
Abstract
Research with domestic dogs provides a unique approach for exploring the evolution of fairness and justice. Not only are dogs descended from highly social canids; they have also been bred for cooperative tasks with humans. Dogs act cooperatively in social play and are skilled on other social cognitive tasks. It is reasonable to ask whether dogs behave in ways similar to primates in other social contexts. In particular, do dogs perceive and respond to unfairness or injustice, a skill potentially borne of long-term affiliation with and selection by humans? Using a revised test of inequity aversion which looks at advantageous and disadvantageous inequity, the current research investigated the behavior of 38 domestic dogs. Subject dogs and a control dog approached two trainers in turn: one who rewarded them equally for sitting on command and one who rewarded them unequally—either over-rewarding or under-rewarding the control dog. After familiarization with the trainers, subjects chose which trainer to approach by themselves. Subjects preferred the over-rewarding trainer over the fair trainer; they had no preference between the under-rewarding and the fair trainer. Further analyses found that length of ownership, subjects’ age, and cooperative work experience reversed the approach preference, predicting preference for the fair trainer—though breed did not. These results suggest that the precursory sensitivity, which dogs showed to iniquitous outcomes in prior research, does not extend to both advantageous and disadvantageous inequity and does not hold when the subject is continually rewarded. Dogs selected a trainer who had treated them “unfairly,” yet who presented a potentially greater opportunity for future rewards. When the stakes were high, dogs showed a greater sensitivity to the quantity of a reward than to the fairness of a reward.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Nobody ever saw a dog make a fair and deliberate exchange of one bone for another with another dog. (Adam Smith, Wealth of Nations)
Do dogs have a sense of fairness? While the founder of modern economics may not have thought so, one suspects that Adam Smith might not have been a dog owner: over 80 % of owners believe their dogs experience jealousy (Morris, Doe, & Godsell, 2008), and jealousy requires, it is thought, some understanding of what is fair and unfair (Reddy, 2010; Salovey & Rodin, 1989). Owners reported that dogs expressed their jealousy when attention was unequally given to themselves and another person or dog (Morris et al., 2008). Anecdotally, owners note that their dogs know when another dog has gotten a treat and they have not (Brosnan, 2006). One study of dogs has tested “inequity aversion,” proposed as a gauge of fairness (Range, Horn, Viranyi, & Huber, 2008). Other than that study, there has been no empirical investigation of this attribution to dogs of a capacity to gauge the fairness of a situation and react thereto (Horowitz & Bekoff, 2007).
There has been a growth of interest in the past decades in whether non-human animals share abilities that used to be considered exclusively human, such as higher emotions and conceptual understanding. Recently, the evolution of morality, also based in an appreciation of fairness and justice, has begun to be explored (e.g., de Waal, 1996). Researchers have proposed that primates may have the rudiments of a moral understanding (Brosnan & de Waal, 2003) based on their natural behavior: chimpanzees can be cooperative with others and vengeful of those who do not cooperate (de Waal, 1996). Notably, some species of primates in the wild are marked by relatively equitable—“fair”—distribution of resources among members (Brosnan, 2006). In experimental studies asking subjects to trade tokens for food, both capuchin monkeys and chimpanzees refused to trade for a lesser reward when able to view a conspecific trading for a preferred reward (Brosnan & de Waal, 2003; Brosnan, Schiff, & de Waal, 2005). Tests of non-human primates’ performance on this kind of “inequity aversion”—the objection to unequal return for one’s actions—have begun to proliferate, as have questions about what experimental elements may change the interpretation of results (Bräuer, Call, & Tomasello, 2009; Wynne, 2004; see also Price & Brosnan, 2012; Bräuer & Hanus, 2012, this issue).
In exploring the evolution of fairness and justice, research with domestic dogs may provide particular insight. Domestic dogs, Canis familiaris, are thought to have been selected in part for cooperativeness with humans (Gácsi, McGreevy, Kara, & Miklósi, 2009). Working dogs engage in elaborate cooperative behavior in, for instance, herding livestock, hunting with humans, search-and-rescue tasks, and assisting disabled humans. And while the nature of the cooperative behavior is less specific, the great majority of dogs who live in human homes develop a bonded relationship with humans. The strong dog–human bond, as well as humans’ tendency to make psychological attributions to dogs, may be founded in part on the dogs’ reliance on and sensitivity to human behavior (Horowitz & Bekoff, 2007; Topál, Miklósi, & Csányi, 1997). Given the social ancestry of dogs, and the possibility that long-term human affiliation (and selection) has changed their attention and/or cognition, recent experiments have asked whether dogs respond in a way similar to primates on various social tasks. The results have found dogs to have socio-cognitive skills in communicative contexts (Miklósi, Polgárdi, Topál, & Csányi, 1998; Soproni, Miklósi, Topál, & Csányi, 2002) and to be attentive to the attention of other dogs (Horowitz, 2009) and humans (Call, Bräuer, Kaminski, & Tomasello, 2003; Gácsi, Miklósi, Varga, Topál, & Csányi, 2004; Schwab & Huber, 2006) at rates that often surpass non-human primates.
It has been suggested that in social play, dogs show cooperative behavior with conspecifics and do not tolerate those who flout the rules of play (Bekoff, 2004), although the evidence for this is anecdotal. Certainly rough-and-tumble play involves coordinated actions (Horowitz, 2009), and social play is cooperative insofar as turn-taking, role-reversal, and self-handicapping—larger or stronger dogs moderating the force they use with smaller or weaker dogs—are common (Bekoff & Allen, 1998; Horowitz, 2009). Cooperative play may even be necessary: coyote pups who did not engage in social play were less closely bonded to their groups and more likely to leave their group, an act which is often fatal (Bekoff, 2002). For these reasons, some have proposed that play, common to most socialized dogs, is the context in which fairness is learned (Bekoff, 2004).
