
Abstract
Aims
Although nitrogen (N) fixation by moss-associated bacteria is the main source of new N in N-limited ecosystems like arctic tundra, we do not know which nutrient, molybdenum (Mo) or phosphorus (P), is rate-limiting for sustaining this process in the long-term. Further, how moss-associated N2 fixation impacts the belowground microbial regulation of decomposition remains unresolved.
Methods
Moss-associated N2 fixation and soil microbial process rates, abundance and community structure were assessed in long-term P and Mo field additions in the Subarctic during three years.
Results
We found tendencies for stimulation of moss-associated N2 fixation by Mo in the short term, by P in the long-term, and tendencies for a stimulation of soil microbial activity by P. However, large variation in microbial activity within and below the moss exceeded any systematic variation induced by the field treatments. Our findings suggest that soil microbial activity is not limited by N at our site, and that Mo and P only occasionally limit N2 fixation during a growing season.
Conclusions
Since increasing CO2 concentrations can induce nutrient limitation, the here reported transient limitation can easily shift into a chronic one with significant implications for ecosystem productivity and biogeochemistry.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Nitrogen (N) fixation performed by moss-associated, N2 fixing bacteria - predominantly cyanobacteria (Leppänen et al. 2013) - is the main pathway through which new N enters pristine ecosystems like boreal forests (DeLuca et al. 2002) and arctic tundra (Rousk and Michelsen 2017). Here, N input from atmospheric N deposition is low (0–2 kg N ha−1 yr−1; Peñuelas et al. 2013), and N commonly limits primary productivity (Mack et al. 2004; LeBauer and Treseder 2008; Wang et al. 2010). The environmental controllers of moss-associated N2 fixation are manifold and interdependent. For instance, both higher temperature and moisture promote N2 fixation in mosses (Rousk et al. 2017a, 2018a), while elevated N input can inhibit this key ecosystem process (Gundale et al. 2011; Ackermann et al. 2012; Rousk et al. 2013).
Another rate limiting factor for N2 fixation may be nutrient availability. Phosphorus (P) and molybdenum (Mo) are key for sustaining N2 fixation due to their importance for activity and structure of the nitrogenase enzyme. A recent meta-analysis of nutrient limitation of terrestrial, free-living N2 fixing bacteria (diazotrophs) found evidence of Mo-limitation of N2 fixation in soil, litter, lichens and mosses in tropical, temperate and boreal forests, and in temperate grasslands, and frequent limitation by P in tropical forests (Dynarski and Houlton 2018). Phosphorus-, but not Mo, limitation of N2 fixation in litter has been repeatedly found in tropical forests (Wurzburger et al. 2012; Reed et al. 2013). Yet, heterotrophic N2 fixation in tropical (Barron et al. 2009) and temperate (Perakis et al. 2017) forest soils has been shown to be limited by Mo. In boreal forests, however, Mo and/ or P limitation of N2 fixation in litter can depend on site and season (Jean et al. 2013).
Surprisingly, there is to date a scarcity of studies that have investigated how moss-associated N2 fixation is regulated by limiting nutrients, and very few studies have been conducted in arctic systems. In the Subarctic, Mo limited moss-associated N2 fixation in the short-term (1 day after nutrient supplements), while P promoted N2 fixation both in the medium-term (several weeks after addition) as well as towards the end of the growing season in the same study (Rousk et al. 2017b). Taken together, these earlier findings suggest that P is more important than Mo for N2 fixation performed by free-living diazotrophs in litter, while Mo seems to promote N2 fixation activity in soil and moss. However, the type of response appears to be dependent on the time scale of the assessment. The previously reported positive and immediate (< 1 day) effects of Mo additions on moss-associated N2 fixation suggests that physiological underpinnings of this functional response are linked to a relieved Mo limitation of enzyme synthesis, which can be fast and can fuel N2 fixation in the short-term (hours) until activity is limited by another factor. Whereas the observed medium and longer-term stimulation by P additions could reflect a P limitation of growth and photosynthesis of both moss and colonizing diazotrophs, enhancing and sustaining biomass and N2 fixation over several weeks (Rousk et al. 2017b). However, it remains unresolved how long these responses in N2 fixation to nutrient supplements can persist, and thus, how they will translate into changes in the overall ecosystem N budget. How long are the positive effects of P maintained? Are the observed patterns consistent across several years?
A consequence of enhanced N2 fixation activity via promotion by Mo and P supplements (Rousk et al. 2017b) is increased N input to the ecosystem. Transfer of fixed N2 from moss to other ecosystem components, including the soil below the moss carpet has been shown in the Arctic (Rousk et al. 2017c). An increase of available N in soil as a result of sustained, increased N2 fixation activity may also affect microbial activity belowground thus regulating the formation and turnover of soil carbon (C) (Liang et al. 2017). Even though soil microbes are commonly limited by C (e.g. Rosinger et al. 2019), they can be limited by N in pristine ecosystems (Sistla et al. 2012), including in those sites close to our study system (Jonasson et al. 1996, 1999; Rinnan et al. 2013). Thus, increased N input via N2 fixation as a result of nutrient supplements may lead to a shift in the limiting nutrient away from N for soil microbial growth and thus for organic matter decomposition, which could transfer C from soil to the atmosphere.
In this study, we addressed the questions of (1) how moss-associated N2 fixation responds to long-term (> 1 year) additions of P and Mo, and (2) if a shift in nutrient limitation of soil microbial activity is induced due to increased N input via a stimulated N2 fixation pathway (see e.g. Rousk et al. 2017c). To accomplish this, we revisited an experiment that started in 2015, with annual field additions of Mo and P in the Subarctic (Rousk et al. 2017b) until 2018. Both long-term (approx. 1 year) and short-term (1 day) responses to the nutrient supplements were resolved. We hypothesized that (H1) P additions would have long-lasting, positive effects on N2 fixation in mosses, that (H2) Mo would promote N2 fixation only in the short-term, and (H3) enhanced N2 fixation and subsequent N input into soil should shift an initially N-limited microbial decomposer community towards P-limitation.
