
Abstract
The effects of alkaline dust emitted from a cement plant for over 40 years on the anatomy of needles of Scots pine and lignin accumulation were analysed. Comparative analytical studies were conducted in stands similar as to their silvicultural indicators, climate and age in alkalised and in a relatively unpolluted area. Cross-section of needles were stained, photographed under microscope and measured. It was found that, due to the alkalisation of the environment, the total area of the needle cross-section, needle width and thickness and the area of mesophyll had decreased. At the same time, the vascular bundles and epidermis had increased. The greatest anatomical and biochemical differences between the needles from trees growing under optimum conditions and in the alkalised area were observed in the oldest needles. Visual analysis of cross-sections and biochemical analysis showed accumulation of lignin in older needles but more intensively in alkalised areas than in control.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
The cement plant in Kunda, Northeast Estonia, was one of the major pollution sources in Estonia, which emitted large quantities of alkaline cement dust for over 40 years. In certain periods, these emissions made up nearly 10% of the cement output of the plant, and dust deposition in the close vicinity of the plant was 1,600–2,700 g/m2 (Mandre 1995). By 1997, thanks to the installation of efficient filters in 1994, the dust emission from the plant had decreased approximately 36-fold of the level of 1992 (Liblik et al. 2000). Today, the emission of dust from the cement plant does not exceed the permissible level (543 tonnes/year; Environmental Review No. 16. 2007). Nevertheless, cement dust belongs to such environmentally hazardous industrial wastes that react with air moisture and become chemically active (Borka 1983). The dust emitted from the cement plant has brought about alkalisation of soil and soil water, which in turn has caused changes in the metabolism and morphological parameters of trees on the surrounding territory at a distance of 5 km (Mandre et al. 1995; Mandre and Ots 1999; Liblik et al. 2000). Earlier studies have shown that in the vicinity of Kunda cement plant the length growth of pine needles and shoots (Mandre et al. 1995), the height growth of trees (Mandre and Ots 1995) and radial increment and bioproduction (Rauk 1995) are inhibited. As the distance from the cement plant increases, the difference in length growth between needles of pines growing in the unpolluted area and in the polluted area decreases (Ots 2000; Ots and Rauk 2000), whereas small quantities of cement dust may even stimulate needle and shoot growth (Mandre et al. 1995). Although dust emissions from the cement plant have practically ceased, the impacts of long-term dust pollution will be evident in the tree physiology and biochemistry for a long time. A study of the dynamics of the pigment system of coniferous trees indicated that their status had not improved by 1999 (Mandre and Korsjukov 2000). The predominant elements of dust pollution prevail also in the mineral composition of trees (Mandre et al. 2008). No noteworthy improvement was observed in the carbohydrate metabolism of needles either (Klõšeiko 2005). Likewise, the density of needles on shoots has not changed significantly (Ots 2005). All this allows us to postulate that the effect of dust pollution from the cement plant is long-lasting and that also the anatomy of trees is affected.
The aim of the research reported in this paper was to study the anatomy and lignification of needles of Scots pine trees in areas affected by cement dust pollution for over 40 years near the cement plant at different distances from Kunda cement plant.
2 Material and Methods
2.1 Study Area and Plant Material
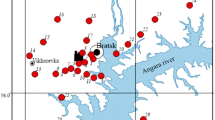
Needle samples were collected in May 2007 before bud break. The research transect is located near the cement plant in the town of Kunda (Northeast Estonia). Five sampling sites (Kunda, K) were in the zone of high pollution 2.5–3.0 km east of the cement plant and were located on the transect at distances of 100 m from one another, beginning from the first site (Fig. 1). Three control sites (control, C) were selected on the unpolluted territory of Lahemaa National Park in the similar climatic and edaphic conditions (38 km W from the cement plant, opposite to prevailing winds; Mandre and Tuulmets 1995).

Sampling sites on the W–E investigation transect. Five sampling sites are located in Kunda (K) E of the cement plant and three control sites (C) W of the cement plant. The distance between two sites is 100 m
2.2 Anatomical Analyses
For morphological and anatomical analyses needles aged 1, 2, and 3 years (n = 15 from each age group) were collected from the middle of the crowns of three trees from each control site and of 12 trees from each sampling site near the cement plant.
