
Abstract
Intraspecific variability in morphological and ecophysiological leaf traits might be theorized to be present in declining populations, since they seem to be exposed to stress and plasticity could be advantageous. Here we focused on declining Persian oaks (Quercus brantii Lindl. var. persica (Jaub and Spach) Zohary) in the Zagros Mountains of western Iran, representing the most important tree species of this region. We selected trees with contrasting crown dieback, from healthy to severely defoliated, to investigate the relationships between canopy dieback and leaf morphology, water content and pigments. We also measured esterase and peroxidase, as enzymatic antioxidants and indicators of contrasting genotypes. Trees showing moderate to severe defoliation showed higher leaf mass area (LMA), reduced relative water content (RWC), and lower stomatal density (SD). Increasing LMA indicates a more sclerophyllic structure, according to drier conditions. We did not find significant differences in leaf pigments (chlorophyll a and b, and carotenoids) among crown dieback classes, suggesting that Persian oak trees are able to maintain accurate photochemical efficiency, while reduced RWC and SD suggest hydraulic limitations. Our results do not provide a consistent pattern as regards enzymatic antioxidant defense in Persian oak. Morphological leaf traits would be important drivers of future adaptive evolution in Persian oak, leading to smaller and thicker leaves, which have fitness benefits in dry environments. Nonetheless, drought responses may be critically affecting carbon uptake, as photosynthetic compounds are less effectively used in leaves with higher sclerophylly.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Intraspecific variability is an important mechanism by which trees may cope with environmental heterogeneity in natural populations (Pigliucci 2005; Hoffmann and Sgrö 2011). Under a climate change scenario, water availability may become a main limiting resource, affecting leaf traits and therefore regulating plant growth and survival at global scale (Ackerly et al. 2000; Atkins and Travis 2010; Matesanz et al. 2010). The functional response of trees to drought involves biochemical, physiological, and structural changes at the leaf and whole-plant level (Reich et al. 1999; Niinemets 1999, 2001; Wright and Cannon 2001; Sperlich et al. 2015). The ability of tree populations to overcome such responses, shifting key functional traits, may determine the survival under the drier and more variable climate expected for drought-prone ecosystems (Lázaro-Nogal et al. 2015; Niinemets 2010, 2015).
As individual trees are exposed to different selective pressures, they may differ in leaf morphology and ecophysiological traits along the distribution range (García-Nogales et al. 2016; Laureano et al. 2016). Environmental heterogeneity may influence forest decline patterns (Kabrick et al. 2008) but also it may be related to differentiation in phenotypic plasticity among populations and local adaptation, modulating species range shifts under climate change (Ackerly et al. 2000; Chevin and Lande 2010; Valladares et al. 2014; Lázaro-Nogal et al. 2015).
Leaf morphological and functional traits related to drought tolerance, as well as their plasticity, may enhance plant fitness in water-limited environments (Corcuera et al. 2002; Chaves et al. 2003). Leaf structure is an important determinant of tree-level carbon uptake and water balance (Reich et al. 1999; Niinemets 1999, 2001; Wright et al. 2004). For instance, leaf life span and leaf mass per area are fundamental traits in the carbon economy of plants, representing the investment required per unit leaf area and the duration of the resulting benefit. Tree species on dry and infertile soils usually converge towards higher leaf mass per area, suggesting that this allows longer average leaf life spans to cope with limiting resources (Niinemets 1999, 2001, 2010; Villar et al. 2013; Sperlich et al. 2015).
Oak trees (Quercus spp.) are adapted to drought-prone sites by an ability either to avoid, or to tolerate, water stress, or both (Corcuera et al. 2002; Niinemets 2015; Gómez-Aparicio et al. 2008; Laureano et al. 2016). Generally, they have deep-penetrating root systems, enabling them to maintain relatively high predawn water potentials during drought (Fotelli et al. 2000; Mészáros et al. 2007). Oaks have thick leaves and some have relatively small stomata, both characteristics that favor high water-use efficiency (Abrams 1990). Nonetheless, episodes of drought-induced tree mortality have been recently observed worldwide (Allen et al. 2010), including oak forests (Haavik et al. 2015), as the Persian oak (Quercus brantii Lindl. var. persica (Jaub and Spach) Zohary) in Zagros Mountains (Hosseini et al. 2017). Persian oak covers more than half of the Zagros forest area (west Iran), representing the most important tree species of this region (Hassanzad-Navroodi et al. 2015).
To better our understanding of forest response to current climate change, we must investigate intraspecific variability in leaf morphological and functional traits related to drought tolerance, as well as their plasticity, in water-limited environments (Ackerly et al. 2000; Fotelli et al. 2000; García-Nogales et al. 2016). The ability to respond to water shortage, by modifying leaf structural traits, may confer drought tolerance by limiting water loss, maintaining to some extent light-capture and increasing water-use efficiency (McDowell et al. 2008). At the same time, biochemical responsiveness of leaf physiology may also indicate intra-specific drought responses.
