
Abstract
Aflatoxin B1 is a potent carcinogen produced by Aspergillus flavus (A. flavus) and Aspergillus. parasiticus (A. parasiticus), mainly during grain storage. The efficacy of the freeze-dried culture filtrate of Streptomyces philanthi (S. philanthi) strain RL-1-178 (DCF) on degradation of aflatoxin B1 (AFB1) were evaluated and its bioactive compounds were identified. The DCF at a concentration of 9.0% (w/v) completely inhibited growth and AFB1 production of A. parasiticus TISTR 3276 and A. flavus PSRDC-4 after 7 days tested in yeast-extract sucrose (YES) medium and on stored maize grains after 28 and 14 days incubation, respectively. This indicated the more tolerance of A. parasiticus over A. flavus. The DCF and bacterial cells of S. philanthi were capable to degrade AFB1 by 85.0% and 100% for 72 h and 8 days, respectively. This confirmed the higher efficacy of the DCF over the cells. After separation of the DCF on thin-layer chromatography (TLC) plate by bioautography bioassay, each active band was identified by liquid chromatography—quadrupole time of flight mass spectrometer (LC-Q-TOF MS/MS). The results revealed two compounds which were identified as azithromycin and an unknown based on mass ions of both ESI+ and ESI− modes. The antifungal metabolites in the culture filtrate of S. philanthi were proved to degrade aflatoxin B1. It could be concluded that the DCF may be applied to prevent the growth of the two aflatoxin-producing fungi as well as the occurrence of aflatoxin in the stored maize grains.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The colonization of maize grains by Aspergillus flavus (A. flavus) and Aspergillus. parasiticus (A. parasiticus) (Link ex Fr. and Speare, respectively; teleomorphs: Petromyces flavus and P. parasiticus) (Fountain et al. 2014, 2015; Okun et al. 2015; García-Díaz et al. 2020) results in contamination of their derived feed, and food products with aflatoxins (AFs) (Diener et al. 1987). Aflatoxin B1 (AFB1) is known as the most toxic aflatoxin, and several studies have demonstrated its hepatotoxic, carcinogenic, and mutagenic effects on human and animals (Guengerich et al. 1996; Hussein and Brasel 2001; Farzaneh et al. 2012; Afsharmanesh et al. 2018).
During the last decades, several studies have been dedicated to physical and chemical strategies for the reduction of AFs in crops, foods and feeds (Kabak et al. 2006; Afsharmanesh et al. 2018; Sipos et al. 2021; Nji et al. 2022). Nevertheless, none of these strategies completely fulfils the necessary efficacy, safety and cost requirements (Mishra and Das 2003; Zhao et al. 2011). These disadvantages encouraged a recent emphasis on the biological degradation of aflatoxins. Biodegradation of aflatoxins using microorganisms, is one of the environmentally friendly strategies to reduce or eliminate the possible contaminations of AFs in foods and feeds (Farzaneh et al. 2012; Sangare et al. 2014; Afsharmanesh et al. 2018). There are two key directions in control of aflatoxin contamination: preventing the growth of toxigenic aflatoxin-producing fungi, namely prevention and if contamination occurs, then detoxify aflatoxin-contaminated commodities by removing the toxic compounds (Xia et al. 2017; Shu et al. 2018). Over the past decades, some bacterial species have been known to degrade aflatoxin, which include Nocardia corynebacterioides (Ciegler et al. 1966; Hormisch et al. 2004), Rhodococcus erythropolis (R. erythropolis), Mycobacterium fluorantheniorans (Teniola et al. 2005), Bacillus licheniformis (B. licheniformis) (Petchkongkaew et al. 2008; Rao et al. 2017), Bacillus. Subtilis (B. subtilis) (Petchkongkaew et al. 2008; Gao et al. 2011; Farzaneh et al. 2012; Siahmoshteh et al. 2017; Xia et al. 2017; Afsharmanesh et al. 2018; Wang et al. 2019; Suresh et al. 2020), Bacillus velezensis (B. velezensis) (Shu et al. 2018; Wang et al. 2021), Bacillus. amyloliquefaciens (Siahmoshteh et al. 2017), Bacillus. Megaterium (B. megaterium) (Wang et al. 2021), Pseudomonas aeruginosa (Sangare et al. 2014). In addition, some Streptomyces sp. strains also appeared as valuable candidates for controlling filamentous fungal growth and inhibiting mycotoxin production (Harkai et al. 2016; Campos-Avelar et al. 2021). The development of efficient antifungal microbial agents could be an alternative method to control the fungi.
Actinobacteria present an interesting, natural, and cost-effective alternative for the effective biodegradation of AFs (Oliveira et al. 2013). Many species of actinomycetes, particularly those belonging to the genus Streptomyces are largely researched for their ability to produce numerous molecules of interest, namely, antibiotics (Igarashi et al. 2005; Quinn et al. 2020), antifungal compounds (Boukaew et al. 2017, 2020a, 2020b, 2021; Chen et al. 2016; Li et al. 2011; Shakeel et al. 2016), and hydrolytic enzymes (glucanase, chitinase) (Prapagdee et al. 2008; Boukaew et al. 2016; VazJauri et al. 2016), which provide them with strong antagonistic capacities against fungal development. The impact of Streptomyces sp. on toxigenic aflatoxin-producing fungi growth has already been assessed, in addition to their ability to degrade AFB1 (Verheecke et al. 2014; Harkai et al. 2016; Campos-Avelar et al. 2021) and to inhibit its production. Indeed, some Streptomyces strains produce aflastatin A, blasticidin A, and dioctatin A, three molecules that inhibit the AFs biosynthetic pathway (Sakuda et al. 1996; Sakuda 2010). In addition, a Streptomyces. roseolus strain was found to reduce AFB1 production by inhibiting aflatoxin gene cluster expression in A. flavus (Caceres et al. 2018). Streptomyces isolates IX45 can effectively restrict the growth of A. flavus growth and remove AFB1 production with 31% as previously described by Campos-Avelar et al. (2021), while 88.34% AFB1 degradation by cell-free supernatant of S. cacaoi sub sp. asoensis K234 was also observed by Harkai et al. (2016). In our previous studies, the antifungal compounds produced in tuna condensate waste medium of the strain Streptomyces philanthi (S. philanthi) showed high efficacy to inhibit mycelial growth and AFB1 production of A. flavus and A. parasiticus (Boukaew et al. 2020b, c), however, the aflatoxin B1 degradation was not investigated. As a consequence, the efficacy of antifungal metabolites of S. philanthi RL-1-178 on AFB1 degradation will be investigated in this study.
