity (Sakakibara et al. 2006). Nitrogen (N) is a central component of the chlorophyll molecule, amino acid, proteins, nucleic acid, and pigments. An adequate supply of N favours the transformation of carbohydrates into proteins and promotes the formation of protoplasm. Basic metabolic activities can continue only in the presence of the optimum N level. Addition of N enhances the vegetative growth while its deficiency leads to stunted growth and low biomass production. N has potential to increase output by improving photosynthetic efficiency through exploiting natural variation of the processes involved, or by manipulation of the biochemical pathway directly, targeting, for example, RuBP regeneration or catalytic properties of RuBisCO (Reynolds et al. 2009). Addition of N to fly ash affects volatilization of N as ammonia (Choi and Moore 2008).
In India, only about 2% of the total fly ash is used for agricultural purposes. Therefore, the limited use of fly ash in agriculture instead of dumping it as a waste material around the power plants can be both economical and effective in its disposal. Taking into consideration the implications of FA and N amendment on the augmentation of soil properties, metal accumulation, and crop yields, the present study deals with the physico-chemical characteristics of FA and its influence along with different doses of FA and N on metal accumulation, growth and yield of two mustard cultivars.
Materials and methods
Experimental site and climatic condition
The pot experiment was conducted in the net house of the Department of Botany, Aligarh Muslim University, Aligarh, India (27° 89′ N, 78° 08′ E and an elevation of 178.45 m above sea level). The Aligarh city has a monsoon-influenced humid-sub-tropical climate with three principle seasons from intense cold winters, monsoons, to severest hot dry summer. The summers (April–May) are moderately hot with temperature extent up to 46–47 °C, winters (December–February) are quite cool as maximum temperature reached up to 32–35 °C. The annual rainfall went from 600 to 650 mm during late June to October. The average monthly rainfall and temperature for the study area are presented in Fig. 1. The experimental soil is sandy loam, alkaline with moderate water holding capacity and light brown in colour.

Meterological data of Aligarh during the years 2014–2015
Experimental condition and pant material
Seeds of brown mustard (Brassica juncea L. Czern & Coss.) cultivars ‘Varuna’ and ‘Pusa Bold’ were surface-sterilized by soaking them in HgCl2 (0.1%) and then rinsed in de-ionized water and then sown in earthen pots (23-cm diameter) filled with reconstituted soil in the net house under natural day/night conditions. The day and night temperatures were 24/18 ± 3 °C, and relative humidity 75 ± 5%. Once seedlings established, a single healthy plant was retained in each pot. The aim of the experiment was to assess the suitable cultivar and utility of FA as the supplement of nutrients in presence of different levels of N. Before sowing, FA was applied at the rate of 0, 20, 40 or 60 t ha−1 along with N in a dose of 0, 60 or 80 kg ha−1 into the soil and mixed well in each pot. A uniform basal dose of P (20 kg ha−1) and K (30 kg ha−1) was also applied on the basis of earlier reports. N, P, and K were given in the form of urea, single super phosphate and muriate of potash. The experiment was arranged in a completely randomized design (CRD), and three replicates (n = 3) for each treatment were maintained. After 35 days, to record some growth, physiological parameters were determined while yield parameters were studied at harvest maturity (150 days).
Sampling and analysis of fly ash and soil
The soil before filing in pots was collected and autoclaved at 137.9 kPa for 20 min and an adequate amount of organic manure was thoroughly mixed in it. FA was collected from a pond near a thermal power station, Kasimpur, located 15 km away from the experimental area, producing 2.71 MT of fly ash/year. It was dried in the sun before use. The physical–chemical characteristics of FA and soil were determined which are presented in Table 1. Texture of soil/FA was determined by the hydrometer method given by Gee and Bauder (1979). EC and pH were estimated as per the methods given by Jackson (1973). Total organic carbon of soil/FA was analysed by method of Walkley and Black (1934) and NO3–N by the method of Ghosh et al. (1983). Analysis of phosphorus (P) was done calorimetrically after HClO4 digestion and potassium (K) was estimated with the help of flame photometer. Scanning Electron Microscopy (Joel JSM 6510LV, USA) analysis and for elemental analysis, Energy-Dispersive X-Ray Analyzer (EDX) profiling (Joel JSM 6510LV, USA) of FA was also done (Fig. 2).

