
Abstract
Nerocila japonica Schioedte & Meinert, 1881 is redescribed based on the holotype and specimens from various localities in Japanese waters. The following fishes are recorded as new hosts: Tribolodon hakonensis (Cyprinidae), Mugil cephalus, Liza affinis, Chelon haematocheilus (Mugilidae), Lateolabrax japonicus, L. latus (Lateolabracidae), Acanthopagrus latus, A. schlegelii schlegelii (Sparidae), Rhyncopelates oxyrhynchus (Terapontidae), Ditrema viride, D. temminckii temminckii (Embiotocidae), Chaenogobius gulosus, Acanthogobius flavimanus (Gobiidae), Pseudolabrus sp. (Labridae) and Aluterus monoceros (Monacanthidae). Specimens previously recorded as ‘Nerocila acuminata’ from Toyama Bay (the Sea of Japan) were examined, when available, and re-identified as N. japonica. As there has been no reliable record of N. acuminata Schioedte & Meinert, 1881 from off Japan, reports of this species in Japanese waters are considered to represent N. japonica.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Nerocila Leach, 1818 is a genus of cymothoid isopod with almost all of its members occurring as the body surface parasites of fishes (Bruce, 1987). This genus comprises 43 species (Bruce & Schotte, 2009), three of which are known from off Japan (Saito et al., 2000): N. acuminata Schioedte & Meinert, 1881, N. japonica Schioedte & Meinert, 1881 and N. phaiopleura Bleeker, 1857. Of these, N. acuminata and N. japonica belong to the ‘Nerocila orbignyi species-group’ (Bruce, 1987).
Nerocila acuminata is known as an East Pacific and Caribbean species (Brusca, 1981). However, it has been recorded from Japanese waters (Iwasa, 1947; Shiino, 1965; Honma et al., 1974; Nunomura, 1981, 1985). Bruce (1987) suspected that N. acuminata, as reported by Nunomura (1981), might actually represent N. japonica.
Nerocila japonica was originally described on the basis of a non-ovigerous female collected from “mare Iaponico” (= Japanese waters) (Schioedte & Meinert, 1881). Since then, N. japonica has been mentioned in the literature several times (Thielemann, 1910; Nierstrasz, 1918, 1931; Gurjanova, 1936; Bruce, 1987). Recently, additional specimens of this species were reported by Yu & Li (2003) from Chinese waters and by Hashimoto (2007) from Pacific waters off Kochi Prefecture, Shikoku, Japan. However, limited information has so far been available on N. japonica.
In this study, we redescribe N. japonica based on the holotype and specimens from various localities in Japanese waters. We also discuss the previous records of N. acuminata from off Japan.
Materials and methods
Specimens were preserved in 70% ethanol or 10% formalin. Appendages were dissected from the isopods with needles. Photographs were made with a digital camera and drawings were made with the aid of drawing tubes attached to microscopes (Nikon SMZ800 and Nikon ECLIPSE 80i).
The isopod classification used follows Brandt & Poore (2003). The common and scientific names of fishes follow those recommended by Froese & Pauly (2009). Those specimens examined are deposited in the following museums: Swedish Museum of Natural History, Stockholm (SMNH), Toyama Science Museum, Toyama (TOYA), Himeji City Aquarium, Himeji (HCA), Osaka Museum of Natural History, Osaka (OMNH), and Kitakyushu Museum of Natural History and Human History, Kitakyushu (KMNH).
Suborder Cymothoida Wägele, 1989
Family Cymothoidae Leach 1814
Genus Nerocila Leach, 1818
Nerocila japonica Schioedte & Meinert, 1881
Syns Nerocila acuminata of Nunomura (1985), Saito et al. (2000), possibly Honma et al. (1974), possibly Nunomura (1981) [nec N. acuminata Schioedte & Meinert, 1881]
Type-material: Holotype: 1 non-ovigerous female (19.5 mm), Japan, coll. R. M. Salmin (SMNH Type-8206 = NRM 4975).
