
Abstract
Three cruises were carried out in Jiaozhou Bay (JZB) in the neap tide in October 2002 (fall) and in both neap and spring tides in May 2003 (spring) to understand the relative importance of external nutrient inputs versus physical transport and internal biogeochemical processes. Nutrients (\( NO_{3} ^{ - } \), \( NO_{2} ^{ - } \), \( NH_{4} ^{ + } \), \( PO_{4} ^{{3 - }} \), silicic acid, total dissolved nitrogen (TDN) and phosphorus (TDP), dissolved organic nitrogen (DON) and phosphorus (DOP)) were measured. The concentrations of nutrients were higher in the northern part than in the southern part. High concentrations of \( NH_{4} ^{ + } \) and DON in JZB demonstrated the anthropogenic input. Ambient nutrient ratios indicated that the potential limiting nutrients for phytoplankton growth were silicon, and then phosphorus. Nutrients showed an obvious tidal effect with low values at flood tide and high values at ebb tide. Nutrient elements were transported into JZB in the north and output in the south (i.e., into the Yellow Sea), which varied with season, tidal cycle and investigation sites. Water exchange between JZB and the Yellow Sea exports \( NO_{3} ^{ - } \), \( NH_{4} ^{ + } \) and DON out of JZB, while it inputs \( PO_{4} ^{{3 - }} \), silicic acid and DOP into JZB. Nutrient budgets demonstrate that riverine input and wastewater discharge are major sources of nutrients, while residual flow is of minor importance in JZB ecosystem. JZB is a sink for the nutrient elements we studied except for DON. Stoichiometric calculations demonstrate that JZB is a net autotrophic system.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Global nutrient cycles have been greatly altered by land-use changes resulting from human disturbance over the last century (Downing et al. 1999). Excessive nutrient discharges and changes in their relative concentrations have been known to result in eutrophication, hence modifying aquatic food webs and causing severe hypoxic events in coastal environments (Turner and Rabalais 1994; Humborg et al. 1997; Ragueneau et al. 2002; Turner 2002; Piehler et al. 2004). Water exchange between coastal and offshore waters can also be a very important factor affecting coastal ecosystems (Aubry and Acri 2004; Mackas and Harrison 1997; Su et al. 2004).
Jiaozhou Bay (JZB) is a typical semi-enclosed water body with a narrow channel (∼2.5 km) connected with the Yellow Sea, with a surface area of 390 km2 and an average depth of 6–7 m. JZB is characterized by semi-diurnal tides with an average tidal range of 2.7–3.0 m and a maximum of 5.1 m (Zhang 2007). Strong tidal turbulent mixing in JZB results in nearly homogeneous vertical profiles of temperature and salinity with a weak stratification only in the summer when the land-source input reaches its maximum (Liu et al. 2004a). About 10 streams empty seasonally into the bay with various amounts of freshwater and sediment loads, including the most important freshwater sources: Yanghe and Daguhe. Most of these rivers, however, have become conduits of industrial and domestic waste discharges along with the economic development and population growth in the region. JZB has been greatly disturbed by human activities, leading to increased nutrient levels in dissolved inorganic nitrogen (DIN) and \( PO_{4} ^{{3 - }} \), but a decrease in Si(OH)4 with higher DIN/ \( PO_{4} ^{{3 - }} \) and lower Si(OH)4/DIN atomic ratios compared to four decades ago (Shen 2001; Liu et al. 2005a). Frequent red tide events have occurred in JZB, for example, Mesodinium rubrum bloom in June 1990 (Sun et al. 1993), Skeletonema costatum and Biddulphia awita bloom in July 1998 (Hao et al. 2000), and Eucampia zoodiacus bloom in July 1999 (Lu et al. 2001; Huo et al. 2001).
This study was aimed at discriminating the relative importance of external nutrient inputs versus physical transport and biogeochemical processes based on the Land – Ocean Interactions in the Coastal Zone (LOICZ) Biogeochemical Modeling Guidelines (cf. Gordon et al. 1996), and the dynamic effects (e.g., tide) regulating the nutrient distribution in JZB.
2 Materials and Methods
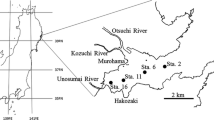
The field observations of this study were carried out in JZB during October 13–15, 2002 (neap tide), May 9–10, 2003 (neap tide), and May 19–20, 2003 (spring tide), respectively. In each of the three cruises, 14 stations were visited and an anchor station (D4-2) in 2002 and two anchor stations (D4-1 and D4-3) during both the neap and spring tides in May 2003 were occupied for over 25 h (Fig. 1). Profiles of temperature, salinity and velocity were measured at grid stations and hourly at the anchor stations during the cruises. Water samples were collected with 5-l Niskin bottles at the grid stations, and every 3 h at anchor station D4-2 in October 2002, and every hour at anchor stations D4-1 and D4-3 in both the neap and spring tides in May 2003. After collection, samples were filtered immediately through pre-cleaned 0.45 μm pore-size acetate cellulose filters, and the filtrates were preserved with the addition of HgCl2. The filters were dried at 50°C and weighed to determine suspended particulate matter (SPM), biogenic silica (BSi), particulate inorganic phosphorus (PIP) and particulate organic phosphorus (POP). Samples for POC/N were filtered through 47 mm GF/F filters that had been pre-combusted at 400°C overnight.

Map of Jiaozhou Bay, which shows grid stations (●), anchor station D4-2 in October 2002 (△), anchor stations D4-1 and D4-3 in May 2003 (⊕), rain water sampling station Fulongshan (star), Haibohe (HB), Licunhe (LC) and Tuandao (TD) sewage treatment plants (▲) and the major streams surrounding Jiaozhou Bay
River water sampling was undertaken in both April (dry season) and August (flood season) 2004. The information about drainage area, water and sediment loads of rivers from the catchment areas of JZB has been reported elsewhere (Liu et al. 2005a). Water samples were collected with 2-l polyethylene bottles attached to a fiberglass reinforced fishing pole in the river. Samples were filtered through pre-cleaned 0.45 μm filters and the filtrates were preserved with the addition of HgCl2.
Nutrients were determined using an Auto-Analyzer Skalar SANplus giving a precision of <3%. The total dissolved nitrogen (TDN) and total dissolved phosphorus (TDP) were measured according to the methods of Grasshoff et al. (1999). The concentration of dissolved inorganic nitrogen (DIN) was the sum of \( NO_{3} ^{ - } \), \( NO_{2} ^{ - } \) and \( NH_{4} ^{ + } \). The concentrations of dissolved organic nitrogen (DON) and phosphorus (DOP) were the differences between TDN and DIN, and between TDP and \( PO_{4} ^{{3 - }} \), respectively.
