
Abstract
The increase of affected river reaches by reservoirs has drastically disturbed the original hydrological conditions, and subsequently influenced the nutrient biogeochemistry in the aquatic system, particularly in the cascade reservoir system. To understand the seasonal variation of nitrogen (N) behaviors in cascade reservoirs, hydrochemistry and nitrate dual isotopes (δ15N-NO3− and δ18O-NO3−) were conducted in a karst watershed (Wujiang River) in southwest China. The results showed that NO3−–N accounted for almost 90% of the total dissolved nitrogen (TDN) concentration with high average concentration 3.8 ± 0.4 mg/L among four cascade reservoirs. Higher N concentration (4.0 ± 0.8 mg/L) and larger longitudinal variation were observed in summer than in other seasons. The relationship between the variation of NO3−–N and dual isotopes in the profiles demonstrated that nitrification was dominated transformation, while assimilation contributed significantly in the epilimnion during spring and summer. The high dissolved oxygen concentration in the present cascade reservoirs system prevented the occurrence of N depletion processes in most of the reservoirs. Denitrification occurred in the oldest reservoir during winter with a rate ranging from 18 to 28%. The long-term record of surface water TDN concentration in reservoirs demonstrated an increase from 2.0 to 3.6 mg/L during the past two decades (~ 0.1 mg/L per year). The seasonal nitrate isotopic signature and continuously increased fertilizer application demonstrated that chemical fertilizer contribution significantly influenced NO3−–N concentration in the karst cascade reservoirs. The research highlighted that the notable N increase in karst cascade reservoirs could influence the aquatic health in the region and further investigations were required.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
With the gradual growth of human demand for water source for irrigation and hydroelectric power, increasing rivers have been impounded to construct reservoirs (Zarfl et al. 2015). Globally, about 58,000 large dams (height > 15 m) have been built during the last 60 years (Mulligan et al. 2020), of which 41% global hydropower/reservoirs were built in China (Chen et al. 2019). Globally, large rivers longer than 1000 km remaining free flowing over their entire length only accounted for 37% and less than 23% of these large rivers are not interrupted throughout their flow to the ocean (Grill et al. 2019). For this reason, the original ecological system in rivers has been disturbed drastically, and the resultant environmental effects have drawn increasing attention (Maavara et al. 2020; Schmutz and Moog 2018). For example, water flow speed decrease, the prolonging of hydraulic retention time (HRT, capacity/average annual discharge), and the increase of water level have resulted in sediment, nutrient trapping, and thermal stratification (Winton et al. 2019), which altered the original nutrient biogeochemical cycle and eventually increased the risk to aquatic health (Maavara et al. 2020; Van Cappellen and Maavara 2016;Wang et al. 2010). Moreover, such a situation is further complicated and remains unclear by the fact that many reservoirs are built in cascade configurations, especially in large rivers (Cheng et al. 2019; Ren et al. 2020; Wang et al. 2010; Shi et al. 2020; Zhou et al. 2016).
Nitrogen (N) is one of the most important elements that maintains the element and energy circulation in the river-reservoir aquatic system. Elevated nitrogenous compound concentrations in the aquatic environment would accelerate algae growth and lead to eutrophication of freshwater systems, which may have detrimental consequences for human and ecosystem health (Finlay et al. 2013; Gao et al. 2019). The increasing nitrogen flux and the changing N biogeochemistry in the reservoir have drawn increasing attention from scientists and policy makers (Akbarzadeh et al. 2019; Chen et al. 2020; Harrison et al. 2009; Qin et al. 2020). Furthermore, N undergoes a variety of biogeochemical processes, e.g., nitrification (producing oxidic nitrogen), denitrification, one of depletion processes (converting NO3− into N2 with N2O as intermediate), assimilation processes, and the released from sediments in reservoirs (Bardhan et al. 2017; Ran et al. 2017). Notably, denitrification can make it a more efficient way to remove nitrogen compared to assimilation for relieving eutrophication, which consequently drew widespread attention to the research on river and lake system (Jin et al. 2019; Wenk et al. 2014). To identify the transformation processes and sources of nitrate (NO3−) in the aquatic system, nitrate dual isotopes (δ15N-NO3− and δ18O-NO3−) analysis has been widely utilized based on various nitrate sources with distinctive isotopic compositions (Mayer and Wassenaar 2012; Wenk et al. 2014; Xing and Liu 2016; Xue et al. 2009; Zhang et al. 2018). Moreover, the source contribution from each endmember could be further calculated by the mixing model (stable isotope analysis in R language, SIAR) (Fadhullah et al. 2020; Li et al. 2019).
Generally, as the long retention time increased the uptake and removal of N in lakes or reservoirs, low to mediate N concentrations (less than 2 mg/L) were widely observed (Akbarzadeh et al. 2019; Bardhan et al. 2017; Harrison et al. 2009). For example, one global dataset showed that 90% of 573 lakes have a total dissolved nitrogen (TDN) concentration less than 2 mg/L (Qin et al. 2020). However, high N concentration (> 2 mg/L) is observed in many karst rivers and reservoirs/lakes around the world, which may result in a high-risk threat to the water quality (Davraz et al. 2009; Fenton et al. 2017; Liu et al. 2011; Yue et al. 2018; Zhao et al. 2020). Actually, about 22 million km2 of karst landforms around the world are sensitive to global climate change, accounting for 20% of terrestrial, ice-free land (Sullivan et al. 2019), while the karst area in China accounts for one-seventh of the worldwide occurrence of this landform and one-third of Chinese land area (Green et al. 2019; Wang et al. 2019). Compared with other landscapes, the karst showed distinctive features, e.g., the thin soil layer, unique hydrological structure, including dual hydrological flow paths (diffusion flow and fast flow), and frequent exchange streams between the surface and subterraneous layers (Green et al. 2019; Huang et al. 2020). All characteristics would facilitate soil erosion and nitrogen leaching to the karst aquatic system, which were observed in Southwest (SW) China by high-frequency sensing (Li et al. 2020a, c; Yue et al. 2019). Therefore, it is necessary to investigate the N cycle in reservoirs, especially for the cascade reservoirs in karst regions.