Material and methods
Field site, experimental set-up and sampling
The field plots were established close to the Abisko Scientific Research Station, Northern Sweden, 560 m above sea level, 68°19 N, 18°50 E in June 2015. The mean annual air temperature in Abisko is 0.2 °C with the warmest and coldest months July and February, with average monthly temperatures 11.9 °C and − 10.0 °C, respectively (30-year mean 1986–2015, ANS 2016). Mean annual precipitation is 337 mm (30-year mean 1986–2015, ANS 2016). The site is a subarctic dry heath and the vegetation is dominated by mosses and a few scattered dwarf shrubs such as Vaccinium uliginosum (L.), Andromeda polifolia (L.) and Rhododendron lapponicum (L.) Wahlenb. The bedrock is a base-rich schist, and the organic soil horizon is up to 15 cm deep with a mean (±SE) soil pH of 6.7 ± 0.02 (Table 1). The plots were 1 × 1 m in size, and the dominant moss is the feather moss Hylocomium splendens (Hedw.) Schimp (Online Resource 1). The annual supplements of molybdate and phosphate were added in three sequential portions distributed evenly over the growing season in 2015 (see Rousk et al. 2017b). In subsequent years, 2016, 2017 and 2018, the annual supplements were added as a single dose early in the growing season (June–July). Molybdenum was added as Na2MoO4.2H2O and P was added as NaH2PO4.H2O. The field additions consisted of nine treatments with five replicates each, and 1 l of each solution was evenly applied onto each plot. The supplements were based on Mo and P content of the moss, which was measured before the initiation of the experiment, 2015. Moss content was 0.07 mg Mo m−2 and 0.4 g P m−2 assessed on dried moss (0.5 g) digested in 40 ml HNO3 on an atomic absorption spectrometer (AAS, PinAAcle 900 T, Perkin Emer, Shelton, CT, USA). Hence, the field treatments were: control (water), 10x moss Mo content (0.7 mg Mo m−2), 100x moss Mo content (7 mg Mo m−2), 10x moss P content (4 g P m−2), 100x moss P content (40 g P m−2), and the full factorial combinations of those: 10xMo + 10xP, 10xMo + 100xP, 100xMo + 10xP, 100xMo + 100xP.
Moss samples were collected before the yearly addition (i.e., 1 year after the last addition) as well as 1 day after the additions. Soil samples were also collected in 2017, just before the next nutrient addition, and hence, one year after the last full addition for measurement of N2 fixation, soil microbial variables (see below) and soil characteristics (Table 1). For this, several composite soil samples (~ 10) were sampled using a stainless-steel soil corer with a diameter of 2 cm to a depth of ca. 5 cm in each plot, including only the O-horizon. In 2018, soil was sampled both before the next addition (i.e., 1 year after the last addition) and one day after the addition for analyses of total Mo and P content. Normalized difference vegetation index (NDVI) was measured in all years (2016–2018) during the sampling campaigns using a Skye 660/730 nm sensor to assess if the treatments lead to changes in vegetation greenness.
Nitrogen fixation in moss and soil
We used the acetylene reduction assay (ARA) as a measure of N2 fixation. For this, mosses (ca. 0.5 g dry weight equivalent) were placed in 50 ml centrifuge tubes in the laboratory in Copenhagen, sealed with a rubber septum (Suba Seal, Sigma) and 10% of the headspace was replaced with acetylene. Mosses were incubated in growth chambers for 24 h in light with an incremental increase in temperature over time: three hours at 4 °C, followed by one hour at 6 °C, one hour at 8 °C up to 10 °C for 14 h and a decrease in temperature after that to mimic the field conditions at the time of sampling. Six ml of the headspace was transferred into 6 ml pre-evacuated, air-tight vials (Labco, Ceredigion, UK) and analysed for ethylene concentration with a gas chromatograph (SRI 310C, FID, SIR Instruments, California). After the assay, moss samples were dried to determine dry weight (dw) (24 h at 70 °C). Acetylene reduction in soil was measured on ca. 1 g dry weight equivalent in 50 ml tubes as described above for the moss samples. Soil samples were incubated with acetylene gas (10%) in the dark at room temperature and gas samples were analysed like the moss gas samples described above. To convert from acetylene reduced (nmol g dw−1 h−1) to N2 fixed (μg N m−2 h−1; Online Resource 2), we used conversion factors established for the same moss species collected in the same plots (Rousk et al. 2017b), assuming a moss biomass of 200 g m−2 (e.g. Sorensen et al. 2006): 2.9 for the control and P-only treatments, 3.3 for the 10Mo (+P) treatments and 4.0 for the 100Mo (+P) treatments for the long-term assessments (1 year after addition); and 1.8 for the control and P-only treatments, 3.0 for the 10Mo (+P) treatments and 4.2 for the 100Mo (+P) treatments for the short-term assessments (1 day after addition; see Rousk et al. 2017b for more details).
Mo and P content of moss and soil
In 2018, dried moss and soil samples were analysed for total Mo and P content using an atomic absorption spectrometer (AAS) for the two time points (1 year and 1 day after a full addition). For this, 0.5 g moss and 1.0 g soil were weighed into 50 ml teflon tubes and digested using a 32% HNO3 solution in a MARS 6 Microwave, CEM. The digested samples were diluted to 8% HNO3 and analysed with flame atomic absorption spectrophotometry using an AAS (PerkinElmer PinAAcle 900 T).
Bacterial and fungal growth and soil characteristics
Bacterial growth was determined by measuring the rate of 3H-Leucine (Leu) incorporation in extracted bacteria (Rousk et al. 2009). One gram fresh soil was mixed with 20 ml demineralized water, vortexed for 3 min and centrifuged (10 min at 1000 g). The resulting bacterial suspension was incubated at 18 °C for 2 h, with 2 μl 1-[4,5-3H]-Leucine (5.7 TBq mmol−1, Perkin Elmer, USA) and unlabelled Leu with a final concentration of 275 nM Leu in the bacterial suspension. Bacterial growth was terminated after 2 h by adding 75 μl of 100% trichloroacetic acid. Centrifugation and washing was performed as described by Bååth et al. (2001). Scintillation cocktail (Ultima Gold; PerkinElmer, USA) was added and the radioactivity was measured using a liquid scintillation counter. The amount of leucine incorporated into extracted bacteria (pmol Leu incorporated g−1 SOM h−1) was used as a measure of bacterial growth.