The biogenic silicon (BSi) in the SPM was determined by leaching with 5% Na2CO3 at 100°C for 100 min. Silicic acid in the leaching solution was analyzed by molybdate-blue spectrophotometry (Mortlock and Froelich 1989), and dissolved aluminum was measured by fluorometry (Ren et al. 2001) for mineral correction (Kamatani and Takano 1984). The coefficient of variability (CV) for BSi measurement in the SPM was better than 10% (Liu et al. 2005b).
Inorganic P (PIP) in the SPM was measured by 1 M HCl extraction (25°C for 24 h), and total P (PTP) was measured by 1 M HCl extraction after ignition of the particulates (550°C for 2 h). The organic P (POP) was determined by the difference between PTP and PIP (Liu et al. 2004c). The extracted \( PO_{4} ^{{3 - }} \) was measured by spectrophotometry (Liu et al. 2003b). The analysis of the Chinese standard of coastal sediment (GBW 07314) gave the PTP concentrations of 19.68 ± 0.08 μmol g−1, which compared well with the certified value (20.85 ± 1.97 μmol g−1). The analytical precision for the P extractions was better than 0.1% for PIP, 17% for POP and 0.5% for PTP (Liu et al. 2004c).
Particulate organic carbon (POC) and nitrogen (PN) were measured using a CHNOS Elemental Analyzer (Model: Vario EL III, Elementar Analysensysteme GmbH) after removal of carbonate. The precision was <5–10%, estimated by repeated analyses (Liu et al. 2005b).
3 Results
3.1 Spatial Distribution of Nutrients in Jiaozhou Bay
In October 2002, the concentrations of dissolved nutrients were higher in the northern than in the southern parts of JZB (Fig. 2). The concentrations of nutrients showed a vertically well-mixed distribution (Table 1). During both the neap and spring tides in May 2003, dissolved nutrients showed higher values in the northern part than in the southern part of JZB (Fig. 3). Vertically, nutrients were slightly higher in the surface than in the near-bottom waters (Table 1).

Nutrients of Jiaozhou Bay in October 2002 (μM). To reduce the number of figures, only the distribution of various nutrient species in surface waters (1 m) is shown

Nutrients of Jiaozhou Bay on 10 May 2003 (neap tide) and 20 May 2003 (spring tide) (μM). It shows the distribution of various nutrient species in surface water (1 m). To reduce the number of figures, the arithmetic mean of two station grids is given in the figure
When comparing the average concentrations of dissolved nutrients between the neap and spring tides in May 2003, it is obvious that nutrients in the surface waters were higher in the spring tide than in the neap tide, except that concentrations of DON and DOP were similar (Table 1). However, comparing nutrient levels at neap tide between 2002 and 2003, the average concentrations of silicic acid and DON were higher in 2002 than in 2003, while the average concentrations of \( NO_{3} ^{ - } \) and \( NH_{4} ^{ + } \) were higher in 2003 than in 2002, and the other nutrient species showed similar distributions (Table 1).
The distributions of PIP, POP, BSi, POC and PN were similar to the dissolved nutrient elements, with higher values in the northern part and lower values in the southern part (plots not shown). A comparison of the average concentrations of particulate elements between the neap and spring tides in May 2003 demonstrated that particulate nutrients were similar in the neap and spring tides, while comparison of the particulate nutrients between 2002 and 2003 indicates that the concentrations were 4-fold higher in fall 2002 than in spring 2003 for BSi, 2-fold higher for POC and PN, and were similar for POP, while the concentrations of PIP were 1.5–2.8 times lower in 2002 than in 2003 (Table 2).
3.2 Variability of Nutrients During the Spring and Neap Tidal Cycles
In October 2002, nutrients showed variations of 2- to 3-fold for \( NO_{3} ^{ - } \), \( NO_{2} ^{ - } \) and \( NH_{4} ^{ + } \), and 1- to 2-fold for \( PO_{4} ^{{3 - }} \), silicic acid, TDN, TDP and DON. Again, the concentrations of nutrients showed a vertically well-mixed distribution (Fig. 4).

Concentrations of nutrient species (μM) at anchor station D4-2 in October 2002, in which changes in tidal elevation, i.e., flood and ebb tides, are shown at the top
During the neap tide of May 2003, the concentrations of nutrients at anchor station D4-1 varied 1- to 2-fold, except for DON, which varied 3- to 4-fold, while the change in nutrient concentrations increased at station D4-3 by 2- to 3-fold for \( NO_{3} ^{ - } \), silicic acid, TDN and DON, and 3- to 8-fold for \( NO_{2} ^{ - } \), \( NH_{4} ^{ + } \), \( PO_{4} ^{{3 - }} \) and TDP. The concentrations of nutrients were comparable at stations D4-1 and D4-3 (Fig. 5a). During the spring tide in May 2003, nutrients varied by 1- to 2-fold for \( NO_{3} ^{ - } \), \( NO_{2} ^{ - } \), \( NH_{4} ^{ + } \), silicic acid and TDN, and 3- to 7-fold for \( PO_{4} ^{{3 - }} \), TDP and DON at anchor station D4-1, while variations of nutrient levels increased at station D4-3 by 1- to 2-fold for silicic acid and TDN, 2- to 5-fold for \( NO_{2} ^{ - } \), \( NH_{4} ^{ + } \) and \( PO_{4} ^{{3 - }} \), 6- to 7-fold for \( NO_{3} ^{ - } \), 4- to 6-fold for DON, and 5- to 9-fold for TDP. The concentrations of nutrients were also similar at stations D4-1 and D4-3 (Fig. 5b).

Concentrations of nutrient species (μM) at anchor stations D4-1 and D4-3 on 9 May (a) and 19 May (b) 2003, in which changes in tidal elevation are shown at the top
The concentrations of nutrients showed a vertically well-mixed distribution with slightly higher values in the surface than in the near-bottom waters during both spring and neap tides, suggesting the nutrients came from land sources, as previously reported by Liu et al. (2005a) based on nutrient budgets. Comparison of nutrient levels between the neap and spring tides in 2003 indicated that concentrations of \( NO_{3} ^{ - } \), \( NO_{2} ^{ - } \), DON and \( NH_{4} ^{ + } \) were higher during the neap tide than during the spring tide, with other nutrients being similar. Examination of the difference of nutrient species in neap tide between 2002 and 2003 revealed that concentrations of \( NO_{3} ^{ - } \) and \( NH_{4} ^{ + } \) increased by 2-fold in 2003 relative to the concentrations in 2002. The TDN level was higher in 2003 than in 2002, while the concentrations of silicic acid decreased by 2-fold in 2003 relative to 2002 because the freshwater discharge was lower in May 2003 than October 2002. The other nutrient species showed similar values.