Scanning electron microscopy (SEM) images and energy dispersion X-ray (EDX) profiling of fly ash
Estimation of MDA, proline and ascorbate content
The lipid peroxidation level in terms of malondialdehyde (MDA) content was determined by Cakmak and Horst (1991). 0.5 g of a fresh leaf was grounded in 10 mL of TCA (0.1%) using mortar and pestle. 4 mL of 0.5% TBA was added to1 mL of the supernatant. The mixture was heated, cooled and centrifuged at 10,000×g for 5 min. The absorbance was read at 532 nm. The MDA content was calculated using the extinction coefficient (155 mM−1 cm−1). Proline content was estimated by the ninhydrin method as described by Bates et al. (1973). 0.3 g of fresh leaves was homogenized in 3% sulphosalicylic acid. The filtrate was acted in response to 1 mL each of glacial acetic acid and acid ninhydrin in a test tube kept in a water bath at 100 °C for 1 h. The reaction was terminated by keeping the test tube on ice. The absorbance of the mixture was read at 520 nm. Ascorbate content was determined by Keller and Schwanger (1977) using 2,6-dichlorophenol-indophenol (DCPIP) dye. Fresh leaves (0.5 g) were homogenized with extracting solution in an ice bath, 1 mL of the supernatant and DCPIP solution were mixed and the OD of the pink solution (Es) was determined at 520 nm wavelength. Now aqueous ascorbic acid solution was added to bleach the pink colour of the dye completely and OD of the bleached solution (Et) was measured at the same wavelength. A blank (Eo), was mixed together and OD was measured at the same wavelength. The total amount of ascorbate was calculated using the following formula:
Estimation of photosynthetic pigments and protein content
The estimation of photosynthetic pigments was done by the method of Arnon (1949) with some modification. Fresh leaves (100 mg) were cut into small pieces and added 10 mL of DMSO in it. The test tubes were incubated at 45 °C for 40 min and read the absorbance at 645 and 663 nm for chlorophyll and at 480 and 510 nm for carotenoid content. Protein content was estimated in each sample through the method of Bradford (1976) using the Coomassie brilliant Blue G250 dye. 200 mg leaves was grounded and centrifuged at 8000×g. Dye solution was added to the supernatant and read the absorbance at 595 nm for not more than 30 min.
Estimation of nitrate reductase activity and carbonic anhydrase activity
Nitrate reductase (NR) activity was determined through the method of Kuo et al. (1982). 1.0 g of fresh leaves was grounded to a powder in liquid N2, and then stored at − 80 °C. The powder was thawed for 10 min at 4 °C and homogenized in 250 mM Tris–HCl buffer. The homogenate was centrifuged at 10,000×g at 4 °C for 30 min. NR activity was analysed by adopting the method of Nakagawa et al. (1984). Carbonic anhydrase (CA) activity was determined by adopting the method of Dwivedi and Randhawa (1974) in the enzyme assay of fresh leaves.
Estimation of growth characteristics, nutrients and heavy metals content
Shoot length and root length were measured by a meter scale. After recording fresh weight, dry weight was determined by drying the plants in oven at 80 °C for 48 h till constant weight. Leaf area per plant was calculated by a leaf area meter (LA 211 Systronics, New Delhi, India). Leaf N, P, and K contents were determined using the method of Lindner (1944), Fiske and Subba Row (1925) and Hald (1946), respectively. For the analysis of Cd, Cr and Pb content, root, leaf and seeds were separately dried in oven at 80 ̊C for 48 h. 1 g of homogenate was digested with a mixture of HNO3, H2SO4, and HClO4 in the ratio of 5:1:1 (v/v) using the method Allen et al. (1986). Cr, Cd and Pb concentration was determined by atomic absorption spectrophotometer (GBC, 932 plus; GBC Scientific Instruments, Braeside, Australia).
Determination of fatty acid content
Oil was extracted from seeds with a Soxhlet apparatus using petroleum ether as a solvent. Fatty acid composition of oil was determined by the method of Kaushik and Agnihotri (1997). The seeds were transmethylated with acetyl chloride. The method is compared with the conventional heating method and extended efficiently for half-seed analysis. The fatty acid methyl esters (FAME) analysis was conducted on Agilent 6890 N gas–liquid chromatography (GLC) equipped with a flame ionization detector (FID).
Statistical analysis
Data were statistically analyzed using SPSS (17.0 Inc. USA) with Analysis of variance (ANOVA) to determine the significance at P < 0.05. In order to determine whether differences among the treatments were significant as compared to control, Duncan’s Multiple Range Test (DMRT) was applied.
Results and discussion
Physicochemical properties of FA and its impact on the performance of Brassica juncea L.
