
Abstract
Global elevated temperatures create uncertainties in crop yield production and sustainability, threatening potatoes’ food security and sustainability roles. This has been among the primary research concerns necessitating the need to evaluate potato genotypes’ response to elevated temperature under field conditions. This study assessed potato genotypes’ agronomic and morphological responses to elevated temperatures. The experiments were conducted under field conditions in the 2022 and 2023 cropping seasons in a split-plot design. Two heat treatments were applied; a control treatment in which the plants received the field temperature of the experimental station, and a heat treatment in which the plants received a temperature of + 6.0–10.0 °C depending on the date and time of day. The agronomic and morphological traits evaluated included tuber yield, mean tuber weight, days to emergence, plant height, number of stems per plant, days to physiological maturity, and tuber size distribution. The results showed significant (P < 0.01) variations in the potato genotypes in response to elevated temperature in all traits except days to emergence. Elevated temperature promoted plant height by 36.94%, and days to physiological maturity by 2.55%, while reducing the number of stems per plant by 11.77% and days to emergence by 2.31%. Furthermore, elevated temperature increased total tuber yield by 25.38%, the number of tubers per plant by 18.75%, mean tuber weight by 8.89%, third-class tuber size distribution by 25.95% and malformed tuber ratio by 1.98%, while decreasing first-class tuber size distribution by 10.11%, second-class size distribution by 1.70% and marketable tuber yield by 4.22%. The results of this study demonstrated the impact of heat stress on potato tuber yield and size distribution. The study showed that temperatures around 27.0 °C to 33.0 °C effectively promoted total tuber yield, number of tubers per plant and mean tuber weight; thus, an increase in temperature within the effective range of potato plants promoted yield and yield-related components. This study demonstrates that open-sided field chambers can be a screening tool for heat tolerance of potato genotypes under field conditions. Potato genotypes with less variation in the traits between the heat and control treatment demonstrate heat tolerance and could be used as selection markers for heat-tolerant genotypes.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Potato (Solanum tuberosum L.) is one of the most widely produced agricultural products globally with an annual production of approximately 388 million tons in around 160 countries (FAO 2024). It is the food and vegetable crop that is considered one of the most promising crops for alleviating inadequate and unbalanced nutrition crisis because of its high yield potential and large production area (Aksoy et al. 2021; Zhou et al. 2023) making it the third most common food crop after corn and rice. The modern potato varieties are known to perform well under moderate temperatures around 20 °C with the optimum aerial biomass and underground tubers growth temperature within 20–25 °C and 15–20 °C respectively (Lee et al. 2020). However, the crop plant is very sensitive to environmental fluctuations, and even the same varieties show substantial variation in terms of morphological structure, yield, and quality characteristics under different ecological conditions (Van Dingenen et al. 2019; Hill et al. 2021; Van Nasir and Toth 2021).
In recent years, global climate dynamics have caused frequent and intense unfavourable weather conditions such as rising average annual temperatures. Heat stress is among the primary factors endangering the yield and productivity of cool-temperature crops including potatoes. The effects of heat stress have been significant on potatoes’ morphology, physiological and biochemical processes, and transcriptional regulation, apart from the negative impact on yield (Momčilović 2019). Zhou et al. (2023) have shown that if the current trend of climate dynamics is not alleviated, potato yield is expected to decline by up to 30% due to heat stress. This threatens the food security role of crops, especially in areas where potatoes contribute about 45% of agriculture production (Naawe and Caliskan 2021; Devaux et al. 2021).
Heat stress phenomena result in morphological, physiological, and biochemical changes (Ávila-Valdés et al. 2020) with varying degrees of effect on potato yield. Potato plants’ response to the impact of temperature depends on the cultivar and growth stage (Mokrani et al. 2023). Studies have reported the effect of the rise in temperature on the leaf area and biomass, tuber initiation, and tuber yield of potatoes (Lee et al. 2020). Demirel et al. (2020) stated that climate change will cause high-temperature stress in potatoes resulting in severe yield loss. In the temperate climate, a temperature rise is speculated to promote potato yield (Ávila-Valdés et al. 2020), while at the same time in the tropical and subtropical climates, a temperature rise negatively impacts potato crop yield (Hancock et al. 2014). Under control conditions, Kim and Lee (2019) found that low night temperatures facilitated tuber initiation and increased tuber sizes, while Lee et al. (2020) observed a positive effect of concurrent elevation of temperature and CO2 concentration within the effective range on potato growth, net photosynthetic rate, leaf area, biomass, and tuber yield. Heat stress applied to the potato’s belowground parts causes distinct tuber deformation, secondary growth, and loss of tuber skin colour without affecting the leaf photosynthetic system (Zhou et al. 2023). However, when heat stress is applied on both aerial and below-ground parts, potato yields and quality are negatively affected by causing a reduction in tuber number and mass, increased tuber disorders ratio, and compromised tuber processing and nutritional quality (Mokrani et al. 2023; Zhou et al. 2023). Heat tolerance potato cultivars have been characterised by higher (plant height, growth rate, chlorophyll b content, photosynthetic rate, stomatal conductance, transpiration rate, tuber number, and tuber yield) and lower levels of cell membrane injury (Zhang et al. 2024) while resulting in drastic yield and quality loss in susceptible varieties. However, very few field studies have been conducted on the impact of high temperatures on potatoes (Ávila-Valdés et al. 2020).