The concentrations of nutrients at anchor stations showed the influence of tidal effects, with low values during the flood tide and an increase during the ebb tide (Figs. 4 and 5). The tidal effects were not significant at stations D4-2 in 2002 or at D4-1 during both the neap and spring tides in 2003. However, the tidal effects were significant, with nutrient levels increasing from the flood to ebb tides at station D4-3 during both the neap and spring tidal cycles in 2003.
For anchor station investigations, particulate nutrient elements were only measured at anchor station D4-2 in October 2002, and nutrients varied by 2- to 3-fold for BSi, POC and PN, and 20- to 50-fold for PIP and POP. Concentrations of particulate nutrients showed a vertically well-mixed distribution (not shown). The tidal effects were not significant in October 2002 (Fig. 6), similar to dissolved nutrient elements.

Concentrations of BSi, PIP and POP (μM), PN and POC (%) at anchor station D4-2 in October 2002, in which changes in tidal elevation, i.e., flood and ebb tides, are shown at the top
3.3 Riverine Input
Table 3 provides recent data on nutrients in the main streams that discharge into JZB. Nutrient concentrations varied considerably, depending on the specific streams (e.g., Licunhe and Yanghe) and nutrients. The concentrations of nutrients in the rivers between the dry and flood seasons varied by a factor of up to 50 for DON, 13 for \( NO_{3} ^{ - } \), 26 for \( NO_{2} ^{ - } \), and 3–5 times for \( NH_{4} ^{ + } \), \( PO_{4} ^{{3 - }} \), silicic acid and DOP. Compared to the nutrient levels in major rivers in 2002 (Liu et al. 2005a), the concentrations of nutrients changed considerably. For example, concentrations of \( NO_{3} ^{ - } \) were seven times higher in the Daguhe and two times lower in the Yanghe in the dry season from 2002 to 2004.
Riverine input of nutrients can be estimated by the annual production of total freshwater discharge and nutrient concentrations. The rivers are characterized by seasonal freshwater discharge and anthropogenic disturbance, which may introduce an uncertainty of 80% to the estimate of nutrient input (Table 4). This is similar to the 2002 investigations (Liu et al. 2005a). The nutrient fluxes into Jiaozhou Bay are mainly from Daguhe for \( NO_{3} ^{ - } \) (92%), \( NO_{2} ^{ - } \) (81%), DON (79%) and silicic acid (67%), and from Moshuihe for \( PO_{4} ^{{3 - }} \) (61%). The fluxes of \( NH_{4} ^{ + } \) into the bay are from Moshuihe (42%), Daguhe (35%) and Licunhe (16%). The fluxes of DOP are from Daguhe (48%) and Moshuihe (40%).
The yields of nutrients over the drainage basin of each seasonal river around JZB can be estimated by the produced nutrient concentration with long-term average discharge divided by the drainage area (Table 5). The yields of \( NO_{3} ^{ - } \) are higher in Yanghe (37%), Bashahe (31%) and Daguhe (24%). The yields of \( NH_{4} ^{ + } \), \( PO_{4} ^{{3 - }} \) and DON are higher in Licunhe (40–60%) and Moshuihe (30–40%). The yields of silicic acid are higher in the Yanghe, Moshuihe and Licunhe, measuring ∼30% for each river. The yields of DOP are higher in Moshuihe (52%) and Yanghe (26%). The resulting yields of nutrients vary considerably among the river catchments, depending upon the element of interest and water discharge.
4 Discussion
4.1 Nutrient Composition
The nutrient concentrations in JZB were higher in the north than the south in October 2002 and May 2003, indicating the effect of riverine inputs, wastewater discharge and tidal patterns. Nutrients were vertically well mixed in 2002, and were slightly higher in the surface than in near-bottom waters in 2003. Yang et al. (2004) reported that the SPM content in the upper water column of JZB was generally coarser than in the bed deposits and the lower part of the water column, which indicated that SPM was modulated differently by settling/resuspension near the seabed and by advection in the surface water. This may explain the pattern of vertical distribution of nutrients, taking into account the active partition of nutrients between the SPM and the solution (Liu et al. 2004b; Rysgaard et al. 1999). The terrestrial inputs exceeded benthic nutrient fluxes (Liu et al. 2005a). It has been reported that the phytoplankton community in JZB is mainly composed of diatoms and dinoflagellates, with diatoms accounting for more than 90% of the phytoplankton cell number in 2003–2004, with high diatom abundance in September and January (Li et al. 2005; Liu 2004; Sun et al. 2002). The higher concentrations of BSi, POC, PN and POP in 2002 than 2003 are related to phytoplankton abundance.
There was an obvious tidal effect, with low values at the flood tide and high values at the ebb tide, depending on the location. A slight tidal regulation of nutrients at stations D4-2 was observed in 2002 and D4-1 in 2003 and significant tidal effects were found at station D4-3 during both the spring and neap tides in 2003 due to flows of different water masses. A similar result has been found in the Chupa Estuary, where concentrations of nutrients showed considerable inter-tidal and spring-neap variability (Dale and Prego 2003). The concentrations of \( NO_{3} ^{ - } \), \( NO_{2} ^{ - } \), DON and \( NH_{4} ^{ + } \) were higher during the neap tide than during the spring tide in 2003, with other nutrients being similar. This is related to the fact that land-source transport is the major source for all nutrient species, and nitrogen species exit from JZB while the other species are imported into JZB (Liu et al. 2005a). The population in Qingdao increased from 4.6 × 106 in the 1960s to 7.0 × 106 in the 1990s (Yin and Lu 2000). The application of chemical fertilizer increased by three times from 1980 to 1997 (Qingdao Municipal Statistics Bureau 1998). The annual wastewater discharge into JZB increased from 70.2 × 106 tons of industrial waste discharge and 14.4 × 106 tons of domestic waste discharge in 1980 to 90.3 × 106 tons of industrial waste discharge and 137.2 × 106 tons of domestic waste discharge in 2002 (Qingdao Municipal Statistics Bureau 2003).
