
Abstract
Investigation of the water sources used by trees of different ages is essential to formulate a conservation strategy for the riparian tree, P. euphratica. This study addressed the contributions of different potential water sources to P. euphratica based on levels of stable oxygen and hydrogen isotopes (δ18O, δ2H) in the xylem of different aged P. euphratica, as well as in soil water and groundwater along the lower Heihe River. We found significant differences in δ18O values in the xylem of different aged P. euphratica. Specifically, the δ18O values of young, mature and over-mature forests were −5.368(±0.252) ‰, −6.033(± 0.185) ‰ and −6.924 (± 0.166) ‰, respectively, reflecting the reliance of older trees on deeper sources of water with a δ18O value closer to that of groundwater. Different aged P. euphratica used different water sources, with young forests rarely using groundwater (mean <15 %) and instead primarily relying on soil water from a depth of 0–50 cm (mean >45 %), and mature and over-mature forests using water from deeper than 100 cm derived primarily from groundwater.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
P. euphratica (Euphrates poplar) is widely distributed in inland river basins of arid zones in northwest China. This tree can tolerate salinity, drought and wind and has important ecological benefits (Chen et al. 2004). The Food and Agriculture Organization has identified P. euphratica as an urgent priority for protection of forests genetic resources; however, a detailed understanding of their water source is essential to their conservation.
Plant water sources can be determined through stable isotope analysis of stem water (Dawson et al. 2002; Asbjornsen et al. 2007; Hasselquist et al. 2010). There is usually no isotopic fractionation during water uptake by terrestrial plants, although examples of plant species that seem to fractionate hydrogen (but not oxygen) during water uptake do exist (Ellsworth and Williams 2007). Water sources can be pinpointed by comparing the isotope ratios of all potential sources with those of water extracted from plant stems (Dawson et al. 2002).
Water sources differ greatly between and within species. Sources used by an individual plant may vary greatly over time, especially in arid regions (Nie et al. 2012). Plant roots interact with soil texture, water availability and salinity to determine which sources are most easily used by the plant (Overdieck et al. 2013). In species capable of growing to a large size, root distribution may vary greatly with plant age (Dawson 1996).
Because P. euphratica is the foundation species of inland riparian forests, its ecological characteristics, stress resistance and water use have been the focus of much research (Chen et al. 2005; Li et al. 2008; Chen et al. 2011; Yang et al. 2012). Studies using the stable isotopes 2H and 18O have shown that sand dune plants in the middle reaches of the Heihe River Basin occasionally use rainwater, but primarily rely on the more stably available phreatic water (Zhou et al. 2011;. Yu et al. 2012). Studies in the lower Heihe River Basin have shown that the growth of woody plants mainly depends on groundwater, while herbs rely on surficial soil water (Zhao et al. 2008). Along the lower reaches of the Tarim River, groundwater is the main water source for survival of P. euphratica (Chen et al. 2006). In this study, we addressed water sources used by different aged P. euphratica in the lower reaches of the Heihe River Basin. We used stable isotopes of 2H and 18O and analysis of soil moisture to assess the contributions of rainfall and groundwater for young, mature and over-mature forests. Our goal was to provide a scientific basis for ecological water configuration measures designed to protect P. euphratica in the lower Heihe River Basin.
Data and methods
Study area
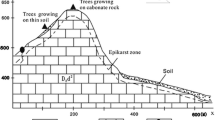
Our study area is located in the lower reaches of the Heihe River, within Ejin Banner, Inner Mongolia, China (Fig. 1). The region is characterized by a dry, continental climate. The average annual precipitation from 1960 to 2010 was 42 mm, with maximum and minimum values of 103 mm and 7 mm, while the mean annual potential evapotranspiration (PET) was 3755 mm, with a maximum of 4035 mm. Therefore, the potential evapotranspiration was 89 times the rainfall. The mean annual temperature was 8.2 °C. The prevailing northwest winds have an average speed of 4.2 m/s, and a maximum of 24.0 m/s. The riparian vegetation is dominated by P. euphratica, as well as the shrubs Tamarix ramosissima and Sophora alopecuroides.

