
Abstract
In the city of Bakersfield, CA, endangered San Joaquin kit foxes (Vulpes macrotis mutica) occur with raccoons (Procyon lotor), striped skunks (Mephitis mephitis), and Virginia opossums (Didelphis virginiana), all of which share the urban landscape with an abundance of domestic dogs (Canis familiaris) and cats (Felis catus). Interactions between domestic and wild carnivores range from ambivalent to competitive and coexistence may be facilitated by partitioning space, time, and resources. An understanding of these mechanisms among increasing urbanization is of particular importance for the conservation of San Joaquin kit foxes. We used annual survey data and quantified urban land uses in multi-scaled χ2 spatial analyses and occupancy modeling to explore how kit foxes and three other mesocarnivores manage cohabitation with dogs and cats. Cat occupancy was driven by a negative association with open spaces, skunk occupancy was driven by a negative association with commercial land uses and a positive association with industrial land uses, and opossum occupancy was driven by a negative association with open spaces as well as a positive association with parks. Kit foxes occurred mostly independently of dogs and cats, though showed some fine-scale spatial avoidance of cats. Opossums exhibited fine-scale spatial avoidance of dogs while raccoons and skunks exhibited fine-scale spatial coexistence with cats. Ultimately, these species select for different urban landscapes and mesocarnivores show varying changes in space use when sharing a landscape immediately with domestic species which may help facilitate coexistence between wild mesocarnivores and domestic species in Bakersfield.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Over half of the world’s human population currently resides in urban areas, with this figure projected to increase to nearly 70% by 2050 (United Nations, Department of Economic and Social Affairs, Population Division 2019). With habitat loss the leading cause of extinction in wild animals, species in areas of encroaching urban development must adapt or risk extinction (Gehrt 2010; Primack and Morrison 2013). As compared to natural ecosystems, urban environments have increased ambient temperature, available surface water, light and noise pollution, non-native species, and habitat heterogeneity and fragmentation (Gehrt 2010). Wildlife in urban areas may benefit from additional available resources including anthropogenic foods and refuge, though may be impeded by unfamiliar dangers such as roads, pollutants, poisons, parasites, and disease (Fuller et al. 2010; Gehrt 2010). Urban wildlife also share the landscape with an abundance of domestic dogs (Canis familiaris) and cats (Felis catus). Dogs and cats are typically the most abundant and ubiquitous carnivores in urban areas due to their close relationship with humans (Baker et al. 2010). Dogs and cats may serve as competitors, predators, or prey to urban wildlife and can be hosts for contagious pathogens (Beck 1973; Bradley and Altizer 2006; Gehrt 2010). Activity patterns of free-roaming dogs and cats concentrate around available food, water, and shelter that are often provided by human caretakers, as well as the availability of reproductive conspecifics (Baker et al. 2010). Both dogs and cats will avoid areas frequented by larger predators and are commonly observed after dark (Berman and Dunbar 1983; Baker et al. 2010).
Wild carnivores in urban environments may exhibit changes in home range sizes, primary food sources, litter sizes, and survival rates as compared to conspecifics in natural environments (Iossa et al. 2010). Several mesocarnivores are found in the city of Bakersfield, California, including coyotes (C. latrans), red foxes (Vulpes vulpes), gray foxes (Urocyon cinereoargenteus), San Joaquin kit foxes (V. macrotis mutica), raccoons (Procyon lotor), striped skunks (Mephitis mephitis), and Virginia opossums (Didelphis virginiana). Red foxes and opossums are not native to the area and San Joaquin kit foxes (hereafter, kit fox) are listed as federally Endangered and Threatened in California due to habitat loss and degradation (Gardner 1982; Lewis et al. 1993; United States Fish and Wildlife Service 1998). In urban environments these species are generally nocturnal, occur in higher densities with smaller home ranges, den in a variety of landscape features including under or within man-made structures, and will opportunistically consume anthropogenic food sources (Rosatte and Lariviére 2003; Cypher 2010; Gehrt and Riley 2010; Hadidian et al. 2010; Rosatte et al. 2010; Soulsbury et al. 2010; Wright et al. 2012; Lombardi et al. 2017). Proclivity for certain landscapes within the urban matrix by carnivores may be a result of microhabitat preferences as well as avoidance of predators or competitors (Crooks et al. 2010). Patterns in past studies on urban carnivores suggest coyotes, red foxes, and gray foxes will select for more natural or undeveloped fragments in the urban landscape (Ansell 2004; Gehrt and Riley 2010; Soulsbury et al. 2010; Kapfer and Kirk 2012). Kit foxes and skunks prefer maintained green spaces with large yards such as schools, golf courses, and churches, and will also select for open space such as water catchment basins and canals (Rosatte et al. 1991, 2010; Frost 2005; Deatherage et al. 2021). Raccoons and opossums prefer areas near water and raccoons further select for tree cover and use residential areas, focusing activity around anthropogenic food sources such as picnic areas and garbage receptacles (Bozek et al. 2007; Hadidian et al. 2010; Fidino et al. 2016).
Interactions among domestic and wild carnivores range from ambivalent to competitive and are dependent on habitat type, species abundances, food habits, resource availability, body size differences, and behavior, and can have dramatic consequences on the structure of carnivore communities (Crooks et al. 2010; Fuller et al. 2010). Larger carnivores will sometimes kill smaller carnivores for food or competitive reasons; coyotes will attack all the smaller species, larger dogs will also attack coyotes and the smaller species, and red foxes will occasionally attack cats, kit foxes, and skunks (Harris 1981; Crooks and Soulé 1999; Kamler et al. 2003; Cypher 2003; Farias et al. 2005; Gosselink et al. 2007; Baker et al. 2010; Hadidian et al. 2010; Shedden et al. 2020). Ecologically similar species can facilitate coexistence with antagonistic competitors by partitioning time, space, and resources (Schoener 1974). Consequently, cats and opossums have been shown to have consistent negative spatial and temporal relationships with coyotes in San Diego, CA (Crooks and Soulé 1999). Conversely, cats and skunks have been observed denning simultaneously with kit foxes and each other (Cypher 2010).