In recent times, many potato production zones have been characterised by unpredictability in their weather conditions (Divya et al. 2021, Ademe et al. 2024), especially regarding temperature rise. The sustainability of potato production is hence an essential target and the concern of potato breeders and farmers. Methods of breeding high-yielding, stable, and stress-tolerant potato genotypes to meet global food needs is a main foresight as a long-term adaptation strategy for sustainable food security. Little is however known about potato genotypes’ response to heat stress under field conditions. Therefore, this study was conducted to determine the morphological and agronomic behaviour of potato breeding lines subjected to two different temperatures, which aimed to develop heat-tolerant climate-adapted potato varieties. This will enhance the understanding of the heat stress-mediated response of potato genotypes under field conditions.
Materials and Methods
Plant Materials, Experimental Area, and Conditions
A total of 29 putative potato genotypes (25 advanced potato lines and 4 standard cultivars) (Supplementary Table 1) were selected from the Potato Breeding Program of the Faculty of Agricultural Sciences and Technologies, Nigde Omer Halisdemir University, Türkiye. These genotypes were subjected to a 2-year (2022 and 2023 potato growing seasons) field experiment at the experimental site of the Faculty of Agricultural Sciences and Technologies at Nigde Ömer Halisdemir University located at 37.9698° N and 34.6766° E and an elevation of 1244 m above sea level. The site is characterised by an annual average temperature of 10.3 °C and precipitation of 338 mm. The soil properties of the experimental area are detailed in Supplementary Table 2, while Fig. 1 presents the average temperatures during the study period. The experiment was conducted in a split-plot in a randomised complete block design (RCBD) with the production environment as the main plot (treatments), a 6-m distance between the treatments and the genotypes as sub-plots, each with four replications. Two heat treatments were applied; a control treatment in which the plants received the field temperature of the experimental station and a heat stress treatment in which the plants received a temperature of + 6.0–10.0 °C depending on the date and time of day. The heat stress treatment was applied by creating a plastic chamber measuring 22.5 m in length, 14.0 m in width, and which had a ridge height of 4.6 m. To ensure proper ventilation, ventilation flaps with 50 mesh nets were installed on all four sides. For the first 3 weeks after planting, the ventilation flaps were adjusted to create a temperature difference of approximately 6.0–10 °C higher than the natural environment in the control area, using the Nigde Omer Halisdemir University Meteorological system as a baseline for the adjustment. This temperature gradient was maintained throughout the field study period. The experiment was manually planted at a density of 10 tubers per plot, with one row per plot and an inter-row and intra-row space of 70 cm × 30 cm. Each block replication comprised a length of 300 cm and 30 cm between replication blocks. The experiment was irrigated using drip irrigation at 4-day intervals, except when it rained, in which irrigation was only supplied to the heat stress treatment, as the plastic chambers prevented rainwater from reaching this plot. An ET-176 temperature and humidity data logger (with a precision of ± 0.1 °C and ± 1%) kept 1 m above ground was used to record the temperature and relative humidity of the heat stress treatment, while the data for the control conditions were obtained from the Nigde Ömer Halisdemir University meteorological station. The experimental field was weeded regularly until 15 days before harvesting time.
Environmental Conditions of the Experiments
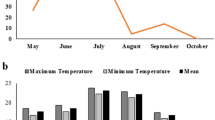
The two growing seasons were characterised by different environmental conditions (Fig. 1). For the control conditions (normal temperature; CT), the first growing season S1 (2022) was characterised by a lower mean temperature, higher relative humidity (RH), and less rainfall (supplementary Table 3). The RH was highest in June and October, while the lowest RH occurred from late July to mid-August. The second season S2 (2023) was characterised by high mean temperature (Fig. 1), higher rains (Supplementary Table 3) and peak RH from planting to mid-June and around the end of October. S1 had a more stable temperature than S2 throughout the season. The mean temperature of the heat stress (elevated temperature treatment; HT) was higher in the S2 than in the S1. The plastic materials used to create the temperature increase in stressful conditions prevented rains in the HT condition (Supplementary Table 3). In S1, the mean temperature for CT and HT was around 25.00 °C and 31.21 °C during the tuber initiation and bulking stages, while in S2, the mean temperature for CT and HT during tuber initiation and bulking was around 28.22 °C and 34.78 °C respectively.