The objectives of the present study were to (i) investigate the efficacy of antifungal compounds produced by S. philanthi on growth and aflatoxin B1 production of the two aflatoxin-producing fungi, (ii) to evaluate the efficacy of antifungal compounds of S. philanthi as potential biocontrol agents of maize grain pathogenic fungi, (iii) to evaluate degradation efficiency of antifungal compounds and S. philanthi on AFB1 and (iv) to identify the active compound responsible for antifungal activity.
Materials and methods
Microorganisms and preparation of freeze-dried culture filtrate
The antagonistic strain S. philanthi RL-1-178 was previously isolated from the rhizosphere of chili pepper in southern Thailand (Boukaew et al. 2011). The culture filtrate of S. philanthi RL-1-178 was prepared by inoculating 10% (v/v) aliquots of the seed culture into 5 l bioreactor (New Brunswick™ BioFlo® 415 Sterilize-in-Place (SIP) Fermentor, Eppendorf North America) containing 4 l tuna condensate medium (pH was adjusted to 7.0 before autoclaving) and stirred (150 rpm) at 30 °C for 10 days (Boukaew et al. 2020c). Then, the culture broth was centrifuged (8880 × g for 20 min) and filtered through a 0.45 mm Millipore membrane. After that, the culture filtrate was freeze-dried by vacuum freeze-dryer at the Office of Scientific Instrument and Testing, Prince of Songkla University (PSU) to obtain the freeze-dried culture filtrate from S. philanthi RL-1-178 (called DCF).
Aflatoxigenic fungal strains and spore inoculum preparation
The A. parasiticus strain TISTR 3276 and A. flavus strain PSRDC-4 were previously found to be as high aflatoxin producers (Boukaew et al. 2020b,c). They were cultivated on potato dextrose agar (39 g l−1; Difco Laboratory) at 30 °C. Spores were collected in 5 ml water from10-day-old culture and counted using a hematocytometer. The inoculum was prepared by dilution in sterilized distilled water to achieve the required concentration.
Efficacy of the DCF concentration against the two aflatoxigenic fungal strains
The efficacy of the DCF concentration against A. parasiticus TISTR 3276 and A. flavus PSRDC-4 was investigated. One gram of the DCF was dissolved in 10 ml of dimethyl sulfoxide (DMSO) and filtered through a 0.45 mm Millipore membrane. The DCF solution for each treatment (at 0.2, 0.4, 1.0, 2.0, 4.0, 5.0, 6.0, 7.0, 8.0, 9.0, and 10.0% (w/v)) were incorporated into melted sterile yeast-extract sucrose (YES) medium at a final concentration of 45 ml. DMSO added to YES medium at equivalent amounts was used as a control. Fifty μl of spore inoculum (105 spore mL−1) of each aflatoxigenic fungal strain was transferred into each flask and incubated on a rotary shaker (150 rpm) at 30 °C for 7 days (Komala et al. 2012). After 7 days cultivation, the mycelial mats were washed (with sterile water) and dried at 80 °C until constant weight (Tolouee et al. 2010; Sangmanee and Hongpattarakere 2014). The inhibition of hyphal growth was calculated as: Percentage of inhibition = {[(Control-treatment)/Control] × 100}.
For aflatoxin B1 production, samples after treatment were extracted with chloroform according to the Association of Official Analytical Chemists (Tosch et al. 1984) and AFB1 concentration was determined as previously described by Boukaew et al. (2020c). For each treatment, three replicates were conducted. AFB1 concentration was determined by indirect competitive ELISA (Enzyme-linked Immunosorbent assay) using a ScreenEZ® Aflatoxin ELISA test kit (Siam Inter Quality Co., Ltd., Thailand).
Evaluation of the DCF efficiency on growth and aflatoxin B 1 production of the two aflatoxigenic fungal strains in stored maize grains
The effect of the DCF on growth and aflatoxin B1 production of the two aflatoxigenic fungal strains on stored maize grains was evaluated using the method as described by Afsharmanesh et al. (2018). Briefly, the 500 g surface-sterilized maize grains were soaked in 200 ml of the DCF solution and dried in a laminar airflow for 30 min. After that, the 10 ml of a spore suspension at 1 × 105 spores ml−1 of each aflatoxigenic fungal strain was aseptically placed in a sterile plastic bag containing 500 g grains and the whole mixed gently for 2 min (modified from Krusong et al. 2015). DMSO was served as a control. The aflatoxigenic fungi inoculated maize grains (AFIMG) samples were then stored at room temperature for 30 days and examined for evidence of the two aflatoxigenic fungal growth after spreading on PDA plates and incubation at 30 °C. Each treatment included three replicates. Aflatoxin estimation following each treatment was carried out at the same time. Extraction of aflatoxin from AFIMG employed a modification of the method described by Sidhu et al. (2009). The AFB1 production was determined as described above.
AFB 1 degradation by the DCF and bacterial cells of S. philanthi
AFB 1 degradation by the DCF
The effect of the DCF solution at a concentration of 10% (w/v) was tested on commercial AFB1 (Siam Inter Quality Co., Ltd., Thailand) degradation in 2-ml-Eppendorftubes a final volume of 1.0 ml according to the method of Teniola et al. (2005) with some modification. Briefly, 50 µl stock solution of AFB1 (an initial AFB1-concentration of 40.0 ppb) was added in 950 µl DCF solution. The mixture was incubated in the dark at 30 °C without shaking for 0, 12, 24, 36, 48, 60, 72, 84, and 96 h. DMSO plus AFB1 served as a control. Samples after treatment were extracted with chloroform according to the Association of Official Analytical Chemists (Tosch et al. 1984) and AFB1 concentration was determined by indirect competitive ELISA (Enzyme-linked Immunosorbent assay) using a ScreenEZ® Aflatoxin ELISA test kit (Siam Inter Quality Co., Ltd., Thailand), as described in details by Boukaew et al. (2020c). The absorbance at 450 nm (OD450) was measured, using a microplate reader (M965 + MetertechInc., Taiwan). The AFB1 concentration was estimated from OD450 using a Stat Fax Reader Model 321. Each treatment included three replicates. The percent degradation = {[(Control-treatment)/Control] × 100} (Branà et al. 2017) where the control was the concentration of AFB1 in the treatment with DMSO and treatment was the concentration of AFB1 treated with DCF solution.
AFB 1 degradation by bacterial cells of S. philanthi
AFB1 degradation ability of bacterial cells of S. philanthi was tested in a test tube with a final volume of 5.0 ml according to the method of Sangare et al. (2014) with some modification. Briefly, 100 µl of a spore suspension at 1 × 107 spores ml−1 of S. philanthi was transferred to sterilized tuna condensate medium and then added with 50 µl stock solution of AFB1 (an initial AFB1-concentration of 40.0 ppb). The detoxification test was conducted at 30℃ for 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, and 10 days. After incubation, the cells were removed by centrifugation (8880 × g for 20 min). Sterilized tuna condensate medium was used to substitute microbial culture in the control. Samples were extracted with chloroform according to the Association of Official Analytical Chemists after treatment (Tosch et al. 1984) and AFB1 degradation analysis was performed as described above. Each treatment included three replicates. The percentage of AFB1 degradation was calculated as described above.