The Wujiang Watershed is in the karst area of SW China, where the cascade reservoirs have been successively impounded since 1979 (Liu 2007; Wang et al. 2010). Notably, those cascade reservoirs have distinctive features including regulation mode, water depth, and running age (Wang et al. 2019b; Yang et al. 2020), which can help to evaluate comprehensively the potential influence of the N cycle in reservoirs with various conditions (Liang et al. 2019; Liu et al. 2011). Therefore, it is necessary to elucidate the seasonal N dynamic variation with varied water level in cascade reservoirs with high N concentration in karst areas, which could help to better understand the environmental effects and to give an indication of the potential risk for further economic evaluation of the river-cascade system (Chen et al. 2020)
In the present study, four cascade reservoirs with different regulation modes in the Wujiang River were studied to explore the spatial-temporal variation on the N biogeochemical processes using the isotopic approach during four seasons. The objectives of this study were to evaluate environmental factors that affect the N dynamics in cascade reservoirs, to clarify the biogeochemical processes against the high N background, and to explore the potential reasons for the high N concentration in karst cascade reservoirs.
Materials and methods
Study area
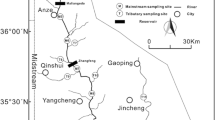
The Wujiang River is the largest tributary on the south upper reaches of the Yangtze River, with a total length of 1050 km and a drainage area of 87,900 km2 including 75.6% karst area (Liu 2007). This watershed is a subtropical monsoon climate with year-on-year average annual temperature of 10–18 °C and rainfall of 800–1400 mm (Li and Ji 2016). The upper reaches of the Wujiang River are in the east of the Yunnan-Guizhou Plateau, while the middle reaches form the main part of the Guizhou Plateau, which are widely exposed with carbonate rocks. The main tributaries include the Liuchong River (the mainstream of the upper stream, W1), the Luojiao River (T1), the Yeji River (T4), and the Pianyan River (T6) in the north, and the Sancha River (T2) and the Maotiao River (T3) in the south (Fig. 1).

Map showing study area and sampling site
Three of the four reservoirs are located in the upper reach, Hongjiadu (HJD), Dongfeng (DF), and Suofengying (SFY), while the Wujiangdu (WJD) is located in middle reaches of the Wujiang River. In particular, HJD is the upper reaches reservoir in Wujiang cascade reservoirs with the longest hydraulic retention time, and WJD is the oldest reservoir built in 1979, which further leaded to the formation of relatively densely populated towns, tourism, and factories in the area. The more detailed characteristics of the four reservoirs are shown in Table S1 (Wang et al. 2010, 2019b).
Sampling and analysis
Water samples were collected in January, April, July, and October 2017, corresponding to winter, spring, summer, and autumn, respectively. These included inflow samples from tributaries samples, profiles samples (at 0.5, 5, 15, 30, 45, 60 m), and outflow samples from reservoir (Fig. 1). These profiles comprised the epilimnion (surface–5 m), metalimnion (5–15 m), and hypolimnion (15–60 m).
Hydrochemistry parameters including water temperature (T), pH, dissolved oxygen (DO), and chlorophyll (Chl) were measured at the sampling site using an automated multiparameter profiler (YSI-EXO). Water samples of about 120 mL were filtered through 0.45-μm cellulose-acetate filters and stored in two 60-mL HDPE bottles at below 4 °C until the laboratory analyses for nutrients concentrations and nitrate dual isotopes were carried out.
The concentrations of nitrogenous species (NO2−, NH4+, NO3−, and TDN) were analyzed using an automatic flow analyzer (Skalar Sans Plus Systems). The dissolved organic nitrogen (DON) concentration was calculated by subtracting DIN (sum of NO2−, NH4+, NO3−) from the TDN. The nitrogen detection limit was 5 μg·L−1 for NO2−–N, 10 μg·L−1 for NO3—N, and NH4+-N. Samples from the profiles (W3, W5, W10, W12, W15, and W17) and all river water samples including inflow water, discharge water, and tributaries were analyzed for nitrate dual isotopes by the denitrifying bacteria method (McIlvin and Casciotti 2011). That is, denitrifying bacteria (Pseudomonas chlororaphis subsp. aureofaciens ATCC 13985) lacking N2O reductase was used to transform nitrate and nitrite to nitrous oxide (N2O) quantitatively, and then N2O was purified using a Trace Gas Pre-concentrator unit (Iso-Prime Ltd., Cheadle Hulme, Cheadle, UK) and analyzed using an isotope ratio mass spectrometer (Iso-Prime, GV, UK). Four international nitrate standards (USGS-32, USGS-34, USGS-35, and IAEA-NO3) were used to calibrate the final isotope data. The standard errors for δ15N-NO3− and δ18O-NO3− were 0.3‰ and 0.5‰, respectively.
Isotope ratios are defined as:
where Rsample and Rstandard represent the ratio of 15N/14N and 18O/16O of samples and standards, respectively. Values of δ15N were reported referred to N2 in the atmosphere, and δ18O values to the Vienna Standard Mean Ocean Water (VSMOW).
Calculation of the extent of denitrification
The isotopic fractionation of denitrification follows the Rayleigh distillation equation which defined as follows (Kendall et al. 2007):
where δr represents the isotope value of the residual nitrate, δ0 is the initial isotope value of nitrate, and ε and f are the isotopic enrichment factor and the remaining fraction of nitrate, respectively (Kendall et al. 2007).
Results
Longitudinal variation of hydrochemistry and DOC concentration
Figure 2 shows the temporal and spatial variation of T, Chl, DO, and DOC. The annual longitudinal variation of hydrochemistry in surface water revealed that T and DO in the reservoir were higher than those of the inflow and outflow water (Wang et al. 2019b). T, DO, and DOC showed more obvious fluctuation in reservoirs than in rivers. Hydrochemistry in profiles showed homogeneity in winter and autumn, while high Chl concentration and significant thermal stratification appeared during the growing season (spring and summer) (Fig. 2). Meanwhile, DO in the profile showed a greater decrease compared to the surface water and were almost higher than 2 mg/L. Nevertheless, the anaerobic water layer was observed in the bottom water around the dam of WJD in winter, and DO concentration was as low as 0.32 mg/L at 60 m.