For anatomical and histochemical analyses cross-sections of needles were made. The needles were pre-fixed with 3% glutaraldehyde in 0.1 mol/L phosphate buffer, pH = 7.3, and fixed in 1% solution of OsO4 (Bozzola and Russell 1992; Ruzin 1999). Then, tissues were dehydrated with ethanol and xylol (Fluka, USA) and embedded in paraffin. The cross-sections of needles (10–15 µm) were made with a microtome (Leitz, Germany) and mounted on glass.
For lignin visualisation, paraffin was removed, and the slices were stained with 5% safranin O and Fast Green FCF (Fluka, USA). The lignified cell walls stained red and the others green. The cross-sections were viewed at ×100 and ×400 magnification with a Micros MC400A (Austria) microscope and photographed with a Nikon Coolpix 5400 camera (Nikon, Tokyo, Japan).
For understanding changes in anatomical peculiarities, the total cross-sectional area, the areas of epidermis, mesophyll, xylem, phloem and sclerenchyma, and needle width and thickness were measured on the cross-sections (Lukjanova and Mandre 2006) from three parts of the needles (top, middle and bottom) to obtain average values describing the needle as a whole. MapInfo Professional for Windows 4.0 (MapInfo Corp. Inc., Troy, NY) and UTHSCSA Image Tool for Windows Version 3.00 (The University of Texas, Health Science Center, San Antonio, TX, USA) were used to measure the tissue area from the images. Average values of anatomical characteristics were calculated for all these tissues. The stained cross-sections revealed the localisation of lignin. The degree of lignification was determined visually by the intensity of staining.
2.3 Biochemical Analyses
2.3.1 Analyses of Lignin
The fresh needles of each investigated tree were carefully cleaned. The fresh mass of the separated needles was weighed, and their dry mass was established after they had been kept at 105°C for 60 h. Lignin was determined as acid-insoluble lignin (Arasimovich and Ermakov 1987; Monties 1989) in the Plant Production Analyses Laboratory of the Estonian Research Agricultural Centre. Air-dried plant material was ground and extracted with acetone (100%) at 5°C, ethanol (96%), ethanol–benzene solution (1:1, v/v) and water at 60–70°C to remove sugars, proteins, interfering phenolics and other soluble compounds. The extraction was repeated several times until the solution remained colourless. The residue was dried at 70°C for 24 h and hydrolysed with H2SO4 (1 ml of 72% H2SO4 was added to each 100 mg of sample). A fraction of acid-soluble lignin and cellulose could be lost during this procedure (Monties 1989). Acid-resistant residue was recovered by filtration on a glass crucible with an asbestos filter, carefully washed and dried at 70°C for 24 h to constant weight. This acid-insoluble residue is insoluble lignin (hereafter called “lignin”) (Monties 1989). After weighing, the residue was ashed at 525 ± 25°C for at least 3 h, and lignin was calculated after correcting for mineral elements (Van Soest 1987). The results for lignin were expressed as a percentage of dry weight (DW) of plant material.
2.3.2 Analyses of Pigments
From three trees in each sampling site (n = 8), needles for analyses of chlorophylls (Chl) were collected and fixed in liquid nitrogen and homogenised in 80% ice-cold acetone solution for 3–5 min under dim light at a low temperature. The extract was filtered through a fritted-glass filter. The content of pigments was measured with a spectrophotometer Helios (Unicam Ltd., UK) at a wavelength of 649 for Chl a and 665 for Chl b and calculated in milligram per gram DW according to formulas of Vernon (1960) and Lichtenthaler and Wellburn (1983).
2.4 Statistical Analyses
The significance (p) of differences between mean anatomical parameters of needles in the control and treatment sample plots was estimated by multiple comparisons of means by the nonparametric Kruskal–Wallis test. For statistical calculations, the Statistica 7.0 software was used. The significance (p) of differences between mean biochemical parameters of needles in the polluted and reference site area was estimated by two-sided t test. For statistical calculations, the Systat 10.0 software for Windows was used.