Separation of bioactive metabolites from the DCF and testing for its efficacy against the two aflatoxigenic fungal strains using bioautography assay and identification of antifungal DCF.
Separation of bioactive metabolites from the DCF
Separation of the bioactive metabolites from the DCF solution was carried out using thin-layer chromatography (TLC) on silica gel (60 RP-18 F254S, Merck, Germany; 2 × 5 cm), with absolute methanol as the mobile phase. After that, the TLC plate was left to dry at 30 °C and sprayed with cerium sulphate (1.0%) and dried at 110 °C. The bioactive metabolite fractions were finally separated, using a bioautography assay. TLC plates were prepared in duplicate, one plate was used for bioautography assay and the other was kept for comparison.
Bioautography assay
Organic compounds, separated by TLC, were evaluated for antifungal properties. First, the TLC plate was sterilized under a UV lamp for 20 min and then placed in a PDA Petri dish, using sterile forceps. Next, it was covered by 4.0 ml of molten PDA (45 °C), containing 100 µl of spore inoculum (106 spore ml−1) of each aflatoxigenic fungal strain. The plate was incubated at 30 °C for 48 h and the inhibition caused by the active band was observed (Azish et al. 2021).
Identification of the antifungal compounds in the DCF
The active band on the TLC plate against the two aflatoxigenic fungal strains was identified by liquid chromatography –quadrupole time of flight mass spectrometer (LC-Q-TOF MS/MS). The TLC active band was scraped and dissolved in methanol plus water (1:1) and centrifuged (at 10,000 × g for 5 min), then filtered through a 0.2 µM nylon Millipore membrane to remove silica gel and debris. The UHPLC column (Zorbax Eclipse Plus C18 Rapid Resolution HD 150 mm length × 2.1 mm inner-diameter, particle size 1.8 μm, Agilent) was used for the UHPLC analysis with a column temperature of 25 °C. The flow rate was 0.4 mL min−1, and the mobile phase comprised 0.1% formic acid in H2O (A) and 0.1% formic acid in acetonitrile (B). The gradient program for the mobile phase was set as follows: 0 min (A:B = 95:5), 2 min (A:B = 95:5), 40 min (A:B = 0:100), and 45 min (A:B = 0:100). The injection volume was 5 μl. The Q-TOF/MS was operated in positive and negative electrospray ionization (ESI) modes. The operating parameters were set as follows: cone voltage of 30 V, capillary voltage of 2 kV, and source temperature of 100 °C. Data were recorded in the mass-to-charge (m/z) range of 50–1200 with a scan time of 0.25 s and an interscan time of 0.02 s for 45 min. In total, 2 LC–MS chromatograms in positive or negative modes were obtained from the active band on the TLC plate and compared by mass hunter METLIN metabolite PCD (Personal Compound Database) and PCDL (Personal Compound Database and Library) version 8.
Statistical analysis
All the experiments were done in replicates (n = 3), and the data were subjected to Analysis of Variance (ANOVA), (SPSS, version 21; IBM Corp, Armonk, NY). The mean values and their significant difference were compared using Tukey’s HSD (Honestly Significant Difference) test at P < 0.05.
Results
Efficacy of the DCF concentrations against the two aflatoxigenic fungal strains
The results showed that the inhibition on growth and AFB1 production of the two aflatoxigenic fungal strains grown in the YES medium was related to the DCF concentrations (Table 1). AFB1 production of A. flavus PSRDC-4 was 3.2 fold higher than that of A. parasiticus TISTR 3276 in the control (1352.18 and 423.12 ppb, respectively). By increasing the DCF concentrations from 0.2 to 7.0% (w/v), the AFB1 production in both fungal strains was decreased up to 96.7% (from 1283.88 to 42.10 ppb) and 85.9% (from 397.52 to 56.18 ppb), respectively. In addition, the growth inhibition in both A. flavus PSRDC-4 and A. parasiticus TISTR 3276 was significantly increased from 8.23% to 93.78%, and 17.25% to 94.32%, respectively. Growth and AFB1 production of the two fungal strains was completely inhibited (100%) at the 9.0% (w/v) DCF concentration.
Evaluation of the DCF efficacy on the growth and aflatoxin B 1 production of the two studied fungal strains in stored maize grains
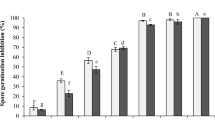
The capability of the DCF to inhibit growth and AFB1 production of the fungal strains during 28 days incubation is shown in Fig. 1. The abundant mold growth of A. parasiticus TISTR 3276 and A. flavus PSRDC-4 occurred on the control seeds in the stored maize grains but no growth occurred on the seeds treated with the DCF for 28 days of A. parasiticus TISTR 3276 (Fig. 1a) and 14 days of A. flavus PSRDC-4 (Fig. 1c). The complete inhibition of AFB1 production on the maize grains was achieved over 28 days of A. parasiticus TISTR 3276 (Fig. 1b) and 14 days of A. flavus PSRDC-4 (Fig. 1d).

Efficacy of the DCF RL-1-178 on growth and AFB1 production of a, b A. parasiticus TISTR 3276 and c, d A. flavus PSRDC-4 on stored maize seeds. The values are means of three replicates and their standard deviation
AFB 1 degradation by the DCF and bacterial cells of S. philanthi RL-1–178
The capability of the DCF and bacterial cells of S. philanthi RL-1-178 to degrade AFB1 production was shown in Fig. 2. The DCF started to degrade the AFB1 at 24 h (10.93%) and significantly increased at 60 h (71.86%) and reached the highest degradation at 72 h (85.0%) (Fig. 2a). The AFB1 concentration in the control tested was rather constant throughout the incubation period.

Time course of in vitro AFB1 degradation activity by the DCF RL-1-178 and S. philanthi RL-1-178 (1 × 107 spores ml−1). a Effect of the DCF RL-1-178 on AFB1 degradation activity during 96 h at 30 °C. b Effect of S. philanthi RL-1-178 (1 × 107 spores ml−1) on AFB1 degradation activity in tuna condensate medium during 10 days at 30 °C. The initial concentration of AFB1 was 40 ppb. The values are means of three replicates and their standard deviation. Means with different letters are significantly different according to Tukey’s HSD test (P < 0.05)
The capability of bacterial cells of S. philanthi RL-1-178 to degrade AFB1 was presented in Fig. 2b. The AFB1 degradation was first observed after 2 days (48 h) incubation (31.80%) and continuously increased to 57.50%, 77.08% degradation at 3 and 4 days, respectively, then insignificantly increased (82% to 86.89%) during 5 to 7 days, respectively. The maximum degradation of AFB1 (100%) was observed after 8 days incubation. There was no obvious change in the content of AFB1 in the control throughout the incubation period.