Longitudinal variation of hydrochemistry (T, Chl, DO, DOC) in surface water (0.5 m) and profile water (5–60 m) of the studied areas during winter (a), spring (b), summer (c), and autumn (d) in 2017. The light grey area corresponds to the reservoir area. Profile water was displayed as “Mean ± SD.” (DOC concentration was displayed in spring, summer, and autumn but not in winter due to a lack of data)
For the surface water of the mainstream, T varied from 13.1 to 29.0 °C and was higher in reservoirs (20.2 ± 4.3 °C) than the river samples (17.3 ± 3.1 °C). The obvious thermal stratification in summer with the main thermocline appeared at 5–15 m depth which can be observed in each reservoir (Fig. S1). The mean longitudinal DO was 9.5 ± 2.6 mg/L, while the maximum and minimum were both observed in WJD during spring (W14, 16.2 mg/L) and winter (W17, 4.6 mg/L), respectively. The concentration of Chl in the mainstream ranged from below detection limit (BDL) to 23.9 μg/L (mean = 1.8 ± 3.8 μg/L), which also varied greatly between those four reservoirs and had the largest value in WJD during April (mean = 14.2 ± 7.4 μg/L). Chl concentration in the outflow water was BDL while high Chl content was mainly observed in the surface water of the reservoir. The DOC concentration ranged from 0.6 to 2.4 mg/L, with a mean value 1.1 ± 0.4 mg/L. The mean DOC concentration during the three measured seasons for the reservoir samples was higher than that in the river samples, and there was a significant increase between the inflow of HJD and the outflow of WJD (~ 50%).
For tributaries, the annual mean values of T and Chl were 18.4 ± 3.6 °C and 1.7 ± 3.6 ug/L, which were consistent with the mainstream. However, the average DO value of tributaries (7.9 ± 2.0 mg/L) was smaller than that of the mainstream (9.4 ± 2.6 mg/L). Meanwhile, the mean DOC concentration in tributaries was 1.3 ± 0.5 mg/L, which was similar with the reservoir samples (1.1 ± 0.4 mg/L).
Longitudinal variation of NO3 − concentration and dual isotopes
Figure 3 shows the NO3−–N concentration in surface water ranged from 1.5 to 6.1 mg/L (mean = 3.4 ± 0.7 mg/L), while other dissolved nitrogenous species showed low concentration. NH4+–N, NO2−–N and DON were 0.1 ± 0.1, 0.07 ± 0.09, and 0.2 ± 0.3 mg/L, respectively. It suggested that NO3−–N was the dominant soluble nitrogen species, accounting for 90% of TDN on average. The seasonal pattern of NO3−–N showed more fluctuation and higher concentration in summer than other seasons. However, such an amplitude of elevating on NO3−–N concentration in summer between surface water in the reservoir and its outflow was spatially varied, and decreased from the upper to the middle reach of the cascade reservoir 28% (HJD), 17% (DF), and 10% (SFY) to 8% (WJD).

Longitudinal variation of nitrate concentrations and nitrate dual isotopes in surface water and profiles during winter (a), spring (b), summer (c), and autumn (d) in 2017. Profile water is displayed statistically as “mean ± SD.” The light grey area corresponds to the reservoir area
The δ15N-NO3− in surface water varied from 4.7 to 9.6‰, which generally showed a contrasting longitudinal trend to NO3−–N. The annual average δ18O-NO3− value is 1.3 ± 1.2‰ (− 1.2 to 3.6‰), whose longitudinal patterns varied during the different seasons with less fluctuation than δ15N-NO3−. δ18O-NO3− gradually increased along the river during spring when the highest δ18O-NO3− was observed, and slightly decreased in the other three seasons. The δ18O-NO3− values in reservoir profiles showed less fluctuation. Also, there was no significant difference (P > 0.05) between the mean values of δ15N-NO3− between the mainstream (7.4 ± 1.3‰) and tributaries (8.2 ± 1.9‰).
Temporal and spatial variation of NO3 −–N and dual isotopes in reservoirs
The nitrate characteristics of the profile in front of each reservoir dam in different seasons are shown in Fig. 4. The average NO3−–N concentrations in HJD, DF, SFY, and WJD reservoirs were 3.9 ± 0.8, 3.6 ± 0.6, 3.7 ± 0.4, and 3.5 ± 0.4 mg/L, respectively (Table S2). More specifically, the maximum NO3−–N concentration was 5.7 mg/L observed in HJD reservoir during summer, while 2 mg/L was the minimum appeared at 60 m in WJD reservoir in winter.

Variation of NO3−–N, δ15N-NO3−, and δ18O-NO3− profiles of W5 (HJD), W10 (DF), W12 (SFY), and W17 (WJD). The light grey-dashed lines divide the water profile into (a) epilimnion, (b) metalimnion, and (c) hypolimnion based on the T profile in summer (Fig. S1)
In contrast to the NO3− variation in profiles, there was no significant variation on the average of δ15N-NO3−, which was 7.4 ± 1.3, 6.8 ± 1.4, 7.0 ± 1.3, and 7.6 ± 1.5‰ in HJD, DF, SFY, and WJD, respectively. This was similar to δ18O-NO3− in HJD, DF, SFY, and WJD, corresponding to 1.5 ± 1.0, 1.6 ± 1.0, 1.3 ± 0.6, and 0.8 ± 1.2 ‰, respectively (Table. S2).
Overall, a wide range of NO3− and δ15N-NO3− was observed in summer, especially in HJD (3.1–5.8 mg/L, 5.4–9.0‰) and DF (1.5–4.8 mg/L, 4.5–7.5‰) at the depths of 0–15 m, while less variation was shown in SFY and WJD reservoirs. Meanwhile, it was observed that NO3− (2.0–3.6 mg/L) and its isotope (7.7–11.9‰) showed significant variation during winter in WJD especially at the 30–60-m depths, while no significant variation trend appeared in HJD, DF, and SFY during winter.
Discussion
Nitrogen biogeochemical processes in karst cascade reservoir system
The spatial and temporal variations of NO3−–N, δ15N-NO3−, and δ18O-NO3− profiles revealed that the studied reservoirs might undergo different NO3− fate (Fig. 4). The source of NO3−-N in the reservoir needed to be considered first, which corresponded to the seasonal variation of nitrate isotopes. The distribution of δ15N-NO3− and δ18O-NO3− values in the reservoir indicated that NO3− was relatively more influenced by soil nitrogen and urban sewage input during dry seasons (winter and spring), while the significant low mean δ15N (< 6.5‰) values indicated more significant influence of chemical fertilizer than other N source endmembers during flood seasons (summer and autumn) (Fig. S2).