Material examined: [Sea of Japan] 1 non-ovig. female (21.5 mm), on pectoral fin of Mugil cephalus Linnaeus, Uozu Aquarium, Uozu, Toyama, 4 Dec. 1979, coll. Y. Kano (TOYA-Cr 23229); 1 ovig. female (19.0 mm), 1 male (16.0 mm), on pectoral and caudal fins of M. cephalus, Uozu, Toyama, 4 Dec. 1974, coll. Y. Kano (TOYA-Cr 23230, 23231); 1 male (19.5 mm), on pectoral fin of Acanthopagrus schlegelii schlegelii (Bleeker), Uozu, Toyama, 2 Jun. 1977, coll. Y. Kano (TOYA-Cr 23232); 1 ovig. female (21.0 mm), on body surface of Ditrema temminckii temminckii Bleeker, off Namerikawa, Toyama, 18 Jul. 1974, coll. Y. Kano (TOYA-Cr 23233); 1 ovig. female (18.5 mm), on caudal fin of D. t. temminckii, Uozu South Port, Uozu, Toyama, 1 Oct. 1975, coll. Y. Kano (TOYA-Cr 23234); 1 non-ovig. female (23.0 mm), on pectoral fin of Pseudolabrus sp. [P. eoethinus (Richardson) or P. sieboldi Mabuchi & Nakabo] (22.0 cm total length [TL]), Uozu South Port, Uozu, Toyama, 24 Aug. 2001, coll. Y. Kano (TOYA-Cr 23235); 2 non-ovig. females (19.0, 22.5 mm), 3 males (15.0, 18.0, 19.0 mm), 1 aegathoid (9.5 mm) from body surface of Tribolodon hakonensis (Günther), 10 Mar. 2003, off Ômizaki, Lake Nakaumi, Shimane, coll. K. Yamaguchi et al. (TOYA-Cr 23236–23241); [Seto Inland Sea] 4 ovig. females (25.0, 25.0, 26.0, 29.0 mm), ex. Mugil cephalus, Dejima, Sakai, Osaka, Jun. 1978, coll. Y. Nabeshima (OMNH Ar6599); 1 non-ovig. female (22.5 mm), from mouth cavity of Lateolabrax japonicus (Cuvier & Valenciennes), off Sakai, Osaka Bay, Osaka, 5 Mar. 1976, coll. Y. Hayashi (TOYA-Cr 23242); 1 non-ovig. female (32.0 mm), on pectoral fin of M. cephalus, Shimo-kamagari Island, Hiroshima, 28 Dec. 2005, coll. T. Umino (TOYA-Cr 23243); 2 females (not measured), on pectoral fin of Acanthopagrus latus (Houttuyn), Hiroshima Bay, Hiroshima, 25 May 2007, coll. T. Umino (TOYA-Cr 23244, 23245); 1 non-ovig. female (23.0 mm), on pectoral fin of A. s. schlegelii, Ôkurokami Island, Hiroshima, 13 Jun. 2006, coll. K. Nagasawa (TOYA-Cr 23246); 1 non-ovig. female (24.0 mm), on the caudal surface of pectoral fin of A. s. schlegelii, Shimo-kamagari Island, Hiroshima, 28 Dec. 2005, coll. T. Umino (TOYA-Cr 23247); 1 ovig. female (33.0 mm), 1 non-ovig. female (20.5 mm), on caudal fin of A. s. schlegelii, Shimo-kamagari Island, Hiroshima, 28 Dec. 2005, coll. T. Umino (TOYA-Cr 23248, 23249); 1 ovig. female (27.0 mm), ex. A.s. schlegeli (29.0 cm TL), Tamano, Okayama, 21 Jun. 2007, coll. A. Nishimura (TOYA-Cr 23250); 1 non-ovig. female (24.0 mm), on body surface of A. s. schlegelii, off Sakai, Osaka, 31 May 1999, coll. H. Ariyama (OMNH Ar6166); 1 female (not measured), on caudal fin of Ditrema viride Oshima, Aboshi, Himeji, Hyogo, 8 Jan. 2003, coll. T. Miki (the specimen is no longer extant); 1 ovig. female (25.0 mm), 5 males (15.0, 16.5, 17.5, 18.0, 20.5 mm), on caudal fin of Lateolabrax japonicus and