The aim of this study was to investigate the relationships between canopy dieback and morphological and ecophysiological leaf traits of Persian oak in the Zagros forests in west of Iran. Specifically, we aim (1) to determine population differentiation in leaf morphology, water content, and pigments among Persian oak trees showing contrasting canopy dieback; (2) to evaluate whether crown dieback is related to esterase and peroxidase markers, as enzymatic antioxidant defense and indicators of contrasting genotypes; and (3) to discuss the extent to that intra-population variation of these traits may provide estimates of its potential to evolve by natural selection.
Materials and methods
Study area and sampling design
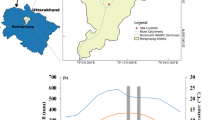
The research was performed in the forested area of the Zagros Range in Shalam, located in the north of Ilam province (Western Iran: latitude 36°31′N, longitude 51°45′E). The region has a Mediterranean climate, mainly affected by westerly disturbances and the Azores High during the cold (November–March) and warm (May–September) seasons, resulting in a clear distinction between a wet winter and a dry summer (Asakereh 2007). Precipitation occurs during an eight-month period from October to May with the maximum during December to March. During June to September, almost no effective precipitation occurs (Supplementary Material, Fig. S1). The mean annual temperature of the region is 16 °C; the mean annual precipitation is 509 mm (Asakereh 2007).
The study area was mainly comprised by forests dominated by Persian oak (Quercus brantii var. persica Lindl). The bedrock was limestone and topography conditions at the sampling sites were homogenous in slope, aspect, and elevation. Crown conditions were evaluated using a crown-dieback classification for each tree, conducted as follows: (1) healthy: canopy damage (defoliation and browning) was less than 5% of the crown length; (2) weak: canopy damage was between 5 and 33% of the crown length; (3) moderate: canopy damage was between 34 and 66% of the crown length; and (4) severe: canopy damage was more than 66% of the crown length (Kabrick et al. 2008; Hosseini et al. 2017). We selected four tree replicates per canopy damage class previously defined (a total of 16 trees were sampled). Within each selected tree, we sampled a representative amount of sun-exposed leaves and 2-year-old branches, randomly selected from the south aspect of the canopy. Thereafter, for each tree, we randomly selected five leaves for morphological traits, water status, and enzyme analysis; and three leaves for chlorophyll and stomatal density. Two-year-old branches were selected to ensure sampling full expanded leaves and full primary-growth woody branches. We randomly selected a total of 10 cm of 2-year-old branches per tree for enzyme analyses. Samples were transferred to the laboratory for further analysis of morphological traits and physiological variables.
Foliar morphology, chemical analyses and assessment of crown condition
Foliar morphological traits were measured on fully expanded leaves from the excised 2-year-old branches (Cornelissen et al. 2003). We measured fresh weight (FW, g) and projected leaf surface area (LA, cm2) with a CI-202 portable laser leaf area meter (CID Bio-Science, Inc., Camas, WA, USA), resolution of 0.1 mm. We oven-dried the leaves at 70 °C for 24 h and weighed the leaves for dry weight (DW, g). We then calculated the percentage of leaf WC as [1 − (DW/FW)] × 100. The relative leaf water content (RWC) was calculated as follows: [(FW − DW)/(TW − DW)] × 100, where FW corresponds to the leaf fresh weight, TW to the leaf turgid weight measured after 12 h of incubation in deionized water at 4 °C in the dark, and DW to the dry weight (24 h at 70 °C). Specific leaf area (SLA; cm2 g−1) was calculated as the ratio of LA to DW. Leaf mass per area (LMA; g m−2) was calculated as the ratio of DW to LA (that is, 1/SLA; Cornelissen et al. 2003).
Chlorophyll a, chlorophyll b, total chlorophylls (a + b), and carotenoids were determined spectrophotometrically, using 80% acetone as a solvent (Lichtenthaler 1987). For quantitative measuring of peroxidase, leaves and 2-year old branches were subjected to a chemical extraction process, followed by optical density readings of the extract, at a wavelength of 420 nm, in a spectrophotometer at the times of 20, 40, 60, 80, and 100 s (Hemeda and Kelin 1990). Average activity of peroxidase per time unit was calculated following Hemeda and Kelin (1990). Qualitative measuring of leaves and 2-year old branches of peroxidase and esterase was performed using vertical polyacrylamide gel electrophoresis (PAGE) method (Hemeda and Kelin 1990). Bands that emerged in each gel were photographed and iso-enzymatic bands were drawn using Microsoft excel software.
Observation and quantification of the number of stomata per surface unit (SD, mm−1) were performed using clear lacquer and transparent adhesive tape (Smith et al. 1989). Due to sclerophyllous leaves of Persian oak, we placed the leaves in boiling water for 10 min to facilitate the quantification of the number of stomata. After removing the leaves from boiling water, we shaved 1 cm2 of the middle part of the leaf and then sprayed the area with clear lacquer (Halfords, Reddith, UK) and left it to dry for 20–25 min. A leaf imprint was obtained with transparent adhesive tape and then placed on a microscope slide (Ferris and Taylor 1994). To count the number of stomata, the leaf impression was observed in an optical microscope at 64 × magnify.