Map of the study area
Sample collection
The study site is located on the flood plain of the Heihe River near Ulan Tug (Fig. 1). There has been no inundation for many years, and there is no significant ground-surface slope. One representative plot was selected within each of three age classifications of P. euphratica: young forests, mature forests and over-mature forests. Three individuals within each plot that met the following requirements were selected: good growth, straight trunk, moderate crown, and no plant diseases or insect pests. A plant stem sample was taken from each selected individual, and a nearby pit was augured to the water table for soil moisture determination. All plots were sampled during August 2012. No rain or water condensation occurred at the study area within a few days before or after sampling. Age group classifications were made according to the standard practice in the forestry sector. Plot characteristics are shown in Table 1, and groundwater parameters are shown in Table 2.
Determination of soil moisture
Soil moisture was measured in vertical layers, with three samples per layer. Layers were 10 cm thick between 0 and 40 cm and 20 cm thick at depths greater than 40 cm. Measurements extended down to the water table, 260 cm in over-mature forests and 320 cm in young and mature forests. Each sample was immediately weighed in the field and then reweighed in the lab after oven-drying at 105 °C to a constant mass.
Xylem and soil water isotope sampling
Stems selected for xylem sampling were more than 2 years old with a diameter of 0.3–0.5 cm and a length of 3–5 cm. The bark, phloem and cambium were removed to expose the xylem, which was quickly loaded into glass bottles, covered with a cork, sealed with parafilm, and placed on ice in a cooler for transport back to the lab. Soil water samples were collected for isotope analysis following the protocol for soil moisture. Groundwater samples were taken from groundwater observation wells.
Laboratory methods
Previous studies have demonstrated that there is no isotope fractionation in plants during water transport from roots to leaves through stems, except in salt-excluding plants in coastal wetlands (Dawson et al. 1991; Zhao et al. 2008). Therefore, as long as the stable isotope compositions of the potential water sources are significantly different, δ2H and δ18O values of xylem water and each water source can be used to determine the relative contribution of each source (Dawson et al. 1991; Busch et al. 1992; Dawson et al. 1993; Phillips et al. 2001; Zhang et al. 2004; Asbjornsen et al. 2007).
In this study, we compared the δ2H and δ18O from xylem water and soil water with the goal of determining from where in the soil profile P. euphratica obtains its water. Values of δ18O and δ2H in soil water decreased monotonically with increasing depth, reflecting the change in water source from precipitation to groundwater; therefore, we used values of δ18O and δ2H to calculate the relative contributions of soil water and groundwater for any age and depth.
Water in the soil and plant stem samples was extracted cryogenically using the approach described by Ehleringer et al. (2000). An LGR liquid water isotope analyzer was used to measure hydrogen and oxygen isotope ratios of groundwater, xylem water and soil water in each layer.
δ2H and δ18O represent hydrogen and oxygen isotope values of samples, while Rsample and Rstandard are the stable isotopic compositions (i.e. the 2H/H molar ratio) of the sample and the standard water (SMOW; Ehleringer et al. 2000). The accuracy (1δ) was 0.1 ‰ for 18O/16O and 0.3 ‰ for 2H/H. The test error was less than 1 ‰ for δ 2H and less than 0.2 ‰ for δ18O.
The IsoSource mixing model (http://www.epa.gov/wed/pages/models/stableIsotopes/isosource/isosource.htm) (Phillips et al. 2003) used stable isotope values to determine the relative contributions of soil water and groundwater as a function of depth. The user supplies the source increment (e.g., entering a value of 1 % specifies examination of all possible combinations of sources contributions from 0 to 100 % in increments of 1 %), and the mass balance tolerance (e.g., entering a value of 0.1 l specifies that all source combinations that result in predicted mixture signatures within 0.1 l of the observed signature are considered feasible solutions). IsoSource provides output files that list each feasible solution, descriptive statistics about the distribution of these solutions (number of solutions, mean, standard deviation, minimum, maximum, 1st percentile, median, and 99th percentile for each source), and histograms of these distributions. Statistical analyses were performed with SPSS17.0 and plotted with origin 8.0.