The physicochemical properties and content of extractable element present in FA and soil selected for amendment are shown in Table 1. SEM images (Fig. 2) showed that FA has small-sized particles of varying sizes and shapes. As indicated, applied FA has an alkaline pH (8.9), high water holding capacity and EC, whereas low organic carbon, NO3−, N, P, and K were found in comparison to field soil used in the experiment. FA amendment improved the water holding capacity by 46.39% over control soil (Table 1). Three HMs viz. Cd, Cr and Pb and other metals viz. Fe, Cu, Al were determined in FA by EDX profiling (Fig. 2). Among these HMs, Cd was lowest in concentration. The elements present in the FA also improve the agronomic properties of soil (Singh and Pandey 2013).
Impact of FA and N fertilizer amendment on heavy metals, malondialdehyde content and antioxidant activity of Brassica juncea L.
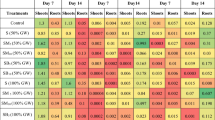
The present study revealed that Cd, Cr, and Pb concentration was significantly higher in different parts of plant studied (viz. root, leaf and seeds) in 60 t ha−1 FA-treated plants than the control (Fig. 3). Lower level of FA showed a decline in uptake of Cd, Cr, and Pb in plant tissue, which increased progressively as the concentration of fly ash increased that might be feasible due to the pH elevation of FA amendment and the results may be with the studies of Petruzzelli et al. (1987). This observation was also corroborated with Pandey et al. (2010); Singh and Pandey (2013) on chickpea and rice, respectively. The heavy metal (Cd, Cr and Pb) concentrations varied in different treatments ranged in roots (0.47–0.91, 1.34–2.35, and 0.78–1.53 mg kg−1 dry mass, respectively), leaves (0.019–0.039, 0.289–0.557, and 0.283–0.563 mg kg−1 dry mass, respectively) and seeds (0.002–0.006, 0.018–0.035, and 0.127–0.244 mg kg−1 dry mass, respectively) of Brassica juncea (Fig. 3). The results revealed that concentrations of these metals were significantly higher in FA60N80 than the control in studied plant parts. The concentration of Cd, Cr, and Pb varies significantly in different parts of plant as the concentration of all three metals was higher in root than the leaves followed by seeds. The level of metals in leaves was found lower as compared to root (Fig. 3), as roots restrict HMs translocation and this could be a probable mechanism of tolerance effective in the root systems (Ernst et al. 1992). Once heavy metals enter into the root, a lesser amount translocate to the shoots because metal sequesters in the vacuoles to make them non-toxic and limits their translocation in the plant system (Shanker et al. 2005; Sinha et al. 2007). Liu et al. (2004) stated that the small proportion of metal accumulated in metal-tolerant plants and subsequently the lowest metal level was detected in the shoot as the present data revealed that ‘Varuna’ accumulated less HMs in the roots, leaves, and seeds than ‘Pusa Bold’ (Fig. 3). This study was further confirmed by Gill et al. (2011) who stated that ‘Varuna’ is tolerant cultivar than ‘Pusa Bold’ in terms of metal accumulation. Greater cross-plant resistance to stress caused by HMs may result from more efficient biosynthesis of sulfur–organic compounds, such as biosynthesis of glucosinolates (Bączek-Kwinta et al. 2011). The present finding is also in line with earlier finding of Yu-Kui et al. (2009) who observed that the Cd content in corn grain was significantly increased with increased concentration of N fertilizer. N application increases the Cu and Pb accumulation in maize and soybean as investigated by Xie et al. (2011); therefore, a decrease in N application may be recommended to decrease the agro-ecological risk associated with Cu and Pb.

Interactive effect of fly ash (FA0, FA20, FA40 and FA60) along with three varying level of N on Cr content (a–c), Pb content (d–f) and Cd content (g–i) of mustard (cv. ‘Varuna’ and ‘Pusa Bold’) at 35 DAS. Data are represented as mean ± SE. Data followed by same letter are not significantly different at p < 0.05
The data indicated that the malondialdehyde content and antioxidant level (ascorbate and proline content) were increased with an increase in FA levels (Fig. 4). HMs present in the higher FA doses could be involved in the overproduction of several oxidative radicals that ultimately leads to oxidative damages to plant cells (Choudhary et al. 2007). The tissue damaged by the oxidative stress due to HMs existing in FA causes increase in production of carbonylated protein and MDA, and the maximum increase of 41.26% MDA content was observed in FA60N80-treated ‘Pusa Bold’ (Fig. 4), free radicals produced by the exclusion of hydrogen from unsaturated fatty acids, thereby triggering disruption of lipid layers and membrane proteins (Navari-Izzo and Quartacci 2001). Plants developed a number of defence system to combat and renovate the damage caused by oxidative stress (Pandey et al. 2010) through the production of antioxidant active compounds that confer the tolerance of plants against HM toxicity. In this study, FA60N80 significantly enhanced ascorbate and proline contents by 69.57% and 55.56%, respectively, in ‘Varuna’ as compared to control (Fig. 4) as these are the important metabolites in plant body responsible for activation of resistance mechanism in plant system under stress conditions which provide stability to plants under the stress condition against free radicals. Sinha et al. (2005) reported that maximum increase was observed in the MDA content at 100% FA-amended soil, and the level of antioxidants, i.e. ascorbate and proline, was also found to increase at 100% FA to combat the oxidative injury.