With respect to the composition of TDN, DIN accounted for ∼40% and DON represented ∼60% in October 2002, while DIN and DON were ∼60 and ∼40% respectively, in May 2003. With respect to DIN, \( NH_{4} ^{ + } \) and \( NO_{3} ^{ - } \) accounted for 60–70%, and ∼30%, respectively, in both 2002 and 2003. The \( NO_{2} ^{ - } \) percentage was low (<4–8%). \( PO_{4} ^{{3 - }} \) represented 70–80% of TDP. The ratios of the DIN percentage over TDN to the \( PO_{4} ^{{3 - }} \) percentage over TDP were 0.56 in 2002 and 0.82 in 2003, implying that anthropogenic activities transport more \( PO_{4} ^{{3 - }} \) than DIN or the regeneration rates of phosphorus are faster than nitrogen. The latter is more likely, as the riverine nutrient transport showed a ratio of the DIN percentage over TDN to the \( PO_{4} ^{3} \) percentage over TDP of 4.44 weighted by river discharge. The high percentage of DON over TDN indicates that nitrogen cycling is important in the ecosystem of JZB. Similar results have been reported in the Brantas River Estuary where organic nitrogen was found to play a more important role in coastal food webs and the nitrogen cycling than previously thought (Jennerjahn et al. 2004). High proportions of DON and DOP reported in the Yellow Sea were found to be important in the modeling of nutrient cycling and in explaining nutrient limitation in the ecosystem (Liu et al. 2003b). High concentrations of \( NH_{4} ^{ + } \) (i.e. 60–70% of DIN) indicated the input of wastewater discharge (as discussed below). Similar results were reported in the Ria Formosa Lagoon where significantly enriched nitrogen (\( NH_{4} ^{ + } \), \( NO_{2} ^{ - } \) and \( NO_{3} ^{ - } \)) with respect to the adjacent coastal waters indicated that inputs from sewage, agricultural runoff and benthic fluxes were not fully assimilated within the lagoon (Newtona and Mudge 2005). Intense shellfish farming also increases the nitrogen concentration within JZB by enhancing the deposition of organic matter to the sediment and promoting dissimilar processes of N cycling, similar to the situation in the Thau Lagoon in the Upper South Cove of Nova Scotia (Gilbert et al. 1997; Hatcher et al. 1994).
The atomic ratio of DIN to \( PO_{4} ^{{3 - }} \) ranged from 6 to 66, with an average of 20 in 2002, while the DIN/\( PO_{4} ^{{3 - }} \) ratios increased to ∼40 in 2003. The Si(OH)4/DIN atomic ratio was 0.3–2.1 with an average of 0.7 in 2002, but was 0.1–0.5 with an average of 0.2 in 2003. The Si(OH)4 to \( PO_{4} ^{{3 - }} \) atomic ratio was 5.8–16.6 with an average of 11.8 in 2002, and 1.2–14.6 with an average of 7.1 in 2003. It has been reported that diatoms represented 60–80% of phytoplankton species, while diatom abundance accounted for >90% of the phytoplankton biomass (Liu et al. 2003a; Liu 2004; Li et al. 2005). Nutrient elemental ratios indicated that the potential limiting nutrients for phytoplankton growth were silicon, and then phosphorus for diatoms. The possible limitation of the phytoplankton biomass by silicon in this region has been proposed based on ambient nutrient ratios (Zhang and Shen 1997; Yang 1999; Shen 2001; Liu et al. 2005a). Zou (2001) reported that the most limiting nutrient for phytoplankton growth in JZB was silicic acid, based on in situ nutrient addition incubation experiments. It has also been reported that in other aquatic systems of the world’s large rivers, the nutrient limitation of phytoplankton growth is shifting towards a higher incidence of P and Si limitation as a result of increased nitrogen loading with an N:P fertilizer use of 26:1 (molar basis) and relatively stable or decreased silicic acid loading, which may alter the phytoplankton community composition and compromise the structure of food webs (Turner et al. 2003).
The particulate nutrient ratios are provided in Table 2. The atomic ratios of BSi to POC were 0.08–0.80 in JZB. This value is within the range of 0.05–1 reported worldwide, with variations depending on the proportion of diatoms in phytoplankton, temperature, light, and trace metal concentrations (Ragueneau et al. 2000). In JZB, the ratios of POC to PN were lower than the Redfield ratio, but PN to POP ratios were higher than the Redfield ratio.
4.2 Nutrient Transport
Figure 7 shows the current vectors at anchor station D4-2 in 2002. The westward current was 1.7 times the eastward current, while the northward current was similar to the southward current. This indicates that water is transported into JZB from the Yellow Sea in the center of the bay mouth. Figure 8 shows the current vectors at stations D4-1 and D4-3 during the neap and spring tides in 2003. The currents were similar in the surface and near-bottom waters. At station D4-1, the current was mainly northward and westward during both the neap and spring tides. At station D4-3, the eastward and westward currents were similar and the northward current was ca. five times the southward flow during the neap tide, while the eastward current was 1.4 times the westward current and the southward current was similar to the northward one in spring tide. On average, the current flowed northward at both stations D4-1 and D4-3; it flowed westward at station D4-1 and eastward at station D4-3. In summary, the flow regime indicates that water is transported into JZB from the north and exits JZB to the Yellow Sea in the south. Based on the observation data at the whole bay mouth from south to north, the residual tidal flow is inflowing at the north and outgoing at the south and central parts. In neap tide, the residual tidal flow is mainly northwestward and is high in magnitude for the north part. In the spring, the residual current is inflowing at the northern part, but out-flowing at the southern and central parts (Wang 2004). The residual current in JZB is different from Tokyo Bay, where strong gravitational circulation at the mouth of the bay flows out of the bay in the upper layer and into the bay in the lower layer (Guo and Yanagi 1998). The residual current in JZB is also unlike Hiuchi-Nada Bay, where the water from the Kurushima Strait intrudes through the middle and bottom layers, while the water from the Bisan Strait intrudes through the surface layer, and water in Hiuchi-Nada Bay may enter the Kurushima Strait through the surface layer and move into the Bisan Strait through the bottom layer (Guo et al. 2004).

Current vectors in surface, middle and near-bottom waters at anchor station D4-2 in 2002

Current vectors in surface (1 m) and near-bottom (nb) waters at anchor stations D4-1 and D4-3 on 9 May (a) and 19 May (b) 2003
The amount of temporal nutrient transport through the entire water column along the east-westward direction (Fu) and the north-southward direction (Fv) at the anchor stations can be estimated, respectively, by:
and
where V and θ stand for the current speed and direction, respectively, h is water depth, and C is the concentration of nutrient species. The sinθ and cosθ represent the flux vector input (i.e., westward) and output of JZB (i.e., eastward), and accumulation northward and southward, respectively. This method for calculating nutrient fluxes has been applied in the Yalujiang Estuary (Liu and Zhang 2004).