In addition to the seasonal changes in nitrate sources, the internal reservoir also underwent different biochemical processes that affected NO3− concentration and dual isotopes (Akbarzadeh et al. 2019; Bardhan et al. 2017). To identify the vertical variation of nitrogen transformation in reservoirs, NO3−–N concentration and δ15N-NO3− at each sampling depth were compared with 5 m in the corresponding profile to calculate Δ δ15N-NO3− and Δ NO3−–N (Fig. 5), as the above analysis has proved that 5 m was the bottom of the epilimnion in the reservoir with similar dual nitrate isotopic values to the inflow river. Therefore, Δ[δ15N-NO3−] and Δ[NO3−–N] can reflect the influence amplitude by nitrogen transformation in reservoirs. Generally, the variation of Δ[δ15N-NO3−] versus Δ[NO3−–N] with a significant negative correlation could theoretically distinguish the biogeochemical processes, such as nitrification which cause Δ[δ15N-NO3−] to decrease and Δ[NO3−–N] to increase, while algal assimilation or denitrification cause Δ[δ15N-NO3−] to increase and Δ[NO3−–N] to decrease (Fenech et al. 2012). Otherwise, the strong mixing processes with high or low isotope signal values caused by dynamic disturbance or external input can also be reflected by no significant negative correlation between Δ[δ15N-NO3−] and Δ[NO3−–N] (Fig. 5).

Δ δ15N-NO3− (Δ δ15Ni= δ15Ni – δ15N5m, i means depth) versus Δ NO3−–N (Δ NO3−–Ni= [NO3−–N]i – [NO3−–N]5m,) of W5 (HJD), W10 (DF), W12 (SFY), and W17 (WJD) in sampling period. Different colors represent samples at different depths. E, epilimnion (0.5 m); M, metalimnion (15 m); H, hypolimnion (30, 45, 60 m). The light blue region is the 95% confidence interval of the linear fitting
The correlation between Δ[δ15N-NO3−] and Δ[NO3−–N] in Fig. 5 indicated that SFY received a significant mixing effect with significant negative correlation that only emerged in summer, while NO3− was significantly affected by the biogeochemical processes in the other three reservoirs. Given that the reservoir profile was aerobic or even oxygen-rich for most time, nitrification was likely to be the major process in the reservoir during four seasons in the other three reservoirs. During nitrification, 1/3 of the oxygen atoms in NO3− originated from atmospheric O2 and the remaining 2/3 originated from the water theoretically (Kendall et al. 2007). The δ18O of the studied area ranged from − 9.7 to − 6.8‰ (Wang et al. 2019a). Therefore, the calculated δ18O-NO3− derived from nitrification in the research area should vary between 1.4 and 3.3‰. The value of δ18O-NO3− of most samples (in spring and summer) lay within this calculated range (Fig. 3), which further confirmed that nitrification was the dominant transformation process. However, rest samples, particularly in winter and autumn, completely below this calculated range could reasonably be inferred from those various biogeochemical processes with the complex O exchange and mixing processes (Buchwald and Casciotti 2010).
Assimilation and denitrification are also the significant removal processes of NO3− in the lake/reservoir system (Swart et al. 2014). Previous research had reported that assimilation could enrich 15N and 18O of residual nitrate with a ratio of δ15N versus δ18O in 1:1 (Granger et al. 2010; Mayer and Wassenaar 2012). There was different extent for assimilation experienced in stratified periods (spring and summer), among which the HJD reservoir was the most severe and the WJD was moderate (Fig. 5). Specifically, our studies showed the ratio of δ15N and δ18O in surface to 15 m in HJD and DF during summer, and WJD during spring were 0.55, 0.65, and 0.72, respectively (Fig. S3).
Meanwhile, the absence of thermal stratification and uniformity in the profile in winter suggested a mixing process. Such environmental conditions were likely to facilitate organic matter mineralization and decomposition which had little impact on δ15N-NO3− variation (Bardhan et al. 2017; Yue et al. 2018). However, there was an anaerobic layer in the hypolimnion of W18 (the oldest reservoir, WJD) during winter, which made it possible for the occurrence of denitrification. Additionally, the ratio of δ15N and δ18O of W18 among 30 m to the bottom was 2.1, which further determined the denitrification (Fig. S3a). Meanwhile, with the 30 m set as initial value and the enrichment factor at − 6.99‰ (Fig. S3b), the denitrification contribution was calculated to be 18–28% in the bottom water using Rayleigh fractionation (Eq. (2)).
Factors controlling nitrogen fate in karst cascade reservoirs
Nitrogen fate is generally constrained by either biogeochemical processes or the process of sources mixing in river-reservoirs system (Nestler et al. 2011), which was reflected by nitrate dual isotopic composition and biogeochemical processes and were further influenced by the environmental characteristics (Finlay et al. 2013; Wenk et al. 2014). The above analysis showed that the dual isotope variation resulted from denitrification in the hypolimnion of WJD during winter, while in other reservoirs, dual isotope variation resulted from assimilation in the epilimnion during the other seasons. Therefore, Pearson correlation analysis was performed in those periods to explore the relationships between the ten parameters in reservoirs, including T, DO, Chl, TDS, pH, DOC, NO2−, NO3−, and dual isotopes (Fig. S4). The results showed that δ15N-NO3− was negatively correlated with NO3− and DO, while positively correlated with TDS and NO2− in winter. These correlations further confirmed DO as the decisive factor for denitrification, and the larger TDS might refer to the supply of more nutrients for denitrification.
The analysis results during other seasons differed from those during winter in WJD. δ15N-NO3− showed a significantly positive correlation with DO, Chl, and pH, with a negative correlation with NO3− and δ18O-NO3−. The higher DO, Chl, and pH referred to the existence of planktonic algae and plants with strong respiration, which strongly promoted assimilation in the epilimnion. Therefore, although the reservoir experienced different biogeochemical processes at various seasons and depths, DO was the key influencing factor for controlling nitrogen transformation in water. Meanwhile, planktonic algae and plants were also an important driving factor for nitrogen transformation in the upper layer of the deep reservoirs studied (Finlay et al. 2013; Glibert et al. 2016).