One theory of the source of dogs’ abilities holds that through the process of domestication, dogs have evolved some human-like skills (Hare, Brown, Williamson, & Tomasello, 2002). Given that dogs are social animals, with a social group expanded to include Homo sapiens as well as conspecifics, it is reasonable to ask whether they respond in way similar to primates in other social contexts. In particular, do they show a rudimentary form of justice, borne of long-term affiliation with and selection by humans? Given their ability to cooperate with and coordinate behavior with humans and conspecifics, dogs are good candidates to have a sense of fairness.
One study has tested dogs’ abilities in this regard. Range et al. (2008; see also Range, Leitner, & Virányi, 2012, this issue) extended the “inequity aversion” paradigm to dogs. They asked dogs to do a simple task—“giving the paw”—for a reward in a social context (two dogs took turns interacting with the experimenter). In some conditions, the experimenter stopped rewarding one of the dogs, although she still requested the dog’s paw-giving behavior. The authors found that these dogs eventually forwent continued participation when no longer rewarded: they refused to give the paw. Further, they refused more quickly when in a social context where another dog was rewarded for performance of that same task, than when alone with the experimenter.
The authors reported their results as evidence for inequity aversion in dogs. The results were also widely reported—not by the authors, but by the media—to be evidence of the dogs’ understanding of the “fairness” or “unfairness” of the situation, and responding so as to “level” the field, by refusing to do the requested action without a reward (Morell, 2009). Indeed, the very interest in studying inequity aversion is to thereby model fairness (Fehr & Schmidt, 1999). However, when considering a non-linguistic population, it is appropriate to ask whether an inequity aversion test is equivalent to having a conceptual understanding of fairness. “Fairness” and “justice” are, for humans, inextricably linked with societal norms or explicit rules regulating the worth of a task or person; for animals who are not full members of human society, their appreciation of fairness may have less to do with such entailments and more to do with adaptive consequences of behavior. It is worth exploring the nature of the dog’s sensitivity to unequal rewards before inequity aversion studies are equated with tests of understanding of fairness.
The present research represents a complementary approach to Range et al. (2008) for testing a dog’s response to inequity. If dogs are truly sensitive to unequal rewards, one would expect that they should remain so in slightly different contexts. In Range et al., the nature of the experimental procedure leads to alternate interpretations of the dog’s behavior. The present research emends the procedure, as described below. Additionally, the prior research explored only the dog’s reaction to a situation unfair to themselves; the current research includes a situation unfair to another dog.
To the procedural point: the order of operations followed in the prior experimental method left room for alternative explanations for the dogs’ behavior. Dogs were always rewarded in their first trial to avoid the “complete frustration” of a subject put in a novel situation and not rewarded for obeying a command (Range et al., 2008). What this created, then, is a simple explanation for the dog’s subsequent refusals to work: extinction. When a behavior is no longer associated with subsequent rewards, the behavior may die out.
Importantly, the subject dogs’ refusal to do the task was accelerated in the presence of a social partner (Range et al., 2008). Thus, there appears to have been a separate or additional element at work in the socially facilitated extinction. Some element of the social setting—simply the appearance of another dog or the presence of a reward given to another dog—hastened the dogs’ uncooperative behavior. The authors experimentally tested and eliminated the first option, so what remains is that the subjects were somehow affected by seeing another dog receive a reward.
A new design can remove any form of extinction as an explanation for the dogs’ behavior. In this research, we created a situation in which the subjects always received a reward for their behavior, thus eliminating the possibility that their subsequent behavior is simply a form of extinction (however, socially provoked). In each trial, the subject dog received the same amount of reward, and it was the control dog’s reward amount which was varied.
Also procedurally, the selection of the “work” behavior for which the dogs get rewarded is important. It cannot be so difficult that all untrained dogs fail at it. “Raising one’s paw” was a good choice. But, as raising one’s paw in a social setting is considered a submissive behavior (Lorenz, 1954), performance of this action could be affected by the social setting alone, regardless of the change in the dog’s fortunes. In the current research, the action requested of the dog was changed from “giving the paw” to “sitting,” a behavior neither submissive nor aggressive, to eliminate the social salience of performing the action.
Additionally, the Range et al. (2008) protocol tested only one of two kinds of inequity aversion: when the subject received an “unfair” reward. Much inequity research examines inequity from both sides: both when the subject receives less of some reward or compensation than another individual—disadvantageous inequity aversion—and when he receives more than another individual—advantageous inequity aversion (Blake & McAuliffe, 2011; Brosnan, 2006; see also Yamamoto & Takimoto, 2012, next issue). Range et al. (2008) covered the disadvantageous situation, but not advantageous inequity aversion: would their subjects have been as averse to unequal rewards if the inequity were not to them, but to a conspecific in their company?
If dogs’ behavior is based on a sense of “fairness” that humans possess, they may also reject this situation. Humans tend to have a negative reaction to both kinds of inequity aversion (Brosnan, 2006). If overcompensated, they may reject the overpayment outright; if undercompensated, they may decrease or cease their effort (Blake & McAuliffe, 2011; Dawes, Fowler, Johnson, McElreath, & Smirnov, 2007; Loewenstein, Thompson, & Bazerman, 1989). Studies of advantageous inequity aversion are rare among non-human primate populations, and even in situations in which apparent advantageous inequity aversion occurs, it is to a far lesser degree than disadvantageous inequity aversion, and the cause of the behavior is ambiguous (Brosnan, Talbot, Ahlgren, Lambeth, & Schapiro, 2010). Instead, a popular gauge of sensitivity to others’ rewards is the “prosocial choice test,” which asks the subject to choose between rewarding just themselves or rewarding themselves and a partner, for no further effort (see also Yamamoto & Takimoto, 2012, next issue). Capuchin monkeys choose prosocial outcomes in a token-exchange task, if their partner is familiar to them (e.g., de Waal, Leimgruber, & Greenberg, 2008). Results of research with chimpanzees have been more ambiguous, with some evidence of helping in the absence of food rewards (e.g., Warneken & Tomasello, 2006), but no evidence of prosocial choice in food donation tasks (e.g., Jensen, Hare, Call, & Tomasello, 2006; Silk et al., 2005). However, recent work has shown that in some contexts chimpanzees might also behave prosocially, regardless of their relationship with the partner (Horner, Carter, Suchak, & de Waal, 2011).