Interactive effect of fly ash (FA0, FA20, FA40 and FA60) along with three varying level of N on ascorbate content (a), proline content (b), malondialdehyde content (c) and protein content (d) of mustard (cv. ‘Varuna’ and ‘Pusa Bold’) at 35 DAS. Data are represented as mean ± SE. Data followed by same letter are not significantly different at p < 0.05
Impacts of FA and N fertilizer amendment on various attributes of Brassica juncea L.
Addition of FA in the soil affects the physical properties and chemical composition of soil due to the presence of various macro- and micronutrients, including Fe, Cu, Zn, Mn, Mg, Ni, P, K etc. in the FA, which help in plant growth when applied at low concentration in contrast at higher level, while they induce toxicity (Gupta et al. 2002). In the present study, growth parameters of Brassica juncea increased as FA and N fertilizer amendments increased up to FA40N60, whereas higher doses of FA together with N, i.e., FA60N80, followed by FA60N60 have deleterious impact on the growth, photosynthetic pigments, metabolism, and yield of both mustard cultivars. The present findings also strengthen the earlier studies (Singh and Siddiqui 2003; Gupta et al. 2010). This is probably due to the reduction of photosynthetic pigments and low availability of N, P, and K at higher FA doses (Tripathi et al. 2004). HMs along with water and nutrients are transported into the aerial plant parts that directly affect cellular metabolism contributing to the reduction of plant height (Shanker et al. 2005). The fact is that HMs bind on the root cell wall, thereby obstruct cell division and cell elongation. The morphological, biochemical and physiological attributes of mustard cultivars were maximally enhanced by FA40N60 as the amendments of FA up to 40% improved the growth and yield of rice plants, whereas 60% and 100% FA-amended soil shows a decline in plant growth and yield attributes (Singh and Siddiqui, 2003). This adverse effect was due to the presence of higher levels of sulphate, chloride, carbonate and bicarbonate in the FA-amended soil which cause salinity (Dwivedi et al 2007). Mengel and Kirkby (2001) stated that insufficient level of N, diminishes the life cycle of plants, and economic yield as the plant matures early. In this experiment, the extent of percentage increase was found to be higher in ‘Varuna’. Therefore, the enhanced growth under N60 supplemented with FA40 proved most effective. An increase of 18.79% and 21.17% in leaf area and leaf area index, respectively, was also observed over control (Table 2). The yield parameters of Brassica juncea significantly increased and could be due to the fact that N is frequently considered as limiting for biomass production in natural ecosystems (Babatunde and Yongabi 2008). The better growth of rice plants and Brassica campestris was observed in soil amended with FA and N fertilizer, respectively (Datta et al. 2009). Similar observation was made by Arivazhagan et al. (2011) who reported that up to 50 MT ha−1, FA not only significantly enhanced the crop yield, but also ensured the translocation of nutrients to edible parts. FA40N60 increases carotenoid and total chlorophyll contents by 39.45% and 26.96%, respectively, as compared with control (Fig. 5), while high doses of the same decreased these parameters. It was observed that the application of N increased the chlorophyll content in the leaves of the Aloe vera plants (Hazrati et al. 2012). Chlorophyll content reflects the activity of photosynthesis as it is the core pigment of photosynthesis. Moreover, FA has essential nutrient ions which are prerequisites for chlorophyll biosynthesis; therefore, photosynthetic pigments increased (Tripathi et al. 2004). The decreased level of chlorophyll may have been due to down-regulation of chlorophyll biosynthesis by restraining the level of precursors or through targeting –SH group of aminolevulinic acid dehydratase (ALAD) when the metals are being there. The results also showed an increase of 10.79% in protein content of the plants growing on FA40N60 as compared to control (Fig. 4). This difference may be due to a defence mechanism of the plant to cope with FA toxicity (Tripathi et al. 2004). Reduction in protein content was also observed by FA60N80 followed by FA60N60. The decrease in protein content may be due to the increase in the activity of protease and other catabolic enzymes which alter the structure and function of proteins (Sinha et al. 1994; Romero-Puertas et al., 2002). In our study, FA40N60 proved effective in enhancing nitrate reductase (NR) and carbonic anhydrase (CA) activities by 10.31% and 21.79%, respectively, as compared with control (Fig. 5). NR is a regulatory enzyme