In 2002, the duration of westward transport of nutrients was longer than eastward transport by a factor of 1.6–2.0 (Fig. 9). The transport along the north-southward direction was different for different nutrients; the northward fluxes were higher than the southward components by a factor of 1.3–1.5 for \( NO_{3} ^{ - } \), \( NH_{4} ^{ + } \) and DOP, and were rather comparable to the southward fluxes with a variation factor of 0.97–1.1 for the other nutrients (Fig. 9).

Temporal transport (mmol m−2 s−1) of nutrient species along east-westward (a) and north-southward (b) directions at anchor station D4-2 on 14–15 October 2002. Both eastward and northward transports are indicated with a positive value and both westward and southward by a negative value
During the neap tide in May 2003, the westward transport of nutrients was longer in duration than the eastward component by a factor of 1.5–2.5, although the instantaneous value of the eastward transport could be higher than the flow of nutrients moving westward at station D4-1, but was similar in the westward and eastward transport of all of the nutrients except for the DOP at station D4-3 (Fig. 10a). The nutrient fluxes along the north-southward direction were mainly northward transport at stations D4-1 and D4-3 (Fig. 10b).

Temporal transport (mmol m−2 s−1) of nutrient species along east-westward (a) and north-southward (b) directions at anchor stations D4-1 and D4-3 on 9–10 May 2003. Both eastward and northward transports are indicated with a positive value and both westward and southward by a negative value
During the spring tide in May 2003, the westward transport of nutrients was longer in duration than the eastward transport by a factor of 1–2 at station D4-1, similar to that during the neap tide, but the eastward transport of nutrients was up to 3-fold higher than the westward flow at station D4-3 (Fig. 11a). This indicates that nutrients were imported to JZB in the north but exported in the south during the spring tide. The nutrient transport in the north-southward direction was mainly northward, and was 1–3 times higher than the southward flow at station D4-1, but the southward transport of nutrients was 1–2 times higher than the northward transport at station D4-3 (Fig. 11b).

Temporal transport (mmol m−2 s−1) of nutrient species along east-westward (a) and north-southward (b) directions at anchor stations D4-1 and D4-3 on 19–20 May 2003. Both eastward and northward transports are indicated with a positive value and both westward and southward by a negative value
In May 2003, generally, the nutrients were transported into JZB at station D4-1 and transported out into the Yellow Sea at station D4-3. The ratios of nutrients coming into the bay to those leaving the bay were 0.7–1.4 for \( NO_{3} ^{ - } \), \( NO_{2} ^{ - } \), \( NH_{4} ^{ + } \) and DOP, and >1.5 for \( PO_{4} ^{{3 - }} \), \( SiO_{3} ^{{2 - }} \) and DON, indicating that \( PO_{4} ^{{3 - }} \) and \( SiO_{3} ^{{2 - }} \) might be transported into the bay from the Yellow Sea. In JZB, cell abundance showed a peak of phytoplankton biomass in September and January but remained low in May and November in 2003–2004 (Liu 2004). A preliminary estimate of nutrient budgets indicates that \( PO_{4} ^{{3 - }} \) and \( SiO_{3} ^{{2 - }} \) were transported into JZB from the Yellow Sea, while \( NO_{3} ^{ - } \) and \( NH_{4} ^{ + } \) were transported out of JZB (Liu et al. 2005a). This was similar to the data of the present study of the transport regime for \( NO_{3} ^{ - } \), \( PO_{4} ^{{3 - }} \), \( SiO_{3} ^{{2 - }} \) and \( NH_{4} ^{ + } \). The near-shore and offshore exchange of dissolved constituents often depends on the nutrient species and the geomorphology of the system, as reported for several European and American systems (Dame et al. 1991; Boorman et al. 1994; Hassen 2001; Bianchi et al. 2004).
4.3 Nutrient Budgets
A steady-state box model based on the LOICZ approach was used to construct nutrient budgets from non-conservative distributions of nutrients and water budgets, which in turn were constrained by the salt balance under a steady-state assumption. It was assumed that either the water volume remained constant or that the change in water volume over time was known, so that net water outflow from the system could be estimated by the difference (Gordon et al. 1996, also at http://data.ecology.su.se/MNODE/). The analysis of nutrient budgets is a common method for assessing the biogeochemical function of estuaries including denitrification and nutrient retention rates (Gordon et al. 1996; Simpson and Rippeth 1998; Chen and Wang 1999; Webster et al. 2000; Hung and Kuo 2002, de Madron et al. 2003; Souza et al. 2003; Liu et al. 2005a).
Nutrient transport fluxes from rivers into JZB in this study were based on the investigations in both dry and flood seasons in 2002 (Liu et al. 2005a) and 2004, estimated by the product of nutrient concentrations and freshwater discharge (Fig. 12). The dissolved inorganic nutrient fluxes from sewage were from investigations in 2002 (Liu et al. 2005a). There was no data for dissolved organic nutrients, so these were assumed to be the average concentrations of stream waters from Licunhe and Moshuihe, as these streams have become conduits of waste discharge.

Nutrient budgets in JZB (106 mol yr−1). In the figure, the bold typeface shows the budgets of model calculation; the light typeface represents the quantities estimated independently of budgetary calculations. The atmospheric deposition (P), riverine input (Q), exchange (X) and net transport (R) between the Yellow Sea (2) and Jiaozhou Bay (1), and net budget (empty triangle) are all shown. Positive values indicate transport into JZB; negative data show the export of nutrients from JZB
Based on rainwater and aerosol sample collection and measurement at Fulongshan in April 2004–March 2005, the atmospheric deposition fluxes of nutrients were 29.2 ± 4.0 × 106 mol yr−1 for \( NO_{3} ^{ - } \), 26.8 ± 3.2 × 106 mol yr−1 for \( NH_{4} ^{ + } \), 0.42 ± 0.07 × 106 mol yr−1 for \( PO_{4} ^{{3 - }} \), 0.93 ± 0.18 × 106 mol yr−1 for \( SiO_{3} ^{{2 - }} \), 29.0 ± 5.8 × 106 mol yr−1 for DON and 0.19 ± 0.04 × 106 mol yr−1 for DOP (unpublished data) (Fig. 12). These data are similar to the atmospheric deposition fluxes of \( NH_{4} ^{ + } \) and \( PO_{4} ^{{3 - }} \), but are a little different from the statistics reported previously for \( NO_{3} ^{ - } \) and \( SiO_{3} ^{{2 - }} \) (Liu et al. 2005a).