Average monthly temperature (°C) during the field studies
Data Measurement
Day to Emergence (Days)
The time to emergence (days) was monitored after planting, and the emergence time of the potato genotypes was determined when 75% of the plants emerged for the control and heat treatment.
Days to Physiological Maturity (Days)
The physiological maturation time was calculated from emergence to the date at which 75% of the plant leaves in each plot turned yellow.
Number of Stems per Plant and Plant Height (cm)
During the flowering stage, the stems of five random plants per plot were counted and averaged as the number of stems per plant (NSP), while the height (PH) of five random plants per plot was measured using a tape measure at ± 1 cm accuracy.
Agronomic Traits
The agronomic traits measured in this study included the tuber size grading (%), malformed tuber ratio, number of tubers per plant, mean tuber weight (g), marketable tuber yield (t/ha), and total tuber yield (t/ha). The potato tubers were graded in three; first-class tubers (> 50 mm), second-class tubers (30–50 mm), and third-class tubers (< 30 mm). The ratio of potato tubers with secondary growth or disordered forms was classified as malformed tubers ratio out of the total number of tubers per genotype. The number of tubers per plant was calculated by dividing the total number of tubers per plot by the number of plants per plot. Mean tuber weight (g) was calculated by dividing the weight of tubers per plot by the total number of tubers per plot. Marketable tuber yield (t/ha) was calculated by adding and converting the yield of first-class and second-class tubers in kilograms to tons per hectare. The total tuber yield (t/ha) was calculated by adding and converting the yield of first-class tubers, second-class tubers, third-class tubers, and malformed tubers in kilograms to tons per hectare.
Data Processing and Analysis
Excel 365 version was used to process all the data. The statistical analysis included analysis of variance (ANOVA) using Jamovi statistical software (version 2.3) and Origin Lab software (student version 2024). The ANOVA results were considered significant at P < 0.05, and Duncan’s multiple-range test was used to perform mean comparisons.
Results
Effect of Elevated Temperature on the Phenological and Morphological Traits
A significant (P < 0.01) variation in the days to emergence among the potato genotypes was observed for both control (CN) and heat (HT) in both seasons (Table 1). A significant (P < 0.05) variation was observed between the years and year × treatment interactions of the days to emergence (DE); however, no significant variation was observed between the CN and HT groups (Table 1). The elevated temperature caused a 2.04% increase in DE of HT over CN in S1 while a 0.79% decrease in DE of the HT over CN was observed in S2 (Table 2). This could suggest that temperature promoted growth by shortening the DE in S2. In both seasons, a significant (P < 0.01) (Table 1) effect was observed among the genotypes, and between the CN and HT for physiological maturity (PM), number of stems per plant (NSP), and plant height (PH). The PM was observed to have significantly varied between the CN and HT with a 10.91% increase in the mean PM for the HT over the CN in S1, while in S2, an 8.36% decline in the mean PM was observed in HT over the CN (Table 2). The significant (P < 0.01) variations observed in the PH in both seasons resulted in a 36.62% and 37.25% increase of the HT over CN in S1 and S2 respectively (Table 2). A 1.43% increase in NSP was observed in HT over the CN in S1; however, in S2, a 13.20% decline in the NSP was observed in HT over the CN (Table 2).
Impact of Temperature on Tuber Yield and Yield Components
A significant variation in tuber number per plant (NTP) was observed among the genotypes in both the control (CN) group and the heat stress (HT) group in both seasons (Table 3). The NTP was consistently and significantly (P < 0.01) higher in the HT group in all the genotypes than CN in S1 while in the S2 nine (9) out of 29 genotypes recorded higher NTP in the CN than the HT (Table 3). In S1, the NTP ranged from 3.96 tubers to 7.05 tubers with an average of 5.0 tubers for the CN, while for HT, NTP ranged from 5.1 tubers to 9.1 tubers with an average of 6.6 tubers, resulting in a 32% increase in NTP of the HT over the CN. In the second season (S2), the NTP ranged from 5.2 tubers to 9.5 tubers for the CN with an average of 7.3, whereas in the HT, the NTP ranged from 5.3 to 10.8 for the HT with an average of 6.9, resulting a 5.5% increase in the NTP of the CN over the HT (Table 3).
The mean tuber weight (MTW) varied significantly (P < 0.01) (Table 1) among the genotypes in both the CN and HT groups with the HT group recording higher average MTW than the CN group in both seasons (Table 4). In S1, the MTW of the CN ranged from 60.33 g to 75.28 g with an average of 68.12 g, while the HT group ranged from 57.96 g to 83.39 g with an average of 73.65 g, resulting in an 8.12% increase in the MTW of HT over the CN group. In S2, MTW varied from 40.60 g to 82.39 g with an average of 61.03 g in the CN, while in the HT group, it ranged from 49.09 g to 95.20 g with an average of 66.93 g, and an average increase of 9.67% in the MTW of the tubers in the HT group over the CN. In the S1, all but one genotype had higher MTW in the HT than in the CN, while in the S2, 13 genotypes recorded higher mean tuber weight in the CN than in the heat treatment (Table 4). The results from the ANOVA (Table 1) revealed that these differences in MTW were due to genotype, treatment, and the interaction between genotypes and treatment (Table 1). Thus, heat stress treatment caused an increase of 8.90% on average in the mean tuber weight of potato genotypes in both seasons.