responsible for plant growth and metabolism (Nazar et al. 2011); however, its activity is inhibited due to reduction in gene expression, the decline in protein synthesis/degradation or by affecting its substrate availability. Inhibition of nitrate reductase activity was also reported in maize by N and Pb, respectively (Sinha et al. 1994). In relation to our results, an increase in carbonic anhydrase activity was also observed in the low level amendment of FA together with N, P, and K fertilizer-grown plants of Brassica nigra (Sahay et al. 2015). This may also be due to the binding ability of these metals with (-SH) groups of enzymes. Sahay et al. (2015) also reported that a higher dose of N fertilizer (N80) together with FA and wastewater reduces the growth, photosynthetic attributes, enzyme activities and yield of mustard plants. The higher photosynthetic rate due to increased leaf area and chlorophyll content leading to enhanced photosynthates under optimum nutrient supply was also evident from the linear regression obtained between (a) shoot dry weight and seed yield, (b) leaf area and seed yield, (c) total chlorophyll content and seed yield, (d) carbonic anhydrase activity and seed yield and (e) nitrogen content and seed yield (Fig. 6).

Interactive effect of fly ash (FA0, FA20, FA40 and FA60) along with three varying level of N on total chlorophyll content (a), total carotenoid content (b), nitrogen, phosphorus and potassium content (c–e), nitrate reductase and carbonic anhydrase content (f and g) of mustard (cv. ‘Varuna’ and ‘Pusa Bold’) at 35 DAS. Data are represented as mean ± SE. Data followed by same letter are not significantly different at p < 0.05

Linear regression between some growth and physiological parameters with seed yield
Impact of FA and N fertilizer amendment on yield and fatty acids of Brassica juncea L.
The results indicated that low level of FA together with N proved beneficial in enhancing seed yield, oil content and oil yield. FA40N60 proved comparatively better as it triggered an increase of 22.90%, 19.91%, and 47.39%, respectively, over control plants (Table 2). An increase of 10.95% and 27.25% in linoleic acid and oleic acid content was observed, whereas decrease of 10.13% of erucic acid over control was noted in the present study (Figs. 7 and 8). The result of present study corroborates with study where a remarkable effect of low levels of FA on yield parameters of rice was noted (Mishra et al. 2007), whereas the higher levels of FA and N, and FA60N80 and FA60N60 decrease the seed yield, oil content, oil yield and fatty acid content in mustard due to the adverse effect (Figs. 7 and 8) as it was also not suitable for yield and production of rice (Singh et al. 2016). The polyunsaturated fatty acids (linoleic and linolenic acids) oxidised resulting reduction in oil quality. Probably metals might disturb lipid synthesis/composition and inactivates thiol-containing enzymes and cofactors (Jones et al. 1987) which adversely affect the oil content and quality. Joshi et al. (1998) reported that supplementation of high N (N90) dose decreased the grain yield, oil content, oil yield as well as fatty acid (oleic acid, linoleic acid, palmitic acid, erucic acid, etc.) content in mustard. Furthermore, Chalkoo et al. (2014) and Iqbal et al. (2015) observed that higher dose of N, P, and K significantly reduced yield characteristics of Brassica. juncea cv. ‘Pusa Bold’ and Capsicum annuum L., respectively. Hence, nutrient quantities and optimization are essential factors to activate better growth and yield performance in plants.

Interactive effect of fly ash (FA0, FA20, FA40 and FA60) along with three varying level of N on oleic acid (a), linoleic acid (b) and erucic acid (c) of mustard (cv. ‘Varuna’ and ‘Pusa Bold’). Data are represented as mean ± SE. Data followed by same letter are not significantly different at p < 0.05

Gas liquid chromatograms showing the effect of fly ash with different dose of N on fatty acid (oleic acid, linoleic acid and erucic acid) content of mustard oil, (a) fatty acid content in control (cv. ‘Varuna’), (b) in optimum combination FA40 × N60 (cv. ‘Varuna’), (c) fatty acid content in control (cv. ‘Pusa Bold’), (d) in optimum combination FA40 × N60 (cv. ‘Pusa Bold’)
Conclusion
It may be concluded from the present study that the lower level of fly ash (FA40) can be useful for agricultural practices as it improved the overall mustard productivity. The result indicated that FA40 N60 proved optimum, it enhanced the growth and yield on the contrary, higher fly ash level (FA60) reduced the above mentioned parameters. Higher doses of fly ash cause oxidative stress with the production of MDA content and HMs accumulation.