D. viride, Aboshi, Himeji, Hyogo, 18 May 2004, coll. T. Miki (HCA-H No.17); 1 ovig. female (24.0 mm), at pectoral fin base of Chaenogobius gulosus (Sauvage), Onomichi Fish Market, Hiroshima, 12 Jul. 1960, coll. Y. Shibata (OMNH); 1 ovig. female (21.0 mm), on pectoral fin of Acanthogobius flavimanus (Temminck & Schlegel) (20.0 cm TL), Misaki, Osaka, 7. Sep. 2005, coll. A. Aimoto (TOYA-Cr 23251); 1 ovig. female (19.0 mm), 1 aegathoid (10.5 mm), on pectoral fin of A. flavimanus (23.5 cm TL), Kawajiri, Hiroshima, 1 Jan. 2004, coll. T. Morihisa (TOYA-Cr 23252, 23253); 1 non-ovig. female (23.0 mm), host unknown, Hiroshima Bay, Hiroshima, Nov. 2007, Coll. K. Nagasawa (TOYA-Cr 23254); 5 non-ovig. female (9.0, 10.0, 16.0, 18.0, 20.0 mm), 1 male (15.0 mm), on pectoral fin of Chelon haematocheilus (Temminck & Schlegel), Kanda Port, Fukuoka, 21 Dec. 1976, coll. K. Izawa (TOYA-Cr 23255–23260); [Ariake Sea] 1 non-ovig. female (27.0 mm), on pectoral fin of Lateolabrax japonicus (50.4 cm TL, age 4, FAKU129257), unknown locality, Dec. 2000, coll. N. Nakadachi (TOYA-Cr 23261); 1 non-ovig. female (16.0 mm), Mar. 2006, on caudal fin of L. japonicus, Yatsushiro, Kumamoto, coll. H. Yoshigou (TOYA-Cr 23262); 1 ovig. female (34.0 mm), on caudal fin of L. latus Katayama (70.0 cm TL), Tomioka, Kumamoto, 3 Apr. 2004, coll. S. Arakaki (TOYA-Cr 23263); 1 non-ovig. female (18.0 mm), on pectoral fin of L. latus (62.0 cm TL), Tomioka, Kumamoto, 19 Jan. 2004, coll. S. Arakaki (TOYA-Cr 23264); 1 non-ovig. female (18.0 mm), on anal fin of L. latus (60.0 cm TL), Tomioka, Kumamoto, 11 Mar. 2004, coll. S. Arakaki (TOYA-Cr 23265); 1 female (19.0 mm), on caudal fin of Liza affinis (Günther) (22.5 cm TL), 11 Mar. 2008, mouth of Saza River, Saza, Nagasaki, coll. H. Yoshigou (TOYA-Cr 23266); [North Pacific Ocean] 1 ovig. female (16.0 mm), on body surface of Acanthogobius flavimanus, 10 Oct. 1992, mouth of Hatauchi River, Shimizu, Shizuoka, coll. T. Kato (KMNH IvR 500,538); 1 ovig. female (41.0 mm), on pectoral fin of Lateolabrax japonicus (73.0 cm TL), Mongawa Bay, Miyazaki, at depth of 8 m, 24 Apr. 2007, coll. M. Wada (the specimen is no longer extent); 1 non-ovig. female (34.0 mm), on caudal fin of L. latus (31.5 cm TL), Mongawa Bay, Miyazaki, at depth of 8 m, 8 Jun. 2007, coll. M. Wada (TOYA-Cr 23267); 1 ovig. female (32.0 mm), on pectoral fin of Acanthopagrus s. schlegelii (30–35 cm TL), Mongawa Bay, Miyazaki, at depth of 6 m, 21 May 2007, coll. M. Wada (TOYA-Cr 23268); [East China Sea] 1 non-ovig. female (20.0 mm), 8 Aug. 2005, on pectoral fin of Rhyncopelates oxyrhynchus (Temminck & Schlegel), off Kataura, Kasasa, Kagoshima, at depth of 27 m, 8 Aug. 2005, coll. M. Ito (TOYA-Cr 23269); 1 ovig. female (23.0 mm), on pectoral fin of Aluterus monoceros (Linnaeus), off Kataura, Kasasa, Kagoshima, at depth of 27 m, 23 Sep. 2005, coll. M. Ito (TOYA-Cr 23270).
