
Abstract
Proteins that misfold into hyper-stable/degradation-resistant species during aging may accumulate and disrupt protein homeostasis (i.e., proteostasis), thereby posing a survival risk to any organism. Using the method diagonal two-dimensional (D2D) SDS-PAGE, which separates hyper-stable SDS-resistant proteins at a proteomics level, we analyzed the plasma of healthy young (<30 years) and older (60–80 years) adults. We discovered the presence of soluble SDS-resistant protein aggregates in the plasma of older adults, but found significantly lower levels in the plasma of young adults. We identified the inflammation-related chaperone protein haptoglobin as the main component of the hyper-stable aggregates. This observation is consistent with the growing link between accumulations of protein aggregates and aging across many organisms. It is plausible higher amounts of SDS-resistant protein aggregates in the plasma of older adults may reflect a compromise in proteostasis that may potentially indicate cellular aging and/or disease risk. The results of this study have implications for further understanding the link between aging and the accumulation of protein aggregates, as well as potential for the development of aging-related biomarkers. More broadly, this novel application of D2D SDS-PAGE may be used to identify, quantify, and characterize the degradation-resistant protein aggregates in human plasma or any biological system.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Aging is a biological process in which the essential physiological functions of an organism deteriorate over time culminating in death. All forms of life—from microorganisms to animals—must deal with the terminal role of aging on life. The mechanism of aging in nature is highly diverse, and lifespans across different species vary tremendously (Jones et al. 2014). It has been suggested the presence of protein aggregates may act as aging factors in many organisms throughout different kingdoms (Koga et al. 2011; Labbadia and Morimoto 2014). Interestingly, studies have shown protein aggregation is likely the most important contributor of bacteria senescence (Baig et al. 2014; Lindner et al. 2008; Maisonneuve et al. 2008), and protein sequences in Escherichia coli exhibit a bias towards protein residues with low aggregation potential (Rousseau et al. 2006). Similarly, longer-lived eukaryotic organisms appear to have proteomes that are less likely to aggregate (Tartaglia et al. 2005). Therefore, the phenomenon of aberrant protein misfolding-aggregation is a constant risk throughout the life span of all organisms and a likely contributor to their aging, consistent with the universal presence of molecular chaperones (David 2012; Labbadia and Morimoto 2014).
In recent years, the loss of protein homeostasis (i.e., proteostasis) has been proposed as a primary factor in aging (Koga et al. 2011; Lepez-Otin et al. 2013). Proteostasis involves a “cradle-to-grave” regulation of protein synthesis, folding, trafficking, and degradation. This quality control system, known as the proteostasis network (PN), is fundamental to the well-being of an organism and consists of molecular chaperones, protein degradation machines, and stress response pathways that sense and respond to protein misfolding in cellular compartments (Balch et al. 2008). An ominous consequence of proteostasis disruption is the accumulation of aggregated proteins, thereby accelerating cellular aging and increasing the risk of disease. Indeed, protein aggregation is associated with many age-related disorders, and increased protein oxidation and aggregation are observed in aged organisms (Knoefler et al. 2012; Morimoto 2008; Rana et al. 2013). An even greater challenge to proteostasis would occur if proteins were to aggregate into hyper-stable/degradation-resistant species, as these would be more likely to persist and further compromise the organism.
Since the aberrant increase of degradation-resistant proteins is likely to result in their accumulation during aging, the detection of hyper-stable protein aggregates may be useful to assess biological/cellular aging. We have shown that hyper-stable (i.e., kinetically stable) proteins are resistant to SDS (Manning and Colon 2004; Xia et al. 2012) and have developed a diagonal two-dimensional (D2D) SDS-PAGE method to identify such proteins at a proteomics level (Xia et al. 2007). In this study, we used human plasma as a model system and carried out D2D SDS-PAGE to determine whether the plasma of older age adults contains higher amounts of SDS-resistant aggregates (SRA) than the plasma of younger adults. The results show the plasma of older adults contains significantly higher amounts of SRA that contain mostly the inflammation-related protein haptoglobin.