Statistical analysis
The Kolmogorov–Smirnov and Levene tests were used to assess normality and homoscedasticity, respectively. We used one-way analysis of variance (ANOVA) for data showing normal distribution of the residuals. The Kruskal–Wallis test by ranks was used when normal distribution of the residuals might be not assumed, as a non-parametric method (Kruskal and Wallis 1952). Duncan’s multiple range test was applied as a multiple comparison procedure that uses the studentized range distribution to compare sets of means. The procedure consists of a series of pairwise comparisons between means, where each comparison is performed at a significance level defined by the number of means separating the two means compared (Duncan 1955). Iso-enzymatic band patterns of peroxidase and esterase, obtained by the PAGE method in leaves and branches, were analyzed by cluster analysis. Significance level was fixed for p ≤ 0.05. Statistical analyses were made with the SPSS 16.0 software package for Windows (SPSS, Inc., Chicago, IL, USA).
Results
Leaf morphological traits in relation with crown dieback
Both FW and DW showed no statistical differences among crown dieback classes (Kruskal–Wallis test; Table 1). WC and RWC were statistically lower in severely defoliated trees, while LA was statistically lower in declining trees as well as moderate and severely defoliated trees (Table 1).
SLA was statistically lower in moderate and severely defoliated trees; as a consequence, LMA (that is, 1/SLA), was statistically higher in those declining trees classes (Table 1). Leaf pigments (chlorophyll a and b, and carotenoids) did not show statistical differences among crown dieback classes (one-way ANOVA; Table 2). Stomatal density was significantly higher in healthy trees, while leaves and branches peroxidase concentrations did not show not statically differences (Table 2).
Peroxidase and esterase bands obtained in leaves and 2 years old branches using vertical polyacrylamide gel electrophoresis (PAGE) are illustrated in the Fig. 1. Due to the low concentration of the esterase enzyme in the samples, PAGE bands of esterase were not quantified, while iso-enzymatic patterns of peroxidase in leaves (Fig. 2a) and branches (Fig. 2b) were analyzed by cluster analysis. Dendrogram obtained by average linkage showed irregular grouping among the studied trees, according to crown dieback classes (Fig. 3).

Polyacrylamide electropherograms of peroxidase (a, b) and esterase (c, d) extracted from leaves and branches of Persian oak trees

Peroxidase bands obtained by polyacrylamide electropherograms in leaves (a) and branches (b) of Persian oak trees with contrasting crown dieback

Dendrogram using average linkage (between groups) based on peroxidase band patterns of leaves and branches of Persian oak trees with contrasting crown dieback. Inset shows a picture illustrating typical leaves and branches morphology of Persian oak.
Discussion
Contrasting responses among declining and non-declining trees have received increasing attention, mainly focused on morphological and physiological response of tree species to abiotic and biotic stressors (McDowell et al. 2008; Allen et al. 2010; Gómez-Aparicio et al. 2008). Forest decline is evident in symptoms such as loss of foliage, change in leaf size and shape and foliage discoloration (Yannis and Kalliopi 2002; Munné-Bosch and Alegre 2000; Sardans et al. 2008). Field surveys in Zagros range have revealed widespread decline symptoms in Persian oak trees, including crown dieback and mortality (Hosseini et al. 2017), while an understanding of how and why forests exhibit decline will depend on a detailed understanding of the physiological basis.
Intraspecific variability in morphological and biochemical traits (i.e., plasticity) might be theorized to be higher at declining populations, since they seem to be exposed to high environmental variability and stress, where adaptive capacity could be advantageous (Pigliucci 2005; Atkins and Travis 2010; Matesanz et al. 2010; Hoffmann and Sgrö 2011; Valladares et al. 2014).
Mediterranean trees acclimate to water deficits with higher investments in structural compounds, thereby increasing leaf density and succulence (Ogaya and Peñuelas 2006; Poorter et al. 2009; Chaves et al. 2003). Foliar traits are known to be good indicators for the ability of drought-adapted species to respond to decreases in rainfall under climate change (Niinemets and Keenan 2014; Sperlich et al. 2015). We would therefore expect that declining trees show leaf traits associated to drought (Valladares et al. 2014; Lázaro-Nogal et al. 2015). Supporting this hypothesis, we found significant leaf traits variation among trees with contrasting crown dieback (Table 1).