Material used in previous identifications: [Sea of Japan] 1 ovig. female (21.5 mm), Ao, Himi, Toyama, 18 Sep. 1979, coll. Toyama Prefectural Fisheries Experiment Station (TOYA Z80-189, recorded as Nerocila acuminata in Nunomura, 1985); 1 ovig. female (26.5 mm), host unknown, Kokufu, Himi, Toyama, 9 Jun. 1974, coll. S. Tanaka (TOYA Z82-76, recorded as N. acuminata in Nunomura, 1985).
Redescription (Figs. 1–7)
Holotype (non-ovigerous female)

Holotype of Nerocila japonica, non-ovigerous female (19.5 mm; SMNH Type-8206). A, habitus, dorsal; B, habitus, ventral; C, habitus, lateral; D, frons, ventral; E, pleon and pleotelson, dorsal. Scales: in mm
Body c.1.75 times as long as maximum width, bilaterally symmetrical (Fig. 1A); pereon dorsum vaulted (Fig. 1C). Cephalon anterior margin evenly rounded (Fig. 1D), without indistinct medial point; eye obscured. Pereonites 1–4 with posterolateral angles produced; pereonites 5–7 with posterolateral angles strongly produced, acute (Fig. 1C). Coxae of pereonites (Fig. 1B,C) with posterior margins acute. Ventrolateral margins of pleonites 1 and 2 (Fig. 1B) posteriorly directed, extending to pleonite 5. Pleotelson (Fig. 1E) shield-shaped, with lateral margins converging to caudomedial point.
Antennule (Fig. 1D) with 8 articles. Antenna with 6 articles (apical articles absent) (Fig. 1D).
Pereopods 1–6 with robust dactyli (Fig. 1B). Pereopod 1 dactylus (Fig. 1B) c.1.6 times as long as propodus. Dactyli of pereopods 4 and 5 with weak swelling (Fig. 1B). Pereopods 6 and 7 with weakly developed carina on bases (Fig. 1B); pereopod 7 with robust setae on posterior margin of ischium, merus, carpus and propodal palm (Fig. 1B).
Uropod exopod curving medially (Fig. 1E), c.1.3 times as long as endopod; endopod straight, with distal margin smoothly tapered.
Ovigerous female
Body c.2.0 times as long as maximum width, bilaterally symmetrical (Fig. 2A,C); pereon dorsum vaulted (Fig. 3A–B). Cephalon anterior margin rounded, with or without indistinct medial point (Fig. 4A); eye obscured. Pereonites 1–4 with posterolateral angles produced or not; pereonites 5–7 with posterolateral angles produced, acute (Figs. 2A,C, 3A,B). Coxae of pereonites 2–4 with posterior margins rounded or acute; coxae of pereonites 5–7 with posterior margins acute. Ventrolateral margins of pleonites 1 and 2 posteriorly directed, extending to or beyond pleonite 5 (Figs. 2A,C, 3A,B). Pleotelson (Fig. 2A,C) shield-shaped, with lateral margins converging to caudomedial point.

Nerocila japonica, habitus, dorsal. A, ‘acuminata form’, ovigerous female (32.0 mm; TOYA-Cr 23268); B, ‘acuminata-aster intermediate form’, non-ovigerous female (19.0 mm; TOYA-Cr 23236); C, ‘aster form’, ovigerous female (18.5 mm; TOYA-Cr 23234); D, male (19.5 mm; TOYA-Cr 23232); E, aegathoid (10.5 mm; TOYA-Cr 23253). Scale-bars: 5 mm

Nerocila japonica, ovigerous female, habitus, lateral. A, ‘acuminata form’ (32.0 mm; TOYA-Cr 23268); B, ‘aster form’ (18.5 mm; TOYA-Cr 23234). Scale-bars: 5 mm

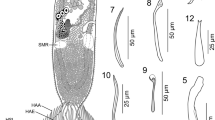
Nerocila japonica, ovigerous female (24.0 mm; OMNH Ar6166). A, head, dorsal; B, frons, ventral; C, left antennule, dorsal; D, left antenna, dorsal; E, left mandible, ventral; F, left maxillule, ventral; G, left maxilla, ventral; H, left maxilliped, ventral. Scale-bars: A–D, 2 mm; E,H, 1 mm; F,G, 0.1 mm
Antennule with 8 articles; article 1 and 2 partly fused (Fig. 4C). Antenna with 9 articles (Fig. 4D). Mandibular palp of article 3 with 8 setae on distolateral margin (Fig. 4E). Maxillule with 4 spines (Fig. 4F). Maxilla with 2 spines, each on medial and lateral lobes (Fig. 4G). Maxilliped article 3 with 6 spines (Fig. 4H).