Separation and identification of bioactive metabolites from the DCF and evaluation of their efficacy against the two fungal strains using a bioautography assay
The bioactive metabolites from the DCF solution was separated in TLC by methanol (Fig. 3). The silica gel chromatograms showed five bands with the Rf of 0.12, 0.20, 0.28, 0.39, and 0.78, respectively (Fig. 3a). The strong band with Rf = 0.78 showed the antifungal activity against A. parasiticus TISTR 3276 (Fig. 3b) and A. flavus PSRDC-4 with positive results appeared as the clear inhibition zones around the marked fractions on the TLC plate (Fig. 3c) using bioautography assay. Identification of the bioactive metabolites using LC-Q-TOF MS/MS exhibited two compounds known as azithromycin (C38H72N2O12, tR = 13.0 min) and an unknown (tR = 20.1 min) (Fig. 4) with the same results based on mass ions of ESI+ (Fig. 4a) and ESI− (Fig. 4b) modes.

Silica gel chromatograms of freeze dry bioactive compounds of Streptomyces philanthi RL-1-178 separated in TLC. The plates were developed in absolute methanol. Chromatograms were a observed after spraying with 1.0% cerium sulphate and further exposure to 110 °C b bioautographed against A. parasiticus TISTR 3276 and c A. flavus PSRDC-4

Identification of bioactive compounds produced by Streptomyces philanthi RL-1-178 against Aspergillus parasiticus TISTR 3276 and A. flavus PSRDC-4 using liquid chromatography—quadrupole time of flight mass spectrometer (LC-Q-TOF MS/MS) based on mass ions of ESI+ (a) and ESI− (b) modes
Discussion
In our previous study, the culture filtrate of S. philanthi RL-1-178 (grown in tuna condensate and molasses medium) at a concentration of 10.0% (v/v) with 1 h exposure time demonstrated a good efficacy to inhibit both mycelial growth and aflatoxin B1 (AFB1) production of A. flavus PSRDC-4 (Boukaew et al. 2020c). In the present study, the culture filtrate of S. philanthi RL-1-178 in the form of freeze dried (DCF) was tested to evaluate its degradation efficiency on the pure aflatoxin. The DCF at 9.0% w/v could effectively inhibit mycelial growth and AFB1 production of A. flavus PSRDC-4 as well as A. parasiticus TISTR 3276. The effective dose (10%, v/v or 9–10%, w/v) of S. philanthi RL-1-178 was much lower concentration than that of Streptomyces. globisporus JK-1 (20%, v/v) and S. philanthi RM-1-138 (20%, v/v) which almost completely inhibited the growth of Magnaporthe oryzae (90.9%) (Li et al. 2011) and Rhizoctonia solani PTRRC-9 (96.0%), respectively. The high potential of the DCF to inhibit growth and aflatoxin B1 production agreed with the results on mycotoxin inhibition with antifungal metabolites of Streptomyces strains and high degradation of AFB1 as reported by Harkai et al. (2016) and Campos-Avelar et al. (2021). In addition, dioctatin A, a metabolite of Streptomyces, could reduce the mRNA level of brlA and inhibit conidiation of A. parasiticus, leading to inhibition of AFB1 production (Yoshinari et al. 2007). Therefore, it is concluded that the substance with antifungal metabolites in the culture filtrate of S. philanthi RL-1-178 can inhibit conidiation of A. parasiticus TISTR 3276 and A. flavus PSRDC-4 as well as AFB1 production. Therefore, besides measuring inhibition potential, evaluation of biological effects is an essential step in eliminating toxins.
A major problem in the storage of foods and feed stocks is spoilage and poisoning caused by fungi, Aspergillus species, and causes great economic losses worldwide (Magnusson et al. 2003). The effects of antifungal metabolites in the form of DCF on the growth of aflatoxigenic strains have never been reported so far. The results of the present study demonstrated for the first time that DCF has great potential in controlling postharvest disease caused by A. parasiticus TISTR 3276 and A. flavus PSRDC-4 on maize grain. High fungal growth and AFB1 production were observed in the control treatment within 3 days. On the other hand, the application of DCF could inhibit the growth and AFB1 production from A. parasiticus TISTR 3276 and A. flavus PSRDC-4, after 28 and 14 days of maize grain storage, respectively. Thus, it could be concluded that the antifungal metabolites of S. philanthi RL-1-178 could be applied in the form of DCF which shortcut the cost of purification stage. This finding suggested the simpler process to produce and apply antifungal compounds in the crude form (DCF) that able to inhibit the Aspergillus species. The results were in good agreement with the experiments conducted by Bressan (2009), in which the treatment of maize seeds with Streptomyces sp. culture filtrate reduced the development of fungi in stored seeds. Biological methods to control of post-harvest diseases in seeds are based on the use of living cells and cell-free compounds, such as Bacillus sp. (Ongena and Jacques 2008; Yánez-Mendizabal et al. 2012; Yánez-Mendizábal and Falconí 2018), Trichoderma sp. (Coşkuntuna and Özer 2008; Reddy et al. 2009; Xue et al. 2017), Pseudomonas sp. (Reddy et al. 2009), and Streptomyces sp. (Sultan and Magan 2011). One of the main factors determining the efficacy of biological control is the method of inoculation of the biological agent. This study indicated that grain inoculation with the DCF provides a significant reduction in the incidence of pathogenic seed fungi and has potential as a biological control agent. Therefore, DCF was found to show significant anti-fungal growth and anti-aflatoxin B1 production, hence, it could be explored as biopreservatives for preventing microbial deterioration and mycotoxins accumulation in food and feedstuffs during pre- and post-harvest and storage.