Our results suggest that severe water deficit resulted in increased leaf thickness and reduced LA and consequently in higher LMA (Niinemets 1999). Individuals showing moderate to severe defoliation showed ecophysiological traits aimed at reducing evaporative water losses and enhancing water-use efficiency, such as smaller LA, lower SLA, higher LMA, and lower stomatal density (Table 1). This intra-specific variability suggests that Persian oak is able to adapt to the local conditions, as has been broadly reported in water-limited environments (Ackerly et al. 2000; Wright et al. 2004). Our results suggest some degree of plasticity in underlying morphological and physiological traits, such as leaf mass area (LMA) and stomatal density, which may be related to changing patterns in water-use efficiency and root-to-leaf biomass ratio in severely defoliated Persian oak trees. Here, it should be hypothesized that changing root-to-leaf biomass ratio results mostly from increased leaf-shedding in response to drought (Chaves et al. 2003).
Oak trees display a wide suite of leaf traits associated to leaf life span (Abrams 1990; Reich et al. 1999; Fotelli et al. 2000; Niinemets 2015) and photosynthetic and respiration rates, stomatal conductance, and nitrogen (Fotelli et al. 2000; Sardans et al. 2008; García-Nogales et al. 2016). In Mediterranean environments, sclerophyllous evergreen oaks tend to dominate in habitats that are drier and poorer in nutrients (Gómez-Aparicio et al. 2008; Laureano et al. 2016).
One way to understand tree adaptive capacity to drought might be to explore leaf-trait relationships in different crown dieback conditions. The ability to respond to water shortage by modifying leaf structural traits may confer drought tolerance by limiting water loss maintaining to some extent light-capture (that is, increasing water use efficiency; Quero et al. 2006; García-Nogales et al. 2016). At the same time, biochemical responsiveness of leaf physiology may also indicate intra-specific drought responses (Bussotti 2008).
Persian oak trees studied here responded plastically as regards foliar morphology, suggesting that harshest microsites seemed to have selected, through foliar traits, to cope with water stress (Corcuera et al. 2002; Laureano et al. 2008; Villar and Merino 2001; Aranda et al. 2005). We postulate that foliar morphological traits are reliable proxies for drought acclimation in Persian oak (Table 1). According to several studies, the plasticity of leaf morphology is generally higher than the plasticity of foliar chemistry and assimilation rates over a wide range of woody species (Niinemets 1999, 2001).
Nonetheless, there is also evidence about relatively low response in structural leaf traits but a high one in physiological traits induced by drought in Mediterranean oak species (Aranda et al. 2005; Quero et al. 2006; Ramírez-Valiente et al. 2010). It has been hypothesized that some trees may have low ability to modify structural leaf traits, which usually have a large ontogenetic component, while they might show a high physiological responsiveness to optimize water use efficiency under drought conditions (but see García-Nogales et al. 2016).
LMA is a fundamental trait in the carbon economy of plants, representing the investment required per unit of leaf area (Wright et al. 2002). Increasing LMA is considered as an important strategy for plants living in resource-poor and harsh environments, besides it is also usually associated with low photosynthetic capacity (Reich et al. 1999 and references therein). In our study, declining trees (i.e. those likely to be located at the harshest microsites within the population range) showed high values of LMA, while stomatal density values were the lowest (Table 1). The relationship found between LMA (that is, 1/SLA) and crown dieback agrees with previous studies for several species, reporting that LMA increases (SLA decreases) in water-limited populations or individuals (Niinemets 1999 and references therein).
Higher LMA in evergreens is mainly due to a greater leaf thickness, caused by a larger volume of mesophyll and air spaces (Sperlich et al. 2015). As LMA increases, leaves present usually thick cell walls, limiting the water loss by transpiration (Bussotti 2008; Villar et al. 2013). On the other hand, lower LMA, observed in healthy Persian oaks, could be related to higher leaf cell-wall elasticity, allowing a water-spending strategy, while higher LMA values should reflect a water-saving strategy (i.e., increasing water-use efficiency by foliar sclerophylly) of declining Persian oak trees (Corcuera et al. 2002; Ogaya and Penuelas 2006; Mészáros et al. 2007).
Increasing LMA is commonly related to higher values of leaf thickness and carbon content, indicating a more sclerophyllic structure according to the drier conditions (Niinemets 1999, 2010). Several studies have reported that leaf carbon assimilation capacity per unit area increases with increasing nitrogen content per unit area. Increases in LMA bring about decreases in assimilative leaf compounds and extensive modifications in leaf anatomy, increasing the intercellular transfer resistance to CO2 (Reich et al. 1999; Niinemets 1999, 2001, 2010; Wright and Cannon 2001). As a consequence, foliar sclerophylly in declining Persian oaks may explain the observed higher LMA, suggesting a greater fraction of support tissues in the leaves in combination with declining N concentrations, limiting CO2 diffusion thorough more densely packed mesophyll cells (Quero et al. 2006; Sperlich et al. 2015).
In summary, higher LMA observed in declining trees should be the result of more densely packed cells, restricting CO2 diffusion within the leaf and lowering mass-based photosynthetic rates (Niinemets 1999). These trade-offs, between the investments in structural and assimilative leaf compounds, may be limiting photosynthetic capacity in declining trees, given that leaf N content and photosynthetic capacity are positively correlated, and N concentrations usually decreases with increasing LMA (Reich et al. 1999; Wright and Cannon 2001). Nevertheless, we did not find significant differences in leaf pigments (chlorophyll a and b, and carotenoids) among crown dieback classes (Table 2).