Due to the significant influence of artificial regulation, the biogeochemical processes of the main nutrients in the reservoir are also affected by factors including hydraulic retention time, storage capacity, and water depth (Schmutz and Moog 2018; Yang et al. 2020; Yu et al. 2014). Recent study further revealed that hydraulic load (ratio of average water depth/HRT) was the key factor for controlling the N2O emissions in the Wujiang cascade reservoirs and showed a negative relationship between N2O flux and hydraulic load in cascade reservoirs (Liang et al. 2019). In the present study, the HJD and WJD reservoirs with longer HRT had more obvious profile variations than SFY with shorter HRT (daily regulation mode) (Table. S1 and Fig. 4), which suggested that the longer HRT was more conducive to the nitrogen transformation process in reservoirs (Finlay et al. 2013).
More specifically, the DF and WJD reservoirs are both regulated monthly, but the variation of δ15N-NO3− in strong thermal stratification period in WJD presented a narrow range relative to DF (Fig. 6). It might be reasonably explained by the quite short monthly HRT (about 10 days) of WJD in summer, which was caused by the larger discharge flow of WJD with orders of magnitude higher than that of DF. The strong hydrodynamic condition in WJD during summer could buffer the variation of δ15N-NO3− inside the reservoir, which further revealed the influence of monthly HRT in each month on N-dynamic processes in the reservoir.

The seasonal variation of the content of δ15N-NO3− and monthly HRT (capacity in each month/discharge in each month) during the sampling period in DF and WJD. Mean monthly discharge is showed in Fig. S5
Additionally, it was found that reservoir running age determined the burial processes, degradation, and source of the organic matter (Barros et al. 2011; Evans et al. 2017). For example, the main source of organic matter was submerged soil and plants in the young reservoirs with an impounded age less than 15 years (e.g., HJD and SFY), while the continuous DOM transport from the inflow river and the endogenous organisms such as algae and plants in the water-level-fluctuating zone would play a more important role as the reservoir aged. This also further confirmed that the impounded age might be positively correlated with the trophic state level (Wang et al. 2018a). Thus, compared with the mesotrophic HJD, SFY, and mild-eutrophication DF, the WJD with eutrophication state was more conducive to the formation of anaerobic environment in the hypolimnion and water-sediment interface (Fig. 2), which might consequently promote the process of denitrification (Wang et al. 2018a).
Long-term variation of TDN in karst cascade reservoirs
The average annual TDN concentrations of the four reservoirs from 1998 to 2017 are shown in Fig. 7. It revealed that the TDN increased at the average rate of 0.1 mg/L-N per year from 2.0 mg/L in 1998 to 3.7 mg/L in 2015 and slightly decreased to 3.5 mg/L in 2017. Such a high content of TDN exceeded the Criterion V of Chinese Environmental Quality Standards for Surface Water (2 mg/L as TDN, Fig. 7) and was much higher than other tributaries and the mainstream in the upper basin of the Yangtze River (Ding et al. 2019; Li et al. 2010). Furthermore, TDN in cascade reservoirs of the upper and middle reaches of the Wujiang River are higher than the other karstic cascade reservoirs densely affected by human activity from agriculture and industry (Maotiao River < 2 mg/L) (Wang et al. 2018b).

TDN concentration of surface water in four reservoirs from 1998 to 2017. The error bars represent the minimum and maximum values. Data was collected in 1988 (Su 2002); 2003–2004 (Zhu 2005); 2007–2008 (Liu et al. 20112017); available summer data in 2014–2015 (Yang et al. 2018); and 2017 during the present study. The blue dot plot data representing the amount of N-fertilizer used in Guizhou Province was collected from http://www.stats.gov.cn/tjsj/ndsj/
Dual isotopes over four seasons showed agricultural fertilizer made more of a contribution in summer and autumn with high runoff flow (Fig. S2), indicating the agricultural effect on water quality (Li et al. 2020a, b; Yue et al. 2020). It showed that the long-term TDN concentration in cascade reservoirs was highly dependent on the long-term N fertilizer usage in Guizhou Province (P < 0.05), which both reached maximum application during 2014 and 2015 (Fig. 7). However, the different increase rate between TDN concentration (1.8-folds in WJD) and N fertilizers (1.3-folds) during the past two decades further indicated that other factors should also be considered from other sources. For example, trapped terrestrial input in cascade reservoirs resulted in new N sources owing to prolonged transport time (Maavara et al. 2020; Van Cappellen and Maavara 2016). Nitrogen biogeochemistry in karst cascade reservoirs may also contribute to an increase of N concentration, e.g., novel production from particle matter, phytoplankton, and even from sediment (Bardhan et al. 2017; Wang et al. 2018b).
Moreover, the studied reservoirs are all deep (> 30 m) with huge elevation drops which provided sufficient hydrodynamic force to deepen the aerobic layer (Qin et al. 2020), which can promote nitrification and reduce the contribution of denitrification simultaneously, resulting in less effectiveness in the removal of NO3− than those in shallow lakes or reservoirs (Liu et al. 2018; Wang et al. 2019a). Overall, the sufficient input of N sources (especially the chemical fertilizer), convenient hydrologic transmission channels, and favorable conditions influenced by strong nitrification and weak nitrogen removal jointly contributed to the high TDN concentration in the present deep cascade reservoirs in karst areas. Additionally, with the previous study showing that phosphate in cascade reservoirs could be retained more efficiently and replenished faster than N (Maavara et al. 2020; Qin et al. 2020). However, the large reserve of high-quality phosphate in the study area (Yang et al. 2018) and potential mining may lead to phosphate transportation to aquatic systems, which may further be a high risk to the aquatic health as high TDN waters in the study area.