3 Results
Analysis of the anatomy of Scots pine needles from the unpolluted control area and an area affected by alkaline cement dust pollution showed that alkalisation of the environment had resulted in a decrease in the total cross-sectional area of needles, needle width and thickness and the mesophyll volume fraction but an increase in the volume of epidermis, vascular bundles and xylem (Table 1). Statistically significant (p < 0.05) differences from control were observed in the width of needle cross-sections (19%), their thickness (16%) and the areas of mesophyll (15%) and epidermis (21%). The decrease of mesophyll was supported by biochemical analyses of the content of pigments participating in photosynthesis and of the content of lignin. The total concentration of chlorophyll was by 16% lower in the pines in the alkalizsed area if compared to the control (Table 2).
Histochemical colour reaction showed stronger staining of needle lignified tissues from the polluted area than in the slide mount from the control area (Fig. 2). Measurements of tissues revealed a 21% larger area of lignifying tissues in the alkalised area compared to the control. To check these results, a biochemical analysis was made, which supported the above-presented findings and showed that the average lignin content in the needles of pines growing in the alkalised area was by 12% larger than the respective data for the control pines (Table 2).

Anatomy of Scots pine needles from the unpolluted (a, c) and polluted area (b, d). General view of 2-year-old needle cross-section at ×100 magnification (a, b) and vascular bundles of needles at ×400 magnification (c, d). Lignified cell walls were stained red. Lignified tissues are more intensively stained in the needles of pines growing in the polluted area
The differences in the anatomical parameters of Scots pine needles between the polluted and unpolluted area depended significantly (p < 0.05) on the age of the needles (Fig. 3). While the total area of needle cross-sectional area and needle width and thickness of pines growing in the control area were greater and increased proportionally with the age of needles. The lowest values were observed in the youngest needles. In the area influenced by the cement plant, the values of these indicators were lower if compared to the control. In the case of younger needles, no statistically significant differences from the control were observed, but for 3-year-old needles, these indicators were lower (total area by 23%, width by 24% and thickness by 27% smaller) than control statistically differed at p < 0.05 level. While in the control area, the vascular bundles and xylem areas did not depend on the age of needles; then in the polluted sites, the area of vascular bundles of 3-year-old needles was larger than that of the youngest needles. The area of xylem showed only a tendency to increase with age.

Mean anatomical parameters (±SE; n = 45) of needles depending on age and location of the sampling site
The proportion of the area of lignified tissues of the total area of cross-sections was estimated 14, 13 and 14% for 1-, 2- and 3 year-old needles of control trees, and in the alkalised area, the proportion of lignified tissues was respectively 16%, 16% and 13%. Although the area of lignified tissues decreased in the 3-year-old needles from the polluted area, in older tissues, the content of lignin analysed by biochemical methods increased (Table 2), which is characteristic of the ageing of plant organisms (Miidla 1989; Polle et al. 1997). Visual study of cross-sections showed that, in 1-year-old needles, all the tissues participating in lignification became stained, but in the older needles, the staining was more intensive (Fig. 4). In addition, the width and thickness of needles and the areas of the epidermis and mesophyll varied between needles of different age. Differently from the needles from the control area, in the 3-year-old needles from the polluted area, these parameters decreased significantly (p < 0.05; Fig. 3). However, with increasing age of needles, the areas of the vascular bundle and xylem increased (both absolutely and as the percentage) compared to needles of the same age from the control area.