Healthy Persian oak trees differed in leaf structure (higher values for LA, SLA and SD), but similar chlorophyll concentrations and physiological activities (similar values of peroxidase activity), compared with declining trees subjected to the same local conditions of climate. As a consequence, these healthy trees tend to have an efficient deployment of leaf area per unit of biomass invested but they might have higher nitrogen content per unit leaf area (although N was not measured in this study), and consequently, higher assimilation and respiration rates than declining trees (Quero et al. 2006; Poorter et al. 2009; Niinemets and Keenan 2014; Sperlich et al. 2015). Within the intra-population variability, trees with a lower SLA, like declining Persian oaks, trade growth efficiency for persistence. Despite leaf toughness is likely energy expensive, it may provide a canopy better able to withstand physical damage and herbivores (Reich et al. 1999; Wright et al. 2004).
Reduction of pigments content, as a result of either slow synthesis or fast breakdown, has been considered as a typical symptom of oxidative stress (Smirnoff 1993). Higher ratio of chlorophyll a to chlorophyll b has been related to stress, since chlorophyll a shows an important role in photoprotection (Munné-Bosch and Peñuelas 2003). Lower leaf chlorophyll concentrations were obtained in declining sugar maple (Acer saccharum) trees, compared with healthy trees, while observations made during the diurnal cycle of both declining and healthy A. saccharum stands showed no apparent leaf water stress (Liu et al. 1997). Larger inter-annual and within-canopy variability in chlorophyll content was observed in Quercus petraea, compared to more stable values observed in Quercus cerris, which has been related to higher sensitivity to drought (Mészáros et al. 2007). Besides Q. petraea had high rates of photosynthesis even under very low leaf water potentials (Yannis and Kalliopi 2002).
Our results support that drought stress do not induce a significant decreases in leaves pigments, suggesting that Persian oak trees are able to maintain accurate photochemical efficiency (Munné-Bosch and Alegre 2000; Bussotti, 2008; Laureano et al. 2008). Notwithstanding, declining trees reduce leaf water content (WC and RWC, Table 1) and stomatal density, suggesting a hydraulic failure limitation (McDowell et al. 2008).
Nevertheless, as we discuss above, increasing LMA looks to bring about decreases in assimilative leaf compounds and extensive modifications in leaf anatomy that may result in increases in intercellular transfer resistance to CO2 (Poorter et al. 2009). Despite the fact that leaf area reduction may be an effective mechanism to reduce transpiring areas, it may also reduce overall carbon uptake in declining trees (Niinemets and Keenan 2014). This hypothesis is also supported by the lower leaf stomatal density observed in declining Persian oak trees, compared to the healthy ones. Persian oak trees might cope with drought-reducing stomata density, which reduces transpiration (Table 1). This finding appears contradictory within anisohydric species, such as oak species from xeric ecosystems, where leaf water potential declines as soil moisture does with drought (Chaves et al. 2003). As a result, mortality occurs in such plants species mainly by hydraulic failure (Mc Dowell et al. 2008).
In addition to hydraulic failure, drought usually leads to oxidative stress due to stomatal closure, which causes the over-reduction of the photosynthetic electron chain and high formation of reactive oxygen species (ROS) in chloroplasts and mitochondria (Bussotti 2008; Laureano et al. 2008; Ozkur et al. 2009). ROS may disrupt normal metabolism of plants through oxidative damages to lipids, proteins, nucleic acids, photosynthetic pigments, and enzymes (Munné-Bosch and Peñuelas 2003; Ozkur et al. 2009). In order to overcome oxidative stress, plants have developed enzymatic and non-enzymatic antioxidant defense mechanisms, with peroxidases among the most important antioxidant enzymes (Smirnoff 1993). Furthermore, non-enzymatic antioxidative carotenoids can also quench ROS and stabilize photosynthetic complexes (Munné-Bosch and Peñuelas 2003; Bussotti 2008).
Our results do not provide a consistent pattern regarding peroxidase and esterase enzymatic antioxidant defense in Persian oak (Table 2; Figs. 2, 3). On the one hand, esterase concentrations were very low, precluding a significant determination, while on the other hand, peroxidase showed non-significant differences in activity among declining and non-declining trees (Table 2). It also lacked a consistent pattern in the obtained by polyacrylamide electropherograms in leaves and branches (Fig. 3).
Contrasting leaf morphological traits observed in declining Persian oak trees (severely defoliated), with healthy ones, allows us to hypothesize that stressful conditions (that led to tree decline) could be selected for drought-adapted genotypes (Maherali et al. 2010; Atkins and Travis 2010). To the extent to that phenotypic plasticity can play an important role in the performance of populations under climate change, higher plasticity in populations subjected to severe drought could confer advantage to cope with the predicted increase in aridity and climatic variability (Matesanz et al. 2010; Valladares et al. 2014; Lázaro-Nogal et al. 2015).