Pereopods 1–6 with robust dactyli (Fig. 5A,B). Dactylus of pereopod 1 (Fig. 5A) c.1.7 times as long as propodus. Dactyli of pereopods 1, 2, 4 and 5 with or without weak swelling (Fig. 5A). Pereopods 6 and 7 with weakly developed carina on bases (Fig. 5B,C); pereopod 7 with 1-2 robust setae on posterior margin of ischium, 3 robust setae on posterior margin of merus, 3–8 robust setae on posterior margin of carpus and 7–11 robust setae on propodal palm (Fig. 5C).

Nerocila japonica, ovigerous female (24.0 mm; OMNH Ar6166). A, left pereopod 1, medial; B, left pereopod 6, medial; C, left pereopod 7, medial; D, pleopod 2, ventral; E, pleopod 5, medial; F, left uropod, dorsal. Scale-bars: 2 mm
Appendix masculina of pleopod 2 (Fig. 5D) c.0.6 length of endopod; pleopod 5 endopod with 2 large folds (Fig. 5E). Uropod exopod curves medially (Fig. 5F), c.1.3 times as long as endopod; endopod straight, with distal margin smoothly tapered.
Male
Body c.2.4 times as long as wide (Fig. 2D) and c.3.6 times in aegathoids (Fig. 2E). Eyes large and distinct in small individuals, but facets indistinct in large individuals. In males and aegathoids, coxae and posterolateral margins of pereonites not as produced as in females. Uropod morphology similar to that of females in large males, but uropod endopod with convex median margin in small males and aegathoids.
Coloration. Dark blue, with 2 faint submedian pale longitudinal bands visible in life and in alcohol.
Size. 15.0–20.5 (mean 17.3) mm in males (n = 11) and 9.0–41.0 (22.9) mm in females (n = 40).
Variation. Bruce (1987) stated that in N. orbignyi (Guerin-Maneville, 1832) there is considerable variation in the degree of prolongation of the posterolateral angles of the pereonites, in coxal shape and in pleonite morphology. This character cannot be considered as of specific significance. The ventrolateral processes of pleonites 1 and 2 may project away from the pleon, and extend posteriorly to pleonite 4 or beyond pleonite 5. The posterior coxae may have concave, convex or straight posterior margins.
There are two distinct morphological forms in N. japonica: the ‘acuminata form’ (Figs. 2A, 3A) and the ‘aster form’ (Figs. 2C, 3B), as occurs in N. acuminata (see Brusca 1981). In addition, intermediates (Fig. 2B) between the two forms were found. Hence, significant variation is found in the posterolateral margins of pereonites 1–5 and the coxae of these pereonites.
Remarks
The general characteristics of the newly collected ovigerous females correspond with the morphology of the holotype of Nerocila japonica.
Nerocila acuminata and N. orbignyi, members of the ‘Nerocila orbignyi group’, have the uropod endopod with a distomedial point (Bruce, 1987). We confirmed that N. japonica is distinguishable from other species in the ‘N. orbignyi group’ by having a uropod endopod with straight margins which converge to an acute apex, as indicated by Bruce (1987). Although the acute lateral margins on pleonites 3–5, the acute posterolateral margins of pereonites 1–4 and the acute coxae of pereonites 2–4 have been reported as diagnostic characters (Bruce, 1987, p. 383, figs. 34G–J), they have proved to be unreliable for separating N. japonica from the other species due to the above-mentioned intraspecific variation.
Sato (2001) showed colour photographs of an unidentified isopod attached on the pelvic fin of Lates japonicus Katayama & Taki collected from Tokushima Prefecture, Shikoku, Japan. There is little doubt that the isopod represents N. japonica based on the shape of its uropod endopod.
Bruce (1987, p. 400) suspected that N. acuminata reported from Japan (Nunomura, 1981) might actually represent N. japonica. Based on the previous records of N. acuminata in Japan, we discuss their synonymy with N. japonica in detail below.