Material and methods
Sample source
Plasma was purchased from Bioreclamation, Inc (Westbury, NY) (individual and pooled control human plasma), and PrecisionMed, Inc. (San Diego, CA) (individual controls) and also collected from St. Peter’s Hospital (Troy, NY) (individual controls) following consensus guidelines for blood biobanking (Teunissen et al. 2011). All participants were recruited with informed consent and approval by the Institutional Review Board. All samples were immediately aliquoted and snap frozen and kept at −80 °C until processed with a depletion kit. Individual samples from young adults and older adults obtained from PrecisionMed were from individuals who reported to be in general good health. Individuals with diabetes; uncontrolled hypertension; cardiac, malignant, or neurological conditions; sleep disorder; psychiatric disorder; substance abuse history; or evidence of chronic disorder were excluded. Likewise, the samples obtained from St. Peter’s Hospital were from healthy individuals with no known disease or chronic condition. The plasma obtained from Bioreclamation was pooled from 5 to 10 individuals who were tested and cleared from infectious disease markers, but lacked information about their health status regarding chronic diseases or conditions. Because of the pooled nature of these samples, the lack of health status information is not a major concern, especially for the plasma from young adults. In terms of gender and ethnicity, the individual samples were of mixed gender and mostly Caucasians. The lack of strict control of gender and ethnicity is a caveat of the study.
Sample processing and depletion kit
To increase the gel resolution and decrease the interference from the top two abundant proteins, plasma samples were processed to remove albumin and IgG proteins. Two different products were used to ensure the results were not influenced by the depletion method used. Pierce Antibody-Based Albumin/IgG Removal Kit from Thermo (Pierce, Rockford, IL) and BIOHPLC Multiple Affinity Removal Column HSA/IgG (Agilent, Santa Clara, CA) were used to remove albumin/IgG from plasma. We strictly followed vendor’s protocol, including using 40 μl from each plasma sample processed.
D2D SDS-PAGE analysis
Processed plasma was concentrated into 150 μl and incubated for 5 min in SDS sample buffer (pH 6.8) to a final concentration of 45 mM Tris HCl, 1 % SDS, 10 % glycerol, and 0.01 % bromophenol blue at room temperature. Sample was loaded without prior heating onto a well of a 12 % acrylamide gel (16 cm × 14 cm × 1.5 mm). Electrophoresis was performed in a Protean II xi cell (Bio-Rad, Hercules, CA) by using 480 V and 50 mA. The gel was kept at 10 °C by using a circulating water bath. Running buffer contained 25 mM Tris base, 0.2 M glycine, and 0.1 % SDS. After the first-dimension run, the gel strip was cut out and incubated for 10 min in equilibration buffer (45 mM Tris HCl, 1 % SDS, 10 % glycerol, 0.01 % bromophenol blue, pH 6.8) at 98 °C. The gel strip was drained briefly and placed on top of a 12 cm × 14 cm × 2 mm 12 % acrylamide gel. A small amount of 12 % acrylamide solution was used to re-polymerize and fuse the strip to the resolving gel. The second-dimension separation was performed under similar conditions as the first-dimension run, except 65 mA was used per gel. Gels were stained with Coomassie blue (Bio-Rad Biosafe). Destained gels were imaged by a Bio-Rad Gel Doc XR+ system and then analyzed by ImageJ software. Background was subtracted by rolling ball subtraction method with radius of 50 pixels. The relative intensity of each spot/field was calculated by multiplying the area × density.
Spots digestion and MS
Protein spots below the diagonal protein band were picked by using a OneTouch 2D gel spot picker (1.5 mm), then digested with trypsin (Promega, Madison, WI). The resulting peptide mixtures were analyzed using an Agilent 1200-Series LC system coupled to an LTQ-Orbitrap mass spectrometer (Thermo Scientific, Bremen, Germany). The LC system was equipped with a 75-μm ID, 15-μm tip, 105-mm picochip (New Objective, Cambridge, MA) bed packed with 5 μm BioBasic (Thermo Scientific, Bremen, Germany) C18, 300A resin. Sample loading was finished in 2 % buffer B (98 % ACN in 0.1 % formic acid) in 10 min. Elution was achieved with a gradient of 15–90 % B in 75 min. The flow rate was passively split from 0.3 ml/min to 200 nl/min. The mass spectrometer was operated in data-dependent mode to switch between MS and MS/MS. The six most intense ions were selected for fragmentation in the linear ion trap using collision-induced dissociation.