A significant (P < 0.01) variation in the total tuber yield (TTY) was observed among the tested potato genotypes in both seasons for the CN and HT (Table 1, Table 5). The TTY was significantly (P < 0.01) higher in the HT than in the CN in S1, while there was no significant difference between the HT and CN in S2. In S1, the TTY ranged from 9.41 to 57.02 t/h for the CN with an average yield of 24.86 t/h, while the HT ranged from 11.79 to 62.46 t/h with an average of 31.45 t/h (Table 5), resulting in a 26.51% increase in TTY in HT over the CN treatment. All the genotypes except MEÇ1302.15 and MEÇ1407.08 recorded higher TTY in HT than in CN in S1 (Table 5). In the S2, the TTY for the CN ranged from 9.17 to 26.42 t/h with an average of 20.38 t/h, while the TTY of the HT ranged from 8.81 to 29.52 t/h with an average of 20.15 t/h (Table 5); no significant (P > 0.05) and consistent variation was observed between the treatments (Table 1). However, an average of 1.13% increase in TTY in CN over the HT was observed. Furthermore, the TTY was higher in S1 than in S2 for both HT and CN treatments, with a 4.48 t/h decline in TTY for CN and an 11.30 t/h decline in HT (Table 5).
Similar to the TTY, the marketable tuber yield (MTY) was significantly (P < 0.05) different between the HT and CN treatment groups in both seasons. In S1, the MTY of the CN ranged from 7.62 to 46.67 t/h with an average of 22.24 t/h while the HT ranged from 8.33 to 50.65 t/h with an average of 25.28 t/h (Table 6). In the S2, the MTY ranged from 9.17 to 27.98 t/h with an average of 17.38 t/h while the HT group ranged from 5.24 to 22.14 t/h with an average of 14.27 t/h. Contrary to S1 where the MTY of the HT group was higher in almost all the genotypes than the MTY of the CN group, in S2, the MTY was higher in CN than in the HT in almost all the genotypes (Table 6). This trend was due to the tuber sizes in S2 where tuber sizes were generally small compared to S1, although the number of tubers was significantly higher in the HT in S2; most of the tubers were unmarketable. This resulted in a 3.03 t//h increase in the MTY of HT over CN in S1 but a 3.12 t/h decline in the MTY of the HT compared to the CN in S2.
Effect of Elevated Temperature on the Tuber Size Distribution and Malformed Tuber Ratio
A significant (P < 0.01) variation was observed for the tuber size distribution and the malformed tuber ratio among the genotypes for both the CN and HT stress treatment groups in both seasons (Table 1). The tuber distribution and malformed tuber ratio also significantly (P < 0.05) varied for the year * treatment, and year * genotype interaction (Table 1). A high percentage of first-class tuber size distribution was observed in the CN over the HT in both seasons (Table 7). In S1, the percentage first-class size distribution ranged from 26.7 to 89.16% with an average of 71.95% for the CN group while the HT ranged from 32.75 to 80.19% with an average of 60.87% (Table 7). In the S2, the percentage first-class size distribution ranged from 23.77 to 54.50% with an average of 40.16% for the CN group while for the HT group, the percentage first-class size distribution ranged from 24.6 to 50.38% with an average of 38.23% (Table 7). In S1, 23 genotypes recorded higher first-class tubers in the CN over the HT, while in S2, 21 genotypes recorded higher first-class tubers in the CN than in the HT. Thus, elevated temperature caused a 15.40% and 4.81% reduction in the first-class tuber size distribution in the first and second seasons respectively.
As revealed in Table 8, in S1, the second-class tuber size ranged from 7.58 to 69.19% with an average of 22.27% for the CN while in the HT it ranged from 4.84 to 54.82% with an average of 24.60%. In S2 on the other hand, the second-class tuber size ranged from 25.33 to 58.45% for the CN and 16.05 to 42.62% for the HT with an average of 34.70% and 30.48% for the CN and HT respectively (Table 8). Thus, the second-class tuber size distribution increased by 10.46% under the HT condition in S1 while in S2, it decreased by 12.16% under the HT conditions. This variation in the second-class tuber size distribution was significant (P < 0.01) among the genotypes for both CN and HT and between the CN and HT in both seasons (Table 8).