Conclusion
Nowadays, the environmental implications and nutrient biogeochemistry in the impounded cascade reservoirs system have drawn more and more attention. The fate of N in four cascade reservoirs located in the upper and middle reaches of the Wujiang River, a karst watershed, was studied. The huge elevation drop provided sufficient hydrodynamic force to facilitate the aerobic layer, resulting in an aerobic aquatic environment for almost all of the reservoirs, except for the last profile of WJD in winter. Such an aerobic aquatic environment facilitated the nitrification process to produce nitrate as the primary species. Nitrate dual isotopes in these four cascade reservoirs showed little spatial variation but more obvious seasonal variation in summer under high discharge, indicating that chemical fertilizer input by agricultural activity was the major contributor. The nitrate dual isotopic signature and high DO concentration demonstrated a weak depletion process, e.g., denitrification, which explained the elevated nitrate concentration in the past two decades. Additionally, the frequent exchange of pollutants between the dual karstic hydrological pathways and the increase of N fertilizer application explained the elevated nitrate concentration in the past two decades.
Data Availability
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Akbarzadeh Z, Maavara T, Slowinski S, Van Cappellen P (2019) Effects of damming on river nitrogen fluxes: a global analysis. Glob Biogeochem Cycles 33:1339–1357
Bardhan P, Naqvi SWA, Karapurkar SG, Shenoy DM, Kurian S, Naik H (2017) Isotopic composition of nitrate and particulate organic matter in a pristine dam reservoir of western India: implications for biogeochemical processes. Biogeosciences 14:767–779
Barros N, Cole JJ, Tranvik LJ, Prairie YT, Bastviken D, Huszar VLM, del Giorgio P, Roland F (2011) Carbon emission from hydroelectric reservoirs linked to reservoir age and latitude. Nat Geosci 4:593–596
Buchwald C, Casciotti KL (2010) Oxygen isotopic fractionation and exchange during bacterial nitrite oxidation. Limnol Oceanogr 55:1064–1074
Chen A, Wu M, McClain ME (2019) Classifying dams for environmental flow implementation in China. Sustainability-Basel 12(1):107
Chen Q, Shi W, Huisman J, Maberly SC (2020) Hydropower reservoirs on the upper Mekong River modify nutrient bioavailability downstream. Natl Sci Rev nwaa026:1–9
Cheng F, Zhang HM, Zhang GL, Liu SM, Song GD, Du GX (2019) Distribution and emission of N2O in the largest river-reservoir system along the Yellow River. Sci Total Environ 666:1209–1219
Davraz A, Karaguzel R, Soyaslan I, Sener E, Seyman F, Sener S (2009) Hydrogeology of karst aquifer systems in SW Turkey and an assessment of water quality and contamination problems. Environ Geol 58:973–988
Ding S, Chen P, Liu S, Zhang G, Zhang J, Dan SF (2019) Nutrient dynamics in the Changjiang and retention effect in the Three Gorges Reservoir. J Hydrol 574:96–109
Evans CD, Futter MN, Moldan F, Valinia S, Frogbrook Z, Kothawala DN (2017) Variability in organic carbon reactivity across lake residence time and trophic gradients. Nat Geosci 10:832–835
Fadhullah W, Yaccob NS, Syakir MI, Muhammad SA, Yue FJ, Li SL (2020) Nitrate sources and processes in the surface water of a tropical reservoir by stable isotopes and mixing model. Sci Total Environ 700:134517
Fenech C, Rock L, Nolan K, Tobin J, Morrissey A (2012) The potential for a suite of isotope and chemical markers to differentiate sources of nitrate contamination: a review. Water Res 46:2023–2041
Fenton O, Mellander PE, Daly K, Wall DP, Jahangir MMR, Jordan P, Hennessey D, Huebsch M, Blum P, Vero S, Richard KG (2017) Integrated assessment of agricultural nutrient pressures and legacies in karst landscapes. Agric Ecosyst Environ 239:246–256
Finlay JC, Small GE, Sterner RW (2013) Human influences on nitrogen removal in lakes. Science 342:247–250
Gao Y, Jia YL, Yu GR, He NP, Zhang L, Zhu B, Wang YF (2019) Anthropogenic reactive nitrogen deposition and associated nutrient limitation effect on gross primary productivity in inland water of China. J Clean Prod 208:530–540
Glibert PM, Wilkerson FP, Dugdale RC, Raven JA, Dupont CL, Leavitt PR, Parker AE, Burkholder JM, Kana TM (2016) Pluses and minuses of ammonium and nitrate uptake and assimilation by phytoplankton and implications for productivity and community composition, with emphasis on nitrogen-enriched conditions. Limnol Oceanogr 61:165–197
Granger J, Sigman DM, Rohde MM, Maldonado MT, Tortell PD (2010) N and O isotope effects during nitrate assimilation by unicellular prokaryotic and eukaryotic plankton cultures. Geochim Cosmochim Acta 74:1030–1040
Green SM, Dungait JA, Tu C, Buss HL, Sanderson N, Hawkes SJ, Xing K, Yue F, Hussey VL, Peng J et al (2019) Soil functions and ecosystem services research in the Chinese karst Critical Zone. Chem Geol 527:119107
Grill G, Lehner B, Thieme M, Geenen B, Tickner D, Antonelli F, Babu S, Borrelli P, Cheng L, Crochetiere H, Ehalt Macedo H, Filgueiras R, Goichot M, Higgins J, Hogan Z, Lip B, McClain ME, Meng J, Mulligan M, Nilsson C, Olden JD, Opperman JJ, Petry P, Reidy Liermann C, Sáenz L, Salinas-Rodríguez S, Schelle P, Schmitt RJP, Snider J, Tan F, Tockner K, Valdujo PH, van Soesbergen A, Zarfl C (2019) Mapping the world’s free-flowing rivers. Nature 569:215–221
Harrison JA, Maranger RJ, Alexander RB, Giblin AE, Jacinthe P-A, Mayorga E, Seitzinger SP, Sobota DJ, Wollheim WM (2009) The regional and global significance of nitrogen removal in lakes and reservoirs. Biogeochemistry 93:143–157