Based on water mass and salinity balance (Liu et al. 2005a), the residual flow and mixing flow transport of nutrients were estimated. The nutrient budgets are summarized in Fig. 12. Considering nutrient inputs, \( NO_{3} ^{ - } \) was mainly from riverine transport, accounting for 73%, then atmospheric deposition (20%); \( NH_{4} ^{ + } \) was mainly from wastewater discharge (86%); \( PO_{4} ^{{3 - }} \) was mainly from wastewater discharge (52%), then riverine transport (39%); \( SiO_{3} ^{{2 - }} \) was mainly from riverine transport (75%), then wastewater discharge (24%); DON was mainly from riverine transport (64%), then wastewater discharge (19%) and atmospheric deposition (17%); and DOP was mainly from riverine transport (61%), then wastewater discharge (31%). The residual flow transport of nutrients from JZB to the Yellow Sea was much less than the input sources; a mixing flow transported large amounts of \( NO_{3} ^{ - } \), \( NH_{4} ^{ + } \) and DON from JZB to the Yellow Sea. \( PO_{4} ^{{3 - }} \), \( SiO_{3} ^{{2 - }} \) and DOP, however, were transported from the Yellow Sea to JZB. Taking these flux estimates as valid, there was a net imbalance. The imbalance suggests that sinks of 163 × 106 mol yr−1 of \( NH_{4} ^{ + } \), 87.7 × 106 mol yr−1 of \( NO_{3} ^{ - } \), 7.52 × 106 mol yr−1 of \( PO_{4} ^{{3 - }} \), 214 × 106 mol yr−1 of \( SiO_{3} ^{{2 - }} \), and 15.7 × 106 mol yr−1 of DOP were buried in the sediment or were transformed into other forms (e.g., phytoplankton cell composition or particles); it also suggests that 23.3 × 106 mol yr−1 of DON is input into JZB. This indicates that JZB is a sink of all these nutrient elements except for DON.
4.4 Stoichiometric Relationships Among Nutrient Budgets
The net ecosystem metabolism (the difference between primary production and respiration [p–r]) can be estimated stoichiometrically from ΔDIP and the C:P ratio in the particulate matter being produced or consumed in JZB; therefore
Note that only DIP is used in the calculation of [p–r] because the production or consumption of DOP is regarded as one of the possible sinks or sources accounting for ΔDIP, and DIP is of primary interest to the budgetary analysis (Gordon et al. 1996). In JZB, the C:P ratio in particulate matter was 113. The annual primary production was higher than respiration, with net primary production of 85 × 107 mol carbon yr−1, indicating an autotrophic system. This value is equivalent to 16% of the annual gross production of 13.6 ± 1.4 mol m−2 yr−1 (Guo and Yang 1992; Wang et al. 1995; Liu et al. 2002).
Nitrogen metabolism in the sea expressed as the net result of nitrogen fixation and denitrification (nfix–denit), which is often quantitatively significant for the nitrogen budget, can be derived from the difference between non-conservative nitrogen flux and expected nitrogen removal through biological uptake:
The particulate N:P ratio in JZB was 20.5. This calculation demonstrates that nitrogen fixation was higher than denitrification, with a net nitrogen metabolism of 24.9 × 107 mol yr–1, indicating a net nitrogen-fixing system.
5 Summary
Jiaozhou Bay is a typical semi-enclosed water body, connected to the Yellow Sea through a narrow channel. JZB has been greatly disturbed by human activities, leading to changes in nutrient levels in past decades. Concentrations of nutrients in JZB were higher in the north than in the south. The eutrophication in JZB was characterized by high concentrations of \( NH_{4} ^{ + } \) (i.e., 60–70% of DIN), mainly coming from wastewater discharge. The high proportion of DON was also important for the JZB ecosystem, which could play an important role in coastal food webs and the nitrogen cycle. Nutrient ratios showed that the potential limiting nutrient for phytoplankton biomass was silicon, and phosphorus was next. The nutrient dynamics showed seasonal variations and a tidal induced pattern, with low values during flood tides and high values during ebb tides. The nutrient transport was estimated based on concentration and current measurements, which changed with tidal cycle, investigation sites, and seasons. Nutrient budgets demonstrated that riverine input and wastewater discharge were major sources of nutrients to the JZB ecosystem. Water exchange between JZB and the Yellow Sea exported \( NO_{3} ^{ - } \), \( NH_{4} ^{ + } \) and DON out of JZB into the Yellow Sea, while it imported \( PO_{4} ^{{3 - }} \), silicic acid and DOP into JZB. JZB was a sink for all the nutrients studied except for DON. Stoichiometric calculations demonstrated that JZB was a net autotrophic system.
References
Aubry, F. B., & Acri, F. (2004). Phytoplankton seasonality and exchange at the inlets of the Lagoon of Venice (July 2001–June 2002). Journal of Marine Systems, 51, 65–76.
Bianchi, F., Ravagnan, E., Acri, F., Bernardi-Aubry, F., Boldrin, A., Camatti, E., et al. (2004). Variability and fluxes of hydrology, nutrients and particulate matter between the Venice Lagoon and the Adriatic Sea. Preliminary results (years 2001–2002). Journal of Marine Systems, 51, 49–64.
Boorman, L. A., Hazelden, J., Loveland, J. P., Wells, J. G., & Levasseur, J. E. (1994). Comparative relationships between primary productivity and organic and nutrient fluxes in four European salt marshes. In W. J. Mitsh (Ed.), Global wetlands. Old world and new (pp. 181–199). New York: Elsevier.
Chen, C.-T. A., & Wang, S.-L. (1999). Carbon, alkalinity and nutrient budgets on the East China Sea continental shelf. Journal of Geophysical Research, 104(C9), 20675–20686.
Dale, A.W., & Prego, R. (2003). Tidal and seasonal nutrient dynamics and budget of the Chupa Estuary, White Sea (Russia). Estuarine, Coastal and Shelf Science, 56, 377–389.
Dame, R. F., Spurrier, J. D., Williams, T. M., Kjerfve, B., Zingmark, R. G., Wolover, T. G., et al. (1991). Annual material processing by a salt marsh-estuarine basin in South Carolina. Marine Ecology Progress Series, 72, 153–166.
de Madron, X. D., Denis, L., Diaz, F., Garcia, N., Guieu, C., Grenz, C., et al. (2003). Nutrients and carbon budgets for the Gulf of Lion during the Moogli cruises. Oceanologica Acta, 26, 421–433.
Downing, J. A., McClain, M., Twilley, R., Melack, J. M., Elser, J., Rabalais, N. N., et al. (1999). The impact of accelerating land-use change on the N-cycle of tropical aquatic ecosystems: Current conditions and projected changes. Biogeochemistry, 46, 109–148.