Author contributions statement
This work was carried out in collaboration between both authors. FA carried out the experimental work, designed the experiment and carried out the analyses and prepared the manuscript. AI supervised the work and was involved in the design of the experiment, both authors read and approved the final manuscript.
References
Allen SE, Grimshaw HM, Rowland AP (1986) Chemical analysis. In: Moore PD, Chapman SB (eds) Methods in plant ecology. Blackwell Scientific Publications, Oxford, London, pp 285–344
Andresen E, Küpper H (2013) Cadmium toxicity in plants. In: Sigel A, Sigel H, Sigel RKO (eds) Cadmium: from toxicity to essentiality metal ions in life sciences. Springer, New York, pp 395–413
Antonkiewicz J (2010) Effect of sewage sludge and furnace waste on the content of selected elements in the sward of legume-grass mixture. J Elementol 15:435–443. https://doi.org/10.5601/jelem.2010.15.3.435-443
Arivazhagan K, Ravichandran M, Dube SK et al (2011) Effect of coal fly ash on agriculture crops: show case project on use of fly ash in agriculture in and around thermal power station area of national thermal power corporation Ltd., India. World of Coal Ash WOCA, Denver
Arnon DI (1949) Copper enzymes in isolated chloroplast polyphenol oxidase in Beta vulgaris. Plant physiol 24:1–15
Babatunde FE, Yongabi KA (2008) Effect of nitrogen on productivity of Aloe barbadensis L. and its inhibitory effect on Trichophyton rubrum. Adv Hort Sci 22:187–190
Bączek-Kwinta R, Bartoszek A, Kusznierewicz B, Antonkiewicz J (2011) Physiological response of plants and cadmium accumulation in heads of two cultivars of white cabbage. J Elementol 16:355–364. https://doi.org/10.5601/jelem.2011.16.3.01
Bates LS, Walden RT, Tearse ID (1973) Rapid determination of free proline for water stress studies. Plant Soil 39:205–207
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of proteins utilising the principle of protein dye binding. Anal Biochem 72:248–254
Cakmak I, Horst JH (1991) Effects of aluminium on lipid peroxidation, superoxide dismutase, catalase, and peroxidase activities in root tips of soybean (Glycine max). Physiol Planta 83:463–468
Chalkoo S, Sahay S, Inam A, Iqbal S (2014) Application of wastewater irrigation on growth and yield of chilli under nitrogen and phosphorus fertilization. J Plant Nutr 37:1139–1147. https://doi.org/10.1080/01904167.2014.881864
Choi IH, Moore P (2008) Effect of various litter amendments on ammonia volatilization and nitrogen content of poultry litter. J Appl Poultry Res 17:454–462
Choudhary M, Jetley UK, Khan MA, Zutshi S, Fatma T (2007) Effect of heavy metal stress on proline, malondialdehyde, and superoxide dismutase activity in the Cyanobacterium spirulina platensis-S5. Ecotoxicol Environ Saf 66:204
Datta JK, Banerjee A, Saha Sikdar M, Gupta S, Mondal NK (2009) Impact of combined exposure of chemical fertilizer, biofertilizer and compost on growth, physiology and productivity of Brassica campestris in old alluvial soil. J Environ Biol 30:797–800
Dwivedi RS, Randhawa NS (1974) Evaluation of a rapid test for the hidden hunger of zinc in plants. Plant Soil 40:445–451. https://doi.org/10.1007/BF00011531
Dwivedi S, Tripathi RD, Srivastava S, Mishra S, Shukla MK, Tiwari KK, Singh R, Rai UN (2007) Growth performance and biochemical responses of three rice (Oryza sativa L.) cultivars grown in fly ash amendment soil. Chemosphere 67:140–151
Ernst WHO, Verkleij JAC, Schat H (1992) Metal tolerance in plants. Acta Bot Neerl 41:229–248
Fiske CH, Subba Row Y (1925) The colorimetric determination of phosphorus. J Biol Chem 66:375–400
Gee GW, Bauder JW (1979) Particle size analysis by hydrometer: a simplified method for routine textural analysis and a sensitivity test of measured parameters. Soil Sci Soc Am J 43:1004–1007
Ghosh AB, Bajaj JC, Hasan R, Singh D (1983) Soil and water testing methods: a laboratory manual. Indian Agricultural Research Institute (IARI), New Delhi