The third-class tuber distribution ranged from 0.34 to 4.28% with an average of 1.72%, and 0.44 to 8.03% with an average of 2.35% for the CN and HT respectively in S1 (Table 9). The percentage of the third-class tuber distribution was higher in S2 than in S1. In S2, third-class tuber ranged from 13.45 to 41.14% with an average of 25.23% for the CN and 13.48 to 54.21% with an average of 29.08% for the HT (Table 9). Thus, the elevated temperature significantly (P < 0.01) (Table 1) increased the third-class tuber distribution by 36.63% and 15.26% in the first and second seasons respectively (Table 9). Heat stress increased tuber secondary growth on the tubers resulting in a significantly (P < 0.01) higher malformed tubers in the HT than the CN in both seasons (Table 1, Table 10). The malformed tuber ratio ranged from 0.0 to 9.47% with an average of 2.68% and 0.89 to 30.49% with an average of 15.87% for the CN and HT, respectively, in S1. In S2, the malformed tuber ratio ranged from 0.0 to 23.693% with an average of 1.28% for the CN and from 0.31 to 42.75% with an average of 2.21% for the HT (Table 10).
Discussion
Our study evaluated the agronomic and morphological responses of 29 putative potato genotypes to different temperatures (control temperature; plants received the field temperature of the experimental station, elevated temperature; plants received a temperature of + 6.0–10.0 °C), which demonstrated that the growth and productivity of different potato genotypes was affected by temperature. Contrary to many short-term or control conditions heat stress treatments, we applied heat stress from planting to harvest compared with the control conditions under field conditions. Wolf et al. (1990), Lizana et al. (2017), and Mokrani et al. (2023) employed similar stress conditions. Heat-tolerant potato genotypes could be screened by inducing potato genotypes with high temperatures (Khan et al. 2015; Trapero‐Mozos et al. 2018) and the tolerance of cultivars evaluated using physiological, biochemical, morphological, and agronomic traits (Rykaczewska 2015; Kim and Lee 2019; Zhang et al. 2020; Mokrani et al. 2023; Gautam et al. 2024). Previous studies differentially demonstrated that different intensities of heat stress have varied effects on the plant’s morphological and agronomic traits (Zhao et al. 2020; Dos Santos et al. 2022).
Although several studies reported that heat stress impaired potato tuber yield in varying degrees (Rykaczewska 2015; Kim and Lee 2019; Zhang et al. 2020; Gautam et al. 2024), in this study, heat stress generally increased plant height, time to physiological maturity, number of stems per plant, tuber number per plant, total tuber yield, and mean tuber weight with slight variation between seasons. In the first season (S1) of this study, when the mean temperature of the control (CN) condition was around 21.0–23.5 °C, and of the heat stress (HT) around 29.5–31.67 °C, total tuber yield, tuber number per plant (NTP), and mean tuber weight (MTW) of the genotypes significantly increased in the HT compared with the CN, while in the second season (S2), when the mean temperature of the control (CN) was 27.0–29.25 °C and of the heat stress (HT) around 34.0–38.34 °C, no significant variation in the TTY, MTY, NTP, and MTW was observed. Gautam et al. (2024) stated that temperatures ranging within 12–28 °C are favourable ranges for higher potato yields, this somehow aligns with the 27 to 33 °C found in this study. Lizana et al. (2017) and Wolf et al. (1990) found increased tuber yield and number of tubers per plant of potato varieties when exposed to higher temperatures under field conditions. Similar results of an increase in potato tuber yield, and mean tuber weight were observed by Mokrani et al. (2023) when assessing the effect of temperature on tuber production and carbohydrate partitioning in potatoes; however, their reduced number of tubers per plant contradicts the current study. A drastic reduction in potato yield was observed by Rykaczewska (2015), Kim and Lee (2019), and Zhang et al. (2020) under heat stress in both control and field conditions. These contradictions could be due to differences in cultivar, experimental conditions, and seasonal changes. In this study, the 2022 season was characterised by low temperatures and high relative humidity while the 2023 season was characterised by high temperature and low relative humidity. This combination of weather conditions could have favoured the high yield in 2022; however, the weather conditions in 2023 could have created unfavourable conditions resulting in a generally low yield and relatively high morphological traits. This aligns with the significant potato yield variations under varying weather conditions (Lee et al. 2020) that changes in climatic conditions could either promote or inhibit tuber yield and quality of potatoes (Gautam et al. 2024).