Relatedly, within the study of social justice in human populations, one arm of organizational research investigates equitable or inequitable payment. Equity theory research suggests that people will accept some undesired payment if they believe that the payment is fair (Tyler, 2001). For example, Pritchard, Dunnette, & Jorgenson (1972) found that subjects who were being fairly paid for their work were more satisfied than others who were unfairly paid—even if these other subjects were overpaid. In their protocol, subjects were hired to a do a job for a fixed rate. The rate was invariant over conditions; however, in some trials, the subjects were told that they were paid fairly with respect to other workers. In other trials, they were told that they were overpaid or underpaid with respect to others doing the same work. Implicit in their result is the human ability to distinguish between fair and unfair outcomes, where unfairness runs in two directions: to themselves (underpayment for a task with respect to others), and to others (overpayment for a task with respect to others).
Given our interest in advancing the understanding of dogs’ inequity aversion to speak to their responses to “fair” or “unfair” situations, we used this research model in the present experimental design. Would a dog confronted not just with a situation in which he was “underpaid” but also with a situation in which he was “overpaid,” still prefer the “fair” situation in every case? In this design, a subject dog and a control dog approached one of two experimental “trainers” who asked the dogs to sit and rewarded them with a dog-food treat for so doing. One trainer was always “fair,” rewarding each dog equally (one treat apiece) and one was always “unfair,” either over-rewarding (three treats) or under-rewarding (zero treats) the control dog. Treat quantities used were determined by prior results. Research has shown that dogs can distinguish two quantities that differ by only one element (West & Young, 2002). Further, in an exchange task, dogs reliably distinguished one from three items (Leonardi, Vick, & Dufour, 2012).
The behavioral measure used in the present study was extrapolated from owner experience: owners report, anecdotally, that dogs are adept at associating some individuals with the treats they give to dogs encountered in dog-run and park settings. Dogs approach (and beg to) people who frequently provide food. Thus, in this study, the test trial asked subjects to make an affiliative choice: to approach one of two individuals to whom they have had recent exposure. Given the option of affiliating with a person who had treated them fairly or unfairly, with respect to another dog, would subjects evince a clear preference? If so, would this preference show a sensitivity to the person’s fair or unfair behavior?
It is worth noting that Range et al. and the current study rely on dogs’ abilities on tasks other than the one at hand: to wit, to form expectations about a person based on past rewarding behavior, and, in the current case, to attribute that behavior to the person—establishing “reputation.” Though non-human primates form expectations about reward, there is research indicating that monkeys do not reject an unreliable partner in an exchange interaction: capuchin monkeys, for instance, did not prefer a reliable partner (Brosnan & de Waal, 2009). Research assessing whether chimpanzees learned the reputation of experimenters found that they discriminated experimenters by behavior (giving/not giving; generous/selfish) whether they observed exchanges passively (Russell, Call, & Dunbar, 2008) or were themselves able to beg to and potentially be rewarded by one experimenter (Subiaul, Vonk, Okamoto-Barth, & Barth, 2008). Dogs also have been seen to learn reputation after observing “giving” and “withholding” third-party interactions and to distinguish intentional from inadvertent behavior (Kundey et al., 2010; Marshall-Pescini, Passalacqua, Ferrario, Valsecchi, & Prato-Previde, 2011). In these studies, dogs’ behavior was gauged by their selection of demonstrator, the same test trial method used in the current study.
If subject dogs in this study have a human-like sense of fairness or justice, they would be expected to choose the fair trainer both when the control dog is under-rewarded and when the control is over-rewarded. If the subjects perform as in Range et al. (2008), they would at least be expected to choose the fair trainer over the one who over-rewarded the control dog. If being continually provisioned with food eclipsed any interest in or attention to the different characteristics of the trainers, subjects’ approaches would be randomly distributed between trainers.
Method
Subjects
Thirty-eight domestic dogs (C. familiaris) participated in the experiment (Table 1). They were recruited through websites, publicly posted flyers, and word of mouth. All dogs were over 6 months of age (range .67–11.5 years) and had lived with their current owners for at least 6 months (range .5–11.0 years). 18 dogs were female and 19 dogs were male (one dog’s sex was unidentified by the owner). 50 % of the subject group (n = 19) were purebred dogs and 50 % (n = 19) were mixed-breed dogs. All subjects responded to basic commands and were able to sit upon request by the owner.
Procedure
The experiment was conducted in two locations on eight separate occasions in the fall of 2010 and 2011. One site was an indoor dog-shelter facility room used for training and puppy classes. The second site was outdoors in a public park in New York City frequented by dogs and owners. No overall difference in performance was seen in these two settings. Each site was sufficiently isolated to avoid interaction with dogs or people not involved in the experiment.
Before the experiment began, the procedure was explained to the subject dog’s owner, who completed a consent form and filled out a background questionnaire. As the owners had a role in the experiment, their role was described to them, but they were not informed of the nature of the experiment.