Results
Soil moisture
Soil moisture profiles varied with age of P. euphratica (Fig. 2). Overall, soil moisture was higher in young forests than mature or over-mature forests. At a finer scale, the soil moisture profile could be divided into 3 zones, 0–100 cm, 100–200 cm and 200 cm to the saturated zone. At depths of 0–100 cm, the soil moisture ranges were 3.86–18.89 % in young forests (Fig. 2a), 0.64–2.08 % in mature forests (Fig. 2b) and 2.09–4.15 % in over-mature forests (Fig. 2c).

Soil water profiles in riparian Populus euphratica forests patches of different ages along the lower Heihe River; a is young forests, b is mature forests and c is over-mature forests
All forests age groups displayed a peak in soil moisture between 100 and 200 cm. For young forests, a peak of 17.35 % occurred at 120–140 cm, while for mature forests, a peak of 7.08 % occurred at 180–200 cm and for over-mature forests a peak of 6.34 % occurred at 100–120 cm. Soil texture profiles did not differ among age groups; therefore, peaks in soil moisture may be related to the distribution of P. euphratica roots. In other words, soil moisture may be more abundant in young forests than mature or over-mature forests because young trees consume less water (Li et al. 2008). As expected soil moisture increased strongly near the water table (Fig. 2), with peak values for young, mature and over-mature forests of 31.8, 24.6 and 28.6 %, respectively. The P. euphratica root distribution was more shallow in young forests (20–40 cm) than in mature forests (60–120 cm) or over-mature forests (60–160 cm) (Table 1).
Water absorption as a function of age and depth
For all three age classifications, the shallowest soil water sample, 5 cm in depth, had the highest or near highest measured value of δ18O (4.4, 4.1 and 1.5 ‰ for young forests, mature forests and over-mature forests, respectively). Values of δ18O decreased with increasing depth, with the greatest rates of decrease occurring near the surface, especially in young forests, reflecting evaporation of soil water near the surface. In young forests, δ18O values decreased rapidly from 4.6 ‰ to −3.4 ‰ from 0 to 30 cm. In contrast, from 60 cm to 320 cm, no trends in δ18O values were observed, forests with all being close to the groundwater δ18O value of −6.9 ‰. The decrease in δ18O values in mature forests occurred at a greater depth than in young forests. Values of δ18O fluctuated around 4 at depths of 0–60 cm, then dropped to −3.6 ‰ at 120 cm. At depths of 120–200 cm, the δ18O values fluctuated around −5 ‰, while at 200–320 cm they were between −7.1 and −7.4 ‰, which was close to the groundwater δ18O value (−6.9 ‰). Over-mature forests showed the same pattern of decreasing values of δ18O with increasing depth, but the gradient was not as strong. The value near the surface was 1.5 ‰, while it decreased to −6.1 ‰ at 100 cm. From 120 to 260 cm, δ18O values fluctuated around the value for groundwater. The similarity between deep soil and groundwater values of δ18O for all forests ages strongly suggests that the deep soil water is derived from groundwater.
Because xylem values of δ18O should equal the volume-weighted average of all the sources used by the tree, comparison of soil and xylem values can suggest from where roots are deriving their water. In young forests, soil water and xylem water values of δ18O were most similar at depths of 30–50 cm, suggesting that young forests is deriving water from shallow layers (Fig. 3a). In mature forests, δ18O values of soil and xylem water were most similar at depths of 120–200 cm (Fig. 3c), suggesting that mature forests uses deeper water than young forests. In over-mature forests, δ18O values of soil and xylem water were similar from 100 to 250 (Fig. 3e) cm, suggesting that over-mature forests is also accessing deep water. This interpretation is supported by comparison of δ18O values in xylem and groundwater. In young forests, xylem values were considerably higher than those of groundwater (Fig. 3a), suggesting that the trees are using considerable amounts of water from near the ground surface. In mature forests, xylem values were only slightly higher than groundwater values (Fig. 3c), suggesting that trees are primarily using water from deep in the soil, where soil and groundwater δ18O values are similar. In over-mature forests, xylem and groundwater δ18O values were essentially the same (Fig. 3e), indicating that these trees rely entirely on deep water from near the water table.