In both seasons, elevated temperature caused an increase in plant height, physiological maturity time, and the number of stems per plant that could be attributed to rapid growth as supported by Siano et al. (2024), heat stress increased plant height and leaf area because of rapid growth. The tuber yield contradiction of this study with the several studies could also be due to differences in the genotype’s response to heat stress and the treatment. Most heat stress studies on potatoes have been done under control conditions (Tang et al. 2018; Lee et al. 2020; Zhang et al. 2024), in which high temperatures caused decreased tuber yield. This could be due to differences in genotypes. The high TTY, NTP, and MTW are buttressed by Siano et al. (2024) who observed high yield in potatoes in a multi-environmental assessment of the impact of heat stress on potato growth and development while contradicting the 13% (Patino-Torres et al. 2021), 4.5 to 34.8% (Mahmud et al. 2021), 67% (Gautam et al. 2024) of yield reduction. The 2022 season produced a higher number of larger tubers compared to the 2023 season while the 2023 season produced a higher number of tubers per plant compared with the 2022 season. The observations of the high number of tubers per plant in this study contradict the decreased formation of tuber number per plant (Zhang et al. 2021) while aligning with the numerous smaller tubers (Mahmud et al. 2021; Gautam et al. 2024) observed in the 2023 season due to delay in tuber initiation due to heat stress as the 2023 season recorded higher temperatures than the 2022 season for both control and heat treatments. Also, the higher number of tubers in 2023 could have resulted in smaller tuber sizes due to strong competition for assimilates partitioned to the tubers. Thus, heat stress could either reduce the average number of tubers per plant or increase the number of tubers per plant based on the severity, timing of heat stress, and genotypic differences for time to tuber initiation of different genotypes (Gautam et al. 2024).
Cultivar, season, and their interaction jointly dictate yield capacity (Benavides-Cardona et al. 2022), and tuber size distribution with the season significantly impacting yield, tuber size distribution, and percentage of malformed tubers. Heat stress had a different and significant impact on the tuber size distribution and malformation of the potato genotypes, resulting in the reduction in marketable tuber yield of the heat stress compared to the control although the heat stress conditions generally had higher total tuber yield than the control. Also, cultivar plays a role in tuber malformation which explains their susceptibility levels to heat stress as was found in this study. Potato cultivars with naturally large and elongated tubers are more susceptible to malformation while cultivars with medium-sized circular tubers are less susceptible to malformation but more susceptible to growth cracks (Siano et al. 2024). Siano et al. 2024 found that the development of a specific tuber physiological disorder in potatoes depends on the environment and the specific (or combination of) abiotic stress present during the growing season. In arid conditions, combined heat and drought stress results in second-growth formation such as heat sprouts, chain tubers, and secondary tuber formation (Zhang et al. 2021). This could explain the highly significant number of malformed tubers in the heat stress conditions compared with the control as influenced by elevated temperature and season.
Periods and duration of heat stress application play an important role in potato yield response (Wolf et al. 1990; Kim and Lee 2019; Lee et al. 2020). Plant exposure to a longer period of heat stress confers heat stress tolerance (Wolf et al. 1990). Studies report that heat stress during the tuber bulking stage has no adverse effect on the tuber yield; however, an increase in temperature for 20 days at the onset of tuber bulking effectively increases the tuber yield of potatoes (Lizana et al. 2017). In this study, the heat stress was applied from the onset of the experiment, this could have conferred stress adaptive mechanisms of the potato genotypes. Also, the observed yield results in this study could be due to the corresponding increase in CO2 as temperature increased. Concurrent elevation of temperature and CO2 within the effective range will exert positive effects on the growth, yield, and photosynthesis of potato crops (Lee et al. 2020) leading to vigorous canopy development that results in enhanced interception of solar radiation.
Conclusion
Heat stress threatens potato production in many regions of the world, and the development of heat-tolerant potato cultivars is needed globally for food sustainability. The impact of elevated temperature on the morphological and agronomic traits was evaluated in this study under field conditions. Plant height, days to physiological maturity, and number of stems per plant were significantly increased under elevated temperatures. These traits can serve as morphological traits for selecting heat-tolerant potato lines. Elevated temperatures around 27.0 to 33.0 °C significantly promoted tuber yield, tuber number per plant, and mean tuber weight compared with temperatures around 20.0 to 24.0 °C but caused no significant effect at 34.0 to 38.0 °C temperature ranges. The tuber size distribution was affected by the range of temperature increase. Temperatures from 27 °C to 33 °C increased the percentage of first and second-grade tubers and the weight of tubers. This may be attributed to an acclimation response to potato to heat stress. Our study demonstrated that the time of heat stress application plays a vital role in determining the heat tolerance capacity of potatoes. In potatoes, elevated temperatures up to 33.0 °C from the onset of planting to harvest confer heat stress tolerance and increase yield and yield-related traits. Thus, the elevation of temperature within the effective range will exert positive effects on the yield, and yield-related components of potato crops.