Each dog participated in two trials (Fig. 1). Each trial had two parts: a familiarization period and a test trial. In the familiarization period, subject dogs (S) were accompanied by a control dog (C) handled by an experimenter. One of two dogs affiliated with the experimenters, both adult mixed-breed dogs, served as the control dog. Dog owners served as subject dog handlers for all trials in order to reduce anxiety for the subjects.

Trial procedure: each subject dog participated in two trials. Each trial had two components: a familiarization period followed by a test trial. Trials differed by kind of familiarization: in the advantageous inequity case, dogs alternated approach to a fair and to an under-rewarding trainer. In the disadvantageous inequity case, dogs alternated approach to a fair and to an over-rewarding trainer. (For instance, for one subject, the order of events went as follows: approach over-rewarding trainer, fair trainer, over-rewarding trainer, fair trainer; test trial. Approach fair trainer, under-rewarding trainer, fair trainer, under-rewarding trainer; test trial.)
Familiarization Period
As each dog participated in two trials, each had two familiarization periods followed by a test trial. In both familiarization periods, S and C and their handlers entered the testing course together. Dogs were held on leash. The course consisted of a designated “start” position approximately 15 ft from and facing two trainers, a “fair” trainer and an “unfair” trainer, standing 8 ft apart from each other (Fig. 2). The trainer held a cardboard plate in each hand bearing dog treats (cubes of dehydrated chicken). The quantity of treats varied by trainer role (see below). Together, the C and S approached one trainer at a time; as they did so, the trainer knelt down, looking straight ahead. When the dogs reached a designated stop point, a foot in front of the trainer, the trainer said “Sit!” in a strong voice. Trainers repeated this request up to five times or until both dogs touched their bottoms to the ground. At that point, the trainer displayed the plates to the dogs, crossing her arms once to display each plate in front of each dog, and again uncrossed their arms. Trainers then released the dogs to eat the dog treats by saying “Okay!” Dogs were permitted to eat the treats, and dogs and handlers returned to the start position. Dogs and handlers then approached the second trainer.

Layout for experimental trials: subject and control dogs and their handlers began at the START position and alternated approach to two TRAINERS 15 ft ahead
Each familiarization period included four approaches, alternating between approaching the “fair” trainer and the “unfair” trainer. We used only two approaches to each trainer in each trial, as we were not only testing a dog’s native ability of sensitivity to inequity but also, implicitly, the dog’s ability to identify the parameters of a novel situation. In pilot testing and experimenter training trials, more than one approach was necessary and two were sufficient for dogs to learn the context of reinforcement and to associate the trainer giving the reward with the reward given: after four trials, dogs did not hesitate to make a choice. We also visually confirmed in these preliminary trials that dogs were able to see—and often tracked with their eyes—the reward being offered to the control dog. However, a minimum of familiarization approaches (4) was used to avoid multiple-trial presentations, which may produce data relating to something other than what is being tested (Hauser, Cominsa, Pytkaa, Cahilla, & Velez-Calderona, 2011), such as learning ability. In particular, Hauser et al. demonstrated that single-trial presentations (of various communicative actions) to subject dogs, as contrasted with the multiple-trial presentation used in many similar studies, could replicate previous studies’ results in some, but not all cases. They suggested that the latter failure indicated that dogs’ successful performance in previous studies may have been based on reinforcement learning over even a few trials.
The order of trials and of trainer approaches within each trial was randomized. Each trainer wore sunglasses in order to avoid making discriminatory eye contact with one dog over another. The trainers themselves were also randomly assigned to the fair or unfair roles in each trial and alternated wearing a gray, white, or black t-shirt. New trainers were used in each of the below two types of trials. These measures were taken to ensure that the subjects were not choosing a trainer based on her personal features.
Advantageous Inequity: Fair Trainer Versus Under-Rewarding Trainer
One trial paired an “advantageous” inequity situation with an equitable situation by having the trainers reward the dogs differently. The “fair” trainer rewarded each dog with one dog treat apiece. The “unfair” trainer under-rewarded the control dog: she gave C no treats and gave S one treat.
Disadvantageous Inequity: Fair Trainer Versus Over-Rewarding Trainer
The second trial paired a “disadvantageous” inequity situation with an equitable situation. In this comparison, the “fair” trainer again rewarded each dog with one dog treat apiece. The “unfair” trainer over-rewarded the control dog: she gave C three treats and gave S one treat.
In every comparison, S received one treat from each trainer. Only C’s reward changed with the trainer roles (Fig. 3).

Examples of the familiarization period. a Subject and control (black dog) approach a “fair” trainer in a trial run indoors. Owner and control-dog handler stand beside the dogs. b Subject and control (black dog) approach an “over-rewarding” trainer in a trial run outdoors. Owner and control-dog handler stand beside the dogs. Another trainer can be seen in the background
Test Trial
In the test trial, a single-approach test was used in order to maintain motivation after repeated approaches and rewards for sitting behavior. C was removed from the course. The trainers each held one empty cardboard plate in front of them (in order to control for a choice based on olfaction, in the test trial neither trainer’s plate held any food). S and handler approached the trainers along a center path, equidistant from the two trainers. After 5 ft, the handler dropped the leash and remained behind S, with the option to hold a loose leash if that felt more natural to the handler. S’s behavior was scored for trainer approached or, if the dog did not approach either trainer within 15 s, was scored “no choice.” “Approach” was scored only if the dog unambiguously walked toward one of the two trainers to within one foot.
Behavior Coding
Both the familiarization periods and the test trials were videotaped and reviewed both to confirm S’s behavior and to determine if there were any handler errors. The coding of handler behavior was important, as the owners were not blinded to the trainers’ behavior in the familiarization period. As Hauser et al. (2011) noted, various handler errors of inadvertent information-providing can affect the subject dogs’ performance. A handler’s verbal cueing, effective pointing, leash guidance (holding or dropping the leash so as to restrict free choice), or walking ahead of the dog would be sufficient to disqualify the subject on a given trial.