Anatomy and lignification of needles of different age. General view of needle cross-section at ×100 magnification (a 1 year, b 2 years, c 3 years) and vascular bundles of needles (d 1 years, e 2 years, f 3 years). Lignified cell walls were stained red (xylem, epidermis) and other tissue (mesophyll) green
4 Discussion
The influence of the alkalisation of the environment resulted in several changes in Scots pine anatomical and biochemical characteristics. Therefore, on the territory influenced by the cement plant, alkalisation may have decreased the growth of top and lateral shoots and radial increment but may have stimulated the proportion of heartwood, precocious maturation and ageing of the stand (Mandre et al. 2008). The growth of conifers under extreme growth conditions is thought to be associated with changes not only in their physiological–biochemical state but also in the anatomy and ratios between volumes of different tissues in needles. In the most alkaline area, a decrease in needle thickness and width, leading to a lower needle cross-sectional area, was established. Lower needle cross-section area could further result in lower needle dry mass, nitrogen content per unit area and lower needle-area-based photosynthetic capacity (Han et al. 2003; Warren et al. 2003). It was shown by Niinemets (1999) that, in many species, the volume fraction of epidermis decreases with increasing leaf thickness, resulting in a greater mesophyll volume fraction and photosynthetic capacity. In the present study, contrary trends to findings of Niinemets (1999) were observed. We found that alkalisation of the environment brought about an increase in the area of epidermis and a decrease in the needle thickness and mesophyll area. Consequently, the needle photosynthetic capacity could be lower in relatively extreme growth conditions than in optimal growth conditions. Therefore, biochemical analyses of pigments participating in photosynthesis showed a significantly lower concentration (p < 0.05) in needles. Furthermore, there are several external and internal factors affecting the biosynthesis of pigments and mesophyll in needles. The relationship between chlorophyll and nitrogen content was found to be the strongest in alkalised growth conditions (r = 0.66; p = 0.001; Mandre and Tuulmets 1995, 1997). However, the concentrations of Chl and N were by 20–23% lower on the alkaline territory than in the needles of Scots pine on the unpolluted area (Mandre and Tuulmets 1995). In addition, needle mesophyll cell volume fraction was shown to be strongly associated with the nitrogen content in leaves, whereas it decreased with increasing volume fraction of support tissues (sum of all tissue fractions with lignified cells; Niinemets et al. 2007). Comparison of lignin content of needles revealed that, with increasing area of lignified tissues, the content of lignin in needles increased in the alkalised environment. It is fairly clear that, in the case of nitrogen deficiency, which appeared in the alkalised areas, high lignin and tannin content may develop in plants (Mandre 2002; Mandre and Tuulmets 1995; Padu et al. 1989; Polle et al. 1997). As lignin incorporation renders plant cell walls mechanically rigid and stops their extension (Polle et al. 1997), the increased and premature lignification in alkalised growth conditions may be one of the reasons of the decrease in the total area of cross-sections, width and thickness of needles and inhibition of the length of needles (Ots 2002a). Hence, as the area of mesophyll decreases, its primary function changes from collecting photosynthates to transporting them from the needles to various sinks through vascular bundles.
A rapid increase of the area of epidermis was detected. Although it was stated by Lambers et al. (1998) that large epidermis cells are characteristic of fast growing species, our results did not support this statement. Moreover, the inhibition of dimensions of the leaves and biomass formation is often observed in alkalised territories (Farmer 1993; Lal and Ambasht 1982; Mandre and Korsjukov 2007; Mandre and Ots 1999; Manning 1971, 2001; Ots 2002a,b). The concentration of epicuticular waxes on pine needles in the alkalised area is about 12% higher than that in optimal growth conditions (Mandre and Bogdanov 2000). This suggests that, under stress, changes occur in the surface (outer) tissues and cells that decrease water evaporation through the cuticle to strengthen protection (Turunen and Huttunen 1990) against the harmful effects of UV radiation (Gordon et al. 1998) and phytopathogens (Neinhuis and Barthlott 1997).
Differences have been observed in the amount of chlorophyll and the intensity of photosynthesis depending not only on needle age but also on tree age. Ladanova and Tuzilkina (1992) found that the amount of chlorophyll increases in the needles of Picea obovata L. until the third to fourth year of age, but the intensity of photosynthesis is the highest in mature 1-year-old needles. Comparison of needle anatomy depending on tree age established great differences in needle width, thickness and cross-sectional area (Lin et al. 2002) and mesophyll volume fraction (Apple et al. 2002; Kovalev 1980). Compared with needles of old-growth Douglas fir trees, needles of saplings are longer and have proportionately smaller vascular cylinders, large resin canals and few hypodermal cells (Apple et al. 2002). Astrosclereids, which sequester lignin in their secondary cell walls and occupy space otherwise filled by photosynthetic cells, are scarce in young Douglas fir tree needles but abundant in needles of old-growth trees (Apple et al. 2002). The spatial and temporal lignification and regulation of the quantity and composition of lignin in plants depends not only on the peculiarities of species and on functions of organs but also on the development stage of organs (Boudet 2000). In our study, only 80-year-old trees were used, so we can conclude that differences in the measured parameters were due to age of needles and growth conditions.