References
Ademe D, Tesfaye K, Simane B, Zaitchik BF, Alemayehu G, Adgo E (2024) Optimizing agronomic practices to harness climate change impacts on potato production in tropical highland regions. Eur J Agron 152:127021. https://doi.org/10.1016/j.eja.2023.1
Aksoy E, Demirel U, Bakhsh A, Zia MAB, Naeem M, Saeed F, Çalışkan S, Çalışkan ME (2021) Recent advances in potato (Solanum tuberosum L.) breeding. In: Al-Khayri JM, Jain SM, Johnson DV (eds) Advances in Plant Breeding Strategies: Vegetable Crops. Springer International Publishing, pp 409–487. https://doi.org/10.1007/978-3-030-66965-2_10
Ávila-Valdés A, Quinet M, Lutts S, Martínez JP, Lizana XC (2020) Tuber yield and quality responses of potato to moderate temperature increase during tuber bulking under two water availability scenarios. Field Crop Res 251:107786. https://doi.org/10.1016/j.fcr.2020.107786
Benavides-Cardona CA, Marcillo-Paguay CA, Gómez-Gil LF, Romero JV (2022) Physiological and yield response to fertilization of short-cycle Solanum tuberosum cultivars in three high-Andean environments. Rev Fac Nac Agron Medellin 75:10009–10021. https://doi.org/10.15446/rfnam.v75n3.99191
Demirel U, Morris WL, Ducreux LJM, Yavuz C, Asim A, Tindas I, Campbell R, Morris JA, Verrall SR, Hedley PE, Gokce ZNO, Caliskan S, Aksoy E, Caliskan ME, Taylor MA, Hancock RD (2020) Physiological, biochemical, and transcriptional responses to single and combined abiotic stress in stress-tolerant and stress-sensitive potato genotypes. Front Plant Sci 11:169. https://doi.org/10.3389/fpls.2020.00169
Devaux A, Goffart J-P, Kromann P, Andrade-Piedra J, Polar V, Hareau G (2021) The potato of the future: opportunities and challenges in sustainable agri-food systems. Potato Res 64(4):681–720. https://doi.org/10.1007/s11540-021-09501-4
Divya KL, Mhatre PH, Venkatasalam EP, Sudha R (2021) Crop simulation models as decision-supporting tools for sustainable potato production: a review. Potato Res 64(3):387–419. https://doi.org/10.1007/s11540-020-09483-9
Dos Santos TB, Ribas AF, De Souza SGH, Budzinski IGF, Domingues DS (2022) Physiological responses to drought, salinity, and heat stress in plants: a review. Stresses 2(1):113–135. https://doi.org/10.3390/stresses2010009
FAO (2024) Food and Agriculture Organization crop production report:2024
Gautam S, Scheuring DC, Koym JW, Vales MI (2024) Assessing heat tolerance in potatoes: Responses to stressful Texas field locations and controlled contrasting greenhouse conditions. Front Plant Sci 15:1364244. https://doi.org/10.3389/fpls.2024.1364244
Hancock RD, Morris WL, Ducreux LJM, Morris JA, Usman M, Verrall SR, Fuller J, Simpson CG, Zhang R, Hedley PE, Taylor MA (2014) Physiological, biochemical and molecular responses of the potato (Solanum tuberosum L.) plant to moderately elevated temperature. Plant Cell Environ 37(2):439–450. https://doi.org/10.1111/pce.12168
Hill D, Nelson D, Hammond J, Bell L (2021) Morphophysiology of potato (Solanum tuberosum) in response to drought stress: Paving the way forward. Front Plant Sci 11:597554. https://doi.org/10.3389/fpls.2020.597554
Khan MA, Munive S, Bonierbale M (2015) Early generation in vitro assay to identify potato populations and clones tolerant to heat. Plant Cell Tissue Organ Cult 121:45–52. https://doi.org/10.1007/s11240-014-0677-z
Kim YU, Lee BW (2019) Differential mechanisms of potato yield loss induced by high day and night temperatures during tuber initiation and bulking: photosynthesis and tuber growth. Front Plant Sci 10:300. https://doi.org/10.3389/fpls.2019.00300
Lee YH, Sang WG, Baek JK, Kim JH, Shin P, Seo MC, Cho JI (2020) The effect of concurrent elevation in CO2 and temperature on the growth, photosynthesis, and yield of potato crops. PLoS ONE 15(10):e0241081. https://doi.org/10.1371/journal.pone.0241081
Lizana XC, Avila A, Tolaba A, Martinez JP (2017) Field responses of potato to increased temperature during tuber bulking: projection for climate change scenarios, at high-yield environments of Southern Chile. Agric for Meteorol 239:192–201. https://doi.org/10.1016/j.agrformet.2017.03.012
Mahmud A, Jahangir Alam M, Kundu BC, Skalicky M, Matiar Rahman M, Shofiur Rahaman EHM et al (2021) Selection of suitable potato genotypes for late sown heat stress conditions based on field performance and stress tolerance indices. Sustain 13:1–14. https://doi.org/10.3390/su13052770
Mokrani K, Kühn C, Tarchoun N (2023) Temperature effects on tuber production and carbohydrates partitioning in different cultivars during consecutive stages of potato (Solanum tuberosum L.) growth. Potato Res 66(1):275–299. https://doi.org/10.1007/s11540-022-09543-2