The utility of isozymes as genetic markers is generally attributed to their frequent polymorphism, codominance, and single gene-Mendelian inheritance (Bussotti 2008). Enzymes with variable substrates, as peroxidase, generally show high variability, but the amount of allozymic polymorphism might be related to environmental conditions (Lázaro-Nogal et al. 2015). Although the selective neutrality of isozymes has been discussed, it should be highly probable that they are adaptive under certain circumstances (Atkins and Travis 2010). However, no evidence of genetic variation for isozymes bands obtained by polyacrylamide electropherograms in leaves and branches was found here within the studied population of Persian oak.
Drought tolerance, like any other trait, may evolve by natural selection, but this requires intra-specific adaptive variability that must be related to fitness and genetically based (Ackerly et al. 2000; Pigliucci 2005; Matesanz et al. 2010). Genetic variation determines the potential for evolutionary response to selection in a specific trait, for instance genetic variation for those plant physiological traits related to drought tolerance (Atkins and Travis 2010; Ramírez-Valiente et al. 2010; Lázaro-Nogal et al. 2015). Lack of significant differences could be partially related to forest decline process itself (Allen et al. 2010). Persian oak forests might be underwent a process of decline leading to smaller and isolated populations, where genetic drift, founder events and population bottlenecks may erode genetic variability and reduce phenotypic plasticity (Atkins and Travis 2010). Moreover, these declining populations, subjected to environmental stress, are subjected to a tradeoff, where the relative metabolic cost of plasticity may increase and exceed the benefits of maintaining adaptive variability (Laureano et al. 2008; Chevin and Lande 2010; Maherali et al. 2010; Ramírez-Valiente et al. 2010).
Conclusion
Variations in leaf structural variables were observed among contrasting crown dieback of Persian oak trees. The current analysis corroborates the already-known findings that leaf morphology plasticity is generally higher than those of foliar chemistry and assimilation rates. We attribute this intra-species variability, observed here among trees with contrasting crown dieback, to acclimation of foliar morphology, although reported for several evergreen sclerophyllic species. Our results support that crown dieback is linked to decreasing RWC and SD, suggesting mainly hydraulic failure limitations, but previous studies also indicates that these relationships may be critically affecting carbon uptake, as photosynthetic compounds are less effectively used in leaves with higher sclerophylly, possibly because of higher diffusive resistance to CO2. No evidence for variation in isozymes obtained by PAGE suggests low evolutionary potential, which may limit future responses of Persian oak to further selective pressures associated with climate change. Despite the lack of intra-population variation for leaves pigments and enzymatic antioxidant, we found intra-population variation for morphological leaf traits, leaves water content, and stomatal density. Therefore, these traits could be subjected to evolution by natural selection and thus allow for further adaptations in Persian oaks.
References
Abrams MD (1990) Adaptations and responses to drought in Quercus species of North America. Tree Physiol 7:227–238
Ackerly D, Dudley S, Sultan SE, Schmitt J, Coleman J, Linder CR, Sandquist DR, Geber MA, Evans AS, Dawson E, Lechowic M (2000) The evolution of plant ecophysiological traits: recent advances and future directions. Bioscience 50:979–995
Allen CD, Macalady AK, Chenchouni H, Bachelet D, McDowell N, Vennetier M, Kitzberger T, Rigling A, Breshears DD, Hogg EH, Gonzalez P, Fensham R, Zhang Z, Castro J, Demidova N, Lim JH, Allard G, Running SW, Semerci A, Cobb N (2010) A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For Ecol Manag 259(4):660–684
Aranda I, Castro L, Pardos M, Gil L, Pardos JA (2005) Effects of the interaction between drought and shade on water relations, gas exchange and morphological traits in cork oak Quercus suber L. seedlings. For Ecol Manag 210:117–129
Asakereh H (2007) Temporal and spatial variations of precipitation in Iran during the last decades. Iran J Geogr Dev 10:145–164
Atkins KE, Travis JMJ (2010) Local adaptation and the evolution of species’ ranges under climate change. J Theor Biol 266:449–457
Bussotti F (2008) Functional leaf traits, plant communities and acclimation processes in relation to oxidative stress in trees: a critical overview. Glob Change Biol 14(11):2727–2739
Chaves MM, Maroco JP, Pereira JS (2003) Understanding plant responses to drought: from genes to the whole plant. Funct Plant Biol 30:239–264