Gill SS, Khan NA, Tuteja N (2011) Differential cadmium stress tolerance in five Indian mustard (Brassica juncea L.) cultivars. Plant Signal Behav 6:293–300
Gupta DK, Rai UN, Tripathi RD, Inouhe M (2002) Impacts of fly ash on soil and plant responses. J Plant Res 115:401–409
Gupta AK, Mishra RK, Sinha S, Lee B (2010) Growth, metal accumulation and yield performance of Brassica campestris L. (cv. Pusa Jaikisan) grown on soil amended with tannery sludge/fly ash mixture. Ecol Eng 36:981–991
Hald PM (1946) Notes on determination and distribution of sodium and potassium in cells and serum of normal human blood. J Biol Chem 163:429–434
Hazrati S, Sarvestani ZT, Salehi A (2012) The effect of differential nitrogen fertilization on morphological and physiological traits of Aloe vera plants. Int Res J Appl Basic Sci 3:682–687
Iqbal S, Tak HI, Inam A, Inam A, Sahay S, Chalkoo S (2015) Comparative effect of wastewater and groundwater irrigation along with nitrogenous fertilizer on growth, photosynthesis and productivity of chilli (Capsicum annuum L.). J Plant Nutr 38:1006–1021. https://doi.org/10.1080/01904167.2014.991032
Jackson ML (1973) Soil chemistry analysis. Prentice Hall, New Delhi
Jones GJ, Nichols PD, Johns RB, Smith JD (1987) The effect of mercury and cadmium on the fatty acid and sterol composition of the marine diatoms Asterionella glacialis. Phytochemistry 26:1343–1348
Joshi NL, Mali PC, Saxena A (1998) Effect of nitrogen and sulphur application on yield and fatty acid composition of mustard (Brassica juncea) oil. J Agron Crop Sci 180:59–63
Kaushik N, Agnihotri A (1997) Evaluation of improved method for determination of rapeseed-mustard FAMES by GC. Chromatographia 44:97–99
Keller T, Schwanger H (1977) Air pollution and ascorbic acid. Eur J Pathol 7:338–350
Kuo TM, Somers DA, Kleinhofs A, Warner RL (1982) NADH-nitrate reductase in barley leaves: identification and amino acid composition of subunit protein. Biochim Biophys Acta Protein Str Mol Enzymol 708:75–81
Lindner RC (1944) Rapid analytical method for some of the more common inorganic constituents of plant tissues. Plant Physiol 19:76–89
Liu J, Li K, Xu J, Zhang Z, Ma T, Lu X, Yang J, Zhu Q (2004) Lead toxicity, uptake and translocation in different rice cultivars. Plant Sci 165:793–802
Mengel K, Kirkby EA (2001) Principles of plant nutrition, 5th edn. Kluwer Acad. Publishers, Dordrecht, p 849
Mishra M, Sahu RK, Padhy RN (2007) Growth, yield, metabolism and elemental status of rice (Oryza sativa L.) Grown in fly ash amended soils. Ecotoxicol 16:271–278
Nakagawa H, Poulle M, Oaks A (1984) Characterization of nitrate reductase from corn leaves (Zea mays cv W64A* W182E): two molecular form of the enzyme. Plant Physiol 75:285–289
Navari-Izzo F, Quartacci MF (2001) Phytoremediation of metals. Tolerance mechanisms against oxidative stress. Minerva Biotechnol 13:73–83
Nazar R, Iqbal N, Syeed S, Khan NA (2011) Salicylic acid alleviates decrease in photosynthesis under salt stress by enhancing nitrogen and sulphur assimilation and antioxidant metabolism differentially in two mungbean cultivars. J Plant Physiol 168:807–815
Noctor G, Mhamdi A, Chaouch S, Han Y, Neukermans J, Marquez-Garcia B, Queval G, Foyer CH (2012) Glutathione in plants: an integrated overview. Plant Cell Environ 35:454–484
Pandey VC, Singh N (2010) Impact of fly ash incorporation in soil systems. Agric Ecosyst Environ 136:16–27
Pandey VC, Abhilash PC, Singh N (2009a) The Indian perspective of utilizing fly ash in phytoremediation, phyto-management and biomass production. J Environ Manag 10:2943–2958
Pandey VC, Abhilash PC, Upadhyay RN, Tewari DD (2009b) Application of fly ash on the growth performance, translocation of toxic heavy metals within Cajanus cajan L.: implication for safe utilization of fly ash for agricultural production. J Hazard Mater 166:255–259