Mass spectrometry data obtained from all LC-MS-MS analysis were searched against Uniprot Homo sapiens Proteome (Proteome ID UP000005640, 2015 June version) using Sequest (Eng et al. 1994) search algorithms through Proteome Discoverer (Thermo Scientific, Bremen, Germany). Enzyme specificity was set as trypsin with a maximum of three missed cleavage allowed. Carbamidomethylation of cysteine and oxidation of methionine were included as variable modifications. The mass error of parent ions was set to 10 ppm and 0.8 Da for fragment ions.
We used commonly accepted criteria for high-confidence peptide identifications (xCorr 1.8 for +1, 2.5 for +2, 3.5 for +3) to screen peptides (Kang et al. 2008). To achieve low false-positive rates of protein identification, each protein included in the results table contains more than two high-confidence unique peptides. Some overlapping spots from different gels contained different proteins. Therefore, in this study, we only report haptoglobin, which was ID in all SRA spots analyzed.
Results
In designing the experiments to compare the presence of SRA between the plasma of older adults and young adults, we took into account various parameters, such as the donor pool and the source of plasma that could affect the results. Most plasma samples were obtained commercially from two different companies, PrecisionMed, Inc. and Bioreclamation, Inc., and some samples were obtained from a local physician. For both young and older adult groups, we included plasma from single adults, as well as plasma pooled from 5 to 10 adults. All plasma samples from single individuals were from healthy controls, but the health status of the donors of the pooled plasma is not known (see “Material and methods” for more details). We also used two different immuno-depletion kits to reduce the two most abundant proteins, human serum albumin (HSA) and gamma globulin (IgG). The conditions for all plasma samples analyzed are summarized in Table 1.
D2D SDS-PAGE of plasma from young and older adults
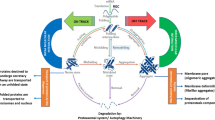
All plasma samples were analyzed by D2D SDS-PAGE (Fig. 1), a method in which (1) unheated protein samples are analyzed by SDS-PAGE, (2) the relevant gel strip is then excised and heated in a boiling buffer solution containing SDS, and (3) the heated strip is analyzed by a second-dimension SDS-PAGE. The resulting 2D gel exhibits a diagonal pattern from the SDS-sensitive proteins migrating the same distance in both SDS-PAGE runs. However, because kinetically stable proteins (KSPs) are SDS-resistant, they migrate less in the first-dimension than in the second-dimension SDS-PAGE. Thus, KSPs migrate on the left side below the gel diagonal (Xia et al. 2007). D2D SDS-PAGE may also be used to identify soluble SRA that enter the stacking gel in the first-dimension gel, but are trapped at the interface of the stacking and resolving gel. This is represented by the red band in the upper part of the 1D gel in Fig. 1. Dissociation of the SRA during boiling in SDS should allow the constitutive proteins to migrate in the 2D gel according to their respective size, as illustrated in Fig. 1 (red bands in 2D gel).

Illustration of how D2D SDS-PAGE may be used for identifying SDS-resistant/kinetically stable aggregates (SRA). Unheated protein samples are analyzed by SDS-PAGE, and the relevant gel strip is then excised and heated in a boiling buffer solution containing SDS. The heated gel strip is analyzed by second-dimension SDS-PAGE. The resulting 2D gel exhibits a diagonal pattern from the SDS-sensitive proteins migrating the same distance in both SDS-PAGE runs. However, SDS-resistant proteins migrate less in the first dimension and thus end up migrating left of the gel diagonal. SDS-resistant soluble aggregates enter the stacking gel in the first-dimension gel but are trapped at the interface of the stacking and resolving gel (red band). Since the boiling step dissociates the SDS-resistant aggregates, the constitutive proteins migrate along the left edge of the 2D gel according to their respective size, as illustrated by the red bands in the 2D gel
All plasma samples from young and older adults were divided into three different groups, labeled in Table 1 as a1–a2, b1–b12, and c1–c8. Figure 2 shows the 2D gels for the a–c plasma groups. Comparison of a1 (young group pooled) and a2 (old group pooled) samples showed an off-diagonal strip in a2 that is nearly absent in a1. The strip contains two obvious protein spots and is located at the interface of the stacking and resolving gels, indicating that it involves a soluble SRA too large to enter the resolving gel. The b1–b12 samples are mostly from young (b1–b4) and old (b5–12) adults whose plasma was HSA- and IgG-depleted with the Pierce antibody-based “top 2” depletion kit. Visual inspection shows the gels from young adults (b1–b4) appear to have less intense SRA compared to gels (b5–b12) from older adults. Panel c shows gels of plasma samples with HSA and IgG depleted by the Agilent bioHPLC Multiple Affinity Removal Column. Young samples (c1–c4) were processed by fast protein liquid chromatography (FPLC) while old (c5–8) samples were processed by HPLC due to the high back-pressure observed for FPLC. As was the case for samples a1 and a2, using HPLC containing metal parts (e.g., loading loop) appears to cause the loss of spots on the diagonal. Despite the experimental differences, gels from young adults (c1–c4) still appear to have less SRA compared to gels (c5–c8) from older adults. Thus, regardless of the plasma source and depletion method used, visual inspections of the gels suggest that the plasma of younger individuals contains less amounts of SRA.