Streptomyces strains have been also applied in detoxification processes against mycotoxins in several ways. Some studies revealed only the antagonist effects of Streptomyces strains against toxin producing fungi (Harkai et al. 2016; Campos-Avelar et al. 2021). According to Verheecke et al. (2014), Streptomyces strains inhibited AFB1 production in A. flavus by gene repression. Toxin degradation is an effective remedy for food that has been infested with toxins (Sun et al. 2023). In this study, the AFB1-degrading capability of the DCF and bacterial cells of S. philanthi RL-1-178 were investigated. Our results confirmed that up to 71.86% AFB1 was eliminated within 60 h of applying the DCF, and > 85% degradation was observed within 72 h. Results implied that the molecules present in DCF (a protein (enzyme) or proteins (enzymes) might be involved in the degradation of AFB1. Many investigators have reported that several laccases produced by Streptomyces are involved in the catabolic pathways of aromatic compounds via a cascade of reactions (Park and Kim 2003; Davis and Sello 2009; Qin et al. 2021). AFB1 is also a polyaromatic compound and could be degraded in a similar manner. Therefore, it is suggested that the DCF can cleave the lactone ring of AFB1 and it is likely an enzymatic degradation. These results suggested that some soluble signal molecules secreted from secondary metabolites by S. philanthi RL-1-178 could inhibit AFB1 biosynthesis, which was similar to the results of Harkai et al. (2016). They showed 88.34% AFB1 degradation by the cell-free supernatant of S. cacaoi sub sp. asoensis K234 after 12 h treatment. Detoxification of AFB1 by cell-free supernatant obtained from many antagonistic bacterial strains, such as R. erythropolis and Mycobacterium fluoranthenivorans sp. nov. DSM44556T (Teniola et al. 2005), Pseudomonas sp. (Sangare et al. 2014), B. licheniformis CFR1 (Rao et al. 2017), B. subtilis (Xia et al. 2017; Suresh et al. 2020), B. velezensis DY3108 (Shu et al. 2018), and B. megaterium (Wang et al. 2021) has previously been reported. The majority of the studied bacterial cells of Streptomyces isolates were able to degrade AFB1, either in solid or liquid media. This is in agreement with the findings of Campos-Avelar et al. (2021) and Harkai et al. (2016) whose degradation assays with Streptomyces strains proved that all of them were able to significantly degrade AFB1. In general, aflatoxin degradation by Streptomyces has been demonstrated by several authors (Verheecke et al. 2015; Harkai et al. 2016; Avelar et al. 2021). Herein, the AFB1 degrading potential value (100%) of S. philanthi RL-1-178 was similar to those of Streptomyces. lividans and Streptomyces. aureofaciens (Eshelli et al. 2015) but higher than the results of other Streptomyces species. These included the Streptomyces isolate MYC (31%) (Campos-Avelar et al. 2021), S. cacaoi subsp. asoensis K234 (88.33%), Streptomyces. sanglieri K139 (61.43%), Streptomyces. luteogriseus K144 (79.93%), Streptomyces. rimosus K145 (79.93%), and Streptomyces. cinereoruber K236 (58.52%) (Harkai et al. 2016). Thus, different AFB1 degradation activities among various Streptomyces species were the result of different levels of active compounds in cell-free supernatant which are responsible for the diminished AFB1. From the experiments conducted, the DCF and bacterial cells of S. philanthi were capable to degrade AFB1 by 85.0% and 100% for 72 h and 8 days, respectively. This confirmed the higher efficacy of the DCF over the cells. This is in agreement with the findings that the cell-free supernatant was predominantly attributed in the AFB1 degrading activity than bacterial cells (Xia et al. 2017; Shu et al. 2018). Dioctatin A (DotA), a metabolite of Streptomyces, inhibited production of norsolorinic acid which is an early biosynthetic intermediate of aflatoxin produced by A. parasiticus (Yoshinari et al. 2007). In addition, it also strongly reduced the mRNA levels of genes responsible for aflatoxin biosynthetic enzymes and the mRNA level of aflR encoding a key regulator protein for aflatoxin biosynthesis. There is limited evidence on the capability of Streptomyces sp. on decrease of AFB1-contents (Sakuda et al. 1996; Zucchi et al. 2008; Harkai et al. 2016), whereas several reports demonstrated that some strains of Streptomyces sp. could inhibit the mycelial growth and aflatoxin production of A. flavus (Sultan and Magan 2011; Caceres et al. 2018; Shakeel et al. 2018; Boukaew et al. 2020b,c; Campos-Avelar et al. 2021) and A. parasiticus (Boukaew et al. 2020b,c). However, the present study exhibited that the DCF and bacterial cells of S. philanthi RL-1-178 had a high potential for degrading AFB1. The DCF had highly inhibitory effects on growth and AFB1 production of A. parasiticus TISTR 3276 and A. flavus PSRDC-4 both in the in vitro and on maize grains.
Separation and localization of the bioactive compounds by TLC on silica gel and bioautography were again proved the presence of antifungal compounds. Based on the TLC results, the DCF with Rf = 0.78 exhibited antifungal activities on both aflatoxin-producing fungi, as confirmed by the bioautography assay. The result is similar to the observation of Azish et al. (2021) that the partial purification of the antifungal metabolites of Streptomyces. libani on TLC with Rf = 0.88 exhibited anti-Aspergillus. fumigatus activities while the bioactive compound of Streptomyces. albidoflavus 321.2 with Rf = 0.85 showed antifungal activity against Aspergillus. niger (Eshelli et al. 2015). Streptomyces produced antimicrobial compounds that showed Rf values ranging from 0.40 to 0.78 in TLC analysis, which confirmed the production of polyene nature of compounds (Selvakumar et al. 2010). The active band was separated as white amorphous powder, which was identified as azithromycin (C38H72N2O12, tR = 13.0 min) and an unknown substance (tR = 20.1 min) based on the mass ions of ESI+ and ESI−modes. Azithromycin, a second generation macrolide, broad-spectrum antifungal substance against Aspergillus sp. (Nguyen et al. 1997; Guo et al. 2018) and Fusarium solani (Guo et al. 2018) and antibacterial substance against Bordetella pertussis and Legionella sp. (Parnham et al. 2014), has received increasing attention in recent years because of additional effects on host-defense reactions and chronic human diseases. It also has activity against Mycoplasma pneumoniae, Treponema pallidum, Chlamydia sp., and Mycobacterium avium complex (Parnham et al. 2014). It was reported that azithromycin exerted its antifungal activity against Aspergillus species by inhibiting mitochondrial and cytoplasmic protein synthesis (Nguyen et al. 1997). The DCF revealed two compounds which were identified as azithromycin and an unknown substance that possessed antifungal activity. Therefore, it could be concluded that the antifungal activity of the integral DFC may be attributable to two compounds resulting from independence or synergistic effect.
In conclusion, the antifungal and anti-mycotoxigenic capabilities of the DCF RL-1-178 both in vitro and on maize grains were evaluated. The DCF was applied to inhibit fungal growth and aflatoxin production, both in vivo and in vitro. These results indicated that the antifungal metabolites of S. philanthi RL-1-178 could be considered as potential biocontrol agents to combat toxigenic fungal growth and subsequent aflatoxin contamination of maize and other agricultural crops in practice.