Fungal growth was determined using the acetate-in-ergosterol (Ac-in-erg) incorporation method (Newell and Fallon 1991) adapted for soil (Bååth 2001; Rousk et al. 2009), which estimates the rate of ergosterol synthesis as a measure of fungal growth. Soil samples (0.5 g) were mixed with 20 μl of 14C-acetate solution ([1-14C] acetic acid, sodium salt, 2.07 GBq mmol−1, Perkin Elmer) and unlabelled sodium acetate, resulting in a final acetate concentration of 220 μM in the soil slurry. Samples were incubated at 18 °C for 4 h in the dark before growth was terminated by addition of formalin. Ergosterol and incorporated acetate were measured according to Rousk and Bååth (2007). The amount of acetate incorporated into ergosterol (pmol g−1 SOM h−1) was used as a measure of fungal growth. Ergosterol concentration was estimated from the UV absorbance at 282 nm compared with external standards.
Respiration rates were estimated as accumulation of headspace CO2 in closed vials. One gram soil was weighed into 20 ml glass vials. The headspace of glass vials was purged with pressurized air, before vials were sealed and incubated at 18 °C for approx. 24 h. The amount of CO2 produced during the incubation was determined using a gas chromatograph equipped with a methanizer and flame ionization detector. Substrate induced respiration was measured as a proxy for microbial biomass. Briefly, 15 mg 4:1 glucose:talcum was vigorously mixed into 1 g soil (corresponding to 4.8 mg glucose-C g−1 soil fwt). After 30 min, vials were purged with pressurized air and incubated at 22 °C for 2 h before the concentration of CO2 was determined. Substrate induced respiration was used to estimate microbial biomass (mg C g−1 SOM) (Anderson and Domsch 1978).
The microbial PLFA composition was determined using 2.0 g frozen soil, according to Frostegård et al. (1993) with modifications (Nilsson et al. 2007). An internal standard (methyl nonadecanoate fatty acid 19:0) was added before the methylation step for quantification. The derived fatty acid methyl esters were quantified on a gas chromatograph with flame ionization detector. Bacterial (i14:0, i15:0, a15:0, i16:0, 16:1ω9, 16.1ω7c, i17:0, a17:0, 17:1ω8, cy17:0, 10Me17:0, 18:1ω7, 10Me18:0 and cy19:0) and fungal-specific (18:2ω6,9) PLFAs were used to estimate the relative abundance of these functional groups (Frostegård and Bååth 1996; Ruess and Chamberlain 2010). The sum total concentration of the PLFAs 14:0, 15:0, 16:1ω5, 16:0, 18:1ω9, and 18:0, in addition to those listed above as bacterial and fungal biomarkers, was used as a measure of total microbial abundance (Frostegård and Bååth 1996; Ruess and Chamberlain 2010).
Soil moisture content was determined gravimetrically (105 °C for 24 h) and SOM content through loss on ignition (600 °C for 12 h). Soil pH and electrical conductivity (EC) were measured in a 1:5 (w:V) water extraction using an electrode and an EC meter, respectively.
Statistical analyses
Differences in N2 fixation rates between the field treatments and time after addition were tested with 3-way repeated measures ANOVA with year (2016–2018) and field treatment (Mo, P) as factors with three levels (0, 10, 100) and interactions, followed by Tukey’s honestly significant difference test. Two-way ANOVAs were run for soil microbial process rates, PLFA abundance (total, bacterial, fungal) and composition with nutrient supplement as factors, followed by Tukey’s honestly significant difference test. We used a principal component analysis (PCA) to screen for differences in the PLFA composition of the soil microbial community using the relative abundances (mol%) of PLFAs, after standardising to unit variance. The scores of the principal components were also subjected to ANOVA. Differences in moss and soil-Mo and P content between the treatments were assessed with 2-way ANOVAs, followed by Tukey’s honestly significant difference test. Data was log-transformed to reach normality and homogeneity of variances prior to the analyses. Analyses were performed in R 3.3.3 (R Core Team 2016) and in JMP 12.0.1 for Mac (SAS Institute, Cary, NC, USA).
Results
Soil and moss characteristics
Soil characteristics, such as pH and SOM were not different between the field treatments, and not even moss pH changed significantly with the high P additions (Table 1). Despite visible changes to the greenness in the plots (Online Resource 1), NDVI did not differ systematically across treatments but was generally lower in 2016 compared to the other years (F2,108 = 18.71, p < 0.0001; Online Resource 3).
Moss-Mo content was significantly different between the treatments for both time points (1 year: F8,27 = 11.10; p < 0.0001; 1 day: F10,34 = 9.71; p < 0.0001), with the highest Mo content in all treatments that contained the high Mo dose. Moss-Mo content doubled just after the application (60.8 μg g dw−1 ± 14.5) compared to moss-Mo content 1 year after the addition in the 100Mo addition plots (26.4 μg g dw−1 ± 7.02) (F1,88 = 4.63; p = 0.34; Fig. 1a, b). Soil-Mo content was almost 10 times lower than moss Mo content (F1, 51 = 6.16; p = 0.02) and did not differ between the treatments, or between the two time points (Fig. 1c, d).

Molybdenum content (μg g dw−1) in moss (a, b) and soil (c, d) 1 year (a, c) and 1 day (b, d) after the Mo field additions in subarctic tundra, N Sweden in 2018. Molybdenum and P were added in two different concentrations (10 and 100x moss content, see text for exact numbers) in a full factorial design. Given are means ±SE (n = 5). Please note the differences in the y-axes
One day after the P additions, moss-P content was higher in the high P addition plots compared to the other plots (F10,34 = 3.41; p = 0.004), except for the 100Mo + 10P treatment, but it did not differ between the treatments 1 year after the additions. Moss-P content was higher right after the additions than 1 year after the additions in all plots that received P (F8,70 = 2.70; p = 0.01; Fig. 2 a, b).

Phosphorus content (mg g dw−1) in moss (a, b) and soil (c, d) 1 year (a, c) and 1 day (b, d) after the P field additions in subarctic tundra, N Sweden in 2018. Molybdenum and P were added in two different concentrations (10 and 100x moss content, see text for exact numbers) in a full factorial design. Given are means ±SE (n = 5)
While there were no significant differences in soil-Mo content between the treatments at either time point (see above), soil-P content was higher in all treatments that contained P compared to the control and Mo only treatments after 1 year (F9,35 = 2.12; p = 0.05; Fig. 2c) -except for the 10Mo + 10P treatment, which was similar to the Mo-only treatments- but was only marginally higher 1 day after the applications (F10,34 = 1.89; p = 0.08; Fig. 2d). Surprisingly, all treatments receiving P, no matter the dose, had similar soil P-content (Fig. 2c, d).