Distribution
Nerocila japonica has been recorded from off Japan (Schioedte & Meinert, 1881; Hashimoto, 2007; Present study), China (Yu & Li, 2003) and Penang, Malaysia (Nierstrasz, 1918). In Japanese waters, N. japonica was collected from the coast of the Sea of Japan off Honshu (Toyama Bay and Lake Nakaumi), the Pacific coast from Suruga Bay to Kyushu, the Seto Inland Sea and the East China Sea coast off Kyushu (Fig. 6). Specimens of this species were found on the fish taken at the depths of 6–27 m.

Distributional records of Nerocila japonica in Japan. Closed and open circles represent localities reported in this paper and by previous authors (Sato, 2001; Hashimoto, 2007), respectively
Hosts
Nerocila japonica is known to infect Lates japonicus (Latidae) (Hashimoto, 2007). In the present study, the following species are newly recorded as hosts: the big-scaled redfin Tribolodon hakonensis (Cyprinidae), the flathead mullet Mugil cephalus (Mugilidae), the eastern keelback mullet Liza affinis (Mugilidae), the so-iny mullet Chelon haematocheilus (Mugilidae), the Japanese seaperch Lateolabrax japonicus (Lateolabracidae), the blackfin seabass L. latus (Lateolabracidae), the yellowfin seabream Acanthopagrus latus (Sparidae), the blackhead seabream A. schlegelii schlegelii (Sparidae), Rhyncopelates oxyrhynchus (Terapontidae), Ditrema viride (Embiotocidae), D. temminckii temminckii (Embiotocidae), Chaenogobius gulosus (Gobiidae), the yellowfin goby Acanthogobius flavimanus (Gobiidae), Pseudolabrus sp. (the red naped wrasse, either P. eoethinus or P. sieboldi) (Labridae) and the unicorn leatherjacket Aluterus monoceros (Monacanthidae). These host records indicate that N. japonica has a low host-specificity. This species attaches at the base of a fin or to the fin proper along the fin rays of its fish host (Fig. 7).

Female of Nerocila japonica (19.0 mm; TOYA-Cr 23266) attached at the base of the caudal fin of Liza affinis. Scale: in mm
Ecological notes
The collection of specimens of N. japonica from a brackish-water lake and the mouth of rivers suggests that this species can tolerate low salinity, as is the case with N. orbignyi and N. acuminata (see Brusca, 1981).
The coloration of the dorsal surface of N. japonica (dark blue with two faint pale submedian longitudinal bands) is almost the same as that of N. orbignyi and N. acuminata (see Brusca, 1981; Bruce, 1987). This species probably camouflages itself on the attachment sites of its host fish using its stable bilaterally symmetrical coloration. A similar suggestion has been made by Körner (1982) for N. bivittata (Risso, 1816).
An extensive skin erosion was often observed at the attachment site of N. japonica. Lesions observed in the present study resemble those caused by N. acuminata (Rand, 1986). N. japonica may cause anaemia and result in a secondary bacterial infection of the infected host. More study is needed on this species, because only limited information is available on its ecology and pathology.
A small bivalve (not identified) was found attached to the left coxa of pereonite 6 of a male N. japonica (19.0 mm) collected from Tribolodon hakonensis in the brackish Lake Nakaumi.
Comments on previous records of Nerocila acuminata in Japanese waters
Nerocila acuminata is known as an East Pacific and Caribbean species and was recorded from about 40 host fish species in the eastern Pacific (Brusca, 1981). On the other hand, this species has also been recorded from off Japan (Iwasa, 1947; Shiino, 1965; Honma et al., 1974; Nunomura, 1981, 1985). Saito et al. (2000) cited these records of N. acuminata in their checklist of isopods of Japan.
Iwasa (1947) recorded N. acuminata from Japanese waters in an encyclopaedia of the fauna of Japan. His description is brief, but it includes a diagnostic feature, i.e. the uropod endopod with a serrate medial margin and a truncate distal margin. This feature strongly suggests that he actually found an unrecorded Nerocila species close to N. serra Schioedte & Meinert, 1881, although it is impossible to determine which species without a re-examination of Iwasa’s material. Shiino (1965) also recorded Iwasa’s (1947) species as N. acuminata from Japan.