Our results showed that the anatomy of needles as well as their biochemical composition depended significantly on needle age. Therefore, the greatest anatomical (total cross-sectional area, needle width and thickness, area of mesophyll and the vascular bundle) and biochemical (chlorophylls, lignin) differences between the needles from trees growing unpolluted conditions and in the alkalised area were observed in the oldest needles. When under optimum conditions anatomical parameters of trees had the highest values in 3-year-old needles, then under alkaline conditions, the maximum values were observed already in 2-year-old needles. This suggests that, under extreme conditions, the physiological ageing of needles may be faster than under optimum conditions. Biochemical studies conducted in the influence zone of the cement plant with spruces in 2000 also revealed more intensive physiological ageing of conifers in the alkaline environment than in unpolluted territory (Mandre and Korsjukov 2000; Mandre et al. 2001). The suggestion was based on the results showing a rapid increase of Ca concentration and lignification of needles, decrease of N (Mandre 2002) and carbohydrates (Mandre 1995) in Norway spruce needles in alkalised territory. Marschner (2002) reported that translocation of N and carbohydrates from older organs to younger ones are typical of the ageing process of plants. Based on the fact that, in the alkaline region, the level of starch and soluble sugars in needles is on average by 20–30% lower than the control (Mandre 1995) and on results of the present study, we concluded that the ageing process of Scots pine might be faster in the polluted area than in the control area. However, several authors have described a shorter lifespan of needles under favourable conditions (Niinemets and Lukjanova 2003; Ziegler 1997). In the course of this study, smaller numbers of 3-year-old and older needles were found in conifers growing in an unpolluted area, which agrees with earlier findings (Ots 2002b). At the same time, in pines growing in the vicinity of the cement plant, up to 5-year-old needles were observed. A noteworthy finding was a decreased area of lignified tissues in the oldest, 3-year-old needles, in the alkalised environment. As it was ascertained that, in the youngest tissues, lignin accumulates as a loose layer, whereas in the oldest tissues the lignin structure becomes denser and accumulation more compact, then the smaller area of lignified tissues in the oldest needles associates with structural changes of lignin under stress (Boudet et al. 1995; Hanisch and Kilz 1990), but the actual amount of lignin in these needles is significantly larger than in the youngest needles.
Although, in pines growing under optimum conditions, needle width, thickness and total cross-sectional area increase proportionally with needle ageing, reaching maximum levels in 3-year-old needles, the areas of vascular bundles and xylem decrease in them. In the alkalised environment, on the contrary, the vascular bundles of 3-year-old needles form a larger amount of conducting tissue for the functioning of needles in translocation nutrients and water and for preserving reserves under stress.
5 Conclusions
In industrial areas where alkaline dust pollutants are emitted, large amounts of solid components cause changes in the soil chemistry and pH. However, the dependence of conifer needle anatomy on the alkalisation of the environment is still a matter of controversy. Differences in anatomical parameters between trees in polluted and control areas were found, and variation in mesophyll and lignified volume fractions was statistically established.
In addition, changes in biochemical characteristics (lignin and chlorophyll) were detected. Responses of Pinus sylvestris physiology and anatomy to alkaline stress are not finally understood. As our results did not support all standpoints in the literature, additional measurements are needed to test the dependence of the general anatomical variability of conifer needles on the peculiarities of growth conditions.
References
Apple, M., Tiekotter, K., Snow, M., Young, J., Soeldner, A., Phillips, D., et al. (2002). Needle anatomy changes with increasing tree age in Douglas-fir. Tree Physiology, 22(2–3), 129–136.
Arasimovich, V. V., & Ermakov, A. I. (1987). Opredelenie polisakharidov i lignina (Measurement of polysaccharides and lignin). In A. I. Ermakov (Ed.), Methods for biochemical studies of plants (Metody biokhimicheskogo issledovaniya rasteniy) (pp. 143–172). Leningrad: Agropromizdat. (in Russian).