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Afsharmanesh H, Perez-Garcia A, Zeriouh H, Ahmadzadeh M, Romero D (2018) Aflatoxin degradation by Bacillus subtilis UTB1 is based on production of an oxidoreductase involved in bacilysin biosynthesis. Food Control 94:48–55
Azish M, Shams-Ghahfarokhi M, Razzaghi-Abyaneh M (2021) Antifungal activity and mechanism of action of dichloromethane extract fraction A from Streptomyces libani against Aspergillus fumigatus. J Appl Microbiol 131:1212–1225
Boukaew S, Chuenchit S, Petcharat V (2011) Evaluation of Streptomyces spp. for biological control of Sclerotium root and stem rot and Ralstonia wilt of chili. Biocontrol 56:365–347
Boukaew S, Petlamul W, Suyotha W, Prasertsan P (2016) Simultaneous fermentative chitinase and β-1,3 glucanase production from Streptomyces philanthi RM-1-1-38 and their antifungal activity against rice sheath blight disease. Biotechnologia 97:271–284
Boukaew S, Prasertsan P, Troulet C, Bardin M (2017) Biological control of tomato gray mold caused by Botrytis cinerea by using Streptomyces spp. Biocontrol 62:793–803
Boukaew S, Petlamul W, Prasertsan P (2020a) Comparison of the biocontrol efficacy of culture filtrate from Streptomyces philanthi RL-1-178 and acetic acid against Penicillium digitatum, in vitro and in vivo. Eur J Plant Pathol 158:939–949
Boukaew S, Petlamul W, Prasertsan P (2020b) Efficacy of Streptomyces philanthi RL-1-178 culture filtrate against growth and aflatoxin B1 production by two aflatoxigenic fungi on maize seeds. Eur J Plant Pathol 156:1041–1051
Boukaew S, Petlamul W, Prasertsan P (2020c) Tuna condensate waste with molasses as a renewable substrate for antifungal compounds by Streptomyces philanthi RL-1-178 against aflatoxingenic B1 (AFB1) Aspergillus flavus. Waste Biomass Valorization 11:1321–1331
Boukaew S, Cheirsilp B, Yossan S, Khunjan U, Petlamul W, Prasertsan P (2021) Utilization of palm oil mill effluent as a novel substrate for the production of antifungal compounds by Streptomyces philanthi RM-1-138 and evaluation of its efficacy in suppression of three strains of oil palm pathogen. J Appl Microbiol 132:1990–2003
Branà MT, Cimmarusti MT, Haidukowski M, Logrieco AF, Altomare C (2017) Bioremediation of aflatoxin B1-contaminated maize by king oyster mushroom (Pleurotus eryngii). PLoS ONE 12:e0182574
Bressan W (2009) Biological control of maize seed pathogenic fungi by use of actinomycetes. Biocontrol 48:233–240
Caceres I, Snini SP, Puel O, Mathieu F (2018) Streptomyces roseolus, a promising biocontrol agent against Aspergillus flavus, the main aflatoxin B1 producer. Toxins 10:442
Campos-Avelar I, de la Noue AC, Durand N, Cazals G, Martinez V, Strub C, Fontana A, Schorr-Galindo S (2021) Aspergillus flavus growth inhibition and aflatoxin B1 decontamination by Streptomyces isolates and their metabolites. Toxins 13:340
Chen YY, Chen PC, Tsay TT (2016) The biocontrol efficacy and antibiotic activity of Streptomyces plicatus on the oomycete Phytophthora capsici. Biol Control 98:34–42
Ciegler A, Lillehoj EB, Peterson RE, Hall HH (1966) Microbial detoxification of aflatoxin. Appl Microbiol 14:934–938
Coşkuntuna A, Özer N (2008) Biological control of onion basal rot disease using Trichoderma harzianum and induction of antifungal compounds in onion set following seed treatment. Crop Prot 27:330–336
Davis JR, Sello JK (2009) Regulation of genes in Streptomyces bacteria required for catabolism of lignin-derived aromatic compounds. Appl Microbiol Biotechnol 86:921–929
Diener UL, Cole RJ, Sanders TH, Payne GA, Lee S, Klich MA (1987) Epidemiology of aflatoxin formation by Aspergillus flavus. Annu Rev Phytopathol 25:249–270
Eshelli M, Harvey L, Edrada-Ebel R, McNeil B (2015) Metabolomics of the bio-degradation process of aflatoxin B1 by Actinomycetes at an initial pH of 6.0. Toxins 7:439–456
Farzaneh M, Shi ZQ, Ghassempour A, Sedaghat N, Ahmadzadeh M, Mirabolfathy M, Avan-Nikkhah M (2012) Aflatoxin B1 degradation by Bacillus subtilis UTBSP1 isolated from pistachio nuts of Iran. Food Control 23:100–106
Fountain JC, Scully BT, Ni X, Kemerait RC, Lee RD, Chen ZY, Guo B (2014) Environmental influences on maize-Aspergillus flavus interactions and aflatoxin production. Front Microbiol 5:1–7
Fountain JC, Khera P, Yang L, Nayak SN, Scully BT, Lee RD, Chen ZY, Kemerait RC, Varshney RK, Guo B (2015) Resistance to Aspergillus flavus in maize and peanut: molecular biology, breeding, environmental stress, and future perspectives. Crop J 3:229–237
Gao X, Ma Q, Zhao L, Lei Y, Shan Y, Ji C (2011) Isolation of Bacillus subtilis: screening for aflatoxins B1, M1, and G1 detoxification. Eur Food Res Technol 232:957–962
García-Díaz M, Gil-Serna J, Vázquez C, Botia MN, Patiño B (2020) A comprehensive study on the occurrence of mycotoxins and their producing fungi during the maize production cycle in Spain. Microorganisms 8:141
Guengerich FP, Johnson WW, Ueng YF, Yamazaki H, Shimada T (1996) Involvement of cytochrome P450, glutathione S-transferase, and epoxide hydrolase in the metabolism of aflatoxin B1 and relevance to risk of human liver cancer. Environ Health Perspect 104:557–562
Guo H, Zhou L, He Y, Gao C, Han L, Xu Y (2018) Natamycin and azithromycin are synergistic in vitro against ocular pathogenic Aspergillus flavus species complex and Fusarium solani species complex isolates. Antimicrob Agents Chemother 62:e00077-e118
Harkai P, Szabó I, Cserháti M, Krifaton C, Risa A, Radó J, Balázs A, Berta K, Kriszt B (2016) Biodegradation of aflatoxin-B1 and zearalenone by Streptomyces sp. collection. Int Biodeterior Biodegradation 108:48–56