Nitrogen fixation in moss and soil
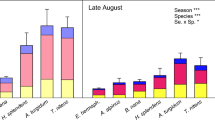
Nitrogenase activity (acetylene reduction) in the common moss Hylocomium splendens did not show a clear pattern in regard to the Mo and P additions (Fig. 3). No treatment effects on moss-associated N2 fixation could be detected in the long-term (Fig. 3 a,b,c). In the short-term, all treatments with high P additions lead to reduced N2 fixation rates -albeit not significantly. Acetylene reduction rates were higher 1 year after the nutrient supplements (overall mean across treatments: 6.33 ± 0.83 nmol g dw−1 h−1) than 1 day after the additions (3.5 ± 0.94 nmol g dw−1 h−1; F1,249 = 24.70; p < 0.0001). Further, the addition of 100xMo in 2017 lead to the highest rates recorded in this study 1 day after the additions (41.80 ± 17.7 nmol g dw−1 h−1; F8,250 = 2.87; p = 0.005), and N2 fixation rates were higher than the rates in all other treatments at the same measuring campaign, except for the low Mo addition. Across years, there was no clear increasing or decreasing trend with time, since N2 fixation rates were higher in 2017 than in 2016 but decreased again in 2018 (Fig. 3). The short-term inhibition by the high P additions disappeared in the long-term.

Nitrogen fixation as acetylene reduction (nmol g dw−1 h−1) in the moss Hylocomium splendens in response to field molybdenum (Mo) and phosphorus (P) additions over 3 years in subarctic tundra, N Sweden. Molybdenum and P were added in two different concentrations (10 and 100x moss content, see the main text for exact numbers) in a full factorial design. Acetylene reduction was measured 1 year as well as after 1 day after a field addition. Shown are means ±SE (n = 5). Different lowercase letters indicate significant differences between treatments (Tukey’s test). Please note the differences in the y-axes
Converted to N2 fixation, rates in Hylocomium splendens were as high as 78 μg N m−2 h−1 (corresponding to 3.4 kg N ha−1 yr−1 assuming half a year of activity at field moisture and temperature regimes as during the ARA incubations) in the 10Mo + 10P treatment 1 year after the additions, and 130 μg N m−2 h−1 (corresponding to 5.7 kg N ha−1 yr−1) in the 100Mo treatment 1 day after the additions (Online Resource 2).
Nitrogen fixation activity in soil was low and variable, and did not differ between treatments expressed per gram soil or per gram SOM, although N2 fixation in the low P addition treatment was high (Fig. 4a, b).

Soil N2 fixation rates as acetylene reduction (a) per g dw and (b) per g SOM in response to field molybdenum (Mo) and phosphorus (P) additions measured 1 year after the last addition in subarctic tundra, N Sweden. Molybdenum and P were added in two different concentrations (10 and 100x moss content, see text for exact numbers) in a full factorial design. Shown are means ±SE (n = 5). Please note the differences in the y-axes
Soil microbial process rates, abundance, and community structure
Soil microbial process rates were variable across the field treatments. For instance, bacterial growth was 10 times higher in the combined 10xMo + 10xP treatments (4544 pmol Leu g−1 SOM h−1) compared to the low Mo addition plots (381 pmol Leu g−1 SOM h−1). Respiration and SIR-biomass ranged from 66 μg CO2 g−1 SOM h−1 and 5.4 mg C g−1 SOM in the low P treatments to 16.8 μg CO2 g−1 SOM h−1 and 2.3 mg C g−1 SOM in control treatments for respiration and SIR-biomass, respectively. Hence, despite high P application rates, and a legacy of high N2 fixation activity in the plots (> 1 kg N ha−1 yr−1; Rousk et al. 2017b), we did not find any systematic and significant differences in microbial growth, SIR-biomass or respiration rates between the treatments (Fig. 5). Similarly, the field treatments had no effect on bacterial and fungal biomass (Fig. 6b, c) or total PLFAs (Fig. 6a), even though bacterial and fungal biomass was high in the 10xP plots (478 and 107 nmol g SOM−1 for bacterial and fungal PLFAs, respectively; Fig. 6b,c).

Bacterial growth estimated with leucine incorporation (a), fungal growth estimated with acetate incorporation into ergosterol (b), the fungal-to-bacterial-growth ratio (c), respiration (d), SIR-biomass (e) and ergosterol concentrations (f) in soils from the Mo and P field treatments. Values are means ±SE (n = 5)

Total (a), bacterial (b) and fungal (c) PLFA concentrations in subarctic soils collected in the Mo and P field addition plots. Given are means ±SE (n = 5)
The two first principal components of the microbial PLFAs together explained 57.5% of the variation (Fig. 7). Although neither nutrient supplement affected the PC1 scores systematically (p > 0.19 for both nutrients), treatments separated in response to P additions (p = 0.006) along PC2, which explained 28.3% of the PLFA variation (Fig. 7). Several bacterial PLFA markers were associated with higher PC2 scores, including most gram-positive markers (i14:0, i15:0, a15:0, i16:0, i17:0, a17:0) and some gram-negative ones (including 18:1ω9, 17:1ω8) (Fig. 7b). In contrast, the fungal marker 18:2ω6,9 was associated with negative PC2 score values (Fig. 7b).

(a) Case scores of the two first components of the PCA on microbial PLFA composition in soils collected in the Mo and P addition plots in subarctic tundra and (b) PCA variable loadings of the microbial community composition derived from PLFA analysis
Discussion
Nitrogen fixation responses towards Mo and P supplements
We hypothesized that the added P would have a long-lasting, positive effect on N2 fixation (H1). While there were tendencies for effects (see discussion below), there was neither a clear long-term (1–3 years) nor a short-term (1 day) positive effect of P additions on N2 fixation in moss. We are aware of only one long-term (7 years) study that assessed the effects of P additions on N2 fixation in free-living bacteria. This study, conducted in tropical forest soils, reported a positive effect of long-term P additions on N2 fixation but this positive effect was attributed to Mo contamination of the P fertilizer (Barron et al. 2009). Our P additions (40 and 400 kg ha−1 yr−1) were several fold higher than deposition estimates for arctic systems (5 g ha−1 yr−1; Peñuelas et al. 2013), but the low P supplements were within the order of magnitude of other studies on nutrient limitation of N2 fixation (~10 kg P ha−1; Wurzburger et al. 2012; Perakis et al. 2017). However, ecosystem comparisons are laden with caveats that challenge interpretation and causal identification. For instance, the dominating N2 fixers in tropical and temperate forest soils likely differ from those colonizing mosses in the Subarctic, and nutrient requirements, utilization and storage capabilities may vary among diazotrophs.