Borka, G. (1983). Wirkung von Zementstaub auf Winterweizen; Entwicklung–Wachstum–Stoffwechsel–Ertrag. Aquilo, Serie Botanica, 19, 233–239.
Boudet, A. M. (2000). Lignins and lignification: selected issues. Plant Physiology and Biochemistry, 38(1–2), 81–96. doi:10.1016/S0981-9428(00)00166-2.
Boudet, A. M., Lapierre, C., & Grima-Pettenati, J. (1995). Biochemistry and molecular biology of lignification. The New Phytologist, 129(2), 203–236. doi:10.1111/j.1469-8137.1995.tb04292.x.
Bozzola, J. J., & Russell, L. D. (1992). Electron microscopy: Principles and techniques for biologists. Sudbury: Jones and Bartlett.
Environmental Review No. 16. (2007). Kunda: Kunda Nordic Heidelberg Cement Group.
Farmer, A. M. (1993). The effects of dust on vegetation—A review. Environmental Pollution, 79(1), 63–75. doi:10.1016/0269-7491(93)90179-R.
Gordon, D. C., Percy, K. E., & Riding, R. T. (1998). Effects of u.v.-B radiation on epicuticular wax production and chemical composition of four Picea species. The New Phytologist, 138(3), 441–449. doi:10.1046/j.1469-8137.1998.00116.x.
Han, Q., Kawasaki, T., Katahata, S., Mukai, Y., & Chiba, Y. (2003). Horizontal and vertical variations in photosynthetic capacity in a Pinus densiflora crown in relation to leaf nitrogen allocation and acclimation to irradiance. Tree Physiology, 23(12), 851–857.
Hanisch, B., & Kilz, E. (1990). Monitoring of forest damage. Spruce and pine. London: Helm.
Klõšeiko, J. (2005). Concentration of carbohydrates in conifer needles near Kunda cement plant, Estonia, nine years after reduced dust pollution. Metsanduslikud Uurimused/Forestry Studies, 42, 87–94.
Kovalev, A. G. (1980). Vozrast dereva i anatomo-morphologicheskoe srtoenie khvoi sosny obyknovennoy (The age of a tree and anatomo-morphological structure of needles in Pinus sylvestris L.). Lesovedenie, 6, 30–35. in Russian with English abstract.
Ladanova, N. V., & Tuzilkina, V. V. (1992). Vozrastnye izmeneniya struktury i funktsii assimilyatsionnogo apparata eli sibirskoy (The age changes of structure and function of the assimilating organs of Siberian spruce). In V. V. Gabukova & E. S. Chavchavadze (Eds.), Anatomiya, fisiologiya i ekologiya lesnykh rasteniy (Anatomy, physiology and ecology of forest plants) (pp. 90–92). Petrozavodsk: Karelian Research Center of RAS. (in Russian).
Lal, B., & Ambasht, R. S. (1982). Impact of cement dust on the mineral and energy concentration of Psidium guayava. Environmental Pollution, 29A(4), 241–247.
Lambers, H., Chapin, F. S., III, & Pons, T. L. (1998). Plant physiological ecology. New York: Springer.
Liblik, V., Pensa, M., & Kundel, H. (2000). Temporal changes in atmospheric air pollution in industrial areas of Ida- and Lääne-Viru counties. Metsanduslikud Uurimused/Forestry Studies, 33, 17–36.
Lichtenthaler, H. K., & Wellburn, A. R. (1983). Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochemical Society Transactions, 11(5), 591–592.
Lin, J., Sampson, D. A., Deckmyn, G., & Ceulemans, R. (2002). Significant overestimation of needle surface area estimated based on needle dimensions in Scots pine (Pinus sylvestris). Canadian Journal of Botany, 80(9), 927–932. doi:10.1139/b02-081.
Lukjanova, A., & Mandre, M. (2006). Anatomical features and localization of lignin in needles of Scots pine (Pinus sylvestris L.) on dunes in South-West Estonia. Biology, Ecology, 55(2), 173–184.
Mandre, M. (1995). Dust emission and deposition. In M. Mandre (Ed.), Dust pollution and forest ecosystems. A study of conifers in an alkalized environment. Publication 3 (pp. 18–22). Tallinn: Institute of Ecology.