Huang SY, Pu JB, Li JH, Zhang T, Cao JH, Pan MC (2020) Sources, variations, and flux of settling particulate organic matter in a subtropical karst reservoir in Southwest China. J Hydrol 586:124882
Jin ZF, Cen JR, Hu YM, Li LJ, Shi YS, Fu GW, Li FL (2019) Quantifying nitrate sources in a large reservoir for drinking water by using stable isotopes and a Bayesian isotope mixing model. Environ Sci Pollut Res 26:20364–20376
Kendall C, Elliott EM, Wankel SD (2007) Tracing anthropogenic inputs of nitrogen to ecosystems. In: Lajtha K, Michener R (eds) Stable Isotopes in Ecology and Environmental Science, 2nd edn. Blackwell, UK, pp 375–449
Li C, Ji H (2016) Chemical weathering and the role of sulfuric and nitric acids in carbonate weathering: isotopes (13C,15N,34S, and18O) and chemical constraints. J Geophys Res Biogeosci 121:1288–1305
Li SL, Liu CQ, Li J, Liu XL, Chetelat B, Wang BL, Wang FS (2010) Assessment of the sources of nitrate in the Changjiang River, China using a nitrogen and oxygen isotopic approach. Environ Sci Technol 44:1573–1578
Li C, Li SL, Yue FJ, Liu J, Zhong J, Yan ZF, Zhang RC, Wang ZJ, Xu S (2019) Identification of sources and transformations of nitrate in the Xijiang River using nitrate isotopes and Bayesian model. Sci Total Environ 646:801–810
Li S-L, Xu S, Wang T-J, Yue F-J, Peng T, Zhong J, Wang L-C, Chen J-A, Wang S-J, Chen X, Liu C-Q (2020a) Effects of agricultural activities coupled with karst structures on riverine biogeochemical cycles and environmental quality in the karst region. Agric Ecosyst Environ 303:107120
Li Y, Abegunrin TP, Guo H, Huang Z, Are KS, Wang H, Gu M, Wei L (2020b) Variation of dissolved nutrient exports by surface runoff from sugarcane watershed is controlled by fertilizer application and ground cover. Agric Ecosyst Environ 303:107121
Li Z, Xu X, Zhu J, Xu C, Wang K (2020c) The contributions of the largest erosive events to sediment yields in karst catchments. Water Resour Res 56. https://doi.org/10.1029/2019WR025839
Liang X, Xing T, Li J, Wang BL, Wang F, He C, Hou LJ, Li SL (2019) Control of the hydraulic load on nitrous oxide emissions from cascade reservoirs. Environ Sci Technol 53:11745–11754
Liu C (2007) Bio–geochemical processes and cycling of nutrients in the earth’s surface: erosion of karstic catchment and nutrients cycling in Southwest China. Science Press, Beijing
Liu XL, Liu CQ, Li SL, Wang FS, Wang BL, Wang ZL (2011) Spatiotemporal variations of nitrous oxide (N2O) emissions from two reservoirs in SW China. Atmos Environ 45:5458–5468
Liu X, Li S, Wang Z, Han G, Li J, Wang B, Wang F, Bai L (2017) Nitrous oxide (N2O) emissions from a mesotrophic reservoir on the Wujiang River, southwest China. Acta Geochim 36:667–679
Liu SS, Wu FC, Feng WY, Guo WJ, Song FH, Wang H, Wang Y, He ZQ, Giesy JP, Zhu P, Tang Z (2018) Using dual isotopes and a Bayesian isotope mixing model to evaluate sources of nitrate of Tai Lake, China. Environ Sci Pollut Res 25:32631–32639
Maavara T, Chen Q, Van Meter K, Brown LE, Zhang J, Ni J, Zarfl C (2020) River dam impacts on biogeochemical cycling. Nat Rev Earth Environ 1:103–116
Mayer B, Wassenaar LI (2012) Isotopic characterization of nitrate sources and transformations in Lake Winnipeg and its contributing rivers, Manitoba, Canada. Journal of Great Lakes Research 38. Supplement 3:135–146
McIlvin MR, Casciotti KL (2011) Technical updates to the bacterial method for nitrate isotopic analyses. Anal Chem 83:1850–1856
Mulligan M, van Soesbergen A, Saenz L (2020) GOODD, a global dataset of more than 38,000 georeferenced dams. Sci Data 7:31
Nestler A, Berglund M, Accoe F, Duta S, Xue DM, Boeckx P, Taylor P (2011) Isotopes for improved management of nitrate pollution in aqueous resources: review of surface water field studies. Environ Sci Pollut Res 18:519–533
Qin B, Zhou J, Elser JJ, Gardner WS, Deng J, Brookes JD (2020) Water depth underpins the relative roles and fates of nitrogen and phosphorus in lakes. Environ Sci Technol 54:3191–3198
Ran XB, Bouwman L, Yu ZG, Beusen A, Chen HT, Yao QZ (2017) Nitrogen transport, transformation, and retention in the Three Gorges Reservoir: a mass balance approach. Limnol Oceanogr 62:2323–2337
Ren J, Zhao M, Zhang W, Xu Q, Yuan J, Dong B (2020) Impact of the construction of cascade reservoirs on suspended sediment peak transport variation during flood events in the Three Gorges Reservoir. Catena 188:104409
Schmutz S, Moog O (2018) Dams: ecological impacts and management, Riverine Ecosystem Management. In: Schmutz S., Sendzimir J. (eds) Riverine Ecosystem Management. Aquatic Ecology Series, vol 8. Springer, Cham, pp 111–127
Shi W, Qiuwen C, Jianyun Z, Dongsheng L, Qitao Y, Yuchen C, Honghai M, Liuming H (2020) Nitrous oxide emissions from cascade hydropower reservoirs in the upper Mekong River. Water Res 173:115582
Su WC (2002) Negative effects of cascade hydropower exploitation environmental in the Wujiang Basin. Resour Environ Yangtze Basin 11:388–392 (In Chinese with English Abstract)
Sullivan PL, Macpherson GL, Martin JB, Price RM (2019) Evolution of carbonate and karst critical zones. Chem Geol 527:119223
Swart PK, Evans S, Capo T, Altabet MA (2014) The fractionation of nitrogen and oxygen isotopes in macroalgae during the assimilation of nitrate. Biogeosciences 11:6147–6157
Van Cappellen P, Maavara T (2016) Rivers in the Anthropocene: global scale modifications of riverine nutrient fluxes by damming. Ecohydrol Hydrobiol 16:106–111