Although there were tendencies for positive (low P additions in all combinations) and negative (high P additions in all combinations) effects of P on moss-associated N2 fixation in our study, the negative effects of the high P additions disappeared in the long-term, while the positive effects of the low P additions only became apparent in the long-term. Any excess P could have been leached from the moss carpet over the period of one year in the high P plots, and could have been taken up by other plants in the same plots via mycorrhiza (Carleton and Read 1991). Alternatively, the added P could have been utilised by moss and cyanobacteria for other processes than N2 fixation, such as photosynthesis, or growth, which may have diluted any effects in the longer term. However, the lack of differences in NDVI between the field treatments, which should reflect growth and photosynthesis, makes this possibility less likely.
High Mo additions promoted N2 fixation in the short-term on only one occasion (Fig. 3e), which does not conclusively falsify or verify our second hypothesis (H2). A previous study on Mo limitation over time reports a positive effect on N2 fixation in litter (Silvester 1989) and moss (K Rousk, unpublished) with maximum rates between 30 and 40 h after the additions in the laboratory, and halved rates between 30 and 50 days after the additions in the field (Silvester 1989) and in the laboratory (K Rousk, unpublished). This suggests that Mo additions can affect nitrogenase synthesis within one day, which was also observed in the plots that we revisited (Rousk et al. 2017b), a pattern that could not be confirmed here for all measuring campaigns. Our Mo additions (70 g ha−1 and 7 g ha−1) were slightly higher, but still within a similar range to additions in other studies. For instance, clear positive effects of Mo additions of 4.7 g Mo ha−1, but not of 0.4 g Mo ha−1 on N2 fixation by free-living bacteria has been found in tropical forests (Barron et al. 2009).
Moss-associated N2 fixation was promoted by additions of Mo and P at the start of the experiment in 2015 (see Rousk et al. 2017b), which could have led to increased biomass of cyanobacteria, increasing the nutrient demand (Mo and P) of the diazotroph community colonizing mosses. Molybdenum is one of the rarest elements on earth and is replenished only via weathering and deposition (Marks et al. 2015). Further, Mo availability is strongly linked to soil organic matter via complexation with plant-derived tannins and tannin-like compounds, which decrease with higher soil pH (Wichard et al. 2009; Marks et al. 2015). Similarly, P in soil is mostly unavailable, and enters the system via deposition, mineralization and weathering. Hence, Mo and P could limit nitrogenase activity in our system. However, given the lack of a positive response to Mo and P, the frequent additions could have saturated demand, until Mo and P were no longer limiting.
If cyanobacterial Mo demand was saturated in our system as a result of repeated Mo supplements, any added Mo not utilised by cyanobacteria could have been used by nitrate reducing bacteria given that nitrate reductase contains Mo as a co-factor. Due to the previously high N2 fixation rates in the Mo-addition plots (Rousk et al. 2017b), nitrate pools may have increased in soils beneath the moss carpet, facilitating denitrification processes. While no link between N2 fixation and denitrification and N2O emission could be detected within the moss carpet in a temperate bog (Rousk et al. 2018b), increased N input to soil could benefit nitrate reducing bacteria in our N-limited system. Intriguingly, moss P content in the Mo addition plots was similar to P content of mosses to which P was added. This could be the results of P acquisition due to high Mo availability, as symbiotic N2 fixers have been found to mine for P in soil (Compton and Cole 2001).
Diazotrophs are able to store large amounts of Mo -up to 10 times more than is required for growth- (Bellenger et al. 2011), reducing potential positive effects of any further additions. Alternatively, by adding Mo, we could have shifted the diversity of nitrogenase enzyme isoforms towards the Mo-based nitrogenase, while the vanadium form of the enzyme (V-nase) may otherwise dominate our system (Rousk et al. 2017b). Darnajoux et al. 2019 suggest a threshold of 250 ng Mo g−1 below which the enzyme shifts from Mo-based to V-based. In our study, this threshold was exceeded in all treatments receiving Mo supplements (5–60 μg g moss−1), and even mosses from the control plots had a Mo content of ca. 1 μg g moss−1, indicating that the Mo-based nitrogenase prevails in our experiment, and adding more Mo did not promote N2 fixation further. However, the threshold proposed by Darnajoux et al. 2019 was established using lichen samples. Diazotroph associated with lichens may have different nutrient requirements and - storage capabilities, and the threshold may not be easily applicable to moss-associated diazotrophs.
We used plot specific conversion ratios to convert from acetylene reduced to N2 fixed (Online Resource 2), which makes the conversion from ARA to units N2 fixed unusually robust (Bellenger et al. 2011), but also adds the inherent variation factor of a unit conversion. Further, conversion factors can vary not only between enzyme isoforms, but also across time (Saiz et al. 2019). Temporal variation in conversion factors could confound treatment effects across years. Nevertheless, no clear treatment differences were found in a direct comparison between the acetylene reduction rates without conversion either (Fig. 3), where we should have been able to detect a promotion of the Mo-based nitrogenase in all measuring campaigns. Hence, our results consistently show that moss-associated N2 fixation is not limited by Mo and P in our study system.