Mandre, M. (2002). Relationships between lignin and nutrients in Picea abies L. under alkaline air pollution. Water, Air, and Soil Pollution, 133(1–4), 361–377.
Mandre, M., & Bogdanov, V. (2000). Epicuticular waxes on the needles of Norway spruce and Scots pine in the area of the Kunda cement plant. Metsanduslikud Uurimused/Forestry Studies, 33, 137–142.
Mandre, M., & Korsjukov, R. (2000). Content of pigments in the needles of Norway spruce and Scots pine in an area of cement production. Metsanduslikud Uurimused/Forestry Studies, 33, 121–128.
Mandre, M., & Korsjukov, R. (2007). The quality of stemwood of Pinus sylvestris in an alkalised environment. Water, Air, and Soil Pollution, 182(1–4), 163–172. doi:10.1007/s11270-006-9329-1.
Mandre, M., & Ots, K. (1995). The height growth of trees. In M. Mandre (Ed.), Dust pollution and forest ecosystems. A study of conifers in an alkalized environment. Publication 3 (pp. 117–118). Tallinn: Institute of Ecology.
Mandre, M., & Ots, K. (1999). Growth and biomass partitioning of 6-year-old spruces under alkaline dust impact. Water, Air, and Soil Pollution, 114(1–2), 13–25. doi:10.1023/A:1005048921852.
Mandre, M., & Tuulmets, L. (1995). Changes in the pigment system. In M. Mandre (Ed.), Dust pollution and forest ecosystems. A study of conifers in an alkalized environment. Publication 3 (pp. 66–77). Tallinn: Institute of Ecology.
Mandre, M., & Tuulmets, L. (1997). Pigment changes in Norway spruce induced by dust pollution. Water, Air, and Soil Pollution, 94(3–4), 247–258. doi:10.1007/BF02406061.
Mandre, M., Rauk, J., & Ots, K. (1995). Needle and shoot growth. In M. Mandre (Ed.), Dust pollution and forest ecosystems. A study of conifers in an alkalized environment. Publication 3 (pp. 103–111). Tallinn: Institute of Ecology.
Mandre, M., Korsjukov, R., & Bogdanov, V. (2001). Okaspuude okaste vananemisega seotud biokeemilistest muutustest (Age related changes in the biochemical composition of conifer needles). Metsanduslikud Uurimused/Forestry Studies, 35, 19–26.
Mandre, M., Kask, R., Pikk, J., & Ots, K. (2008). Assessment of growth and stemwood quality of Scots pine on territory influenced by alkaline industrial dust. Environmental Monitoring and Assessment, 138(1–3), 51–63. doi:10.1007/s10661-007-9790-3.
Manning, W. J. (1971). Effects of limestone dust on leaf condition, foliar disease incidence, and leaf surface microflora of native plants. Environmental Pollution, 2(1), 69–76. doi:10.1016/0013-9327(71)90038-3.
Manning, W. J. (2001). Air pollution and forest health: establishing cause and effect in the forest. TheScientificWorldJournal, 1, 391–392. doi:10.1100/tsw.2001.73.
Marschner, H. (2002). Mineral nutrition of higher plants. London: Academic.
Miidla, H. (1989). Biochemistry of lignin formation. The formation of lignin in wheat plants and its connection with mineral nutrition. Publications in plant physiology and plant biochemistry, 5. Acta et Commentationes Universitatis Tartuensis, 845, 11–23).
Monties, B. (1989). Lignins. In P. M. Dey & J. B. Harborne (Series Eds.), Methods in plant biochemistry. In J. B. Harborne (Ed.), Plant phenolics (vol. 1, pp. 113–157). London: Academic.
Neinhuis, C., & Barthlott, W. (1997). The tree leaf surface: structure and function. In H. Rennenberg, W. Eschrich & H. Ziegler (Eds.), Trees—contributions to modern tree physiology (pp. 3–19). Leiden: Backhuys.
Niinemets, Ü. (1999). Components of leaf dry mass per area—thickness and density—alter leaf photosynthetic capacity in reverse directions in woody plants. The New Phytologist, 144(1), 35–47. doi:10.1046/j.1469-8137.1999.00466.x.