The author coded all trials. Four trials were eliminated because of handler error: three in the disadvantageous inequity comparison; one in the advantageous inequity comparison. An independent coder, naive to the role of the trainers in each test trial, separately coded all test trials except those excluded for handler error. The two coders agreed on 71 of 72 trials, Cohen’s kappa = .9753.
Results
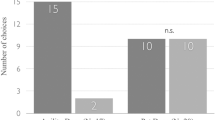
Seventy-two trials were coded and scored. In 65 trials, the subject dog made a choice of trainer; on seven trials, the subject made no choice. In the disadvantageous inequity trial, in which subjects received less reward than the control, more dogs (25; 78.1 %) chose the over-rewarding trainer than the fair trainer (7; 21.9 %), χ2(1) = 10.13, p = .001 (Fig. 4). In the advantageous inequity trial, in which subjects received more reward than the control, dogs were equally likely to choose the fair trainer (16; 48.5 %) and the under-rewarding trainer (17; 51.5 %), χ2(1) = .03, p = .86.

Main results: subjects selected the over-rewarding trainer significantly more often than the fair trainer (p < .001); subjects did not show a preference between the under-rewarding and the fair trainer (p = .86)
Additional analyses were performed using generalized estimating equations to account for the repeated measures nature of the data (i.e., each dog made two choices between a fair and an unfair trainer).
Trial order had no effect on choice of trainer, χ2(1) = 1.12, p = .29, nor did order interact with trial type, χ2(1) = 1.04, p = .31. Additionally, there was no main effect of the side that the fair trainer was standing on (right vs. left) on choice, χ2(1) = 2.44, p = .12.
There was no main effect of dog gender on choice, χ2(1) = .69, p = .41, nor did gender interact with trial type, χ2(1) = .09, p = .77. Neither was there a main effect of purebred versus mixed-breed on choice, χ2(1) = 2.38, p = .12, nor did this variable interact with trial type, χ2(1) = .24, p = .62.
Length of ownership (included as a continuous variable) was a significant predictor of the likelihood of choosing the fair trainer, χ2(1) = 3.74, p = .05. There was also a trend for the interaction between the length of ownership and trial type, χ2(1) = 2.85, p = .09, indicating that the relationship between length and probability of choosing the fair trainer was stronger (and was only significant) for the choice between the over-rewarding and fair trainers, b = .49 (.24), χ2(1) = 4.31, p = .04, than for the choice between the under-rewarding and fair trainers, b = .06 (.13), χ2(1) = .22, p = .64.
Dog age was highly correlated with length of ownership (r = .91). Accordingly, age showed a similar main effect on the likelihood of choosing the fair trainer, b = .24 (.12), χ2(1) = 4.99, p = .046. Unlike with length of ownership, this main effect was not qualified by an interaction with trial type, χ2(1) = 1.97, p = .16.
Eighteen percent of the participant dogs (n = 7) had experience at a cooperative task (i.e., working as a therapy dog or doing agility). The dogs with cooperative work experience were more likely than those with no such experience to choose the fair trainer over the unfair trainer,Footnote 1 χ2(1) = 4.11, p = .04, and this difference was not moderated by the type of comparison, χ2(1) = .92, p = .34.
To investigate possible breed contributions to the preference for fairness, we classified the purebred dogs of relevant breeding as either independent workers or cooperative workers, using the scheme proposed by Gácsi et al. (2009). This yielded five independent workers (Brussels Griffon, Jack Russell, Manchester terrier, Petit Basset Griffon Vendéen, Wheaten Terrier) and seven cooperative workers [Border Collie (2), Golden Retriever, Labrador Retriever, Poodle (2), Shetland Sheepdog]. Using breed classification as a predictor, the cooperative worker breeds were more likely to choose the fair trainer across trials, b = .86 (1.13), although the effect was not significant, χ2(1) = .57, p = .45.
Discussion
Quantity may Eclipse “Fairness”
In this study, subject dogs were not averse to individuals who had treated them unfairly. Indeed, dogs most often preferred to approach a trainer who had repeatedly treated the subjects unfairly with respect to another dog, rather than one who treated the two dogs equivalently to one another. The disadvantageous inequity aversion seen in Range et al. (2008) was not seen here—although their preference for the trainer who gave their partner more implies that they were aware of the disadvantageous inequity and that this subsequently changed their behavior. Neither did dogs show advantageous inequity aversion: they neither preferred nor rejected the unfair trainer who rewarded them and failed to reward the control dog. These subjects’ behavior in an inequity situation contrasts with human behavior and, correspondingly, it is appropriate to suggest that dogs’ sense of “fairness” is not the same as humans’ in a clearly delineated payment scenario.
What explains this result? While dogs selected a trainer who had treated them “unfairly” in the disadvantageous inequity aversion trial, this trainer had another salient characteristic: she distributed the most dog treats. Over the two familiarization approaches, she distributed eight treats to the two dogs; the fair trainer gave out half as many. Additionally, she distributed the largest quantity to any one dog (three treats as compared to either one or none). Possibly, it was this feature which led dogs to affiliate with the unfair trainer, as her distribution rate may have suggested a greater opportunity for future rewards. The present result suggests that, when the rewards are great, dogs have a sensitivity to quantity which over-rides any interest in the fairness of the distributer or distribution.
This conclusion must also take into account the dogs’ apparent indifference to the quantity distinction in the advantageous inequity (under-rewarding) trials. If quantity was the only behavioral motivator, then dogs should also have chosen to approach the fair trainer over the under-rewarding trainer. They did not. It is possible that the three treats distributed each approach by the over-rewarding trainer simply overshadowed all other salient characteristics of the experimental situation for the subjects.