When we compare our results to the relevant time points in the same plots measured in 2015 (Rousk et al. 2017b) overall patterns match well. This includes N2 fixation activity as well as the response pattern of low rates in the high P addition plots, and promotion in all plots that received low P additions. Nonetheless, large variation in N2 fixation between plots as well as climatic variation between measurement years may overshadow significant effects of the nutrient supplements. For instance, cumulative precipitation from the start of the growing season until the time of measurement (April–July) was 174.6, 150.8, 150.4 and 120.5 mm for the years 2015, 2016, 2017, 2018, respectively (SMHI; Fig. 8). In the wetter years (2015, 2017), treatment effects could be detected and N2 fixation was higher compared to the other, drier years. Precipitation was similar in 2016 and 2017, but air temperatures were lower in 2015 and 2017, indicating that the optimal conditions for moss-associated N2 fixation are a combination of high moisture (e.g. > 150 mm precipitation) and low temperatures (< 5 °C mean monthly temperature). The effects of moss-moisture on cyanobacterial activity may override the effects of the nutrient supplements, since no N2 fixation is taking place in dry mosses (Rousk et al. 2014).

Nitrogen fixation as acetylene reduction (nmol g dw−1 h−1) ± SE (n = 5–15) across the Mo and P field additions close to Abisko, N Sweden between years in relation to (a) cumulative growing season precipitation (mm) and (b) mean monthly temperature (°C) in the months April to July
The sensitivity of resolving limiting factors (e.g. Mo, P) for processes (including N2 fixation) will depend on the overall rates of the process for two reasons. First, the signal-to-noise ratio will increase with high values, and second, a biological process will only consume resources rapidly when it occurs at high rates. For these reasons, it is possible that the effects of nutrient supplements on N2 fixation will depend on the overall rates of the process, and that it will only be detectable when it reaches high levels (e.g. in 2017). We assessed links between N2 fixation activity in control samples and increased activity as a result of nutrient additions by regressing the difference in N2-fixation between Mo supplemented samples and the controls against the rate in the control. But there was only one positive correlation (out of 48) between rates in control samples and an increase in rates due to nutrient supplements. Hence, we could exclude this possibility.
Soil N2 fixation was very low - two orders of magnitude lower than moss-N2 fixation. Hence, free-living, heterotrophic N2 fixing bacteria below the moss carpet seems negligible at our sites, probably due to inhibitory effects of mosses (Klavina et al. 2015). Adding a large amount of Mo alone inhibited N2 fixation in soil, while adding it together with low P, or adding low Mo and high P together increased N2 fixation rates in soil. This indicates a limitation by P, rather than by Mo, of N2 fixation in soil. Reed et al. (2013) suggested that N2 fixation in litter is limited by Mo if litter Mo content is below 100 ng g−1, and limited by P when litter Mo content is above 250 ng g−1 (see also Darnajoux et al. 2019). In our experiment, soil Mo content was mostly above 100 ng g−1 (Fig. 1c, d), suggesting P-limitation rather than Mo limitation of free-living N2 fixation in our soils.
Soil microbial responses to towards Mo and P supplements
We expected that we would induce P-limitation of soil microbes in the Mo addition plots, (H3), and alleviate this putatively induced P-limitation in the P addition plots (i.e. higher soil microbial process rates in the Mo + P treatments than in the Mo-only treatments). Although N2 fixation stimulation did not occur in the Mo or P treated plots in 2016–2018, earlier work in these plots showed high N2 fixation activity in 2015 (Rousk et al. 2017b). Given that Mo did not systematically stimulate N2 fixation, and subsequent delivery of N belowground, the expected microbial stimulation by P additions were subtle if responses could be observed at all. While some microbial measures (PLFA and SIR-biomass, respiration, N2 fixation) were slightly increased by low P additions, the Mo-only treatments did not result in lower process rates. Any systematic effects by the added nutrients needed to be sufficiently large to be detected from the high background variation in our soil microbial measures between plots. For instance, bacterial growth showed 15-fold differences between treatments, ranging from 300 to 4500 pmol Leu g−1 SOM h−1. Variation in bacterial growth could be explained by differences in pH (R 2 = 0.48; p = 0.02) and soil organic matter (SOM) content (R2 = 0.46; p = 0.03), and fungal growth by differences in SOM (R2 = 0.33; p = 0.05), independent of treatment. This suggests that variation in other factors exceeded the effects of P and Mo supplements, thus constraining the responses we can expect to be smaller than that assigned to other environmental factors. Similarly, N2 fixation as well as NDVI differed significantly between field replicates within the treatments indicating that the variation caused by the field nutrient additions was smaller than that caused by variation in other factors at the site. Although it has been shown that microbial decomposition in arctic tundra soils can be N-limited (Sistla et al. 2012), and microorganisms have even been reported to be N-limited in study sites proximal to those here assessed (Jonasson et al. 1996, 1999), it remains unknown how widespread this is. In most soils, the microbial community is predominantly limited by lack of C rather than nutrients (Rosinger et al. 2019), even in boreal forest soils with high C/N ratios (Demoling et al. 2007). If these suggestions can be generalised to also hold true here, a soil microbial community growth-limited by C would be consistent with the results obtained. By increasing N and P availability in the plots via increased N2 fixation following the Mo and P additions, we may have merely strengthened the already manifested microbial C limitation (Demoling et al. 2007; Rosinger et al. 2019).
Not surprisingly, Mo and P content was higher in moss than in soil (more pronounced for Mo than for P), and the added nutrients reached the soil after a delay (> 1 day). Further, differences in P and Mo content in the moss were most obvious just after the additions, while treatment differences prevailed in the soil for at least 1 year for the P additions. These results highlight the high nutrient absorption capacity of mosses (Aldous 2002), while at the same time, the substantial loss of nutrients from the moss carpet via leaching (Carleton and Read 1991; Coxson 1991; Wilson and Coxson 1999). However, moss-Mo content was two orders of magnitude higher than soil-Mo content, while P contents of soil and moss were similar. This indicates that Mo is not transferred easily from moss to soil, in contrast to P. Molybdate is strongly held by polyphenolic complexes that decrease availability, not only in soil (Wichard et al. 2009), but potentially also within the moss carpet.