Statistical analysis of gel data
Because of potential differences in loading amounts and other factors, visual inspection of raw band intensity is not reliable for determining whether the amount of SRA is indeed higher in the plasma of older adults. Therefore, each gel was digitalized and analyzed by ImageJ to quantitatively determine the amount of SRA relative to the whole gel. Table 2 shows the results of the analysis for each gel shown in Fig. 1. The statistical significance of the differences of relative intensity between young and old groups was determined by both ANOVA and t test methods (Bailey 2008; Casella 2008). The statistical analyses show SRA/diagonal band (SRA/dia), and SRA/whole protein density in the gel exhibits a significant difference between young and old groups. The average SRA/dia percentage in the old group is 17.7 %, 2.5-fold higher than for the young group average (7.0 %), p < 0.00004. The average SRA/whole in the old group is 7.8 %, about 1.9-fold higher than for the young group average (4.1 %), p < 0.0004. The resulting p value is significantly better than the p = 0.01 threshold for high statistical significance and clearly indicates older adults have higher amounts of SRA than younger adults. The relative amount of SRA in all gels from old and young plasma, respectively, was averaged and is shown in Fig. 3 along with their respective standard deviation values.

Relative amount of SRA present in the plasma of young (blue) and older (red) adults. The bar plot shows the percentage of SRA relative to the proteins on the gel diagonal (SRA/Dia) and all the proteins present (SRA/whole) in the D2D gel. Error bars represent the standard deviation from all the gels shown in Fig. 2. See Table 2 for detailed data and statistical analysis
Protein identification by LC-MS/MS
Several major spots of SRA were chosen for identification (Fig. 4). Protein spots a1–a3 were picked and digested with trypsin, and the resulting peptide mixtures were analyzed by LC-MS/MS (see “Material and methods” and Supplementary Fig. 1). Mass spectrometry data were searched against Uniprot Homo sapiens Proteome using Sequest search algorithms through Proteome Discoverer. Commonly accepted criteria for high-confidence peptide identifications (×Corr 1.8 for +1, 2.5 for +2, 3.5 for +3) were used to screen peptides. To achieve low false-positive rates of protein identification, each protein included in the results table contains more than two high-confidence unique peptides. It was clear various proteins were present in different gels, but only one protein, haptoglobin (Hp), was consistently identified with high confidence in different gels. The various Hp bands identified are consistent with the heterogenous structure of Hp in plasma. Hp is a highly abundant dimeric plasma protein that contains various alpha and beta subunits. The alpha subunit has two versions of 83 (alpha 1) and 142 (alpha 2) residues, whereas the beta subunit has 245 residues. The alpha and beta subunits interact via disulfide bonds to form the Hp monomer, which further dimerizes via two disulfide linkages. Depending on which alpha subunit interacts with the beta subunit, three different Hp dimer isoforms, Hp1-1, Hp2-1, and Hp2-2, may be found in human plasma.