Hormisch D, Brost I, Kohring GW, Giffhorn F, Kroppen-stedt RM, Stackebrandt E, Färber P, Holzapfel WH (2004) Mycobacterium fluoranthenivorans sp. nov., a fluoranthene and aflatoxin B1 degrading bacterium from contaminated soil of a former coal gas plant. Syst Appl Microbiol 27:653–660
Hussein SH, Brasel JM (2001) Toxicity, metabolism and impact of mycotoxins on humans and animals. Toxicology 167:101–134
Igarashi M, Takahashi Y, Shitara T, Nakamura H, Naganawa H, Miyake T, Akamatsu Y (2005) Caprazamycins, novel lipo-nucleoside antibiotics, from Streptomyces sp. J Antibiot 58:327–337
Kabak B, Dobson AD, Var I (2006) Strategies to prevent mycotoxin contamination of food and animal feed: a review. Crit Rev Food Sci Nutr 46:593–619
Komala VV, Ratnavathi CV, Kumar BV, Das IK (2012) Inhibition of aflatoxin B1 production by an antifungal component, eugenol in stored sorghum grains. Food Control 26:139–146
Krusong W, Jindaprasert A, Laosinwattana C, Teerarak M (2015) Baby corn fermented vinegar and its vapor control postharvest decay in strawberries. N Z J Crop Hortic Sci 43:193–203
Li Q, Jiang Y, Ning P, Zheng L, Huang J, Li G, Jiang D, Hsiang T (2011) Suppression of Magnaporthe oryzae by culture filtrates of Streptomyces globisporus JK-1. Biol Control 58:139–148
Magnusson J, Stro”m K, Roos S, Sjo”gren J, Schnu”rer J (2003) Broad and complex antifungal activity among environmental isolates of lactic acid bacteria. FEMS Microbiol Lett 219:129–135
Mishra HN, Das C (2003) A review on biological control and metabolism of aflatoxin. Crit Rev Food Sci Nutr 43:245–264
Nguyen MH, Clancy CJ, Yu YC, Lewin AS (1997) Potentiation of antifungal activity of amphotericin B by azithromycin against Aspergillus species. Eur J Clin Microbiol Infect Dis 16:846–848
Nji QN, Babalola OO, Mwanza M (2022) Aflatoxins in maize: can their occurrence be effectively managed in Africa in the face of climate change and food insecurity? Toxins 14:574
Okun DO, Khamis FM, Muluvi GM, Ngeranwa JJ, Ombura FO, Yongo MO, Kenya EU (2015) Distribution of indigenous strains of atoxigenic and toxigenic Aspergillus flavus and Aspergillus parasiticus in maize and peanuts agro-ecological zones of Kenya. Agric Food Secur 4:14
Oliveira CAF, Bovo F, Humberto C, Vincenzi A, Ravindranadha K (2013) Recent trends in microbiological decontamination of aflatoxins in foodstuffs. In: Razzaghi-Abyaneh M (ed) Aflatoxins—recent advances and future prospects. In Tech, Rijeka, Croatia, pp 59–62
Ongena M, Jacques P (2008) Bacillus lipopeptides: versatile weapons for plant disease biocontrol. Trends Microbiol 16:115–125
Park H-J, Kim E-S (2003) An inducible Streptomyces gene cluster involved in aromatic compound metabolism. FEMS Microbiol Let 226:151–157
Parnham MJ, Erakovic Haber V, Giamarellos-Bourboulis EJ, Perletti G, Verleden GM, Vos R (2014) Azithromycin: mechanisms of action and their relevance for clinical applications. Pharmacol Ther 143:225–245
Petchkongkaew A, Taillandier P, Gasaluck P, Lebrihi A (2008) Isolation of Bacillus spp. from Thai fermented soybean (Thua-nao): screening for aflatoxin B1 and ochratoxin A detoxification. J Appl Microbiol 104:1495–1502
Prapagdee B, Kuekulvong C, Mongkolsuk S (2008) Antifungal potential of extracellular metabolites produced by Streptomyces hygroscopicus against phytopathogenic fungi. Int J Biol Sci 4:330–337
Qin X, Xin Y, Zou J, Su X, Wang X, Wang Y, Zhang J, Tu T, Yao B, Luo H, Huang H (2021) Efficient degradation of aflatoxin B1 and zearalenone by laccase-like multicopper oxidase from Streptomyces thermocarboxydus in the presence of mediators. Toxins 13:754
Quinn GA, Banat AM, Abdelhameed AM, Banat IM (2020) Streptomyces from traditional medicine: sources of new innovations in antibiotic discovery. J Med Microbiol 69:1040–1048
Rao KR, Vipin AV, Hariprasad P, Anu Appaiah KA, Venkateswaran G (2017) Biological detoxification of aflatoxin B1 by Bacillus licheniformis CFR1. Food Control 71:234–241
Reddy KRN, Reddy CS, Muralidharan K (2009) Potential of botanicals and biocontrol agents on growth and aflatoxin production by Aspergillus flavus infecting rice grains. Food Control 20:173–178
Sakuda S (2010) Mycotoxin production inhbitors from natural products. Mycotoxins 60:79–86
Sakuda S, Ono M, Furihata K, Nakayama J, Suzuki A, Isogai A (1996) Aflastatin A, a novel inhibitor of aflatoxin production of Aspergillus parasiticus, from Streptomyces. J Am Chem Soc 118:7855–7856
Sangare L, Zhao Y, Minnie Y, Folly E, Chang J, Li J, Selvaraj JN, Xing F, Zhou L, Wang Y, Liu Y (2014) Aflatoxin B1 degradation by a Pseudomonas Strain. Toxins 6:3028–3040
Sangmanee P, Hongpattarakere T (2014) Inhibitory of multiple antifungal components produced by Lactobacillus plantarum K35 on growth, aflatoxin production and ultrastructure alterations of Aspergillus flavus and Aspergillus parasiticus. Food Control 40:224–233
Selvakumar D, Arun K, Suguna S, Kumar D, Dhevendaran K (2010) Bioactive potential of Streptomyces against fish and shellfish pathogens. Iran J Microbiol 2:157–164
Shakeel Q, Lyu A, Zhang J, Wu M, Chen S, Chen W, Li G, Yang L (2016) Optimization of the cultural medium and conditions for production of antifungal substances by Streptomyces platensis 3–10 and evaluation of its efficacy in suppression of clubroot disease (Plasmodiophora brassicae) of oilseed rape. Biol Control 101:59–68
Shakeel Q, Lyu A, Zhang J, Wu M, Li G, Hsiang T, Yang L (2018) Biocontrol of Aspergillus flavus on peanut kernels using Streptomyces yanglinensis 3–10. Front Microbiol 9:1049
Shu X, Wang Y, Zhou Q, Li M, Hu H, Ma Y, Chen X, Ni J, Zhao W, Huang S, Wu L (2018) Biological degradation of aflatoxin B1 by cell-free extracts of Bacillus velezensis DY3108 with broad pH stability and excellent thermostability. Toxins 10:330