Changes in frequency and intensity of interspecific interactions are anticipated in urban areas as distributions are limited by impeding development and population densities tend to be elevated (Cypher 2010; Hadidian et al. 2010; Riley et al. 2010). The fragmentation of urban landscapes tends to concentrate some carnivores into remaining natural or green spaces, leading to an increase in interactions (Parsons et al. 2019). Little is known about how species respond to frequent encounters in urban areas, particularly between wild mesocarnivores and novel domestic competitors. While coexistence between domestic and wild carnivores is a necessary adaptation for urban living, risks of high population densities include increased competition, predation, and disease transmission. These risks are of particular concern for endangered species like the kit fox.
We used presence/absence data to investigate how kit foxes and three other mesocarnivores: raccoons, skunks, and opossums, manage cohabitation with ubiquitous domestic dogs and cats in the urban environment of Bakersfield. We used chi-square analyses to test spatial relationships between domestic species and wild mesocarnivores on two spatial scales. We also used occupancy modeling to investigate urban landscape preferences for the species and further co-occurrence patterns between domestic species and kit foxes. We expected cats and opossums to prefer similar urban landscapes that included easy access to resources such as parks and other maintained green space and expected skunks would select for more undeveloped or open areas. We also expected kit foxes and the other mesocarnivores would occur independently of cats while showing some spatial avoidance of dogs due to the tendency of dogs to kill the smaller species.
Methods
Study area
The city of Bakersfield is located in Kern County, within the southeastern San Joaquin Valley of Central California. The area is generally flat with elevations roughly between 30 and 183 m above sea level (Stachelski and Sanger 2008). Hot, dry summers, and cool, wet winters characterize the desert climate, with average daily maximum temperatures ranging from 14°C in December to 37°C in July and average daily minimum temperatures ranging from 4°C in December to 21°C in July (National Oceanic and Atmospheric Administration 2020). Annual precipitation averages 16.4 cm but varies greatly between years (National Oceanic and Atmospheric Administration 2020).
Bakersfield is a heavily developed city with a growing population of nearly 380,000 people. Only 25 – 30% of Bakersfield abuts natural environments including arid grasslands, shrublands, and riparian areas, with the remainder bordered by agriculture (Cypher 2010). Bakersfield encompasses a variety of urban land uses including commercial developments, preserved green spaces, agriculture, campuses, residential, recreational, and industrial areas. The Kern River runs through the city though typically only portions of the corridor contain water year-round due to diversion for agriculture. Vegetation in the city consists primarily of a mix of planted native and non-native ornamental trees, shrubs, and flowering plants. A multitude of free-roaming dogs and cats inhabit the area, with approximately 6,600 dogs and 5,500 cats reported as stray intakes at Kern County Animal Services in 2019 (Kern County Animal Services 2019).
Study design
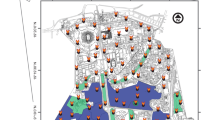
We conducted annual camera surveys throughout Bakersfield from 2015 to 2021 using camera station methodology developed specifically to survey for kit foxes and other sympatric mesocarnivores (Westall and Cypher 2017). We established a sampling grid of up to 111 1-km2 cells to be surveyed each year, randomly selected from a total of 358 cells that contained ≥ 50% urban development, thus sampling approximately 31% of the city (Fig. 1). Each cell was roughly representative of the average kit fox home range of 0.78 km2 in urban areas (Cypher et al. 2023). Studies have shown the other species to also have home ranges smaller than or close to 1 km2 in urban areas (Rosatte et al. 1991; Gehrt 2004; Baker et al. 2010; Wright et al. 2012). We placed one motion-triggered trail camera (Black Flash model C3 or E3, Cuddeback, Green Bay, WI, USA) in each cell, secured within protective cases (CuddeSafe E Model 3327, Cuddeback, Green Bay, WI, USA) to t-posts, fences, or vegetation with zip-ties and cable locks. We selected camera locations based on permission to survey properties, accessibility for mesocarnivores, and minimal risk of human disturbance. We attempted to find locations toward the center of cells to place cameras such that stations in adjacent cells would be separated by approximately 1 km, though cameras were not placed in locations without a means of entry by our target species (e.g., areas completely enclosed within tall, solid walls) and were hidden out of view (e.g., within vegetation or behind structures) in areas with consistent high human activity. With a few exceptions due to human disturbance, cells surveyed and camera locations remained consistent each year. In locations where cameras were tampered with or stolen, efforts were made to place cameras in different locations within the same cell; however, if this was not possible another cell was used as a replacement or the cell was omitted in any given year. This resulted in 112 different cells being surveyed over the seven-year sampling period.

112 1-km2 grid cells throughout Bakersfield, CA surveyed annually from 2015 to 2021 for urban mesocarnivores
We used 30-cm nails to secure a perforated 163-ml can of cat food approximately 2 m in front of each camera, added several drops of a carnivore scent lure (Carman’s Canine Call, Minnesota Trapline Products Inc., Pennock, MN, USA) as bait, and clipped back excess vegetation from the camera’s field of view. We programmed cameras to wide view and the highest resolution (20 MP) to collect one image following a 15-s delay during daylight (to reduce images of vegetation and human activity) and three images as fast as possible after dark. We deployed cameras in up to 15 cells at a time between June and September each year. Cameras were deployed for seven nights and then images were reviewed for the number of species detections each night, defined as 1200 through 1159 the following day. Images went through two rounds of review, first by two to three researchers concurrently to confirm species presence and a second time by a single researcher to record the specific number of individuals and number of nights they were detected in each cell. Individual animals were distinguished from others of the same species based on size, sex, fur patterns, or markings. If no distinguishing characteristics were discernable, we counted multiple appearances of a species on camera during the 7-n session as the same individual.