Gilbert, F., Souchu, P., Brianchi, M., & Bonin, P. (1997). Influence of shellfish farming activities on nitrification, nitrate reduction to ammonium and denitrification at the water-sediment interface of the Thau lagoon, France. Marine Ecology Progress Series, 151, 143–153.
Gordon, D. C., Boudreau, P. R., Mann, K. H., Ong, J.-E., Silvert, W. L., Smith, S. V., Wattayakorn, G., Wulff, F., & Yanagi, T. (Eds.) (1996). LOICZ biogeochemical modelling guidelines. LOICZ reports and studies 5. The Netherlands: Texel, LOICZ.
Grasshoff, K., Kremling, K., & Ehrhardt, M. (Eds.) (1999). Methods of seawater analysis, 3rd ed. Weinheim: Wiley.
Guo, X., Futamura, A., & Takeoka, H. (2004). Residual currents in a semi-enclosed bay of the Seto Inland Sea, Japan. Journal of Geophysical Research, 109, C12008. doi:10.1029/2003JC002203.
Guo, X., & Yanagi, T. (1998). Variation of residual current in Tokyo Bay due to increase of fresh water discharge. Continental Shelf Research, 18, 677–693.
Guo, Y.-J., & Yang, Z.-Y. (1992). The phytoplankton in Jiaozhou Bay. In R. Liu (Ed.), Ecology and living resources of Jiaozhou Bay (pp. 136–139). Beijing: Science. (in Chinese).
Hao, J.-H., Huo, W.-Y., & Yu, Z.-M. (2000). Preliminary study on red tide occurrence in relation to nutritional condition in aquaculture seawater of Jiaozhou Bay. Marine Science, 24(4), 37–41 (in Chinese).
Hassen, M. B. (2001). Spatial and temporal variability in nutrients and suspended material processing in the Fier d’Ars Bay (France). Estuarine, Coastal and Shelf Science, 52, 457–469.
Hatcher, A., Grant, J., & Schofield, B. (1994). Effect of suspended mussel culture (Mytilus spp.) on sedimentation, benthic respiration and sediment nutrient dynamics in a coastal bay. Marine Ecology Progress Series, 115, 219–235.
Humborg, C., Ittekkot, V., Cociasu, A., & Bodungen, B. V. (1997). Effect of Danube River dam on Black Sea biogeochemistry and ecosystem structure. Nature, 386, 385–388.
Hung, J.-J., & Kuo, F. (2002). Temporal variability of carbon and nutrient budgets from a tropical lagoon in Chiku, southwestern Taiwan. Estuarine, Coastal and Shelf Science, 54, 887–900.
Huo, W.-Y., Yu, Z.-M., Zou, J.-Z., & Han, X.-T. (2001). Analysis of dynamic process and the causes of Eucampia zoodiacus red tide in Jiaozhou Bay. Journal of Fisheries of China, 25(3), 222–226 (in Chinese).
Jennerjahn, T. C., Ittekkot, V., Klöpper, S., Adi, S., Nugroho, S. P., Sudiana, N., et al. (2004). Biogeochemistry of a tropical river affected by human activities in its catchment: Brantas River estuary and coastal waters of Madura Strait, Java, Indonesia. Estuarine, Coastal and Shelf Science, 60, 503–514.
Kamatani, A., & Takano, M. (1984). The behavior of dissolved silica during the mixing of river and seawaters in Tokyo Bay. Estuarine Coastal and Shelf Science, 19, 505–512.
Li, Y., Li, R.-X., Wang, Z.-L., Zhu, M.-Y., Sun, P.-X., & Xia, B. (2005). A preliminary study on phytoplankton community structure and its changes in the Jiaozhou Bay. Advances in Marine Science, 23(3), 328–334 (in Chinese with English abstract).
Liu, D. (2004). Community structure succession study of phytoplankton and sediment diatom in Jiaozhou Bay. Dissertation, Ocean University of China (in Chinese, with English abstract).
Liu, D., Sun, J., & Qian, S. (2002). Study on the phytoplankton in Jiaozhou Bay II: Influence of the environmental factors on the phytoplankton community. Journal of Ocean University of Qingdao, 32(3), 415–421 (in Chinese, with English abstract).
Liu, D., Sun, J., & Zhang, L. (2003a). Structural characteristics of phytoplankton community during harmful algae bloom in Jiaozhou Bay. Chinese Journal of Applied Ecology, 14(11), 1963–1966 (in Chinese).
Liu, Z., Wei, H., Liu, G., & Zhang, J. (2004a). Simulation of water exchange in Jiaozhou Bay by average residence time approach. Estuarine, Coastal and Shelf Science, 61(1), 25–35.
Liu, S. M., & Zhang, J. (2004). Nutrient dynamics in the macro-tidal Yalujiang Estuary. Journal of Coastal Research, Special Issue, 43, 147–161.
Liu, S. M., Zhang, J., Chen, S. Z., Chen, H. T., Hong, G. H., Wei, H., et al. (2003b). Inventory of nutrient compounds in the Yellow Sea. Continental Shelf Research, 23, 1161–1174.
Liu, S. M., Zhang, J., Chen, H. T., & Rabbe, T. (2004b). Benthic nutrient recycling in shallow coastal waters of the Bohai. Chinese Journal of Oceanology and Limnology, 22(4), 365–372.
Liu, S. M., Zhang, J., Chen, H. T., & Zhang, G. S. (2005a). Factors influencing nutrient dynamics in the eutrophic Jiaozhou Bay, North China. Progress in Oceanography, 66, 66–85.
Liu, S. M., Zhang, J., & Li, D. J. (2004c). Phosphorus cycling in sediments of the Bohai and Yellow Sea. Estuarine, Coastal and Shelf Science, 59, 209–218.
Liu, S. M., Zhang, J., & Li, R. X. (2005b). Ecological significance of biogenic silica in the East China Sea. Marine Ecology Progress Series, 290, 15–26.
Lu, M., Zhang, L.-J., Li, C., Zou, L., & Zhang, J. (2001). Analysis of the ecological environment elements in the red tide generating and vanishing process in the eastern Jiaozhou Bay in July 1999. Journal of Oceanography of Huanghai and Bohai Seas, 19(4), 43–50 (in Chinese).
Mackas, D. L., & Harrison, P. J. (1997). Nitrogenous nutrient sources and sinks in the Juan de Fuca Strait/Strait of Georgia/Puget Sound estuarine system: Assessing the potential for eutrophication. Estuarine coastal Shelf Science, 44, 1–21.
Mortlock, R. A., & Froelich, P. N. (1989). A simple method for the rapid determination of biogenic opal in pelagic marine sediments. Deep-Sea Research, 36(9), 1415–1426.