LC-MS/MS analyses reveal the inflammation-related and chaperone protein haptoglobin as the main component of SRA in the plasma of older adults. Representative D2D gel of plasma sample from older adult (c7 in Fig. 2) shows location of SRA-related proteins. Numbered protein strip/bands inside the red box were analyzed by LC-MS/MS (see “Material and methods”). The main protein identified in bands a1–a3 was haptoglobin (Swiss Pro ID p00738), on the basis of 10, 18, and 10 unique peptides, respectively. For MS data, see Supplementary Fig. 1
Discussion
The goal of this study was to determine whether we could use D2D SDS-PAGE to identify differences in the presence of SRA between the plasma of young and older adults. Although other differences may be present, statistical analyses of our gel data demonstrate the most significant difference between the plasma of young and older adults is the presence of a strip of aggregated proteins that migrated at the interface of the stacking and resolving gel. It should be noted as a caveat that the health status of the individuals associated with the pooled plasma sample is not known, and there was no strict gender control. Since only three samples were from pooled plasma, including two for young adults (Table 1: a1 and b1 plasma samples) and one for older adult (Table 1: a2 plasma sample), it seems unlikely the limitations of the present study would have confounded the results. Nevertheless, future studies under more rigorous match control conditions will be needed, especially in a larger study, to explore further the potential biomedical significance of the amount of SRA in plasma.
It is not clear whether the higher amounts of soluble aggregates in the plasma of older adults reflect a deficiency of the proteostasis network (PN) related to aging. However, Hp identified in the SRA is linked to inflammation and chaperone activities. Hp exerts a broad range of anti-inflammatory activities and acts indirectly as a bacteriostatic agent and an antioxidant by binding free hemoglobin (Hb) in plasma and facilitating its clearance by macrophages (Theilgaard-Monch et al. 2006). Hp is also an extracellular chaperone that inhibits the precipitation of stressed proteins by forming solubilized high molecular weight complexes with them. It has been suggested its mode of action is similar to clusterin, which is associated with the clearance of cellular debris and apoptosis (Yerbury et al. 2005). Thus, it is plausible the higher amounts of Hp-containing SRAs identified in older adults may arise from stressed proteins chaperoned by Hp. If that were the case, then reducing the burden of SRA would be a desirable objective that may be achieved through small molecules able to promote the clearance of SRA or reduce their formation in the first place. For example, trehalose, a biologically common disaccharide, has been shown to stabilize proteins, reduce protein aggregation via a chaperone-like function, and activate autophagy (Emanuele 2014). These properties of trehalose make it a promising compound for reducing protein aggregates that contribute to aging and aging-related diseases.
Hp has been proposed as an aging biomarker in mouse (Ding and Kopchick 2011) and has also been shown to increase with age in rat hippocampus (Spagnuolo et al. 2014b). In humans, high levels of Hp in older adults have been observed in some studies (Miura et al. 2011; Shamsi et al. 2012) but not in others (Byerley et al. 2010; Napolioni et al. 2011). Thus, it appears that direct measurement of total Hp expression may not serve as a reliable biomarker of aging. In older adults, high concentration of Hp in plasma is correlated with infection or inflammation (Katz et al. 1990). Since increased inflammation is associated with aging (Ferrucci et al. 2006), the presence of Hp in SRA shown in this study may indicate a state of increased inflammation in older adults. Thus, the presence of degradation-resistant aggregates in plasma, as we have identified in this study, may have better prospects as a biomarker for cellular aging, although this remains to be seen. In humans, it is known that the aging process results in a significant increased risk of disease beginning during middle age. However, it is known that health status does not always correlate with chronological aging, as some people are in good overall health at the age of 80, whereas others are physically and mentally frail by age 70. Therefore, there is a need for fundamental research to discover molecular markers that reflect the true “biological age” (Baker and Sprott 1988; Rowe and Kahn 1987). The existence of centenarians, individuals >100 years old who escape common aging-related diseases, is a fascinating group for future studies (Pareja-Galeano et al. 2015). It would be very interesting to analyze the plasma of such individuals to determine whether they lack the higher amounts of SRA observed here for typical older adults. Such a study could help establish the usefulness of SRA levels in plasma to assess aging-related disease risk during aging.