Wang FS, Yu YX, Liu CQ, Wang BL, Wang YC, Guan J, Mei HY (2010) Dissolved silicate retention and transport in cascade reservoirs in Karst area, Southwest China. Sci Total Environ 408:1667–1675
Wang B, Qiu X-L, Peng X, Wang F (2018a) Phytoplankton community structure and succession in karst cascade reservoirs, SW China. Inland Waters 8:229–238
Wang ZJ, Yue FJ, Li SL, Li XD, Wang SL, Li C, Tao FX (2018b) Nitrate dynamics during impoundment and flood periods in a subtropical karst reservoir: Hongfeng Lake, Southwestern China. Environ Sci Process Impacts 20:1736–1745
Wang KL, Zhang CH, Chen HS, Yue YM, Zhang W, Zhang MY, Qi XK, Fu ZY (2019) Karst landscapes of China: patterns, ecosystem processes and services. Landsc Ecol 34(12):2743–2763
Wang BL, Zhang HT, Liang X, Li XD, Wang FS (2019a) Cumulative effects of cascade dams on river water cycle: evidence from hydrogen and oxygen isotopes. J Hydrol 568:604–610
Wang W, Li S, Zhong J, Maberly SC, Li C, Wang FS, Xiao HY, Liu CQ (2019b) Climatic and anthropogenic regulation of carbon transport and transformation in a karst river-reservoir system. Sci Total Environ 707:135628
Wenk CB, Zopfi J, Blees J, Veronesi M, Niemann H, Lehmann MF (2014) Community N and O isotope fractionation by sulfide-dependent denitrification and anammox in a stratified lacustrine water column. Geochim Cosmochim Acta 125:551–563
Winton RS, Calamita E, Wehrli B (2019) Reviews and syntheses: dams, water quality and tropical reservoir stratification. Biogeosciences 16:1657–1671
Xing M, Liu WG (2016) Using dual isotopes to identify sources and transformations of nitrogen in water catchments with different land uses, Loess Plateau of China. Environ Sci Pollut Res 23:388–401
Xue D, Botte J, De Baets B, Accoe F, Nestler A, Taylor P, Van Cleemput O, Berglund M, Boeckx P (2009) Present limitations and future prospects of stable isotope methods for nitrate source identification in surface- and groundwater. Water Res 43:1159–1170
Yang D, Wang S, Lu W, Xiang P, Yang Y, Tan D, Guo M, Yeager KM (2018) Impoundment-induced nitrogen–phosphorus imbalance in cascade reservoirs alleviated by input of anthropogenic nutrients. Inland Waters 8:196–206
Yang M, Li X-D, Huang J, Ding S, Cui G, Liu C-Q, Li Q, Lv H, Yi Y (2020) Damming effects on river sulfur cycle in karst area: a case study of the Wujiang cascade reservoirs. Agric Ecosyst Environ 294:106857
Yu Z, Yang J, Amalfitano S, Yu X, Liu L (2014) Effects of water stratification and mixing on microbial community structure in a subtropical deep reservoir. Sci Rep-Uk 4:5821
Yue F-J, Li S-L, Liu C-Q, Mostofa KMG, Yoshida N, Toyoda S, Wang S-L, Hattori S, Liu X-L (2018) Spatial variation of nitrogen cycling in a subtropical stratified impoundment in southwest China, elucidated by nitrous oxide isotopomer and nitrate isotopes. Inland Waters 8:186–195
Yue F-J, Waldron S, Li S-L, Wang Z-J, Zeng J, Xu S, Zhang Z-C, Oliver DM (2019) Land use interacts with changes in catchment hydrology to generate chronic nitrate pollution in karst waters and strong seasonality in excess nitrate export. Sci Total Environ 696:134062
Yue F-J, Li S-L, Waldron S, Wang Z-J, Oliver DM, Chen X, Liu C-Q (2020) Rainfall and conduit drainage combine to accelerate nitrate loss from a karst agroecosystem: insights from stable isotope tracing and high-frequency nitrate sensing. Water Res 186:116388
Zarfl C, Lumsdon AE, Berlekamp J, Tydecks L, Tockner K (2015) A global boom in hydropower dam construction. Aquat Sci 77:161–170
Zhang HY, Yang YS, Zou JY, Wen YJ, Gao CP (2018) The sources and dispersal of nitrate in multiple waters, constrained by multiple isotopes, in the Wudalianchi region, northeast China. Environ Sci Pollut Res 25:24348–24361
Zhao HJ, Xiao Q, Miao Y, Wang ZJ, Wang QG (2020) Sources and transformations of nitrate constrained by nitrate isotopes and Bayesian model in karst surface water, Guilin, Southwest China. Environ Sci Pollut Res 27:21299–21310
Zhou X, Chen N, Yan Z, Duan S (2016) Warming increases nutrient mobilization and gaseous nitrogen removal from sediments across cascade reservoirs. Environ Pollut 219:490–500
Zhu J (2005) Effect of dams on the biogeochemical cycles of nutrients in the Wujiang River. Institute of Geochemistry, Chinese Academy of Sciences, Guiyang (China) (In Chinese with English Abstract)
Funding
This research was funded by the National Key Research and Developmental Program of China (grant no. 2016YFA0601002) and National Natural Science Foundation of China (grant nos. 41672351, 42073076, 419250020).
Author information
Authors and Affiliations
Contributions
Sai-Nan Chen: investigation, data curation, formal analysis, writing original draft. Fu-Jun Yue: conceptualization, supervision, writing, review, and editing. Xiao-Long Liu: formal analysis, methodology, investigation, writing, review, and editing. Jun Zhong: data curation, writing, and review. Yuan-Bi Yi and Yulin Qi: investigation, review, and editing. Wan-Fa Wang: data curation and investigation. Si-Liang Li and Hua-Yun Xiao: conceptualization, supervision, writing, review, and editing.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Responsible editor: Philippe Garrigues
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
ESM 1
(DOCX 775 kb)
Rights and permissions
About this article

Cite this article
Chen, SN., Yue, FJ., Liu, XL. et al. Seasonal variation of nitrogen biogeochemical processes constrained by nitrate dual isotopes in cascade reservoirs, Southwestern China. Environ Sci Pollut Res 28, 26617–26627 (2021). https://doi.org/10.1007/s11356-021-12505-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-021-12505-9