Without positing anything further about the dogs’ perception of the experimental task, it does seem clear that subjects changed their behavior in reaction to a disadvantageous arrangement. While they did not respond to it with a call for fairness, it changed the nature of their approach preference. In some ways, this is similar to the dog and primate literature which showed inequity aversion: when a subject’s partner is treated better, the subject’s behavior is changed—although also in line with this literature, there seems to be little concern about being over-rewarded (see Bräuer & Hanus, 2012; Price & Brosnan, 2012; Range et al., 2012, this issue and Yamamoto & Takimoto, 2012, next issue). In the current study, it was simply changed to reflect, perhaps, the dog’s interest in getting what his partner had.
When considering the contributions of specific subject subgroups, the picture of dog ability and behavior changes somewhat. In particular, older dogs and those owned longer (often the same dogs) preferred to approach fair trainers in the disadvantageous inequity trial. Perhaps living in a social familial group contributes to perception of the distinction between fair and unfair situations. Social living involves distribution of resources, and over time some “fair” apportionment might be learned. Living in a human family, dogs might learn that a “fair” distribution of resources is exactly that which is provided by their human owners. Research with chimpanzees has found inequity aversion to be contingent on social group membership (Brosnan et al., 2005). Notably, in the current study, the (small) sub-group of subjects who lived with other dogs did not perform differently from the main group: 88 % [n = 7 (/8)] chose the over-rewarding trainer over the fair trainer; 44 % [n = 4 (/9)] chose the under-rewarding trainer over the fair trainer. The age and social relationship of the dogs in the Range et al. study may have been relevant to their performance, however. Much research on dogs’ socio-cognitive abilities highlights the relevance of prior experience on performance (see, e.g., Udell & Wynne, 2010).
Similarly, dogs trained for cooperative tasks more often chose the equitable trainer in both trials—i.e., regardless of who was being treated unequally by the unfair trainers. The same did not hold for the so-called “cooperative” breeds, though. These results may be reconciled (and are delimited by) the small samples from which they drew (n = 7 in both cases; one subject appeared in both groups). Still, they argue for more investigation of what is behind the application of the word “cooperation” when applied to a behavior or a group of dogs.
The Nature of Cooperation
To that point: what is called “cooperative” between animals should be acknowledged to involve heterogeneous abilities in differing contexts. When a human adult cooperates with a colleague to solve a problem, we can attribute to him an understanding that the other individual has his own perspective, abilities, and desideratum that must be taken into account to work together successfully. This kind of attribution would be hasty in description of toddlers working together to push a door open. For a domesticated animal, simply to live peaceably among another species is in some sense cooperative: this point hints at, but does not in and of itself demonstrate, though, that dogs understand the nature of cooperation. Similarly with “fairness”: in this study, the dogs’ sensitivity to an element of an iniquitous situation—the preference for who had “more” in one comparison—surely indicates that they noticed inequity in some way. They did not seem to make a judgment that bore on whether it was “fair” to themselves or to the other dog, however. To use these words—“cooperative”; “fairness”—carefully should not be taken as an undermining of an animal’s skills. Instead, it is a note of caution when assuming that behavior which is indicative of a mental ability in human adults is indicative of an identical ability in animals with different sensory systems, brain organization, and phylogenetic histories.
Experimental Design with the Subject in Mind
Differing results such as those of the current research and Range et al. (2008; see also Range et al., 2012, this issue) exemplify how a non-human subject may not see, attend to, or comprehend the nature of the task asked of the subject in an experimental test. For instance, though we assigned the trainers the names “fair” and “unfair,” in one scenario the quantity of food provided by trainers appears to have been more salient than their fair or unfair behavior, even in a social setting. The subject dogs may have been labeling them “less” and “more” trainers.
Similarly, the behavioral measure can lead to the testing of an ability other than the desired one. In the current study, the measurement was dogs’ performance on a single trial. In Range et al. (2008), the dogs’ performance over many trials was scored. The authors report that dogs who were no longer rewarded, whether in a social context or not, did not stop cooperating at once; instead, they gradually stopped, giving their paws from 5 to 30 more times before ceasing to do so (Range et al., 2008, Fig. 2). Scoring the dog’s behavior this way is relevant to the conclusions that can be drawn from the results. The dog’s behavior is now affected not only by a single response of the trainer (not rewarding him), but also by the (recent) past experience of what his own behavior can yield. That is, the dog is learning that “raising my paw no longer produces a reward,” and the presence of a dog whose paw does produce rewards highlights the inadequacy of one’s own act. In the present study, there is no such learning across test trials: each dog had only one test trial.
It should be noted that had dogs rejected the over-rewarding trainer to affiliate with the fair trainer, the method of the current study could, at most, have shown a weak form of inequity aversion. Some human research on the topic asks subjects to demonstrate their aversion to an iniquitous situation by sacrificing some of their rewards (Blake & McAuliffe, 2011). In this study, dogs were not asked to make a choice that reflected consideration of the fate of their social companion, the control dog, at all, and the control dog was not affected by the subject’s choices. Adam Smith’s assertion of inequitable dog resource trading remains unexamined.
“Non-data” Data
One way to remedy the effects on the data of the possible divide between the subject’s and experimenter’s perception of the task is to begin to videotape, score, and code what are ordinarily considered non-data. Following Hauser et al. (2011), handler behavior was scored. Given the salience of human presence, gestures, and eye contact to dogs, the handler’s behavior, even at a distance, could easily affect the dog’s behavior. Those trials removed for handler error included examples of inadvertent pointing and of dropping the leash in a way that might induce the dog to walk toward one trainer over the other. While these data, when included, do not change our main result (and we cannot be sure that they changed the dogs’ behavior), they represented unintentional elements of the trial that may be meaningful for the subjects.