Landscape attributes
We used satellite imagery maps to quantify urban landscape attributes in surveyed cells to use in analyses. We superimposed cells with a 10 × 10 m point grid (100 points/cell) in Google Earth Pro and used 2018 imagery at an eye altitude of 300-m above-ground-level to characterize points and camera locations scaled to 1.0. We characterized each point by the landscape that best described that location (i.e., the terrestrial land use on which the point was located on or nearest to), recorded if the point fell on a mature tree, and noted the presence of stable water sources within cells (e.g., water catchment basins, recreational ponds and lakes, agricultural water supply, the Kern River, and other large bodies of water). Point counts approximated percentages of 12 landscape attributes in cells (Table 1).
Course-scale spatial analyses
To assess spatial relationships between domestic species and mesocarnivores within urban landscapes, we compared the total number of detections in landscape attributes specifically describing camera locations for each pairwise relationship between dogs or cats and kit foxes as well as other smaller mesocarnivores (raccoons, skunks, and opossums combined into one account of “smaller mesocarnivores” due to low detections) using a χ2 contingency table test for association and Yate’s corrected χ2 goodness of fit tests (Gotelli and Ellison 2013).
Fine-scale spatial analysis
To assess spatial relationships between domestic species and mesocarnivores on a more refined scale, we tested for associations among species presence on nights when they occurred for each pairwise relationship between dogs or cats and kit foxes, raccoons, skunks, or opossums using χ2 two-way contingency tables (Gotelli and Ellison 2013). We tested these relationships across all cells from all years collectively, as well as each year individually for associations with cats. All tests were analyzed at an α-level of 0.05 for significance.
Occupancy modeling
We used occupancy modeling to investigate associations between urban land uses and species occupancy in surveyed cells, as well as interaction patterns between domestic species and kit foxes within the urban landscape (MacKenzie et al. 2018). We used detection histories to estimate single-species occupancy probabilities (ψ) for cats, opossums, and skunks in relation to landscape attributes in a single-season framework (MacKenzie et al. 2018). Dogs and raccoons performed poorly in single-species occupancy modeling so were excluded from this analysis. Campuses with large maintained green yards (e.g., schools and churches) and paved roads, particularly highways, have been previously described as likely positive and negative, respectively, influential landscape attributes for urban kit fox occupancy (Deatherage et al. 2021; Cypher et al. 2023). These attributes were used in co-occurrence modeling for kit foxes. Important attributes identified for cats from single-species modeling were also included in co-occurrence modeling for cats and kit foxes. We calculated species co-occurrence (φ) and co-detection (δ) factors in a two-species, single-season framework for dogs or cats and kit foxes (MacKenzie et al. 2018).
We used PRESENCE 2.13.10 (Hines 2006) to fit occupancy models using all surveyed cells (s = 112). We set each year to represent a survey (K = 7) and set detection (p or r) parameters to remain constant over surveys (MacKenzie et al. 2018). We felt this model design was appropriate for our data because changes in habitat composition in the urban environment are minimal once developed and the specific year a cell was surveyed would therefore have minimal effect on occupancy. Further, covariates affecting detection probabilities were not measured in this study as we were primarily concerned with occupancy probabilities and felt detection variation was limited year-to-year due to consistencies in cells surveyed each year, time of year cells were surveyed, use of bait and lure, and a relatively short survey period. We used alternate parameterization in PRESENCE and set landscape attribute covariates to vary between the two species for co-occurrence models (Hines 2006; MacKenzie et al. 2018). Single-species candidate models included a null model (no covariates), a global model (including all covariates considered most likely to influence occupancy for a given species), univariate models, and additive pairwise combinations of covariates following four a priori categories representing varying degrees of urban development: high, intermediate, low, and green space (Table 1). We tested pairwise correlations between covariates using Spearman’s Rank tests and corrected the resulting P-values (Legendre and Legendre 1998). We considered an rs value > ± 0.50 as moderately correlated and such covariates were not included in the same model (Burnham and Anderson 2002).
We calculated Pearson’s χ2 goodness-of-fit statistic and an overdispersion factor (\(\widehat{c}\)) using 1,000 bootstraps for global models at an α-level of 0.05 (Burnham and Anderson 2002). We used Akaike’s Information Criterion adjusted for overdispersion (QAIC, χ2 P-value < 0.05 and \(\widehat{c}\) ⪆ 1.00) and small sample sizes (AICc, sample size/maximum number of model parameters < 40) where appropriate (Burnham and Anderson 2002). We examined discontinuity in ΔAIC values (the difference between the AIC of the top model in the set and the AIC of the given model) to rate model adequacy and then examined the direction and relative magnitude of covariate coefficient estimates (β) to evaluate covariate effects for a given species (Burnham and Anderson 2002). For co-occurrence models, we considered φ or δ values < 1.00 to signify a negative association between species (i.e., avoidance or exclusion), φ or δ values > 1.00 to signify a positive association, and φ or δ values ≈ 1.0 to signify independence (MacKenzie et al. 2018).
Results
Camera results are summarized over all surveyed cells from all seven years in Table 2. Detections were highest for cats (58% of detections), followed by kit foxes (17%), dogs (8%), skunks and opossums (both 5%), and finally raccoons (4%). Cats occurred in more cells and on more nights than any other species each year (80% of surveyed cells and 43% of surveyed nights) followed by kit foxes (35% of surveyed cells and 16% of surveyed nights). Dogs occurred in frequencies similar to raccoons, skunks, and opossums, with dogs occurring in the most cells (17%) and opossums occurring on the most nights (6%).