Niinemets, Ü., & Lukjanova, A. (2003). Needle longevity, soot growth and branching frequency in relation to site fertility and within-canopy light conditions in Pinus sylvestris. Annals of Forest Science, 60(3), 195–208. doi:10.1051/forest:2003012.
Niinemets, Ü., Lukjanova, A., Turnbull, M. H., & Sparrow, A. D. (2007). Plasticity in mesophyll volume fraction modulates light-acclimation in needle photosynthesis in two pines. Tree Physiology, 27(8), 1137–1151.
Ots, K. (2000). Morphometric parameters of conifer needles and shoots in the areas near the Kunda cement plant. Metsanduslikud Uurimused/Forestry Studies, 33, 158–176.
Ots, K. (2002a). Impact of air pollution on the growth of conifers in the industrial region of Northeast Estonia. Doctoral thesis. Tartu: Estonian Agricultural University.
Ots, K. (2002a). The length growth of needles of Scots pine in the industrial area of Northeast Estonia. Metsanduslikud Uurimused/Forestry Studies, 36, 39–46.
Ots, K. (2005). Changes in the density of needles on Scots pine shoots in the area polluted by Kunda cement plant. Metsanduslikud Uurimused/Forestry Studies, 42, 79–86.
Ots, K., & Rauk, J. (2000). Increment of conifers and its air pollution-related tendencies in Ida-Viru and Lääne-Viru counties. Metsanduslikud Uurimused/Forestry Studies, 33, 87–97.
Padu, E., Meiner, L., & Selgis, R.(1989). The activity of L-phenylalanine ammonia-lyase and peroxidase, and the biosynthesis of phenolic compounds in wheat under different condition of mineral nutrition. The Formation of lignin in wheat plants and its connection with mineral nutrition. Publications in plant physiology and plant biochemistry, 5. Acta et commentationes Universitatis Tartuensis, 845, 85–108.
Polle, A., Otter, T., & Sandermann, J. (1997). Biochemistry and physiology of lignin synthesis. In H. Rennenberg, W. Eschrich & H. Ziegler (Eds.), Trees—contributions to modern tree physiology (pp. 455–475). Leiden: Backhuys.
Rauk, J. (1995). Bioproductivity. In M. Mandre (Ed.), Dust pollution and forest ecosystems. A study of conifers in an alkalized environment. Publication 3 (pp. 119–123). Tallinn: Institute of Ecology.
Ruzin, S. E. (1999). Plant microtechnique and microscopy. New York: Oxford University Press.
Turunen, M., & Huttunen, S. (1990). A review of the response of epicuticular wax of conifer needles to air pollution. Journal of Environmental Quality, 19(1), 35–45.
Van Soest, P. J. (1987). Nutritional ecology of the ruminant. Ruminant metabolism, nutritional strategy, the cellulytic fermentation and the chemistry of forages and plant fibers. Ithaca: Cornell University Press.
Vernon, L. P. (1960). Spectrophotometric determination of chlorophylls and pheophytins in plant extracts. Analytical Chemistry, 32(9), 1144–1150. doi:10.1021/ac60165a029.
Warren, C. R., Dreyer, E., & Adams, M. A. (2003). Photosynthesis–Rubisco relationships in foliage of Pinus sylvestris in response to nitrogen supply and the proposed role of Rubisco and amino acids as nitrogen stores. Trees. Structure and Function, 17, 359–366.
Ziegler, H. (1997). Some open questions in tree physiology. In H. Rennenberg, W. Eschrich & H. Ziegler (Eds.), Trees—contributions to modern tree physiology (pp. 531–544). Leiden: Backhuys.
Acknowledgements
This research was supported by the Estonian Science Foundation (grant no. 6022) and by the Estonian Ministry of Education and Research (project no. 0170021s08).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Lukjanova, A., Mandre, M. Effects of Alkalization of the Environment on the Anatomy of Scots Pine (Pinus sylvestris) Needles. Water Air Soil Pollut 206, 13–22 (2010). https://doi.org/10.1007/s11270-009-0080-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11270-009-0080-2