Similarly, it is worth considering coding the behavior of the control dog. In Range et al. (2008), videotape of one reward-inequity trial shows the control dog beginning to offer the paw when it was asked of the subject (who was refusing to cooperate for no reward). Did all controls act this way? Could this behavior have either accentuated or diminished the subject dogs’ responses? In the current study, it was noted that after many trials, the control dog stopped reaching for the reward when in front of the under-rewarding trainer (who gave the control dog no reward), even though this role was played by a different experimenter on every trial. This demonstrates that the control dog certainly perceived the reward element of the situation. It is an open question whether his behavior could have had any effect on the subject dog’s perception and behavior. Coding control behavior would also be a way to begin to draw the picture of subject experience and perception in experimental trials.
These results should serve as a reminder that when considering the concept of fairness or justice across species, the data can also reveal that the subjects’ perception of the situation may be different than our own. In experimental design, we assume that the features of the social and reward situation are identical for the designers and our subjects. This is likely not the case.
Notes
Three owners did not complete this section of the questionnaire and so for these analyses n = 34 (one non-completer brought two subject dogs).
References
Bekoff, M. (2002). Virtuous nature. New Scientist, 175, 34.
Bekoff, M. (2004). Wild justice and fair play: Cooperation, forgiveness, and morality in animals. Biology and Philosophy, 19, 489–520.
Bekoff, M., & Allen, C. (1998). Intentional communication and social play: How and why animals negotiate and agree to play. In M. Bekoff & J. A. Byers (Eds.), Animal play: Evolutionary, comparative, and ecological perspectives (pp. 97–114). Cambridge, New York: Cambridge University Press.
Blake, P. R., & McAuliffe, K. (2011). “I had so much it didn’t seem fair”: Eight-year-olds reject two forms of inequity. Cognition, 120, 215–224.
Bräuer, J., Call, J., & Tomasello, M. (2009). Are apes inequity averse? New data on the token-exchange paradigm. American Journal of Primatology, 71, 175–181.
Bräuer, J., & Hanus, D. (2012). Social Justice Research, 25(3) (forthcoming).
Brosnan, S. F. (2006). Nonhuman species’ reactions to inequity and their implications for fairness. Social Justice Research, 19, 153–185.
Brosnan, S. F., & de Waal, F. B. M. (2003). Monkeys reject unequal pay. Nature, 425, 297–299.
Brosnan, S. F., & de Waal, F. B. M. (2009). Cebus apella tolerate intermittent unreliability in human experimenters. International Journal of Primatology, 30, 663–674.
Brosnan, S. F., Schiff, H. C., & de Waal, F. B. M. (2005). Tolerance for inequity may increase with social closeness in chimpanzees. Proceedings of the Royal Society of London B, 272, 253–258.
Brosnan, S. F., Talbot, C., Ahlgren, M., Lambeth, S. P., & Schapiro, S. J. (2010). Mechanisms underlying responses to inequitable outcomes in chimpanzees, Pan troglodytes. Animal Behaviour, 79, 1229–1237.
Call, J., Bräuer, J., Kaminski, J., & Tomasello, M. (2003). Domestic dogs (Canis familiaris) are sensitive to the attentional state of humans. Journal of Comparative Psychology, 117, 257–263.
Dawes, C. T., Fowler, J. H., Johnson, T., McElreath, R., & Smirnov, O. (2007). Egalitarian motives in humans. Nature, 446, 794–796.
de Waal, F. B. M. (1996). Good natured: The origins of right and wrong in human and other animals. Cambridge, MA: Harvard University Press.
de Waal, F. B. M., Leimgruber, K., & Greenberg, A. R. (2008). Giving is self-rewarding for monkeys. Proceedings of the National Academy of Sciences of the United States of America, 105, 13685–13689.
Fehr, E., & Schmidt, K. M. (1999). A theory of fairness, competition, and cooperation. Quarterly Journal of Economics, 114, 817–868.
Gácsi, M., McGreevy, P., Kara, E., & Miklósi, Á. (2009). Effects of selection for cooperation and attention in dogs. Behavioural and Brain Functions, 5, 31.
Gácsi, M., Miklósi, A., Varga, O., Topál, J., & Csányi, V. (2004). Are readers of our face readers of our minds? Dogs (Canis familiaris) show situation-dependent recognition of human’s attention. Animal Cognition, 7, 144–153.
Hare, B., Brown, M., Williamson, C., & Tomasello, M. (2002). The domestication of social cognition in dogs. Science, 298, 1634–1636.
Hauser, M. D., Cominsa, J. A., Pytkaa, L. M., Cahilla, D. P., & Velez-Calderona, S. (2011). What experimental experience affects dogs’ comprehension of human communicative actions? Behavioural Processes, 86, 7–20.
Horner, V., Carter, J. D., Suchak, M., & de Waal, F. (2011). Spontaneous prosocial choice by chimpanzees. Proceedings of the National Academy of Sciences of the United States of America, 108, 13847–13851.
Horowitz, A. (2009). Attention to attention in domestic dog (Canis familiaris) dyadic play. Animal Cognition, 12, 107–118.
Horowitz, A. C., & Bekoff, M. (2007). Naturalizing anthropomorphism: Behavioral prompts to our humanizing of animals. Anthrozoös, 20, 23–35.
Jensen, K., Hare, B., Call, J., & Tomasello, M. (2006). What’s in it for me? Self-regard precludes altruism and spite in chimpanzees. Proceedings of the Royal Society of London. Series B: Biological Sciences, 273, 1013–1021.