Honma et al. (1974) recorded N. acuminata in a faunal list of a brackish-water lake (Kamo-ko) on Sado Island located in the Sea of Japan, but gave no diagnostic information. Nunomura (1981) also recorded N. acuminata from Sado Island (two specimens on the body surface of Tribolodon hakonensis (as Leuciscus (Tribolodon) hakonensis) and two specimens without host data), but he did not give any diagnostic information. In his annotated list of the species in Nerocila, Bruce (1987, p. 400) suspected that N. acuminata reported by Nunomura (1981) might actually represent N. japonica. The specimens recorded by Nunomura (1981) were deposited at the Sado Marine Biological Station (SMBS), Niigata University (N. Nunomura, pers. comm.). At our request Mr T. Shimotani of the SMBS tried to find these specimens, and one of us (KN) also examined invertebrate specimens preserved at the SMBS, but no cymothoid material could be found. The specimens recorded by Honma et al. (1974) and Nunomura (1981) are presumed not to be extant; therefore, the identity of those specimens is uncertain. Based on current information on the geographical ranges of the two Nerocila species, it is likely that both Honma et al. (1974) and Nunomura (1981) actually misidentified specimens of N. japonica as N. acuminata.
Nunomura (1985) recorded two females of N. acuminata from Toyama Bay (the Sea of Japan). We re-examined these specimens deposited in the TOYA and concluded that these are N. japonica (see above under Material used in previous identifications).
There has so far been no reliable record of N. acuminata from Japanese waters. Therefore, records of N. acuminata from Japan should not be regarded as being N. acuminata. In addition, Iwasa’s (1965) record of N. acuminata from off Taiwan is questionable in view of current information on the geographical range of this species.
References
Brandt, A., & Poore, G. C. B. (2003). Higher classification of the flabelliferan and related Isopoda based on a reappraisal of relationships. Invertebrate Systematics, 17, 893–923.
Bruce, N. L. (1987). Australian species of Nerocila Leach, 1818, and Creniola n. gen. (Isopoda: Cymothoidae), crustacean parasites of marine fishes. Records of the Australian Museum, 39, 355–412.
Bruce, N. L., & Schotte, M. (2009). Nerocila Leach, 1818. In: Schotte, M., Boyko, C. B., Bruce, N. L., Poore, G. C. B., Taiti, S., & Wilson, G. D. F. (Eds) World marine, freshwater and terrestrial isopod crustaceans database. Accessed through: World Register of Marine Species at http://www.marinespecies.eu/aphia.php?p=taxdetails&id=118417 on 2010-01-20.
Brusca, R. C. (1981). A monograph on the Isopoda Cymothoidae (Crustacea) of the eastern Pacific. Zoological Journal of the Linnean Society, 73, 117–199.
Froese, R., & Pauly, D. (Eds) (2009). FishBase. World Wide Web electronic publication. www.fishbase.org, version (10/2009).
Gurjanova, E. F. (1936). Isopods of the Eastern Seas. Fauna of the USSR. Crustacea Vol. 7 (Part 3). Moscow-Leningrad: Academy of Sciences of the USSR, 278 pp. (in Russian).
Hashimoto, K. (2007). Record of Nerocila japonica (Isopoda, Cymothoidae), parasitic on Japanese snook from Kochi Prefecture, Japan. Nankiseibutu, 49, 183–185 (in Japanese).
Honma, Y., Kitami, T., & Ito, S. (1974). Fauna of Lake Kamo-ko, Sado Island in the Japan Sea (A preliminary report). Proceedings of the Japanese Society of Systematic Zoology, 10, 63–73. (in Japanese with English abstract).
Iwasa, M. (1947). Nerocila acuminata Schioedte et Meinert. In: Uchida, S. (Ed.) Revised and enlarged illustrated encyclopedia of the fauna of Japan. Tokyo: Hokuryukan, p. 815 (in Japanese).
Iwasa, M. (1965). On a small collection of amphipods and isopods from Korea, Formosa, and the Loo-Choo Islands. Researches on Crustacea, 2, 56–59 (in Japanese).
Körner, H. K. (1982). Countershading by physiological colour change in the fish louse Anilocra physodes L. (Crustacea: Isopoda). Oecologia, 55, 248–250.