Landscape attributes
Of the 112 cells, most were dominant in residential land use, followed by other open space, paved road, industrial, commercial and agriculture, and finally park land uses (Table 1). Over half the cells had a stable water source available and nearly one third had ≥ 20% mature tree cover. Camera stations were located in one of eight different landscape characterizations (Table 1). There were 119 camera locations due to seven cameras being moved to different locations within the cell throughout the study. The average distance between camera locations was 1.06 km (± 0.10 km), with a minimum distance of 0.41 km. Cameras that were classified in the roadway attribute were always located on the side of the road and cameras that were classified in the other open space attribute were specified as either undeveloped lot (13 cameras) or other open space (primarily water catchment basins, 44 cameras). Most cameras were located in other open space (37% of cameras) and campuses (34%), followed by undeveloped lots (11%), commercial areas (8%), and industrial areas, side of the roadway, and parks (all 3%), with the least number of cameras located in residential areas (< 1%).
Course-scale spatial analyses
We found associations between species occurrences and land use at camera locations (χ2 = 358.15, df = 21, P < 0.001). Dogs occurred more frequently than kit foxes along roadways than would be expected if they occurred equally in the attribute (i.e., 50:50 frequencies; χ2 = 24.45, df = 1, P < 0.001). Dogs also occurred more frequently than other smaller mesocarnivores along roadways as well as in parks (χ2 range = 5.33 – 19.45, all df = 1, P range = < 0.001 – 0.02). Dogs occurred less frequently than kit foxes in all other land uses apart from park and industrial (χ2 range = 4.98 – 49.77, all df = 1, P range = < 0.001 – 0.03) and less frequently than other smaller mesocarnivores in all other land uses apart from residential areas or undeveloped lots (χ2 range = 4.98 – 99.05, all df = 1, P range = < 0.001 – 0.03). Dogs did not have an association with kit foxes in parks or industrial areas (χ2 range = < 0.01 – 0.05, all df = 1, P range = 0.83 – 1.00), nor with other smaller mesocarnivores in residential areas or undeveloped lots (χ2 range = < 0.01 – 0.05, all df = 1, P range = 0.48 – 1.00). Cats occurred more frequently than kit foxes in most land uses (χ2 range = 9.80 – 381.03, all df = 1, all P < 0.001) and more frequently than other smaller mesocarnivores in all land uses than would be expected if the species occurred equally (χ2 range = 9.60 – 283.45, all df = 1, P range = < 0.001 – 0.02). Cats did not have an association with kit foxes in residential areas or undeveloped lots (χ2 range = 0.01– 0.70, all df = 1, P range = 0.40 – 0.94).
Fine-scale spatial analysis
Opossum nightly presence was lower when dogs were also present across all years combined (χ2 = 7.91, df = 1, P = 0.005); kit fox nightly presence was lower when cats were also present in 2016, 2020, and across all years (χ2 range = 6.68 – 23.86, all df = 1, P range = < 0.001 – 0.004); raccoon nightly presence was higher when cats were also present in 2020 and across all years (χ2 range = 4.32 – 14.28, all df = 1, P range = < 0.001 – 0.04); skunk nightly presence was higher when cats were also present in 2016, 2018, and across all years (χ2 range = 5.62 – 36.60, all df = 1, P range = < 0.001 – 0.02) though was lower when cats were present in 2015 (χ2 = 28.88, df = 1, P = < 0.001); and opossum nightly presence was higher when cats were also present in 2016 (χ2 = 13.01, df = 1, P = < 0.001) though the opposite was found in 2021 (χ2 = 6.21, df = 1, P = 0.01). The remaining 24 pairwise relationships between nightly presence were independent (χ2 range = < 0.01 – 3.29, all df = 1, P range = 0.07 – 0.96).
Occupancy modeling
Five of the 12 landscape attributes: water, road, rail, KR, and ag, were excluded from single-species occupancy analyses due to overall poor performance in models (i.e., did not converge or produced large standard errors for β values across the species). Of the viable covariates, five of the pairwise combinations were moderately correlated: tree + OOS (rs = -0.57), res + ind (rs = -0.56), res + OOS (rs = -0.55), tree + ind (rs = -0.52), and tree + res (rs = 0.75; all df = 356, all P (corrected) < 0.001). The other 23 combinations were less than moderately correlated (rs range = -0.39 – 0.37, all df = 356, P (corrected) range = < 0.001 – 1.736).
Single-species
Including the global and null models, we developed 29 candidate models for cats and 25 candidate models for skunk and opossum single-species modeling (Online Resource 1). The res covariate was differentiated into low-to-medium density residential and high-density residential attributes to test for more fine-scale effects of residential land use on cats (Table 1). The cat global model included HDR, LMDR, com, camp, and park covariates and resulted in the use of QAIC for cats (K = 8, χ2 = 800.03, P = 0.03, \(\widehat{c}\) = 2.31). The skunk global model included OOS, tree, camp, and park covariates (K = 6, χ2 = 395.80, P = 0.36, \(\widehat{c}\) = 1.07), and the opossum global model included res, com, camp, and park covariates (K = 6, χ2 = 400.34, P = 0.31, \(\widehat{c}\) = 1.10). The number of sites divided by the highest number of parameters led to the use of AICc for all single-species models (112/8 = 14; Table 3). The dog global model did not converge and resulted in very poor fit (K = 7, χ2 = 10456.71, P < 0.001, \(\widehat{c}\) = 35.44) and raccoon modeling resulted in no candidate model outperforming the null and large standard errors, so these two species were excluded from our single-species occupancy analysis.
We considered models with ΔAICc < 4.00 for cat single-species occupancy which resulted in nine models with a cumulative w of 95% (Table 3). These models included OOS and tree univariate models followed by the additive model tree + camp, four additive models including OOS and one other covariate, and finally two additive models including tree and one other covariate (Table 3). However, all covariates apart from OOS had β confidence intervals that crossed 0 so were not considered reliable covariates for cats (Table 4). In the top model, OOS had a relatively weak negative effect on cat occupancy (Table 4). We considered other open space to be the most important land use for cat occupancy in our analysis. Considering the OOS univariate model, the lowest occupancy probability for cats (0.08) occurred in the cell with the most open space (82% OOS) and the highest occupancy probability (1.00) occurred in cells where there was no open space (Table 5). The lowest occupancy probability for cats was still higher than the other modeled species and cats also had a higher detection probability than the other modeled species (Table 5).