Chevin LM, Lande R (2010) When do adaptive plasticity and genetic evolution prevent extinction of a density-regulated population? Evolution 64:1143–1150
Corcuera L, Camarero JJ, Gil-Pelegrin E (2002) Functional groups in Quercus species derived from the analysis of pressure–volume curves. Trees 16:465–472
Cornelissen JHC, Lavorel S, Garnier E, Diaz S, Buchmann N, Gurvich DE, Reich PB, ter Steege H, Morgan HD, van der Heijden MGA, Pausas JG, Poorter H (2003) A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust J Bot 51:335–380
Duncan DB (1955) Multiple range and multiple F tests. Biometrics 11:1–42
Ferris R, Taylor G (1994) Stomatal characteristics of native herbs following exposure to elevated CO2. Ann Bot 73:447–453
Fotelli MN, Radoglou KM, Constantinidou HIA (2000) Water stress responses of seedlings of four Mediterranean oak species. Tree Physiol 20:1065–1075
García-Nogales A, Linares JC, Laureano RG, Seco JI, Merino J (2016) Range-wide variation in life-history phenotypes: spatiotemporal plasticity across the latitudinal gradient of the evergreen oak Quercus ilex. J Biogeogr 43(12):2366–2379
Gómez-Aparicio L, Pérez-Ramos IM, Mendoza I, Matías L, Quero JL, Castro J, Zamora R, Marañón T (2008) Oak seedling survival and growth along resource gradients in Mediterranean forests: implications for regeneration in current and future environmental scenarios. Oikos 117:1683–1699
Haavik LJ, Billings SA, Guldin JM, Stephen FM (2015) Emergent insects, pathogens and drought shape changing patterns in oak decline in North America and Europe. For Ecol Manag 354:190–205
Hassanzad-Navroodi I, Zarkami R, Basati M, Mohammadi-Limaei S (2015) Quantitative and qualitative characteristics of Persian oak along altitudinal gradation and gradient (case study: Ilam province, Iran). J For Sci 61(7):297–305
Hemeda HM, Kelin BP (1990) Effects of naturally occurring antioxidants on peroxidase activity of vegetables extracts. Food Sci 55:184–185
Hoffmann AA, Sgrö CM (2011) Climate change and evolutionary adaptation. Nature 470:479–485
Hosseini A, Hosseini SM, Linares JC (2017) Site factors and stand conditions associated with Persian oak decline in Zagros mountain forests. For Syst 26(3):e014
Kabrick JM, Dey DC, Jensen RG, Wallendorf M (2008) The role of environmental factors in oak decline and mortality in the Ozark Highlands. For Ecol Manag 255(5–6):1409–1417
Kruskal WH, Wallis WA (1952) Use of ranks in one-criterion variance analysis. J Am Stat Assoc 47(260):583–621
Laureano RG, Lazo YO, Linares JC, Luque A, Martínez F, Seco J, Merino J (2008) The cost of stress resistance: construction and maintenance costs of leaves and roots in two populations of Quercus ilex. Tree Physiol 28:1721–1728
Laureano RG, García-Nogales A, Seco JI, Linares JC, Martínez F, Merino J (2016) Plant maintenance and environmental stress: summarising the effects of contrasting elevation, soil, and latitude on Quercus ilex respiration rates. Plant Soil 409(1):389–403
Lázaro-Nogal A, Matesanz S, Godoy A, Pérez-Trautman F, Gianoli E, Valladares F (2015) Environmental heterogeneity leads to higher plasticity in dry-edge populations of a semi-arid Chilean shrub: insights into climate change responses. J Ecol 103:338–350
Lichtenthaler HK (1987) Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Methods Enzymol 148:350–382
Liu X, Ellsworth DS, Tyree MT (1997) Leaf nutrition and photosynthetic performance of sugar maple (Acer saccharum) in stands with contrasting health conditions. Tree Physiol 17:169–178
Maherali H, Caruso CM, Sherrard ME, Latta RG (2010) Adaptive value and costs of physiological plasticity to soil moisture limitation in recombinant inbred lines of Avena barbata. Am Nat 175:211–224
Matesanz S, Gianoli E, Valladares F (2010) Global change and the evolution of phenotypic plasticity in plants. Ann N Y Acad Sci 2:35–55
McDowell N, Pockman WT, Allen CD, Breshears DD, Cobb N, Kolb T, Plaut J, Sperry J, West A, Williams DG, Yepez EA (2008) Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought? New Phytol 178:719–739
Mészáros I, Veres S, Kanalas P, Oláh V, Szöllösi E, Sárvázi E, Lévai L, Lakatos G (2007) Leaf growth and photosynthetic performance of two co-existing oak species in contrasting growing seasons. Acta Silv Lignaria Hung 3:7–20
Munné-Bosch S, Alegre L (2000) Changes in carotenoids, tocopherols and diterpenes during drought and recovery, and the biological significance of chlorophyll loss in Rosmarinus officinalis plants. Planta 207:925–931
Munné-Bosch S, Peñuelas J (2003) Photo- and antioxidative protection during summer leaf senescence in Pistacia lentiscus L. grown under Mediterranean field conditions. Ann Bot 92:385–391
Niinemets Ü (1999) Components of leaf dry mass per area–thickness and density–alter leaf photosynthetic capacity in reverse directions in woody plants. New Phytol 144:35–47