Momčilović I (2019) Effects of heat stress on potato productivity and nutritive quality. Hrana i Ishrana 60(2):43–48. https://doi.org/10.5937/hraIsh1902043M
Naawe EK, Caaliskan EM (2021) Effects of genotype by environment interaction on the yield and quality traits of potato breeding lines in the Central Anatolia region of Turkey. In: III 3rd Balkan agriculture congress 2021, Edirne, pp 769–790
Nasir MW, Toth Z (2021) Response of different potato genotypes to drought stress. Agriculture 11(8):763. https://doi.org/10.3390/agriculture11080763
Patino-Torres AJ, Andrade MHML, Guedes ML, Cavallin IC, Pinto CABP, Souza JC et al (2021) Performance of superior potato clones under high and mild temperatures in tropical climates. Agron J 113:2349–2360. https://doi.org/10.1002/agj2.20704
Rykaczewska K (2015) The effect of high temperature occurring in subsequent stages of plant development on potato yield and tuber physiological defects. Am J Potato Res 92(3):339–349. https://doi.org/10.1007/s12230-015-9436-x
Siano AB, Roskruge N, Kerckhoffs H, Sofkova-Bobcheva S (2024) Effects of abiotic stress associated with climate change on potato yield and tuber quality under a multi-environment trial in New Zealand. Potato Res. https://doi.org/10.1007/s11540-024-09695-3
Tang R, Niu S, Zhang G, Chen G, Haroon M, Yang Q, Rajora OP, Li XQ (2018) Physiological and growth responses of potato cultivars to heat stress. Botany 96(12):897–912. https://doi.org/10.1139/cjb-2018-0125
Trapero-Mozos A, Morris WL, Ducreux LJM, McLean K, Stephens J, Torrance L, Bryan GJ, Hancock RD, Taylor MA (2018) Engineering heat tolerance in potato by temperature-dependent expression of a specific allele of HEAT-SHOCK COGNATE 70. Plant Biotechnol J 16(1):197–207. https://doi.org/10.1111/pbi.12760
Van Dingenen J, Hanzalova K, Abd Allah Salem M, Abel C, Seibert T, Giavalisco P, Wahl V (2019) Limited nitrogen availability has cultivar-dependent effects on potato tuber yield and tuber quality traits. Food Chem 288:170–177. https://doi.org/10.1016/j.foodchem.2019.02.113
Wolf S, Olesinski AA, Rudich J, Marani A (1990) Effect of high temperature on photosynthesis in potatoes. Ann Bot 65(2):179–185. https://www.jstor.org/stable/42758299
Zhang G, Tang R, Niu S, Si H, Yang Q, Bizimungu B, Regan S, Li XQ (2020) Effects of earliness on heat stress tolerance in fifty potato cultivars. Am J Potato Res 97(1):23–32. https://doi.org/10.1007/s12230-019-09740-9
Zhang G, Tang R, Niu S, Si H, Yang Q, Rajora OP, Li XQ (2021) Heat-stress-induced sprouting and differential gene expression in growing potato tubers: comparative transcriptomics with that induced by postharvest sprouting. Horticulture Research 8(1):226. https://doi.org/10.1038/s41438-021-00680-2
Zhang S, Ye H, Kong L, Li X, Chen Y, Wang S, Liu B (2024) Multivariate analysis compares and evaluates heat tolerance of potato germplasm. Plants 13(1):142. https://doi.org/10.3390/plants13010142
Zhao J, Lu Z, Wang L, Jin B (2020) Plant responses to heat stress: physiology, transcription, noncoding RNAs, and epigenetics. Int J Mol Sci 22(1):117. https://doi.org/10.3390/ijms22010117
Zhou J, Li K, Li Y, Li M, Guo H (2023) Responses of aerial and belowground parts of different potato (Solanum tuberosum L.) cultivars to heat stress. Plants 12(4):818. https://doi.org/10.3390/plants12040818
Acknowledgements
This research is part of Eric Kuopuobe Naawe’s PhD research. The authors acknowledge the financial support of Nigde Omer Halisdemir University Research Project Units under the project TGT 2021/24-ÖNAP. The help of the Potato Research team of the Faculty of Agricultural Sciences and Technologies during the planting, field maintenance, and harvesting is duly acknowledged.
Funding
Open access funding provided by the Scientific and Technological Research Council of Türkiye (TÜBİTAK).
Author information
Authors and Affiliations
Contributions
M.E.Ç conceived and supervised the research, and M.E.Ç and S.Ç, sourced for funding. E.K.N, I.K, R. I.A., and O.L.G. conducted the research and analysed the data. E. K. N and M.E.Ç drafted the manuscript, and all authors read and approved the final draft.
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Naawe, E.K., Köken, I., Aytekin, R.I. et al. Effects of Elevated Temperature on Agronomic, Morphological, Physiological and Biochemical Characteristics of Potato Genotypes: 1. Agronomic and Morphological traits. Potato Res. (2024). https://doi.org/10.1007/s11540-024-09774-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11540-024-09774-5