We considered models with ΔAICc < 3.00 for skunk single-species occupancy which resulted in three models with a cumulative w of 72% (Table 3). The three top models included the additive models com + ind followed by res + com and OOS + ind. Across the top models, com and res had a weak negative effect on skunk occupancy and ind and OOS had a weak positive effect (Table 6). We considered the additive effect of commercial and industrial to be the most important land uses for skunk occupancy in our analysis. Considering the com + ind additive model, the lowest occupancy probability (0.02) occurred in a cell with no industrial land use and 41% commercial land use, while the highest occupancy probability (0.96) occurred in a cell with 66% industrial land use and 8% commercial land use (Table 5; Fig. 2).

Occupancy probability (ψ) for striped skunks in relation to percentage of industrial and commercial land uses in 112 1-km2 grid cells, using detection histories from annual surveys from 2015 to 2021 in Bakersfield, CA
We considered models with ΔAICc < 4.00 for opossum single-species occupancy which resulted in four models with a cumulative w of 75% (Table 3). These models included the additive model OOS + park followed by the park univariate model, and two other additive models, res + park and tree + park. However, tree had a β confidence interval that crossed 0 by 0.02 so was not considered a reliable covariate for opossums (Table 7). Across the considered models, OOS had a weak negative effect on opossum occupancy while res had a weak positive effect and park had a moderate positive effect (Table 7). We considered the additive effect of other open space and parks to be the most important land use for opossum occupancy in our analysis. From this model, the lowest occupancy probability (0.06) occurred in a cell with 74% open space and no parks, while the highest occupancy probability (0.98) occurred in a cell with 31% open space and 32% parks (Table 5; Fig. 3).

Occupancy probability (ψ) for Virginia opossums in relation to percentage of other open space and parks land uses in 112 1-km2 grid cells, using detection histories from annual surveys from 2015 to 2021 in Bakersfield, CA
Co-occurrence
We developed three co-occurrence models for dogs and kit foxes and four for cats and kit foxes (Online Resource 2). We designated the domestic species as the dominant species (Species A) in both model sets. The number of sites divided by the highest number of parameters also led to the use of AICc for co-occurrence modeling (112/10 = 11.20). Both co-occurrence model sets resulted in the null model as the single top model holding 71% of the model weight for dogs and kit foxes and 100% for cats and kit foxes (Table 8). Considering the null models, kit foxes had a positive occupancy probability given dog or cat presence (Table 9). Cats being present and detected had a slight negative effect on kit fox detection, though the same effect was not observed with dogs (Table 9). Ultimately, kit foxes occurred independently of dogs and cats (Table 10).
Discussion
Overall, kit foxes appear to occur independently of dogs and cats though there is some evidence of fine-scale spatial avoidance of cats by kit foxes. Cats are generally two or three times heavier than kit foxes and in Bakersfield, kit foxes have been observed deferring to cats at feeding stations established for feral cats (Cypher 2010). These results somewhat contradict our prediction that kit foxes would occur independently of cats but might avoid dogs. However, while there may be some avoidance of cats by kit foxes where possible, both are highly ubiquitous and probably mostly ignore each other (Cypher 2010). Despite the risk of predation by dogs, kit foxes may occur independently of dogs due to the influence humans have over dog location and activity. Because many people own pet dogs, they occur ubiquitously throughout the city and are often confined on leashes or within fences. Therefore, dogs may not be as threatening to kit foxes as other predators.
Among the other smaller mesocarnivores, there was some evidence of fine-scale spatial avoidance of dogs by opossums. Interactions between dogs and opossums are not well documented, though opossums may avoid dogs to some extent due to risk of predation. Opossums have also shown negative associations with dog presence in San Francisco Bay Area, CA (Reilly et al. 2016). Raccoons and skunks exhibited fine-scale spatial coexistence with cats. Positive associations between cats and smaller mesocarnivores may be explained by low competitive pressure due to none of the species being overtly territorial and all having opportunistic feeding patterns (Hopkins and Forbes 1980; Gardner 1982; Leyhausen 1979; Pearre and Maass 1998; Hadidian et al. 2010; Rosatte et al. 2010).
Cats are the most frequently detected urban carnivore in Bakersfield, followed by kit foxes and then dogs. The pervasive distributions of urban mesocarnivores, immense heterogeneity of the urban environment, and human influence over animal activity, particularly for domestic species, makes it difficult to discern clear occupancy patterns for the species in our study; however, we found open space to be an important negative predictor of cat and opossum occupancy. Cats and opossums are likely avoiding open spaces due to a lack of cover and increased risk of predation by coyotes or red foxes. Less human presence in open spaces also makes it less likely that cats and opossums will find accessible meals from people leaving out pet food. Cats and opossums may find refuge from predators in isolated fragments of more natural environments (e.g., parks and green space) surrounded by development in part due to mesopredator release, in which smaller carnivores are freed from larger, competitive predators that are unfit to inhabit small and isolated fragments (Crooks and Soulé 1999; Crooks 2002). It is unsurprising that dogs occurred more frequently along roadways than other species as pet dogs are routinely guided along sidewalks adjacent to roads while on walks with their owners. Raccoons, skunks, and opossums have been known to avoid smaller roadways while cats and kit foxes primarily display avoidance of major roadways; however, these avoidances are likely a result of the inhospitality and risk of mortality at roads and not a result of frequent dog occurrences (Barratt 1997; Prange et al. 2003; Bjurlin et al. 2005; Greenspan et al. 2018; Cypher et al. 2023).