Siahmoshteh F, Siciliano I, Banani H, Hamidi-Esfahani Z, Razzaghi-Abyaneh M, Gullino ML, Spadaro D (2017) Efficacy of Bacillus subtilis and Bacillus amyloliquefaciens in the control of Aspergillus parasiticus growth and aflatoxins production on pistachio. Int J Food Microbiol 254:47–53
Sidhu OP, Chandra H, Behl HM (2009) Occurence of aflatoxins in mahua Madhuca indica Gmel.) seeds: synergistic effect of plant extracts on inhibition of Aspergillus flavus growth and aflatoxin production. Food Chem Toxicol 47:774–777
Sipos P, Peles F, Brassó DL, Béri B, Pusztahelyi T, Pócsi I, Gyori Z (2021) Physical and chemical methods for reduction in aflatoxin content of feed and food. Toxins 13:204
Sultan Y, Magan N (2011) Impact of a Streptomyces (AS1) strain and its metabolites on control of Aspergillus flavus and aflatoxin B1 contamination in vitro and in stored peanuts. Biocontrol Sci Technol 21:1437–1455
Sun F, Yu D, Zhou H, Lin H, Yan Z, Wu A (2023) CotA laccase from Bacillus licheniformis ZOM-1 effectively degrades zearalenone, aflatoxin B1 and alternariol. Food Control 145:109472
Suresh G, Cabezudo I, Pulicharla R, Cuprys A, Rouissi T, Brar SK (2020) Biodegradation of aflatoxin B1 with cell-free extracts of Trametes versicolor and Bacillus subtilis. Res Vet Sci 133:85–91
Teniola OD, Addo PA, Brost IM, Farber P, Jany KD, Alberts JF, van Zyl WH, Steyn PS, Holzapfel WH (2005) Degradation of aflatoxin B (1) by cell-free extracts of Rhodococcus erythropolis and Mycobacterium fluoranthenivorans sp. nov. DSM 44556(T). Int J Food Microbiol 105:111–117
Tolouee M, Alinezhad S, Saberi R, Eslamifar A, Zad SJ, Jaimand K, Taeb J, Rezaee MB, Kawachi M, Shams-Ghahfarokhi M, Razzaghi-Abyaneh M (2010) Effect of Matricaria chamomilla L. flower essential oil on the growth and ultrastructure of Aspergillus niger van Tieghem. Int J Food Microbiol 139:127–133
Tosch D, Waltking AE, Schlesier JF (1984) Comparison of liquid chromatography and high performance thin layer chromatography for determination of aflatoxin in peanut products. J Associ Anal Chem 67:337–339
VazJauri P, Altier N, Kinkel LL (2016) Streptomyces for sustainability. In: Castro-Sowinski S (ed) Microbial models: from environment to industrial sustainability, microorganism for sustainability 1. Springer, Berlin, pp 251–276
Verheecke C, Liboz T, Darriet M, Sabaou N, Mathieu F (2014) In vitro interaction of actinomycetes isolates with Aspergillus flavus: impact on aflatoxins B1 and B2 production. Lett Appl Microbiol 58:597–603
Verheecke C, Liboz T, Anson P, Zhu Y, Mathieu F (2015) Streptomyces-Aspergillus flavus interactions: impact on aflatoxin B accumulation. Food Addit Contam Part A 32:572–576
Wang X, Bai Y, Huang H, Tu T, Wang Y, Wang Y, Luo H, Yao B, Su X (2019) Degradation of aflatoxin B1 and zearalenone by bacterial and fungal laccases in presence of structurally defined chemicals and complex natural mediators. Toxins 11:609
Wang L, Huang W, Sha Y, Yin H, Liang Y, Wang X, Shen Y, Wu X, Wu D, Wang J (2021) Co-cultivation of two Bacillus strains for improved cell growth and enzyme production to enhance the degradation of aflatoxin B1. Toxins 13:435
Xia X, Zhang Y, Li M, Garba B, Zhang Q, Wang Y, Zhang H, Li P (2017) Isolation and characterization of a Bacillus subtilis strain with aflatoxin B1 biodegradation capability. Food Control 75:92–98
Xue AG, Guo W, Chen Y, Siddiqui I, Marchand G, Liu J, Ren C (2017) Effect of seed treatment with novel strains of Trichoderma spp. on establishment and yield of spring wheat. Crop Prot 96:97–102
Yánez-Mendizábal V, Falconí CE (2018) Efficacy of Bacillus spp. to biocontrol of anthracnose and enhance plant growth on andean lupin seeds by lipopeptide production. Biol Control 122:67–75
Yánez-Mendizábal V, Zeriouh H, Viñas I, Torres R, Usall J, De Vicente A, Pérez-García A, Teixidó N (2012) Biological control of peach brown rot (Monilinia spp.) by Bacillus subtilis CPA-8 is based on production of fengycinlike lipopeptides. Eur J Plant Pathol 132:609–619
Yoshinari T, Akiyama T, Nakamura K, Kondo T, Takahashi Y, Muraoka Y, Nonomura Y, Nagasawa H, Sakuda S (2007) Dioctatin A is a strong inhibitor of aflatoxin production by Aspergillus parasiticus. Microbiol 153:2774–2780
Zhao LH, Guan S, Gao X, Ma QG, Lei YP, Bai XM, Ji C (2011) Preparation, purification and characteristics of an aflatoxin degradation enzyme from Myxococcus fulvus NSM068. J Appl Microbiol 110:147–155
Zucchi TD, De Moraes LAB, De Melo IS (2008) Streptomyces sp. ASBV-1 reduces aflatoxin accumulation by Aspergillus parasiticus in peanut grains. J Appl Microbiol 105:2153–2160
Funding
This research work was financially supported by the Agricultural Research Development Agency (Public Organization) (PRP6405030400) and Thailand Research Fund (TRF) under Research Team Promotion Grant No. (RTA6080010).
Author information
Authors and Affiliations
Contributions
SB: contributed to the design and implementation of the research, to the analysis of the results, and the writing of the manuscript. PP: contributed to the design of the research project and editing the manuscript. PM, TS and WP: contributed to the interpretation of the research data. All authors discussed the results and contributed to the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have not disclosed any competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Boukaew, S., Prasertsan, P., Mahasawat, P. et al. Efficacy of the antifungal metabolites of Streptomyces philanthi RL-1-178 on aflatoxin degradation with its application to prevent aflatoxigenic fungi in stored maize grains and identification of the bioactive compound. World J Microbiol Biotechnol 39, 24 (2023). https://doi.org/10.1007/s11274-022-03470-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11274-022-03470-7