Hp has been recently linked to the pathogenesis of Alzheimer’s disease (AD) (Song et al. 2015; Spagnuolo et al. 2014a). In one study, the serum levels of Hp were found to be higher in individuals with AD compared to healthy controls (Song et al. 2015). The higher levels of Hp in AD patients suggest high levels of oxidative stress and inflammation, potentially caused by a compromised proteostasis network, leading to increased misfolded and aggregated proteins. In another study, it was found that Hp promotes complex formation between Apo E and beta-amyloid by binding to both of them (Spagnuolo et al. 2014a). The formation of the Apo E-beta-amyloid complex is believed to play an important role in beta-amyloid homeostasis in the brain. Thus, if Hp levels are syphoned to deal with protein aggregation, as suggested by our findings here, this could negatively impair the clearance of amyloid-beta in the brain. Our results showing higher amounts of Hp-containing SRA in the plasma of older adults are consistent with these prior findings linking Hp and AD. Because of the known link between aging-related neurodegenerative disease and protein aggregation, the putative link between Hp and neurodegenerative diseases is particularly intriguing, and our approach for identifying SRA in plasma, CSF, or brain tissue could be applied to further probe this link. Overall, our findings and application of D2D SDS-PAGE provide a unique opportunity to directly probe the amount of SRA and content of Hp and other proteins, not only in older adults with AD but also in people with other types of aging-related neurodegenerative diseases.
This study is to our knowledge the first example of a biophysical property, kinetic stability, being used to discover degradation-resistant protein aggregates in an organism. Because protein aggregation has been suggested to be a common phenomenon associated with aging in all organisms (Koga et al. 2011; Labbadia and Morimoto 2014), D2D SDS-PAGE is a convenient method to discover degradation-resistant aggregates that may accumulate during the aging process of any organism. Furthermore, it offers novel possibilities for exploring the link between Hp-SRA complexes and aging-related ailments, in particular neurodegenerative diseases.
References
Baig UI, Bhadbhade BJ, Watve MG (2014) Evolution of aging and death: what insights bacteria can provide. Q Rev Biol 89:209–223
Bailey RA (2008) Design of comparative experiments. Cambridge University Press, New York
Baker GT, Sprott RL (1988) Biomarkers of aging. Exp Gerontol 23:223–239
Balch WE, Morimoto RI, Dillin A, Kelly JW (2008) Adapting proteostasis for disease intervention. Science 319:916–919
Byerley LO, Leamy L, Tam SW, Chou CW, Ravussin E, Study LHA (2010) Development of a serum profile for healthy aging. Age 32:497–507
Casella G (2008) Statistical design. Springer-Verlag, New York
David DC (2012) Aging and the aggregating proteome. Front Genet 3:247
Ding J, Kopchick JJ (2011) Plasma biomarkers of mouse aging. Age 33:291–307
Emanuele E (2014) Can trehalose prevent neurodegeneration? Insights from experimental studies. Curr Drug Targets 15:551–557
Eng JK, McCormack AL, Yates JR (1994) An approach to correlate tandem mass spectral data of peptides with amino acid sequences in a protein database. J Am Soc Mass Spectrom 5:976–989
Ferrucci L et al. (2006) Relationship of plasma polyunsaturated fatty acids to circulating inflammatory markers. J Clin Endocrinol Metab 91:439–446
Jones OR et al. (2014) Diversity of ageing across the tree of life. Nature 505:169–173
Kang R et al. (2008) Neural palmitoyl-proteomics reveals dynamic synaptic palmitoylation. Nature 456:904–909
Katz PR, Karuza J, Gutman SI, Bartholomew W, Richman G (1990) A comparison between erythrocyte sedimentation rate (ESR) and selected acute-phase proteins in the elderly. Am J Clin Pathol 94:637–640
Knoefler D, Thamsen M, Koniczek M, Niemuth NJ, Diederich AK, Jakob U (2012) Quantitative in vivo redox sensors uncover oxidative stress as an early event in life. Mol Cell 47:767–776
Koga H, Kaushik S, Cuervo AM (2011) Protein homeostasis and aging: the importance of exquisite quality control. Ageing Res Rev 10:205–215