We also found commercial and industrial areas to be important negative and positive predictors, respectively, of skunk occupancy. In Toronto, Ontario, both industrial and commercial areas have supported skunk densities as high as 36 individuals/km2, though it was presumed that close proximity to empty fields is what attracted skunks to those areas (Rosatte et al. 1991). In Bakersfield, industrial areas tend to abut open spaces or even encompass such land uses (e.g., large dirt roads or power line lots), and open space was also a positive predictor for skunks in our study. Skunks may refine their selection for industrial areas as refuge from neighboring heavily populated and developed commercial areas in Bakersfield, which might not have been possible for skunks at densities as high as 36 individuals/km2 in Toronto. These results, as well as our prediction that cats and opossums would share landscape preferences, are consistent with our predictions on urban land use selection by these species.
Overall positive or indifferent associations between domestic species and kit foxes or other smaller mesocarnivores could enhance the risk of disease transmission. Feeding stations for feral cats throughout Bakersfield are known to attract foxes, raccoons, skunks, and opossums (Cypher 2010). Primary transmittable diseases between the species include rabies, distemper, parvovirus, leptospirosis, and a variety of endo and ectoparasites (Baker et al. 2010). Disease may spread through the environment, direct contact, waste material, or arthropod vectors (Baker et al. 2010). An aggressive skin disease called sarcoptic mange, caused by an infectious mite (Sarcoptes scabiei), has been detected in urban kit foxes in Bakersfield since 2013 and is fatal for kit foxes without intervention (Rudd et al. 2020). Our camera surveys have documented a decline in the once robust Bakersfield kit fox population by nearly 70% since 2015 (Kelly et al. 2022). The outbreak in kit foxes has been attributed to a potential spillover event from dogs or coyotes (Rudd et al. 2020). Efforts are underway to monitor, capture, and treat kit foxes with sarcoptic mange, though response can be costly and time consuming as advanced cases require keeping individuals in captivity for at least one month of intensive treatment (Cypher et al. 2017). Vaccinations have proven to be an effective management technique against the spread of other diseases, particularly rabies, among these species, though large-scale vaccinations and treatment of wild carnivores in urban areas is novel and challenging due to the fragmented landscapes and risks to humans, pets, and other wildlife (Baker et al. 2010; Hadidian et al. 2010; Soulsbury et al. 2010).
Wild mesocarnivores living in Bakersfield exhibited varying degrees of spatial partitioning with domestic dogs and cats. The apparent indifference of kit foxes towards domestic species may help kit foxes persist more widely and in higher densities in the urban environment than other urban mesocarnivores. The conservation of urban populations of kit foxes can be beneficial for the long-term persistence of this species as their natural habitat continues to be developed. Conservation initiatives should explore disease prevention in detail, particularly at cat feeding stations or other urban sites where the species may congregate. Because both wild and domestic carnivores commonly co-occur within urban environments, understanding their adaptations to urbanization and novel sympatry allows for management strategies that facilitate coexistence and promote conservation of the wild species, particularly San Joaquin kit foxes.
Availability of data and material
Data for this manuscript are provided in supplementary information and can be available upon request.
References
Ansell RJ (2004) The spatial organization of a red fox (Vulpes vulpes) population in relation to food resources. Ph.D. Dissertation, University of Bristol, Bristol, England
Baker PJ, Soulsbury CD, Iossa G, Harris S (2010) Domestic cat (Felis catus) and domestic dog (Canis familiaris). In: Gehrt SD, Riley SPD, Cypher BL (eds) Urban carnivores, ecology, conflict, and conservation. Baltimore, Maryland, pp 156–171
Barratt DG (1997) Home range size, habitat utilisation and movement patterns of suburban and farm cats Felis catus. Ecography 20:271–280. https://doi.org/10.1111/j.1600-0587.1997.tb00371.x
Beck AM (1973) The Ecology of stray dogs: a study of free-ranging urban animals. West Lafayette, Indiana
Berman M, Dunbar I (1983) The social behavior of free-ranging suburban dogs. Appl Anim Ethol 10:5–17
Bjurlin CD, Cypher BL, Wingert CM, Van Horn Job CL (2005) Urban roads and the endangered San Joaquin kit fox. California Department of Transportation, Endangered Species Recovery Program Final Report, Contract Number 65A0136. Fresno, California
Bozek CK, Prange S, Gehrt SD (2007) The influence of anthropogenic resources on multi-scale habitat selection of raccoons. Urban Ecosyst 10:413–425. https://doi.org/10.1007/s11252-007-0033-8
Bradley CA, Altizer S (2006) Urbanization and the ecology of wildlife diseases. Trends Ecol and Evol 22:95–102. https://doi.org/10.1016/j.tree.2006.11.001
Burnham KP, Anderson DR (2002) Model selection and multimodal inference: a practical information theoretic, 2nd edn. New York, New York
Crooks KR (2002) Relative sensitivities of mammalian carnivores to habitat fragmentation. Conserv Biol 16:488–502. https://doi.org/10.1046/j.1523-1739.2002.00386.x
Crooks KR, Riley SPD, Gehrt SD, Gosselink TE, Van Deelen TR (2010) Community ecology of urban carnivores. In: Gehrt SD, Riley SPD, Cypher BL (eds) Urban carnivores, ecology, conflict, and conservation. Baltimore, Maryland, pp 184–196