Our findings highlight the importance of moss-associated N2 fixation as a main N source for arctic systems, with N2 fixation rates higher than 5 kg N ha−1 yr−1 in Mo supplemented plots – and up to 2.4 kg N ha−1 yr−1 in control plots. More importantly, moss-associated N2 fixation does not seem to be limited by a lack of Mo or P, and thus, the ecosystem influx of N via N2 fixation by cyanobacteria is not chronically rate limited by a lack of these nutrients. Instead, short-term climate variability seems to be a stronger driver of moss-associated N2 fixation (see also e.g. Rousk et al. 2015, 2018a). Further, the transient stimulation of N2 fixation by cyanobacteria does not impact the soil microbial decomposer community or processes in a detectable way. This suggests that the microbial decomposers were not limited by N at our sites. Generalising from our here obtained results, subarctic systems are differently regulated than e.g. tropical systems. The physical climate sets the threshold for arctic systems, while mineral nutrient limitation sets the threshold for tropical systems (e.g. Vitousek 1984). Yet, the occasional Mo and P limitation we observed here may shift into a chronic limitation given the rise in atmospheric CO2 that can intensify P and Mo limitation of N2 fixation (Trierweiler et al. 2018). Such a change would reduce the N input to pristine ecosystems via the N2 fixation pathway, which will ultimately lead to increased N limitation for plant growth.
References
Ackermann K, Zackrisson O, Rousk J, Jones DL, DeLuca TH (2012) N2 fixation in feather mosses is a sensitive indicator of N deposition in boreal forests. Ecosystems 15:986–998
Aldous AR (2002) Nitrogen translocation in Sphagnum mosses: effects of atmospheric nitrogen deposition. New Phytol 156:241–253
Anderson JPE, Domsch KH (1978) A physiological method for the quantitative measurement of microbial biomass in soils. Soil Biol Biochem 10:215–221
Bååth E (2001) Estimation of fungal growth rates in soil using 14C-acetate incorporation into ergosterol. Soil Biol Biochem 33:2011–2018
Bååth E, Pettersson M, Söderberg KH (2001) Adaptation of a rapid and economical microcentrifugation method to measure thymidine and leucine incorporation by soil bacteria. Soil Biol Biochem 33:1571–1574
Barron AR, Wurzburger N, Bellenger JP, Wright SJ, Kraepiel AML, Hedin LO (2009) Molybdenum limitation of asymbiotic nitrogen fixation in tropical forest soils. Nat Geosci 2:42–45
Bellenger JP, Wichard T, Xu Y, Kraepiel AML (2011) Essential metals for nitrogen fixation in a free-living N2-fixing bacterium: chelation, homeostasis and high use efficiency. Environ Microbiol 13:1395–1411
Carleton TJ, Read DJ (1991) Ectomycorrhizas and nutrient transfer in conifer-feather moss ecosystems. Can J Bot 69:778e785
Compton JE, Cole DW (2001) Fate and effects of phosphorus additions in soils under N2-fixing red alder. Biogeochem 53:225–247
Core Team R (2016) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria https://www.R-project.org
Coxson DS (1991) Nutrient release from epiphytic bryophytes in tropical montane rain forest (Guadeloupe). Can J Bot 69:2122–2129
Darnajoux R, Magain N, Renaudin M, Lutzoni F, Bellenger JP, Zhan X (2019) Molybdenum threshold for ecosystem scale alternative vanadium nitrogenase activity in boreal forests. PNAS 116(49):24682–24688
DeLuca TH, Zackrisson O, Nilsson MC, Sellstedt A (2002) Quantifying nitrogen-fixation in feather moss carpets of boreal forests. Nature 419:917–919
Demoling F, Figueroa D, Bååth E (2007) Comparison of factors limiting bacterial growth in different soils. Soil Biol Biochem 39:2485–2495
Dynarski KA, Houlton BZ (2018) Nutrient limitation of terrestrial free-living nitrogen fixation. New Phytol 217:1050–1061
Frostegård Å, Bååth E (1996) The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil. Biol Fert Soils 22:59–65
Frostegård Å, Tunlid A, Bååth E (1993) Phospholipid fatty acid composition, biomass, and activity of microbial communities from two soil types experimentally exposed to different heavy metals. Appl Environ Microb 59:3605–3617
Gundale MJ, DeLuca TH, Nordin A (2011) Bryophytes attenuate anthropogenic nitrogen inputs in boreal forests. Glob Chan Biol 17:2743–2753
Jean ME, Phalyvong K, Foster-Drolet J, Bellenger JP (2013) Molybdenum and phosphorus limitation of asymbiotic nitrogen fixation in forests of eastern Canada: influence of vegetative cover and seasonal variability. Soil Biol Biochem 67:140–146
Jonasson S, Michelsen A, Schmidt IK, Nielsen EV, Callaghan TV (1996) Microbial biomass C, N and P in two arctic soils and responses to addition of NPK fertilizer and sugar: implications for plant nutrient uptake. Oecologia 106:507–515
Jonasson S, Michelsen A, Schmidt IK, Nielsen EV (1999) Responses in microbes and plants to changed temperature, nutrient, and light regimes in the arctic. Ecology 80:1828–1843
Klavina L, Springe G, Nikolajeva V, Martsinkevich I, Nakurte I, Dzabijeva D, Steinberga I (2015) Chemical composition analysis, antimicrobial activity and cytotoxicity screening of Moss extracts (Moss Phytochemistry). Molecules 20:17221–17243
LeBauer DS, Treseder KK (2008) Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 89:371–379
Leppänen S, Salemaa M, Smolander A, Mäkipää R, Tiirola M (2013) Nitrogen fixation and methanotrophy in forest mosses along a N deposition gradient. Environ Exp Bot 90:62–69
Liang C, Schimel JP, Jastrow JD (2017) The importance of anabolism in microbial control over soil carbon storage. Nat Microbiol 2(8):17105
Mack MC, Schuur EA, Bret-Harte MS, Shaver GR, Chapin FS III (2004) Ecosystem carbon storage in arctic tundra reduced by long-term nutrient fertilization. Nature 431:440–443
Marks JA, Perakis SS, King KE, Pett-Ridge J (2015) Soil and organic matter regulates molybdenum storage and mobility in forests. Biogeochem 125:167–183
Newell SY, Fallon RD (1991) Toward a method for measuring instantaneous fungal growth rates in field samples. Ecology 72:1547–1559