Labbadia J and Morimoto RI. (2014) Proteostasis and longevity: when does aging really begin? F1000Prime Rep 6: 7
Lepez-Otin C, Blasco MA, Partridge L, Serrano M, Kroemer G (2013) The hallmarks of aging. Cell 153:1194–1217
Lindner AB, Madden R, Demarez A, Stewart EJ, Taddei F (2008) Asymmetric segregation of protein aggregates is associated with cellular aging and rejuvenation. Proc Natl Acad Sci U S A 105:3076–3081
Maisonneuve E, Ezraty B, Dukan S (2008) Protein aggregates: an aging factor involved in cell death. J Bacteriol 190:6070–6075
Manning M, Colon W (2004) Structural basis of protein kinetic stability: resistance to sodium dodecyl sulfate suggests a central role for rigidity and a bias toward beta-sheet structure. Biochemistry 43:11248–11254
Miura Y et al. (2011) Proteomic analysis of plasma proteins in Japanese semisuper centenarians. Exp Gerontol 46:81–85
Morimoto RI (2008) Proteotoxic stress and inducible chaperone networks in neurodegenerative disease and aging. Genes Dev 22:1427–1438
Napolioni V, Gianni P, Carpi FM, Concetti F, Lucarini N (2011) Haptoglobin (HP) polymorphisms and human longevity: a cross-sectional association study in a Central Italy population. Clin Chim Acta 412:574–577
Pareja-Galeano H, Alis R, Sanchis-Gomar F, Lucia A, Emanuele E (2015) Vitamin D, precocious acute myocardial infarction, and exceptional longevity. Int J Cardiol 199:405–406
Rana A, Rera M, Walker DW (2013) Parkin overexpression during aging reduces proteotoxicity, alters mitochondrial dynamics, and extends lifespan. Proc Natl Acad Sci U S A 110:8638–8643
Rousseau F, Serrano L, Schymkowitz JW (2006) How evolutionary pressure against protein aggregation shaped chaperone specificity. J Mol Biol 355:1037–1047
Rowe JW, Kahn RL (1987) Human aging—usual and successful. Science 237:143–149
Shamsi KS, Pierce A, Ashton AS, Halade DG, Richardson A, Espinoza SE (2012) Proteomic screening of glycoproteins in human plasma for frailty biomarkers. J Gerontol A Biol Sci Med Sci 67:853–864
Song IU, Kim YD, Chung SW, Cho HJ (2015) Association between serum haptoglobin and the pathogenesis of Alzheimer’s disease. Intern Med 54:453–457
Spagnuolo MS et al. (2014a) Haptoglobin interacts with apolipoprotein E and beta-amyloid and influences their crosstalk. ACS Chem Neurosci 5:837–847
Spagnuolo MS et al. (2014b) Haptoglobin increases with age in rat hippocampus and modulates Apolipoprotein E mediated cholesterol trafficking in neuroblastoma cell lines. Front Cell Neurosci 8:1–13
Tartaglia GG, Pellarin R, Cavalli A, Caflisch A (2005) Organism complexity anti-correlates with proteomic beta-aggregation propensity. Protein Sci 14:2735–2740
Teunissen CE et al. (2011) Consensus guidelines for CSF and blood biobanking for CNS biomarker studies. Mult Scler Int 2011:246412
Theilgaard-Monch K et al. (2006) Haptoglobin is synthesized during granulocyte differentiation, stored in specific granules, and released by neutrophils in response to activation. Blood 108:353–361
Xia K, Manning M, Hesham H, Lin Q, Bystroff C, Colon W (2007) Identifying the subproteome of kinetically stable proteins via diagonal 2D SDS/PAGE. Proc Natl Acad Sci U S A 104:17329–17334
Xia K, Zhang S, Bathrick B, Liu S, Garcia Y, Colon W (2012) Quantifying the kinetic stability of hyperstable proteins via time-dependent SDS trapping. Biochemistry 51:100–107
Yerbury JJ, Rybchyn MS, Easterbrook-Smith SB, Henriques C, Wilson MR (2005) The acute phase protein haptoglobin is a mammalian extracellular chaperone with an action similar to clusterin. Biochemistry 44:10914–10925
Acknowledgments
This work was supported in part by a grant (#1158375) from the National Science Foundation to W. Colón.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
All participants were recruited with informed consent and approval by the Institutional Review Board.
Electronic supplementary material
About this article

Cite this article
Xia, K., Trasatti, H., Wymer, J.P. et al. Increased levels of hyper-stable protein aggregates in plasma of older adults. AGE 38, 56 (2016). https://doi.org/10.1007/s11357-016-9919-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11357-016-9919-9