Niinemets Ü (2001) Global-scale climatic controls of leaf dry mass per area, density, and thickness in trees and shrubs. Ecology 82:453–469
Niinemets Ü (2010) Responses of forest trees to single and multiple environmental stresses from seedlings to mature plants: past stress history, stress interactions, tolerance and acclimation. For Ecol Manag 260:1623–1639
Niinemets Ü (2015) Is there a species spectrum within the world-wide leaf economics spectrum? Major variations in leaf functional traits in the Mediterranean sclerophyll Quercus ilex. New Phytol 205:79–96
Niinemets Ü, Keenan T (2014) Photosynthetic responses to stress in Mediterranean evergreens: mechanisms and models. Environ Exp Bot 103:24–41
Ogaya R, Penuelas J (2006) Contrasting foliar responses to drought in Quercus ilex and Phillyrea latifolia. Biol Plant 50:373–382
Ozkur O, Ozdemir F, Bor M, Turkan I (2009) Physiochemical and antioxidant responses of the perennial xerophyte Capparis ovata Desf to drought. Environ Exp Bot 66:487–492
Pigliucci M (2005) Evolution of phenotypic plasticity: where are we going now? Trends Ecol Evol 20:481–486
Poorter H, Niinemets Ü, Poorter L, Wright IJ, Villar R (2009) Causes and consequences of variation in leaf mass per area (LMA): a meta-analysis. New Phytol 182:565–588
Quero JL, Villar R, Marañón T, Zamora R (2006) Interactions of drought and shade effects on seedlings of four Quercus species: physiological and structural leaf responses. New Phytol 170:819–834
Ramírez-Valiente JA, Valladares F, Delgado-Huertas A, Granados S, Aranda I (2010) Factors affecting cork oak growth under dry conditions: local adaptation and contrasting additive genetic variance within populations. Tree Genet Genomes 7:285–295
Reich PB, Ellsworth DS, Walters MB, Vose JM, Gresham C, Volin JC, Bowman WD (1999) Generality of leaf trait relationships: a test across six biomes. Ecology 80:1955–1969
Sardans J, Penuelas J, Ogaya R (2008) Drought-induced changes in C and N stoichiometry in a Quercus ilex Mediterranean forest. For Sci 54:513–522
Smirnoff N (1993) The role of active oxygen in the response of plants to water deficit and desiccation. New Phytol 125:27–58
Smith S, Weyers JDB, Berry WG (1989) Variation in stomatal characteristics over the lower surface of Commelina communis leaves. Plant Cell Environ 12:653–659
Sperlich D, Chang CT, Penuelas J, Gracia C, Sabate S (2015) Seasonal variability of foliar photosynthetic and morphological traits and drought impacts in a Mediterranean mixed forest. Tree Physiol 35(5):501–520
Valladares F, Matesanz S, Guilhaumon F, Araujo MB, Balaguer L, Garzón M, Cornwell W, Gianoli E, van Kleunen M, Naya DE, Nicotra AB, Poorter H, Zavala MA (2014) The effects of phenotypic plasticity and local adaptation on forecasts of species range shifts under climate change. Ecol Lett 17:1351–1364
Villar R, Merino J (2001) Comparison of leaf construction costs in woody species with differing leaf life-spans in contrasting ecosystems. New Phytol 151:213–226
Villar R, Ruiz-Robleto J, Ubera JL, Poorter H (2013) Exploring variation in leaf mass per area (LMA) from leaf to cell: an anatomical analysis of 26 woody species. Am J Bot 100:1969–1980
Wright IJ, Cannon K (2001) Relationships between leaf lifespan and structural defences in a low-nutrient, sclerophyll flora. Funct Ecol 15:351–359
Wright IJ, Westoby M, Reich PB (2002) Convergence towards higher leaf mass per area in dry and nutrient-poor habitats has different consequences for leaf lifespan. J Ecol 90:534–543
Wright IJ, Reich PB, Westoby M, Ackerly DD, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen JHC, Diemer M, Flexas J, Garnier E, Groom PK, Gulias J, Hikosaka K, Lamont BB, Lee T, Lee W, Lusk C, Midgley JJ, Navas ML, Niinemets U, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov VI, Roumet C, Thomas SC, Tjoelker MG, Veneklaas EJ, Villar R (2004) The worldwide leaf economics spectrum. Nature 428:821–827
Yannis RY, Kalliopi R (2002) Physiological responses of beech and sessile oak in a natural mixed stand during a dry summer. Ann Bot 89:723–730
Author information
Authors and Affiliations
Corresponding author
Additional information
Project funding: Ministerio de Economía y Competitividad, Spain (CGL2013-48843-C2-2-R); European Union FEDER 0087 (TRANSHABITAT); Tarbiat Modares University (Doctorate support to AH).
The online version is available at http://www.springerlink.com
Corresponding editor: Hu Yanbo.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Hosseini, A., Hosseini, S.M. & Linares, J.C. Linking morphological and ecophysiological leaf traits to canopy dieback in Persian oak trees from central Zagros. J. For. Res. 30, 1755–1764 (2019). https://doi.org/10.1007/s11676-018-0805-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11676-018-0805-4