Crooks KR, Soulé ME (1999) Mesopredator release and avifaunal extinctions in a fragmented system. Nat 400:563–566. https://doi.org/10.1038/23028
Cypher BL (2003) Foxes. In: Feldhamer GA, Thompson BC, Chapman JA (eds) Wild mammals of North America: Biology, management, and conservation, 2nd edn. Baltimore, Maryland, pp 511–546
Cypher BL (2010) Kit foxes (Vulpes macrotis). In: Gehrt SD, Riley SPD, Cypher BL (eds) Urban carnivores, ecology, conflict, and conservation. Baltimore, Maryland, pp 48–61
Cypher BL, Deatherage NA, Westall TL, Kelly EC, Philips SE (2023) Potential habitat and carrying capacity of endangered San Joaquin kit foxes in an urban environment: implications for conservation and recovery. Urban Ecosyst 26:173–183. https://doi.org/10.1007/s11252-022-01287-8
Cypher BL, Rudd JL, Westall TL, Woods LW, Stephenson N, Foley JR, Richardson D, Clifford DL (2017) Sarcoptic mange in endangered kit foxes (Vulpes macrotis mutica): Case histories, diagnoses, and implications for conservation. J Wildl Dis 53:46–53. https://doi.org/10.7589/2016-05-098
Deatherage NA, Cypher BL, Murdoch J, Westall TL, Kelly EC, Germano DJ (2021) Urban landscape attributes affect occupancy patterns of the San Joaquin kit fox during an epizootic. Pac Conserv Biol 27:256–266. https://doi.org/10.1071/PC20059
Farias V, Fuller TK, Wayne RK, Sauvajot RM (2005) Survival and cause-specific mortality of grey foxes (Urocyon cinereoargenteus) in southern California. Zool Soc Lond 266:249–254. https://doi.org/10.1017/S0952836905006850
Fidino MA, Lehrer EW, Magle SB (2016) Habitat dynamics of the Virginia opossum in a highly urban landscape. Am Midl Nat 175:155–167. https://doi.org/10.1674/0003-0031-175.2.155
Frost N (2005) San Joaquin kit fox home range, habitat use, and movements in urban Bakersfield. MS Thesis, Humboldt State University, Arcata, California
Fuller TK, Destefano S, Warren PS (2010) Carnivore behavior and ecology, and relationship to urbanization. In: Gehrt SD, Riley SPD, Cypher BL (eds) Urban carnivores, ecology, conflict, and conservation. Baltimore, Maryland, pp 18–19
Gardner AL (1982) Virginia Opossum. In: Chapman JA, Feldhamer GA (eds) Wild mammals of North America. Baltimore, Maryland, pp 3–36
Gehrt SD (2004) Ecology and management of striped skunks, raccoons, and coyotes in urban landscapes. In: Fascione N, Delach A, Smith M (eds) Predators and people: from conflict to conservation. Washington, DC, pp 81–104
Gehrt SD (2010) The urban ecosystem. In: Gehrt SD, Riley SPD, Cypher BL (eds) Urban carnivores, ecology, conflict, and conservation. Baltimore, Maryland, pp 2–11
Gehrt SD, Riley SPD (2010) Coyotes (Canis latrans). In: Gehrt SD, Riley SPD, Cypher BL (eds) Urban carnivores, ecology, conflict, and conservation. Baltimore, Maryland, pp 85–89
Gosselink TE, Van Deelen TR, Warner RE, Mankin PC (2007) Survival and cause-specific mortality of red foxes in agricultural and urban areas of Illinois. J Wildl Manag 71:1862–1873. https://doi.org/10.2193/2006-020
Gotelli NJ, Ellison AM (2013) A primer of ecological statistics. Sunderland, Massachusetts
Greenspan E, Nielsen CK, Cassel KW (2018) Potential distribution of coyotes (Canis latrans), Virginia opossums (Didelphis virginiana), striped skunks (Mephitis mephitis), and raccoons (Procyon lotor) in the Chicago Metropolitan Area. Urb Ecosyst 21:983–997. https://doi.org/10.1007/s11252-018-0778-2
Hadidian J, Prange S, Rosatte R, Riley SPD, Gehrt SD (2010) Raccoons (Procyon lotor). In: Gehrt SD, Riley SPD, Cypher BL (eds) Urban carnivores, ecology, conflict, and conservation. Baltimore, Maryland, pp 34–47
Harris S (1981) The food of suburban foxes (Vulpes vulpes) with special reference to London. J Zool 187:195–199. https://doi.org/10.1111/j.1365-2907.1981.tb00003.x
Hines JE (2006) PRESENCE– Software to estimate patch occupancy and related parameters. USGS–PWRC. http://www.mbr-pwrc.usgs.gov/software/presence.html
Hopkins DD, Forbes RB (1980) Dietary patterns of the Virginia opossum in an urban environment. Murrelet 61:20–30
Iossa G, Soulsbury CD, Baker PJ, Harris S (2010) A taxonomic analysis of urban carnivore ecology. In: Gehrt SD, Riley SPD, Cypher BL (eds) Urban carnivores, ecology, conflict, and conservation. Baltimore, Maryland, pp 172–180
Kamler JF, Keeler K, Wiens G, Richardson C, Gipson PS (2003) Feral dogs, Canis familiaris, kill coyote, Canis latrans. Can Field-Nat 177:123–124
Kapfer JM, Kirk RW (2012) Observations of gray foxes (Urocyon cinereoargenteus) in a suburban landscape in the Piedmont of North Carolina. Southeast Nat 11:507–516. https://doi.org/10.1656/058.011.0313
Kelly EC, Cypher BL, Westall TL, Deatherage NA (2022) Sarcoptic mange in urban kit foxes: potential for cross-species transmission. Proceedings of the Vertebrate Pest Conference 30:18
Kern County Animal Services (2019) Director’s yearly report. https://www.kerncountyanimalservices.org/media/1795/monthlyreport201912.pdf. Accessed 16 Mar 2023
Legendre P, Legendre L (1998) Numerical ecology, 2nd edn. The Netherlands, Amsterdam
Lewis JC, Sallee KL, Golightly RT (1993) Introduced red fox in California. Department of Fish and Game Wildlife Management Division, Nongame Bird and Mammal Section Report 93–10. Sacramento, California
Leyhausen P (1979) Cat behaviour: The predatory and social behaviour of domestic and wild cats. England, London
Lombardi JV, Comer CE, Scognamillo DG, Conway WC (2017) Coyote, fox, and bobcat response to anthropogenic and natural landscape features in a small urban area. Urban Ecosyst 20:1239–1248. https://doi.org/10.1007/s11252-017-0676-z
MacKenzie DI, Nichols JD, Royle JA, Pollock KH, Bailey LL, Hines JE (2018) Occupancy estimation and modeling, inferring patterns and dynamics of species occurrence, 2nd edn. San Diego, California