Nilsson LO, Bååth E, Falkengren-Grerup U, Wallander H (2007) Growth of ectomycorrhizal mycelia and composition of soil microbial communities in oak forest soils along a nitrogen deposition gradient. Oecologia 153:375–384
Peñuelas J, Poulter B, Sardans J, Ciais P, van der Velde M, Bopp L, Janssens IA (2013) Human-induced nitrogen–phosphorus imbalances alter natural and managed ecosystems across the globe. Nature comm 4:2934. https://doi.org/10.1038/ncomms3934
Perakis SS, Pett-Ridge JC, Catricala CE (2017) Nutrient feedbacks to soil heterotrophic nitrogen fixation. Biogeochem 134:41–55
Reed SC, Cleveland CC, Townsend AR (2013) Relationship among phosphorus, molybdenum and free-living nitrogen fixation in tropical rain forests: results from observational and experimental analyses. Biogeochem 114:135–147
Rinnan R, Michelsen A, Bååth E (2013) Fungi benefit from two decades of increased nutrient availability in tundra heath soil. PLoS One 8(2):e56532
Rosinger C, Rousk J, Sanden H (2019) Can enzymatic stoichiometry be used to determine growth-limiting nutrients for microorganisms? -a critical assessment in two subtropical soils. Soil Biol Biochem 128:115–126
Rousk J, Bååth E (2007) Fungal biomass production and turnover in soil estimated using the acetate-in-ergosterol technique. Soil Biol Biochem 39:2173–2177
Rousk K, Michelsen A (2017) Ecosystem nitrogen fixation throughout the snow-free period in subarctic tundra: effects of willow and birch litter addition and warming. Glob Chan Biol 23:1552–1563
Rousk J, Brookes PC, Bååth E (2009) Contrasting soil pH effects on fungal and bacterial growth suggest functional redundancy in carbon mineralization. Appl Environ Microb 75:1589–1596
Rousk K, Rousk J, Jones DL, Zackrisson O, DeLuca TH (2013) Feather moss nitrogen acquisition across natural fertility gradients in boreal forests. Soil Biol Biochem 61:86–95
Rousk K, Jones DL, DeLuca TH (2014) The resilience of nitrogen fixation in feather moss (Pleurozium schreberi)-cyanobacteria associations after a drying and rewetting cycle. Plant Soil 377:159–167
Rousk K, Sorensen PL, Lett S, Michelsen A (2015) Across habitat comparison of diazotroph activity in the subarctic. Microb Ecol 69:778–787
Rousk K, Pedersen PA, Dyrnum K, Michelsen A (2017a) The interactive effects of temperature and moisture on nitrogen fixation in two temperate-arctic mosses. Theor Exp Plant Phys 29:25–36
Rousk K, Degboe J, Michelsen A, Bradley R, Bellenger JP (2017b) Molybdenum and phosphorus limitation of moss-associated nitrogen fixation in boreal ecosystems. New Phytol 214:97–107
Rousk K, Sorensen PL, Michelsen A (2017c) Nitrogen fixation in the high Arctic: a source of ‘new’ nitrogen? Biogeochem 136:213–222
Rousk K, Sorensen PL, Michelsen A (2018a) What drives biological nitrogen fixation in high arctic tundra: moisture or temperature? Ecosphere 9:e02117
Rousk K, Vestergård M, Christensen S (2018b) Are nitrous oxide emissions and nitrogen fixation linked in temperate bogs? Soil Biol Biochem 123:74–79
Ruess L, Chamberlain PM (2010) The fat that matters: soil food web analysis using fatty acids and their carbon stable isotope signature. Soil Biol Biochem 42:1898–1910
Saiz E, Sgouridisb F, Drijfhoutc FP, Ullahd S (2019) Biological nitrogen fixation in peatlands: comparison between acetylene reduction assay and 15N2 assimilation methods. Soil Biol Biochem 131:157–165
Silvester WB (1989) Molybdenum limitation of asymbiotic nitrogen fixation in forests of pacific Northwest America. Soil Biol Biochem 21:283–289
Sistla SA, Asao S, Schimel JP (2012) Detecting microbial N-limitation in tussock tundra soil: implications for Arctic soil organic carbon cycling. Soil Biol Biochem 55:78–84
SMHI-The Swedish Meteorological and Hydrological Institute. www.smhi.se Accessed May 2019
Sorensen PL, Jonasson S, Michelsen A (2006) Nitrogen fixation, denitrification, and ecosystem nitrogen pools in relation to vegetation development in the subarctic. Arct Antarct Alp Res 38:263–272
Trierweiler AM, Winter K, Hedin LO (2018) Rising CO2 accelerates phosphorus and molybdenum limitation of N2 fixation in young tropical trees. Plant Soil 429:363–373
Vitousek PM (1984) Litterfall, nutrient cycling, and nutrient limitation in tropical forests. Ecology 65:285–298
Wang YP, Law RM, Pak B (2010) A global model of carbon, nitrogen and phosphorus cycles for the terrestrial biosphere. Biogeosci 7:2261–2282
Wichard T, Mishra B, Myneni SC, Bellenger JP, Kraepiel AML (2009) Storage and bioavailability of molybdenum in soils increased by organic matter complexation. Nat Geosci 2:625–629
Wilson JA, Coxson DS (1999) Carbon flux in a subalpine spruce-fir forest: pulse release from Hylocomium splendens feather-moss mats. Can J Bot 77:564–569
Wurzburger N, Bellenger JP, Kraepiel AML, Hedin LO (2012) Molybdenum and phosphorus interact to constrain asymbiotic nitrogen fixation in tropical forests. PLoS One 7:e33710
Acknowledgements
We thank M. H. Wahlgren and G. Sylvester for assistance with laboratory analyses. Abisko Scientific Research Station kindly provided logistic support. Funding was provided by the Independent Research Fund Denmark (IRFD) “Research Project 1” (Grant ID: DFF—6108-00089), by the IRFD Sapere Aude Grant (Grant id: 7027-00011B), and by the Knut and Alice Wallenberg Foundation (KAW 2017.0171).
Author information
Authors and Affiliations
Contributions
KR designed the field study. KR and JR conducted the field additions and field sampling. KR (moss related analyses) and JR (soil related analyses) performed the laboratory assays and analysed the data. KR and JR wrote the manuscript.
Corresponding author
Additional information
Responsible Editor: Ulrike Mathesius.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
ESM 1
(PDF 1784 kb)
Rights and permissions
About this article

Cite this article
Rousk, K., Rousk, J. The responses of moss-associated nitrogen fixation and belowground microbial community to chronic Mo and P supplements in subarctic dry heaths. Plant Soil 451, 261–276 (2020). https://doi.org/10.1007/s11104-020-04492-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-020-04492-6