Comparison of δ 18O and δ2H profiles of soil water, xylem water and ground water in Populus euphratica forests of different age. Figures a and b are young forests, c and d are mature forests and e and f are over-mature forests
The patterns of δ2H in xylem water were most similar to those of δ18O, but the decrease in δ2H with depth was not as strong as that in δ18O. In young forests, soil water δ2H decreased from 0 to 150 cm, increased from 150 to 250 cm and then slightly decreased at greater depths (Fig. 3b). In mature forests, δ2H values of soil water increased from 0 to 50 cm, decreased from 50 to 125 cm, and showed no consistent trend at greater depths (Fig. 3d). In over-mature forests, δ2H decreased from 0 to 70 cm, but then increased from 170 to 200 cm (Fig. 3f). Because of the relatively weak trend of decreasing δ2H with depth, δ2H was less useful than δ18O for determining where in the soil profile P. euphratica obtained its water.
Modeled contributions by different water sources
We used a mixing model to estimate contributions to xylem water made by different water sources, first considering only δ18O and then considering both δ18O and δ2H. When only δ18O was considered, young forests primarily used (40–48 %) soil water from 0 to 50 cm (Table 3), while mature forests used little water from depths of 0–50 cm and 50–100 cm (0–10 % and 0–13 %), and instead derived most of its water from deeper layers and groundwater. Over-mature forests primarily used soil water from 100 to 200 cm, 200 to 260 cm and groundwater (0–92, 0–93 and 0–94 %, respectively), while it used little water from 0 to 50 cm and 50 to 100 cm (0–4 % and 0–18 %, respectively).
Similar results were observed upon consideration of both the δ18O and δ2H values (Table 3). Specifically, young forests primarily used (45–48 %) soil water from 0 to 50 cm depth, while it used little from deeper layers. Mature forests used mostly (84–88 %) groundwater, and little soil water from shallower depths. Over-mature forests used mostly (35–54 %) soil water from 100 to 200 cm.
Discussion
All ages of P. euphratica along the Heihe River exploit soil layers with the most available water. For all ages of forests, we found a correspondence between the depths of high moisture availability, the depths from which the trees derived most of their water, and the depths with the highest concentration of roots (Brunel et al. 1995). Stable isotopes of oxygen and hydrogen indicated that only young forests derived a large percentage of its water (45–48 %) from near the soil surface (0–50 cm; Table 3). Young forests was also the only age group with significant moisture near the surface (Fig. 2), and the only age group with roots concentrated near the surface (20–40 cm; Table 1). In mature and over-mature forests, stable isotopes indicated that water was primarily derived from deep soil layers and groundwater (Table 3), soil moisture was highest in deeper layers (Fig. 2) and roots were concentrated in deeper layers (Table 1). In contrast, Liu et al. (2012) found that soil water content decreased in layers containing the most absorbing roots of Caragana intermedia in Alpine sandland.
Transpiration, evaporation and soil water infiltration could all affect hydrogen and oxygen isotope fractionation in the soil moisture (Gat et al. 1996; DePaolo et al. 2004). Precipitation is low and evaporation is high along the Heihe River, resulting in little downward infiltration of surface moisture to the water table. Except for certain halophytes, plant transpiration has no effect on soil moisture isotopes (Dawson and Ehleringer, 1991; Mensforth et al. 1994). Therefore, surface evaporation is the main cause of hydrogen and oxygen isotope enrichment in soil moisture along the lower Heihe River. For all ages of forests, soil water δ18O values decreased monotonically with depth. High values near the surface reflected strong isotope fractionation resulting from evaporation, while low values were similar to those for groundwater, strongly suggesting that deep soil water is derived from groundwater. These results are consistent with those of previous studies along arid land rivers (e.g., Dawson et al. 1993; Phillips et al. 2001).