Kundey, S. M. A., De Los Reyes, A., Royer, E., Molina, S., Monnier, B., German, R., et al. (2010). Reputation-like inference in domestic dogs (Canis familiaris). Animal Cognition, 14, 291–302.
Leonardi, R. J., Vick, S.-J., & Dufour, V. (2012). Waiting for more: The performance of domestic dogs (Canis familiaris) on exchange tasks. Animal Cognition, 15, 107–120.
Loewenstein, G. F., Thompson, L., & Bazerman, M. H. (1989). Social utility and decision making in interpersonal contexts. Journal of Personality and Social Psychology, 57, 426–441.
Lorenz, K. (1954). Man meets dog. London: Methuen.
Marshall-Pescini, S., Passalacqua, C., Ferrario, A., Valsecchi, P., & Prato-Previde, E. (2011). Social eavesdropping in the domestic dog. Animal Behaviour, 81, 1177–1183.
Miklósi, Á., Polgárdi, R., Topál, J., & Csányi, V. (1998). Use of experimenter-given cues in dogs. Animal Cognition, 1, 113–121.
Morell, V. (2009). Going to the dogs. Science, 325, 1062–1065.
Morris, P. H., Doe, C., & Godsell, E. (2008). Secondary emotions in non-primate species? Behavioural reports and subjective claims by animal owners. Cognition and Emotion, 22, 3–20.
Price, S. A., & Brosnan, S. F. (2012). To each according to his need? Variability in the responses to inequity in non-human primates. Social Justice Research, 25(2). doi:10.1007/s11211-012-0153-z.
Pritchard, D., Dunnette, M. D., & Jorgenson, D. O. (1972). Effects of perceptions of equity and inequity on worker performance and satisfaction. Journal of Applied Psychology, 56, 75–94.
Range, F., Horn, L., Viranyi, Z., & Huber, L. (2008). The absence of reward induces inequity aversion in dogs. Proceedings of the National Academy of Sciences of the United States of America, 106, 340–345.
Range, F., Leitner, K., & Virányi, Z. (2012). The influence of the relationship and motivation on inequity aversion in dogs. Social Justice Research, 25(2). doi:10.1007/s11211-012-0155-x.
Reddy, V. (2010). Green eyes in bio-cultural frames. In S. Hart & M. Legerstee (Eds.), Handbook of jealousy: Theory, research and multidisciplinary approaches. Chichester: Wiley-Blackwell.
Russell, Y. I., Call, J., & Dunbar, R. I. M. (2008). Image scoring in great apes. Behavioural Processes, 78, 108–111.
Salovey, P., & Rodin, J. (1989). Envy and jealousy in close relationships. Review of Personality and Social Psychology, 10, 221–246.
Schwab, C., & Huber, L. (2006). Obey or not obey? Dogs (Canis familiaris) behave differently in response to attentional states of their owners. Journal of Comparative Psychology, 120, 169–175.
Silk, J., Brosnan, S. F., Vonk, J., Henrich, J., Povinelli, D. J., Shapiro, S., Richardson, A., Lambeth, S. P., & Mascaro, J. (2005). Chimpanzees are indifferent to the welfare of unrelated group members. Nature, 437, 1357–1359.
Soproni, K., Miklósi, Á., Topál, J., & Csányi, V. (2002). Dogs’ (Canis familiaris) responsiveness to human pointing gestures. Journal of Comparative Psychology, 116, 27–34.
Subiaul, F., Vonk, J., Okamoto-Barth, S., & Barth, J. (2008). Do chimpanzees learn reputation by observation? Evidence from direct and indirect experience with giving and withholding strangers. Animal Cognition, 11, 611–623.
Topál, J., Miklósi, Á., & Csányi, V. (1997). Dog-human relationship affects problem solving behaviour in the dog. Anthrozoös, 10, 214–224.
Tyler, T. R. (2001). Procedural strategies for gaining deference: Increasing social harmony or creating false consciousness? In J. M. Darley, D. M. Messick, & T. R. Tyler (Eds.), Social influences on ethical behavior in organizations (pp. 69–87). Mahwah, NJ: Lawrence Erlbaum.
Udell, M. A. R., & Wynne, C. D. L. (2010). Ontogeny and phylogeny: Both are essential to human-sensitive behavior in the genus Canis. Animal Behaviour, 79, e9–e14.
Warneken, F., & Tomasello, M. (2006). Altruistic helping in human infants and young chimpanzees. Science, 311, 1301–1303.
West, R. E., & Young, R. J. (2002). Do domestic dogs show any evidence of being able to count? Animal Cognition, 5, 183–186.
Wynne, C. D. L. (2004). Fair refusal by capuchin monkeys. Nature, 428, 140.
Yamamoto, S., & Takimoto, A. (2012). Social Justice Research, 25(3) (forthcoming).
Acknowledgments
This study was approved by Columbia University’s Institutional Animal Care and Use Committee (protocol AC-AAAC2044). Thanks are due to those who were integral to the running of the trials: Julie Hecht, Emily Cherenack, Adam Chapman, Orellana del Fierro, Rebekka Dohme, Rebecca Johnson, Meredith Leeman, Jennifer Oh, Shoshana Schoenfeld, Hannah Solomon, and Ilana Yablonovich. Drs. Heather Barry Kappes and Tom Tyler, previously at New York University, initiated this research and provided the conceptual foundation; Heather additionally provided great statistical support. Many thanks to Animal Haven, a non-profit shelter in New York City that generously donated use of their facility for running trials. The reviewers of this manuscript improved it, for which I give my thanks.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Horowitz, A. Fair is Fine, but More is Better: Limits to Inequity Aversion in the Domestic Dog. Soc Just Res 25, 195–212 (2012). https://doi.org/10.1007/s11211-012-0158-7
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11211-012-0158-7