Pandey VC, Singh JS, Kumar A, Tewari DD (2010) Accumulation of heavy metals by chick pea grown in fly ash treated soil: effects on antioxidants. Clean-Soil Air Water 38:1116–1123
Petruzzelli G, Lubrano L, Cervelli S (1987) Heavy metal uptake by wheat seedlings grown in fly ash amended soils. Water Air Soil Pollut 32:389
Ram LC, Jha SK, Tripathi RC, Masto RE, Selvi VA (2008) Remediation of fly ash landfills through plantation. Remediat J 18:71–90
Reynolds M, Foulkes MJ, Slafer GA, Berry P, Parry MAJ, Snape JW, Angus WJ (2009) Raising yield potential in wheat. J Exp Bot 60:1899–1918
Romero-Puertas MC, Palma JM, Gómez M, del Río LA, Sandalio LM (2002) Cadmium causes the oxidative modification of proteins in pea plants. Plant Cell Environ 25:677–686
Sahay S, Inam A, Inam S, Iqbal S (2015) Modulation in growth, photosynthesis and yield attributes of black mustard (B. nigra cvIC247) by interactive effect of wastewater and fly ash under different NPK levels. Cogent Food Agric 1:1087632
Sakakibara H, Takei K, Hirose N (2006) Interactions between nitrogen and cytokinin in the regulation of metabolism and development. Trends Plant Sci 11:440–448
Shanker AK, Carlos C, Loza-Tavera H, Avudainayagam S (2005) Chromium toxicity in plants. Environ Int 31:739–753
Singh JS, Pandey VC (2013) Fly ash application in nutrient poor agriculture soils: impact on methanotrophs population dynamics and paddy yields. Ecotoxicol Environ Saf 89:43–51
Singh LP, Siddiqui ZA (2003) Effects of fly ash and Helminthosporium oryzae on growth and yield of three cultivars of rice. Bioresour Technol 86:73–78
Singh RP, Tripathi RD, Sinha SK, Maheshwari R, Srivastava HS (1997) Response of higher plants to lead contaminated environment. Chemosphere 34:2467–2493
Singh PK, Tripathi P, Dwivedi S, Awasthi S, Shri M, Chakrabarty D, Tripathi RD (2016) Fly ash augmented soil enhances heavy metal accumulation and phytotoxicity in rice (Oryza sativa L.); a concern for fly ash amendments in agriculture sector. Plant Growth Regul 78:21–30. https://doi.org/10.1007/s10725-015-0070-x
Sinha SK, Srivastava HS, Tripathi TD (1994) Influence of some growth regulators and divalent cations on the inhibition of nitrate reductase activity by lead in maize leaves. Chemosphere 29:1775–1782
Sinha S, Rai UN, Bhatt K, Pandey K, Gupta AK (2005) Fly ash induced oxidative stress and tolerance in Prosopis juliflora L. grown on different amended substrates. Environ Monit Assess 102:447–457
Sinha S, Gupta AK, Bhatt K (2007) Uptake and translocation of metals in fenugreek grown on soil amended with tannery sludge: involvement of antioxidants. Ecotoxicol Environ Saf 672:67–277
Tripathi RD, Vajpayee P, Singh N et al (2004) Efficacy of various amendments for amelioration of fly ash toxicity: growth performance and metal composition of Cassia siamea Lamk. Chemosphere 54:1581–1588
Walkley A, Black IA (1934) An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci 37:29–38. https://doi.org/10.1097/00010694-193401000-00003
Xie W, Zhou J, Wang H, Liu Q, Xia J, Lv X (2011) Cu and Pb accumulation in maize (Zea mays L.) and soybean (Glycine max L.) as affected by N, P and K application. Afr J Agric Res 6:1469–1476
Yu-Kui R, Fu-Suo Z, Jian-bo S (2009) Effects of nitrogen fertilization on heavy metal content of corn grains. Phyton 78:101–104
Acknowledgement
Authors are thankful to the University grant Commission, New Delhi for providing financial assistance to carry out this work.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by R. Baczek-Kwinta.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Ashfaque, F., Inam, A. Accumulation of metals, antioxidant activity, growth and yield attributes of mustard (Brassica juncea L.) grown on soil amendments with fly ash together with inorganic nitrogen fertilizer. Acta Physiol Plant 42, 150 (2020). https://doi.org/10.1007/s11738-020-03139-4
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11738-020-03139-4