Newtona, A., & Mudge, S. M. (2005). Lagoon-sea exchanges, nutrient dynamics and water quality management of the Ria Formosa (Portugal). Estuarine, Coastal and Shelf Science, 62, 405–414.
Piehler, M. F., Twomey, L. J., Hall, N. S., & Paerl, H. W. (2004). Impacts of inorganic nutrient enrichment on phytoplankton community structure and function in Pamlico Sound, NC, USA. Estuarine, Coastal and Shelf Science, 61, 197–209.
Qingdao Municipal Statistics Bureau (1998). Qingdao statistical yearbook. Beijing: China Statistics Press.
Qingdao Municipal Statistics Bureau (2003). Qingdao statistical yearbook. Beijing: China Statistics Press.
Ragueneau, O., Lancelot, C., Egorov, V., Vervlimmeren, J., Cociasu, A., Déliat, G., et al. (2002). Biogeochemical transformations of inorganic nutrients in the mixing zone between the Danube River and the Northwestern Black Sea. Estuarine, Coastal and Shelf Science, 54, 321–336.
Ragueneau, O., Tréguer, P., Anderson, R. F., Brzezinski, M. A., DeMaster, D. J., Dugdale, R. C., et al. (2000). A review of the Si cycle in the modern ocean: Recent progress and missing gaps in the application of biogenic opal as a paleoproxy. Global and Planetary Change, 543, 315–366.
Ren, J. L., Zhang, J., Luo, J. Q., Pei, X. K., & Jiang, Z. X. (2001). Improved fluorimetric determination of dissolved aluminium by micelle-enhanced lumogallion complex in natural waters. The Analyst, 126, 698–702.
Rysgaard, S., Thastum, P., Dalsgaard, T., Christensen, P. B., & Sloth, N. P. (1999). Effects of salinity on \( NH_{4} ^{ + } \) adsorption capacity, nitrification, and denitrification in Danish Estuarine sediments. Estuaries, 22(1), 21–30.
Shen, Z.-L. (2001). Historical changes in nutrient structure and its influences on phytoplankton composition in Jiaozhou Bay. Estuarine, Coastal and Shelf Science, 52, 211–224.
Simpson, J. H., & Rippeth, T. P. (1998). Non-conservative nutrient fluxes from budgets for the Irish Sea. Estuarine, Coastal and Shelf Science, 47, 707–714.
Souza, M. F. L., Kjerfve, B., Knoppers, B., de Souza, W. F. L., & Damasceno, R. N. (2003). Nutrient budgets and trophic state in a hypersaline coastal lagoon: Lagoa de Araruama, Brazil. Estuarine, Coastal and Shelf Science, 57, 843–858.
Su, H.-M., Lin, H.-J., & Hung, J.-J. (2004). Effects of tidal flushing on phytoplankton in a eutrophic tropical lagoon in Taiwan. Estuarine, Coastal and Shelf Science, 61, 739–750.
Sun, Y., Chen, J. F., & Zhang, Y. B. (1993). Analysis of chemical indexes of nutritional condition in Jiaozhou Bay waters. Marine Environmental Science, 12, 25–31 (in Chinese).
Sun, S., Liu, G.-M., Zhang, Y.-S., Wu, Y.-L., Pu, X.-M., & Yang, B. (2002). Community composition of distribution characteristics of phytoplankton in the Jiaozhou Bay in the 1990s. Oceanologia et Limnologia Sinica, 33, 37–44 (in Chinese with English abstract).
Turner, R. E. (2002). Element ratios and aquatic food webs. Estuaries, 25, 694–703.
Turner, R. E., & Rabalais, N. N. (1994). Coastal eutrophication near the Mississippi River delta. Nature, 368, 619–621.
Turner, R. E., Rabalais, N. N., Justic, D., & Dortch, Q. (2003). Future aquatic nutrient limitations. Marine Pollution Bulletin, 46, 1032–1034.
Wang, Z. Y. (2004). Observation and analysis of tidal current in the mouth of the Jiaozhou Bay. Dissertation, Ocean University of China (in Chinese).
Wang, R., Jiao, N. Z., Li, C. L., Shen, Z. L., & Ji, P. (1995). Primary production and new production in Jiaozhou Bay. Studia Marina Sinica, 36, 181–194 (in Chinese).
Webster, I. T., Smith, S. V., & Parslow, J. S. (2000). Implications of spatial and temporal variation for biogeochemical budgets of estuaries. Estuaries, 23(3), 341–350.
Yang, D. F. (1999). Influences of silicate, light and temperature on phytoplankton growth. Dissertation, Institute of Oceanology, Chinese Academy of Sciences (in Chinese).
Yang, S., Meng, Y., Zhang, J., Xue, Y., Chen, H., Wei, H., et al. (2004). Suspended particulate matter in Jiaozhou Bay: Properties and variations in response to hydrodynamics and pollution. Chinese Science Bulletin, 49(1), 91–97.
Yin, P., & Lu, Y.-X. (2000). Environmental evolution and sustainable utilization of Jiaozhou Bay. Coastal Engineering, 19(3), 14–22 (in Chinese).
Zhang, J. (2007). Watershed nutrient loss and eutrophication of the marine recipients: A case study of the Jiaozhou Bay. Water, Air, and Soil Pollution: Focus. (this volume).
Zhang, J. S., & Shen, Z. L. (1997). A study of changes in the nutrient structure of Jiaozhou Bay. Oceanologia et Limnologia Sinica, 28, 529–535 (in Chinese).
Zou, L. (2001). An experimental study on the relationship between nutrients and phytoplanktons in the coastal waters of North China. Dissertation, Ocean University of China (in Chinese).
Acknowledgements
The authors would like to thank Dr. Z.Y. Wang for his assistance in data processing, Drs. X. Luo, X.W. Ye, X.H. Qi and G.S. ZHang and other colleagues from the Laboratory of Marine Biogeochemistry, OUC for their help in field and laboratory experiments. This study was funded by the Chinese Natural Sciences Foundation (Nos. 40036010 and 40206017), and the Ministry of Science and Technology, P. R. China (No. 2001CB409703). Additional financial support was provided by SKLEC/ECNU for C and N determinations. Dr. P.Y. Tishchenko and two anonymous reviewers are appreciated for their comments and suggestions, which improved the original manuscript. We are grateful to Ms. J. Willson for the syntax corrections of English.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Liu, S.M., Li, X.N., Zhang, J. et al. Nutrient Dynamics in Jiaozhou Bay. Water Air Soil Pollut: Focus 7, 625–643 (2007). https://doi.org/10.1007/s11267-007-9125-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11267-007-9125-y