Using multiple lines of evidence strengthens investigation of plant water sources. In this study, we combined analysis of stable isotopes of oxygen and hydrogen in xylem water, soil water and groundwater with measurements of soil moisture levels and tree root distribution. The results indicate that the water sources exploited by P. euphratica vary with forests age. Young forests uses soil water from 0 to 50 cm, while mature and over-mature forests use soil water from deeper than 100 cm. Water used by young forests is primarily derived from local precipitation, while water used by mature and over-mature forests is derived from groundwater. These results are similar to those reported by Zhao et al. (2008), who found that mature P. euphratica in this area use up to 93 % groundwater. These findings are also consistent with water use by trees in other arid and semi-arid desert riparian forests (Scott et al. 2000). In contrast, Zhu et al. (2007) found that both young and mature P. euphratica in Ejinaqi exploit deep soil water. This discrepancy could be explained if the young forests of Zhu et al. (2007) was older than the young forests examined here.
The age-related shift in water sources used by P. euphratica efficiently exploits the water resource and reduces competition for water between age classes. In some situations, mature and over-mature forests can take up water at depth and release it at shallower levels (Hao et al. 2009), increasing the water available to young forests. Such strategies may increase survival of this species with a high water requirement in a xeric environment.
References
Asbjornsen H, Mora G, Helmers MJ (2007) Variation in water uptake dynamics among contrasting agricultural and native plant communities in the Midwestern US. Agric Ecosyst Environ 121:343–356
Brunel JP, Walker GR, Kennett-Smith AK (1995) Field validation of isotopic procedures for determining sources of water used by plants in a semi-arid environment. J Hydrol 167:351–368
Busch DE, Ingraham NL, Smith SD (1992) Water uptake in woody riparian phreatophytes of the southwestern United States: a stable isotope study. Ecol Appl 2:450–459
Chen Y, Xu Z (2005) Plausible impact of global climate change on water resources in the Tarim River Basin. Sci China Earth Sci 48:65–73
Chen YP, Chen YN, Li WH, Zhang HF (2004) Effect of ecological water input on MDA of Populus euphratica Oliv leaf in the lower reaches of the Tarim River. Chin J Appl Envion Biol 10:408–411
Chen YN, Li WH, Chen YP, Zhao RF, Wan JH (2006) Ecological response and ecological regeneration of transfusing stream water along the dried-up watercourse in the lower reaches of the Tarim River, Xinjiang. Arid Zone Res 4:521–530
Chen YN, Chen YP, Xu CC, Li WH, Fu AH (2011) Effects of groundwater depth on the gas exchange and chlorophyll fluorescence of Populus euphratica in the lower reaches of Tarim River. Acta Ecol Sin 31:344–353
Dawson TE (1996) Determining water use by trees and forests from isotopic, energy balance and transpiration analyses: the roles of tree size and hydraulic lift. Tree Physiol 16:263–272
Dawson TE, Ehleringer JR (1991) Streamside trees that do not use stream water. Nature 350:335–337
Dawson TE, Ehleringer JR (1993) Isotopic enrichment of water in the “woody” tissues of plants: implications for plant water source, water uptake, and other studies which use the stable isotopic composition of cellulose. Geochim Cosmochim Acta 57:3487–3492
Dawson TE, Mambelli S, Plamboeck AH, Templer PH, Tu KP (2002) Stable isotopes in plant ecology. Ann Rev Ecol Syst 33(1):507–559
DePaolo DJ, Conrad ME, Maher K, Gee GW (2004) Evaporation effects on oxygen and hydrogen isotopes in deep vadose zone pore fluids at Hanford, Washington. Vadose Zone J 3:220–232