National Oceanic and Atmospheric Administration (2020) Bakersfield climate. https://www.weather.gov/hnx/bflmain. Accessed 28 June 2020
Parsons AW, Rota CT, Forrester T, Baker-Whatton MC, McShea WJ, Schuttler SG, Millspaugh JJ, Kays R (2019) Urbanization focuses carnivore activity in remaining natural habitats, increasing species interactions. J Appl Ecol 56:1894–1904. https://doi.org/10.1111/1365-2664.13385
Pearre S Jr, Maass R (1998) Trends in prey size-based trophic niches of feral and house cats Felis catus L. Mammal Rev 28:125–139. https://doi.org/10.1046/j.1365-2907.1998.00030.x
Prange S, Gehrt SD, Wiggers EP (2003) Demographic factors contributing to high raccoon densities in urban landscapes. J Wildl Manag 67:324–333. https://doi.org/10.2307/3802774
Primack RB, Morrison RA (2013) Extinction, causes of. In: Levin SA (ed) Encyclopedia of biodiversity, 2nd (edn). The Netherlands, Amsterdam, pp 401–412
Reilly ML, Tobler MW, Sonderegger DL, Beier P (2016) Spatial and temporal response of wildlife to recreational activities in the San Francisco Bay ecoregion. Biol Conserv 207:117–126. https://doi.org/10.1016/j.biocon.2016.11.003
Riley SPD, Gehrt SD, Cypher BL (2010) Urban carnivores: Final perspectives and future directions. In: Gehrt SD, Riley SPD, Cypher BL (eds) Urban carnivores, ecology, conflict, and conservation. Baltimore, Maryland, pp 222–232
Rosatte RC, Lariviére S (2003) Skunks (Genera Mephitis, Spilogale, and Conepatus). In: Feldhamer GA, Thompson BC, Chapman JA (eds) Wild mammals of North America: biology, management, and conservation, 2nd edn. Baltimore, Maryland, pp 692–707
Rosatte RC, Power MJ, MacInnes CD (1991) Ecology of urban skunks, raccoons, and foxes in metropolitan Toronto. In: Adams LW, Leedy DL (eds) Wildlife conservation in metropolitan environments. Columbia, Maryland, pp 31–38
Rosatte R, Sobey K, Dragoo JW, Gehrt SD (2010) Striped skunks and allies (Mephitis spp.). In: Gehrt SD, Riley SPD, Cypher BL (eds) Urban carnivores, ecology, conflict, and conservation. Baltimore, Maryland, pp 96–106
Rudd JL, Clifford DL, Cypher BL, Hull JM, Jane Riner A, Foley JE (2020) Molecular epidemiology of a fatal sarcoptic mange epidemic in endangered San Joaquin kit foxes (Vulpes macrotis mutica). Parasit Vectors. https://doi.org/10.1186/s13071-020-04328-3
Schoener TW (1974) Resource partitioning in ecological communities. Science 185:27–39. https://doi.org/10.1126/science.185.4145.27
Shedden JM, Bucklin DM, Quinn NM (2020) Do coyotes eat mesocarnivores in Southern California? A molecular genetic analysis. Proceedings of the Vertebrate Pest Conference 29:64
Soulsbury CD, Baker PJ, Iossa G, Harris S (2010) Red foxes (Vulpes vulpes). In: Gehrt SD, Riley SPD, Cypher BL (eds) Urban carnivores, ecology, conflict, and conservation. Baltimore, Maryland, pp 63–75
Stachelski C, Sanger G (2008) The climate of Bakersfield, California. NOAA Technical Memorandum NWS WR-281. Salt Lake City, Utah
United Nations, Department of Economic and Social Affairs, Population Division (2019) World urbanization prospects: The 2018 revision (ST/ESA/SER.A/420). New York: United Nations
United States Fish and Wildlife Service (1998) Recovery plan for upland species of the San Joaquin Valley, California. United States Fish and Wildlife Service, Region 1. Portland, Oregon
Westall TL, Cypher BL (2017) Latency to first detection of kit foxes (Vulpes macrotis) during camera surveys. Canid Biol Conserv 20:32–37
Wright JD, Burt MS, Jackson VL (2012) Influences of an urban environment on home range and body mass of Virginia opossums (Didelphis virginiana). Northeast Nat 19:77–86. https://doi.org/10.1656/045.019.0106
Acknowledgements
We thank Christine Van Horn Job and Larry Saslaw with the Endangered Species Recovery Program for assistance with camera surveys. We appreciate the dozens of local business owners and employees, school districts and campuses, churches, city and county staff, and many local residents for use of their property to place cameras. Funding for this project was provided by The Morris Animal Foundation (grant D17ZO-066), the U.S. Bureau of Reclamation’s Central Valley Project Conservation Program (grant R19AP000068), the Sacramento Field Office of the United States Fish and Wildlife Service, and the California Department of Fish and Wildlife.
Funding
Funding for this project was provided by The Morris Animal Foundation (grant D17ZO-066), the U.S. Bureau of Reclamation’s Central Valley Project Conservation Program (grant R19AP000068), the Sacramento Field Office of the United States Fish and Wildlife Service, and the California Department of Fish and Wildlife.
Author information
Authors and Affiliations
Contributions
Nicole A. Deatherage and Brian L. Cypher contributed to the study conception and design and performed data analysis. Material preparation and data collection were performed by all authors. The first draft of the manuscript was written by Nicole A. Deatherage and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval
No approval of research ethics committees was required to accomplish the goals of this study because all data was collected non-invasively via remote cameras and did not require direct interaction with the species.
Competing interests
The authors declare no competing interests.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Deatherage, N.A., Cypher, B.L., Westall, T.L. et al. Spatial relationships between mesocarnivores and domestic species in an urban environment and implications for endangered San Joaquin kit foxes. Urban Ecosyst 27, 321–334 (2024). https://doi.org/10.1007/s11252-023-01447-4
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-023-01447-4