Nierstrasz, H. F. (1918). Alte und neue Isopoden. Zoologische Mededeelingen, Rijksmuseum van Natuurlijke Historie te Leiden, 4, 103–142. plates 9, 10.
Nierstrasz, H. F. (1931). Die Isopoden der Siboga-Expedition. III. Isopoda Genuina II. Flabellifera. Siboga Expeditie Monographie, 32c, 123–232.
Nunomura, N. (1981). Isopod crustaceans from Sado Island in the Sea of Japan. Annual Report of the Sado Marine Biological Station, Niigata University, 11, 43–62.
Nunomura, N. (1985). Marine isopod crustaceans in the coast of Toyama Bay. Memoirs of the Natural Science Museum, Tokyo, 18, 121–139.
Rand, T. G. (1986). The histopathology of infestation of Paranthias furcifer (L.) (Osteichthyes: Serranidae) by Nerocila acuminata (Schioedte and Meinert) (Crustacea: Isopoda: Cymothoidae). Journal of Fish Diseases, 9, 143–146.
Saito, N., Itani, G., & Nunomura, N. (2000). A preliminary check list of isopod crustaceans in Japan. Bulletin of the Toyama Science Museum, 23, 11–107. (in Japanese with English summary).
Sato, Y. (2001). Lates japonicus collected from southern part of Tokushima Prefecture. Museum News (Tokushima Prefectural Museum), 44, 6 (in Japanese).
Schioedte, J. C., & Meinert, F. W. (1881). Symbolae ad Monographiam Cymothoarum Crustaceorum Isopodum Familiae 2. Anilocridae. Naturhistorisk Tidsskrift, (3), 13, 1–166.
Shiino, S. M. (1965). Nerocila acuminata Schioedte et Meinert. In: Okada, Y., Uchida, S., & Uchida, T. (Eds) New illustrated encyclopedia of the fauna of Japan. Tokyo: Hokuryukan, Part 2, p. 543. (in Japanese).
Thielemann, M. (1910). Beiträge zur Kenntnis der Isopodenfauna Ostasiens. Abhandlungen der Mathematisch-Physikalischen Classe der Königlich Bayerischen Akademie der Wissenschaften. München, 2(3), 1–109.
Yu, H., & Li, X. (2003). Study on the Cymothoidae from Chinese waters. Studia Marina Sinica, 45, 223–238 (in Chinese with English summary).
Acknowledgements
We wish to express our gratitude to Dr Y. Kano (Uozu Aquarium), Dr T. Kato (Kyoto University), Mr H. Konishi (Suita City), Dr H. Ariyama (Osaka Prefectural Fisheries Experimental Station), Mr A. Aimoto (Kankyo Kagaku Co., Ltd), Mr A. Nishimura (Okayama City), Mr K. Yamaguchi (Shinjiko Nature Museum), Dr T. Umino (Hiroshima University), Mr N. Nakadachi (Hiroshima University), Mr H. Yoshigou (Chugai Technos Co. Ltd), Mr T. Morihisa (Shimane University), Dr Y. Sato (Tokushima Prefectural Museum), Dr S. Arakaki (Kyushu University), Mr M. Wada (Kadogawa Town) and Mr M. Ito (Minamisatuma City) for providing specimens or valuable information. We are also much indebted to Dr Anders Warén (SMNH), Mr N. Nunomura (TOYA), Dr R. Yamanishi (OMNH), Mr T. Miki (HCA) and Dr M. Shimomura (KMNH) for lending specimens, and to Mr T. Shimotani (SMBS) for providing information on the specimens preserved at the SMBS. We also thank Dr I. Madinabeitia (Hiroshima University) for her comments on an early version of the manuscript. Part of the present study was supported by grants from the TaKaRa Harmonist Foundation, the Fujiwara Natural History Foundation and Grant-in-Aids for Scientific Research (B) (No. 18380116 to KN, No. 20380110 to TY) from the Japan Society for the Promotion of Science.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Yamauchi, T., Nagasawa, K. Redescription of the fish parasite Nerocila japonica Schioedte & Meinert, 1881 (Crustacea: Isopoda: Cymothoidae), with comments on previous records of N. acuminata in Japanese waters. Syst Parasitol 81, 147–157 (2012). https://doi.org/10.1007/s11230-011-9336-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11230-011-9336-5