Ehleringer JR, Roden J, Dawson TE (2000) Assessing ecosystem-level water relation through stable isotope analyses. In: Sala OE, Jackson RB, Mooney HA, Howarth RW (eds) Methods in ecosystem science. Springer, New York, pp 181–198
Ellsworth PZ, Williams DG (2007) Hydrogen isotope fractionation during water uptake by woody xerophytes. Plant Soil 291:93–107
Gat JR (1996) Oxygen and hydrogen isotopes in the hydrologic cycle. Annual Rev Earth Planet Sciences 24:225–262
Hao XM, Chen YN, Li WH, Gou B, Zhao RF (2009) Evidence and ecological effects of hydraulic lift in Populus euphratica. J Plant Ecol 33:1125–1131
Hasselquist NJ, Allen MF, Santiago LS (2010) Water relations of evergreen and drought-deciduous trees along a seasonally dry tropical forests chronosequence. Ecologia 164:881–890
Li JL, Feng Q, Si JH (2008) Distribution and Simulation of uptake roots of P. euphratica Oliv in extreme arid region, China. Arid Land Geo 31:97–101
Liu L, Jia Z, Zhu Y, Liu J, Yang D, Wei D (2012) Water use strategy of different stand ages of Caragana intermedia in Alpine sandland. J Arid Land Res Environ 26:119–125
Mensforth LJ, Thorburn PJ, Tyerman SD, Walker GR (1994) Sources of water used by riparian Eucalyptus camaldulensis overlying highly saline groundwater. Oecologia 100:21–28
Nie YP, Chen HS, Wang KL, Yang J (2012) Water source utilization by woody plants growing on dolomite outcrops and nearby soils during dry seasons in karst region of Southwest China. J Hydrol 420:264–274
Overdieck D, Ziche D, Yu RD (2013) Gas exchange of Populus euphratica leaves in a riparian zone. J Arid Land 5:531–541
Phillips DL (2001) Mixing models in analysis of diet using multiple stable isotopes: a critique. Oecologia 127:166–170
Phillips DL, Gregg JW (2003) Source partitioning using stable isotopes: coping with too many sources. Oecologia 136:261–269
Scott RL, Shuttleworth WJ, Goodrich DC, Maddock T III (2000) The water use of two dominant vegetation communities in a semiarid riparian ecosystem. Agric For Meteorol 105:241–256
Yang YH, Chen YN, Cai BY, Jie WG, Lv DY (2012) Arbuscular mycorrhizae in roots of Populus euphratica in the lower reaches of Tarim River in extreme arid area. Arid Land Geo 35:260–266
Yu SW, Sun ZY, Zhou AG, Zhang X, Duan LJ, Xu SG (2012) Determination of water source of Gobi plants by D and 18O stable isotopes in middle reaches of the Heihe River. J Desert Res 32:717–723
Zhang GX, Yan H, Wei D (2004) Progresses of the applied study on the isotopic D and 18O in aqueous environment. Arid Zone Res 21:225–229
Zhao LJ, Xiao HL, Cheng GD, Song YX, Zhao L, Li CZ, Yang Q (2008) A preliminary study of water sources of riparian plants in the lower reaches of the Heihe basin. Acta Geosci Sin 29:709–718
Zhou CX, Sun ZY, Yu SW (2011) Using D and 18O stable isotopes to determine the water sources of sand dune plants in Linze, middle reaches of the Heihe River. Geol Sci Technol Inf 30:103–109
Zhu JM (2007) Studies on selective utilization of water by plants in arid land region. Chinese Academy of Forestry, Beijing, pp 78–79
Acknowledgments
This study was jointly supported by the National Natural Science Foundation of China (Grants No. 41371515) and the National Science and Technology support plan (2014BAC15B02).
Author information
Authors and Affiliations
Corresponding author
About this article

Cite this article
Liu, S., Chen, Y., Chen, Y. et al. Use of 2H and 18O stable isotopes to investigate water sources for different ages of Populus euphratica along the lower Heihe River. Ecol Res 30, 581–587 (2015). https://doi.org/10.1007/s11284-015-1270-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11284-015-1270-6